
Vesicular MicroRNA as Potential Biomarkers of Viral Rebound

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
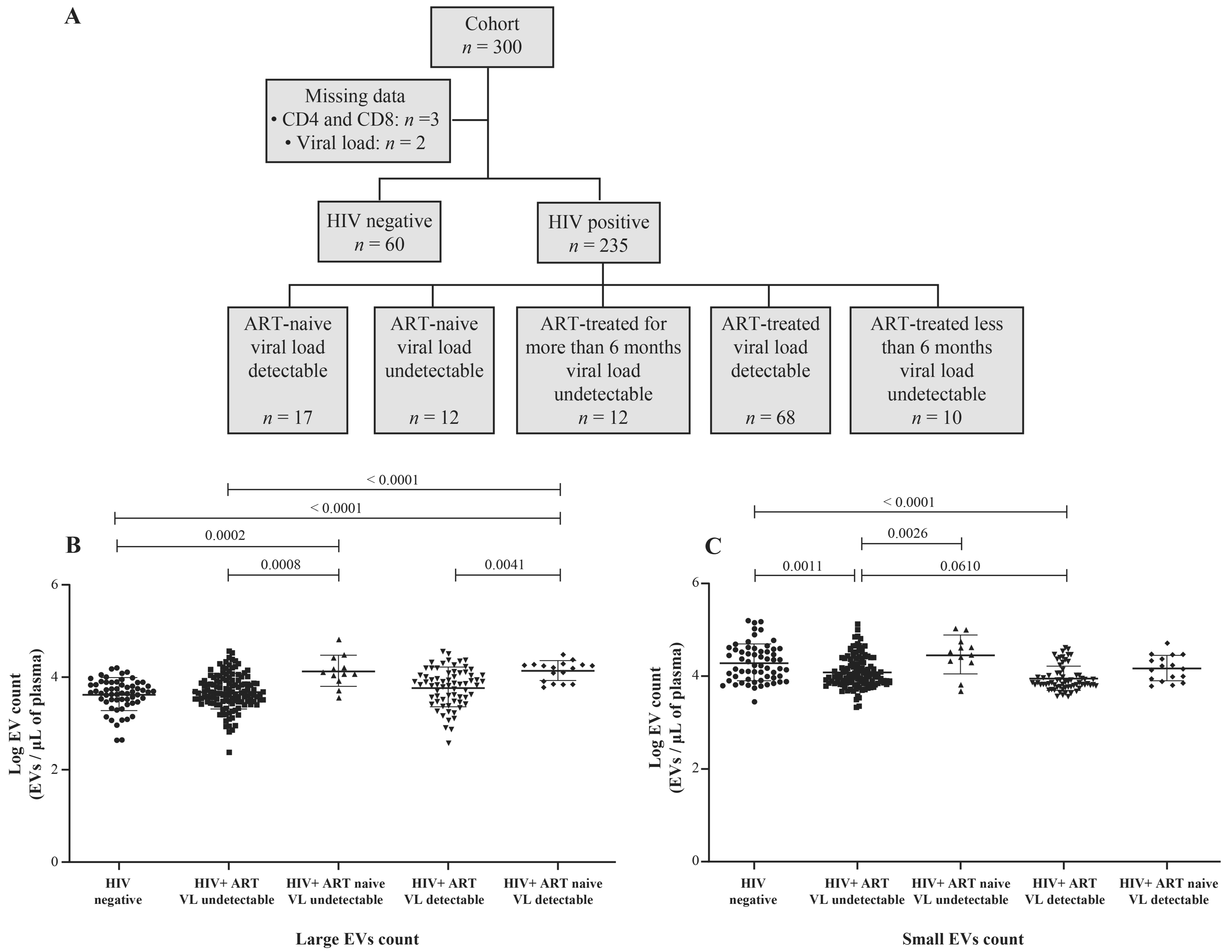
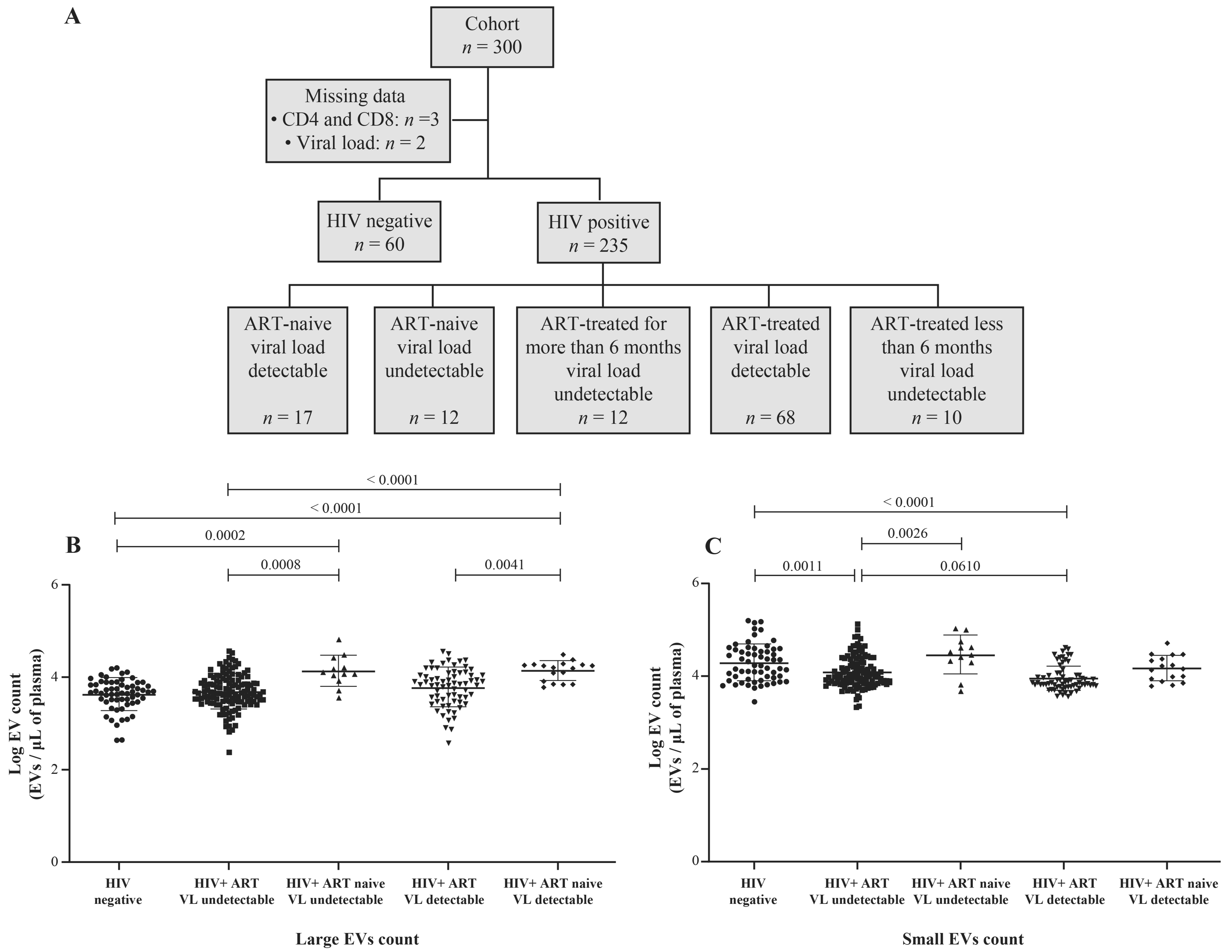
2.1. Study Subjects
2.2. Quantitation of HIV-1 RNA, CD4 and CD8 T Lymphocytes
2.3. Purification of Extracellular Vesicles
2.4. EV Flow Cytometry Analysis
2.5. MicroRNA Quantification
2.6. Transmission Electron Microscopy
2.7. Statistical Analysis
3. Results
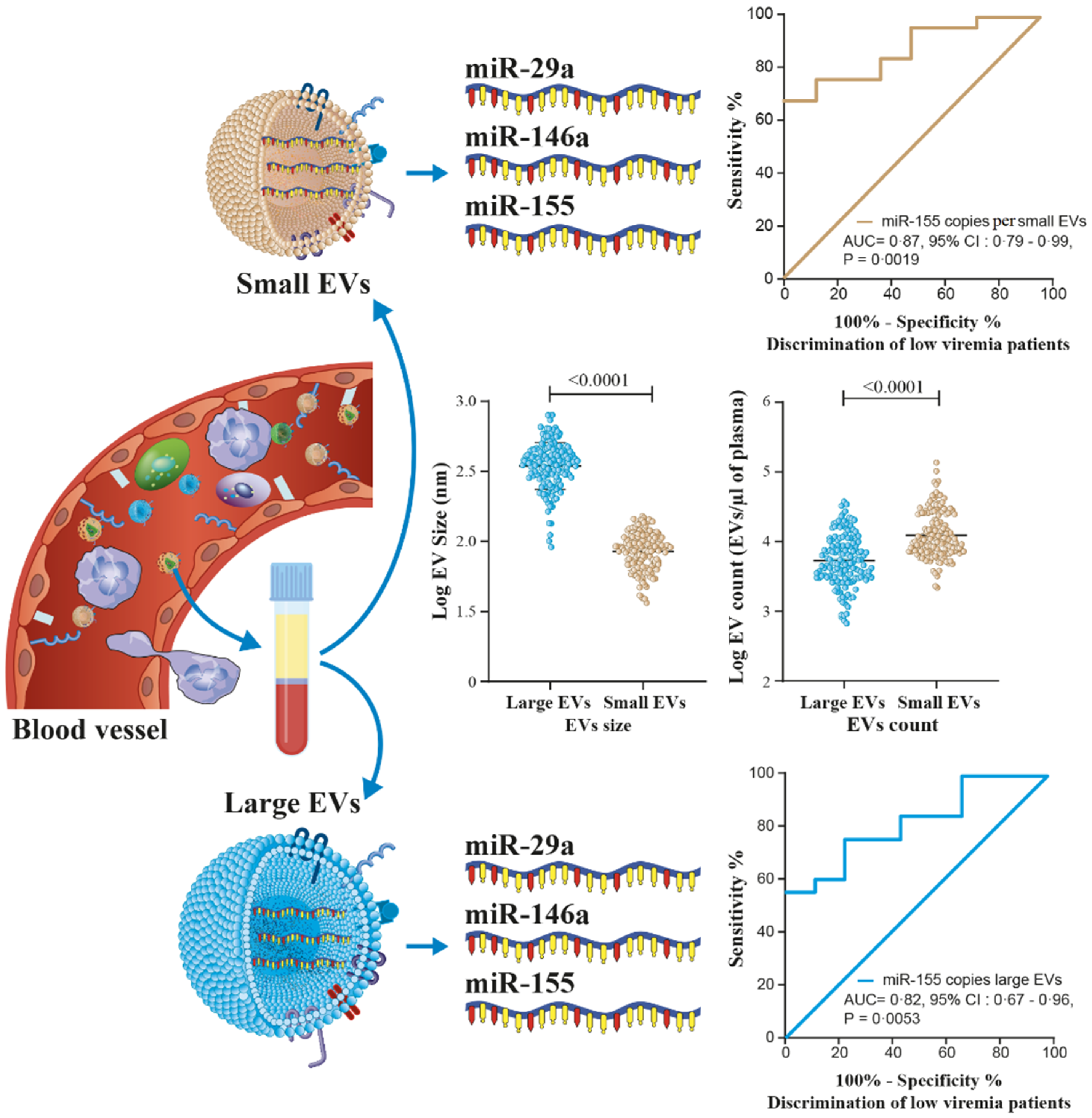
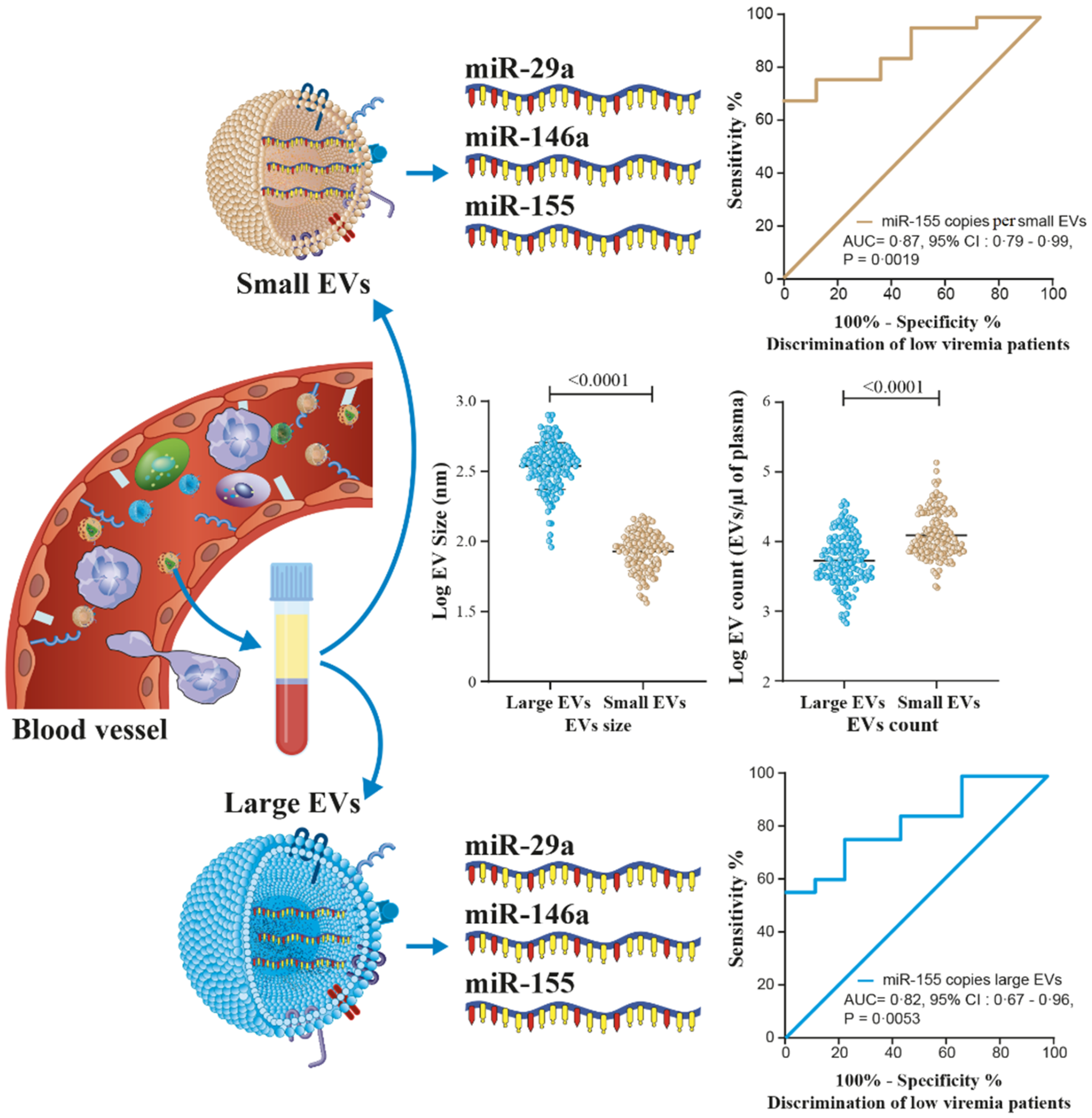
3.1. HIV-1 Replication Could Be Induces the Production of Large EVs
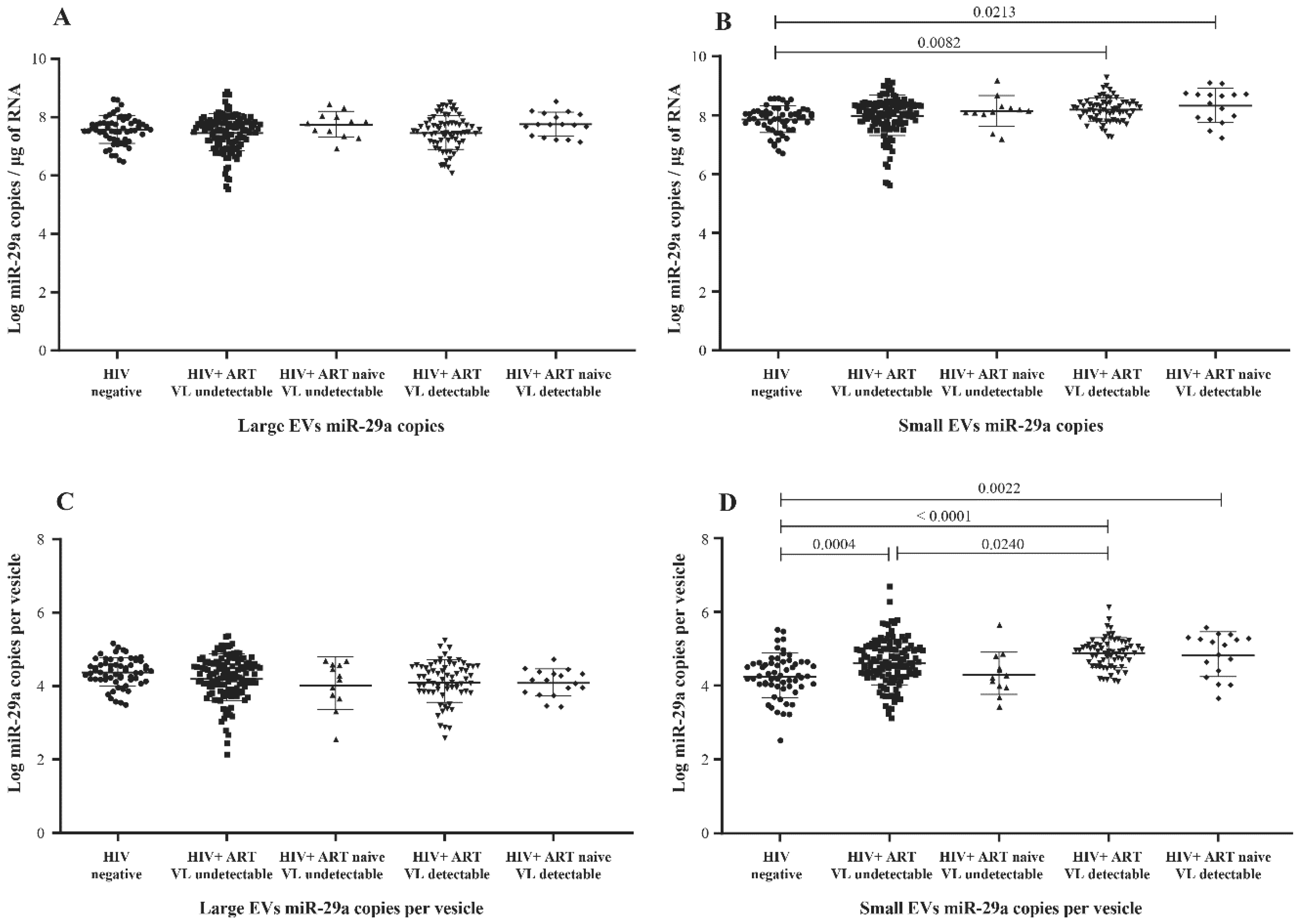
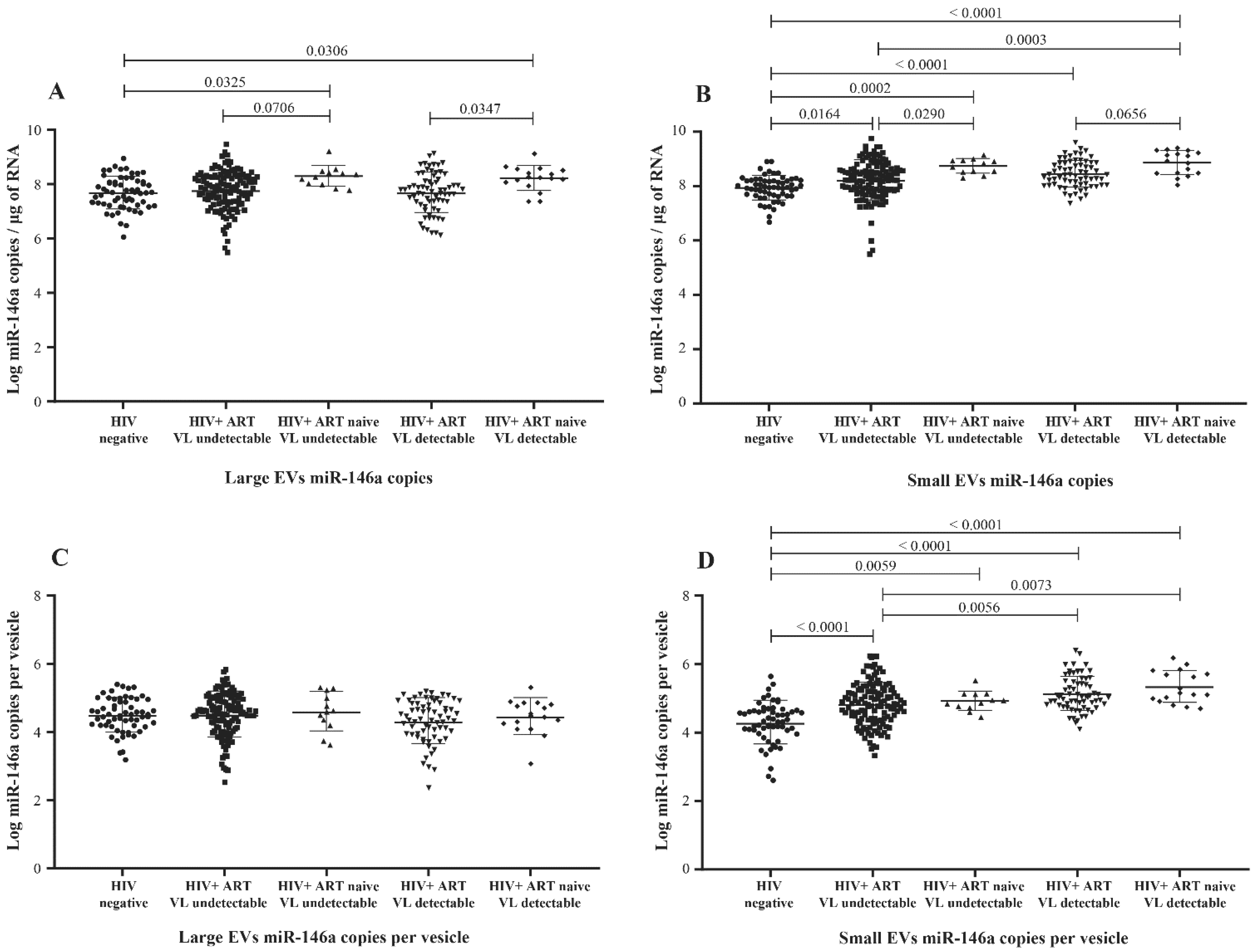
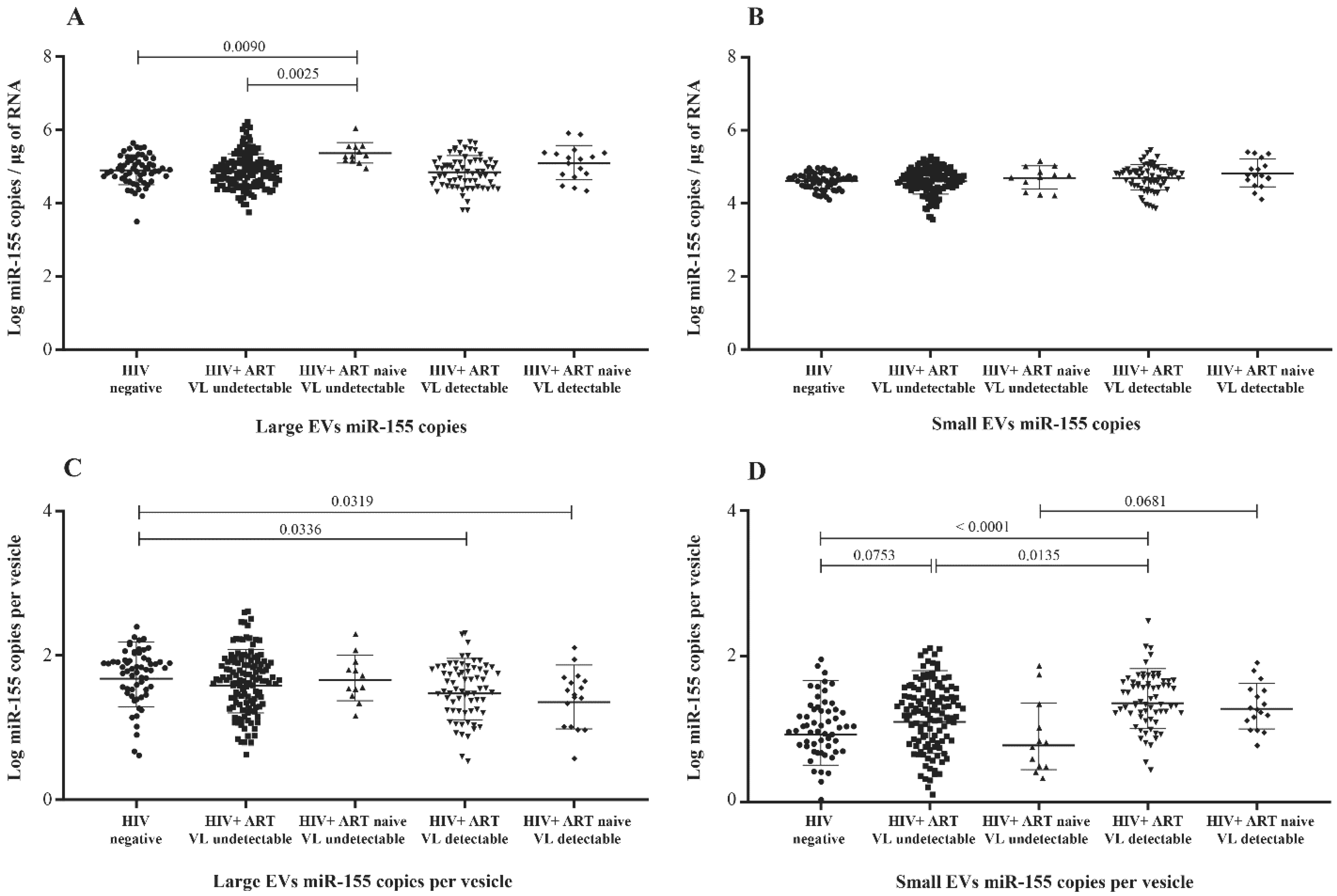
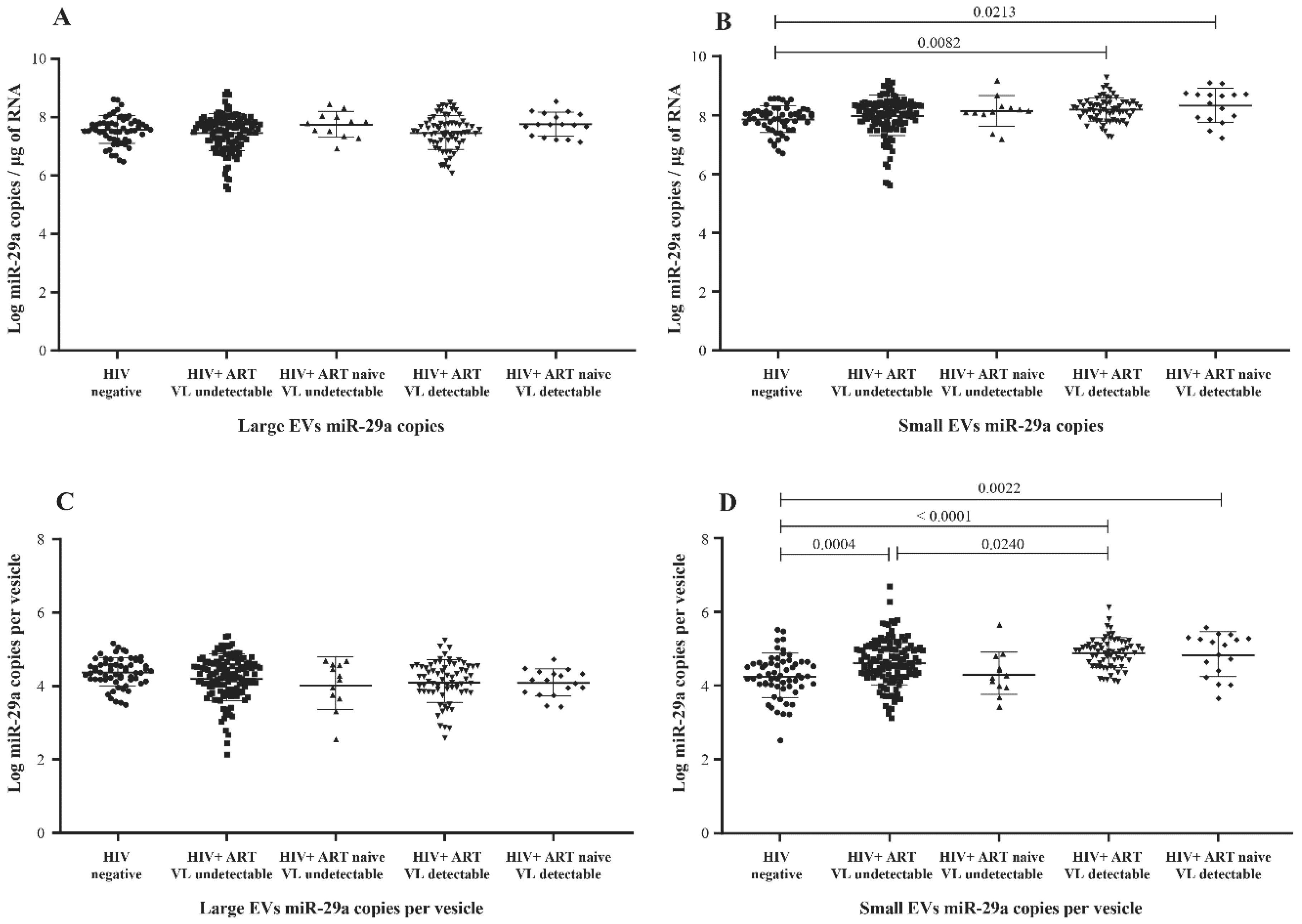
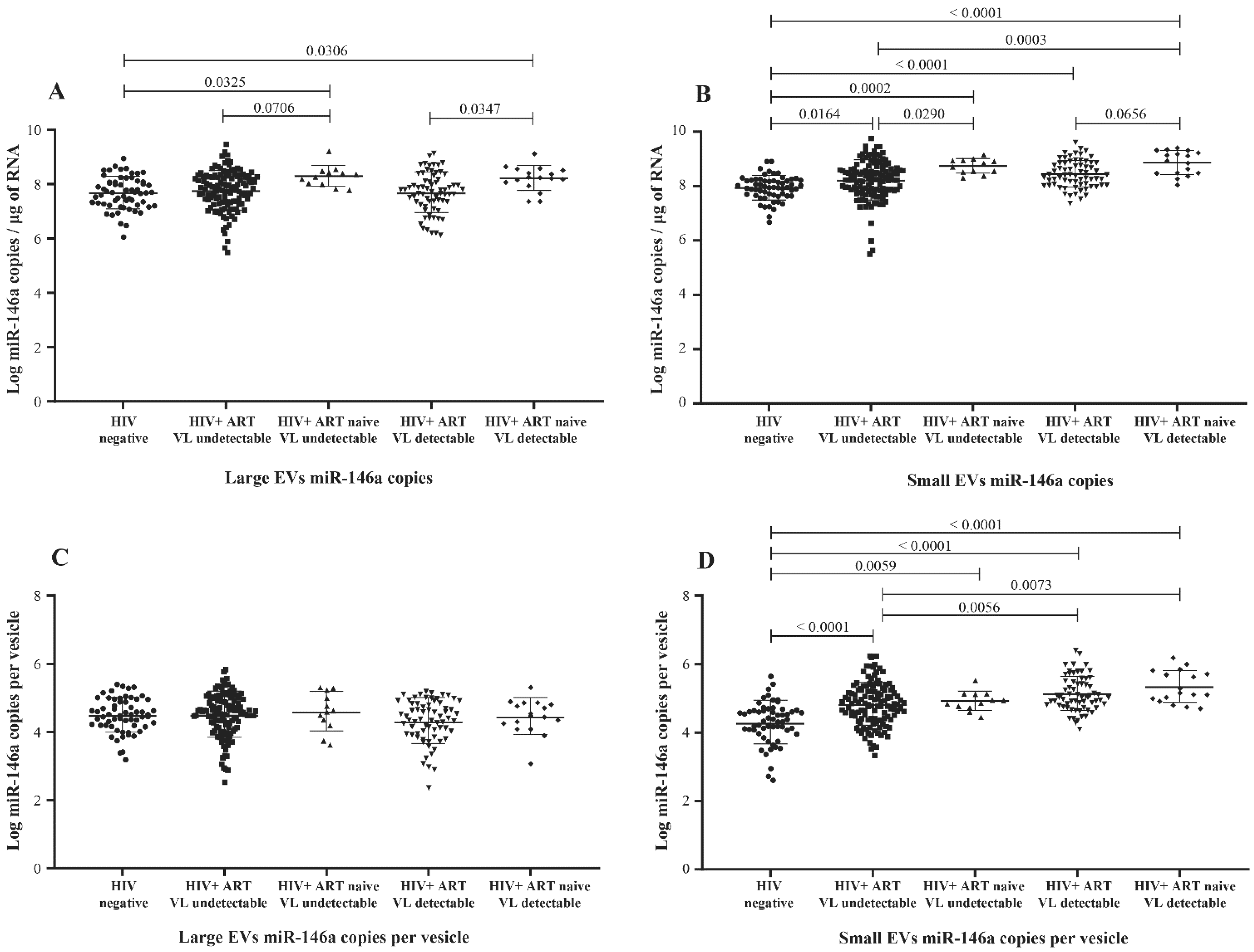
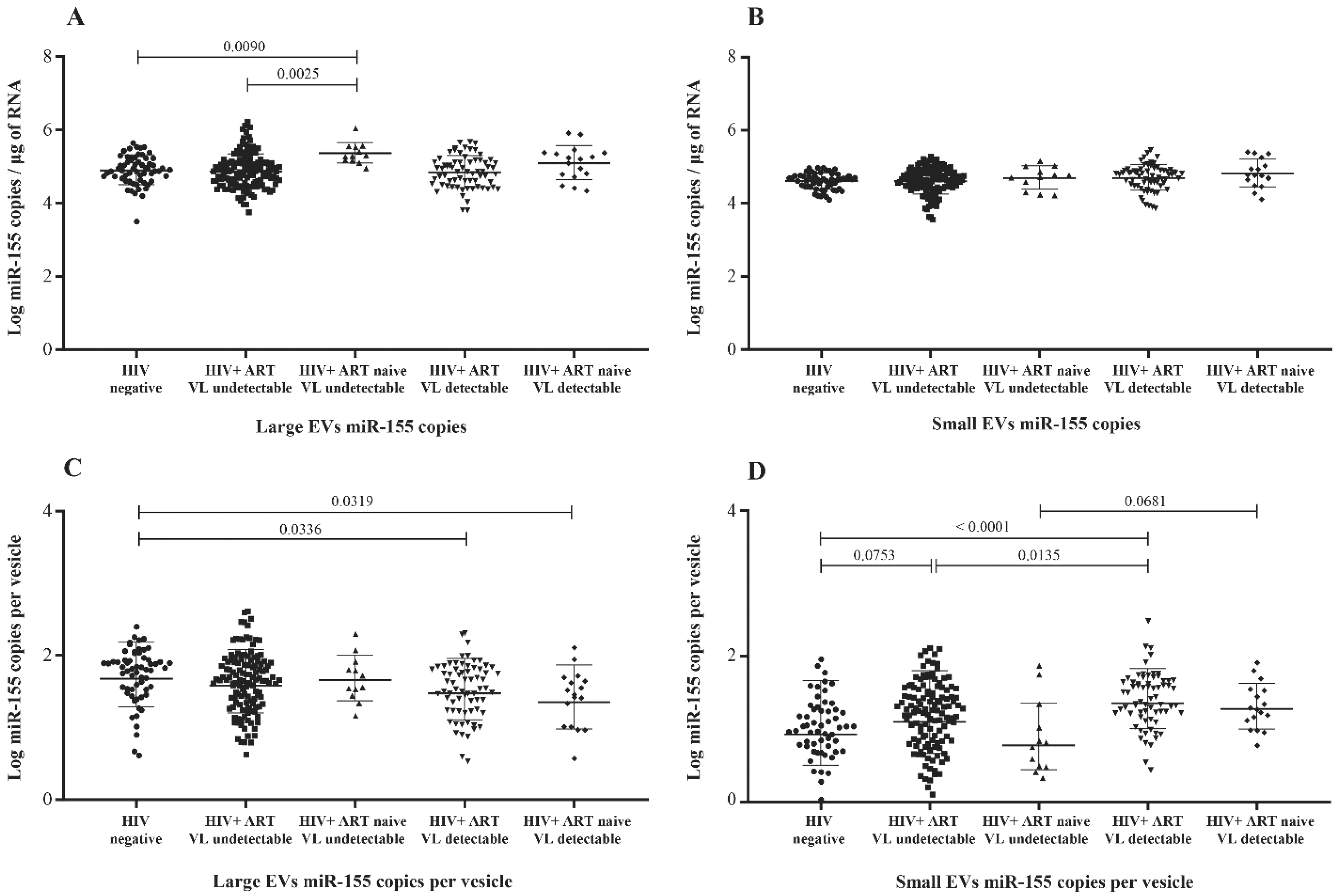
3.2. MicroRNA Content of Large versus Small EVs
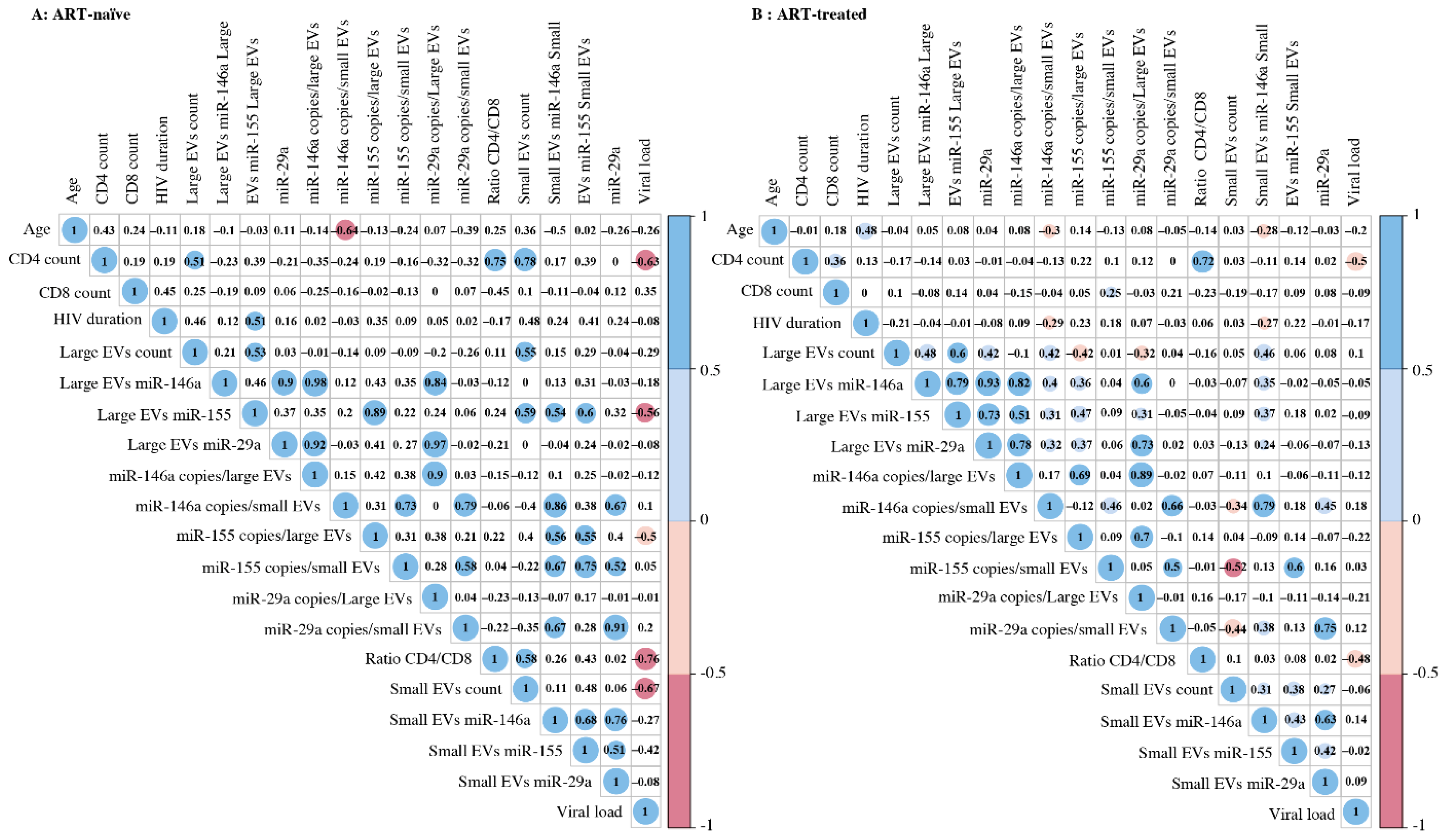
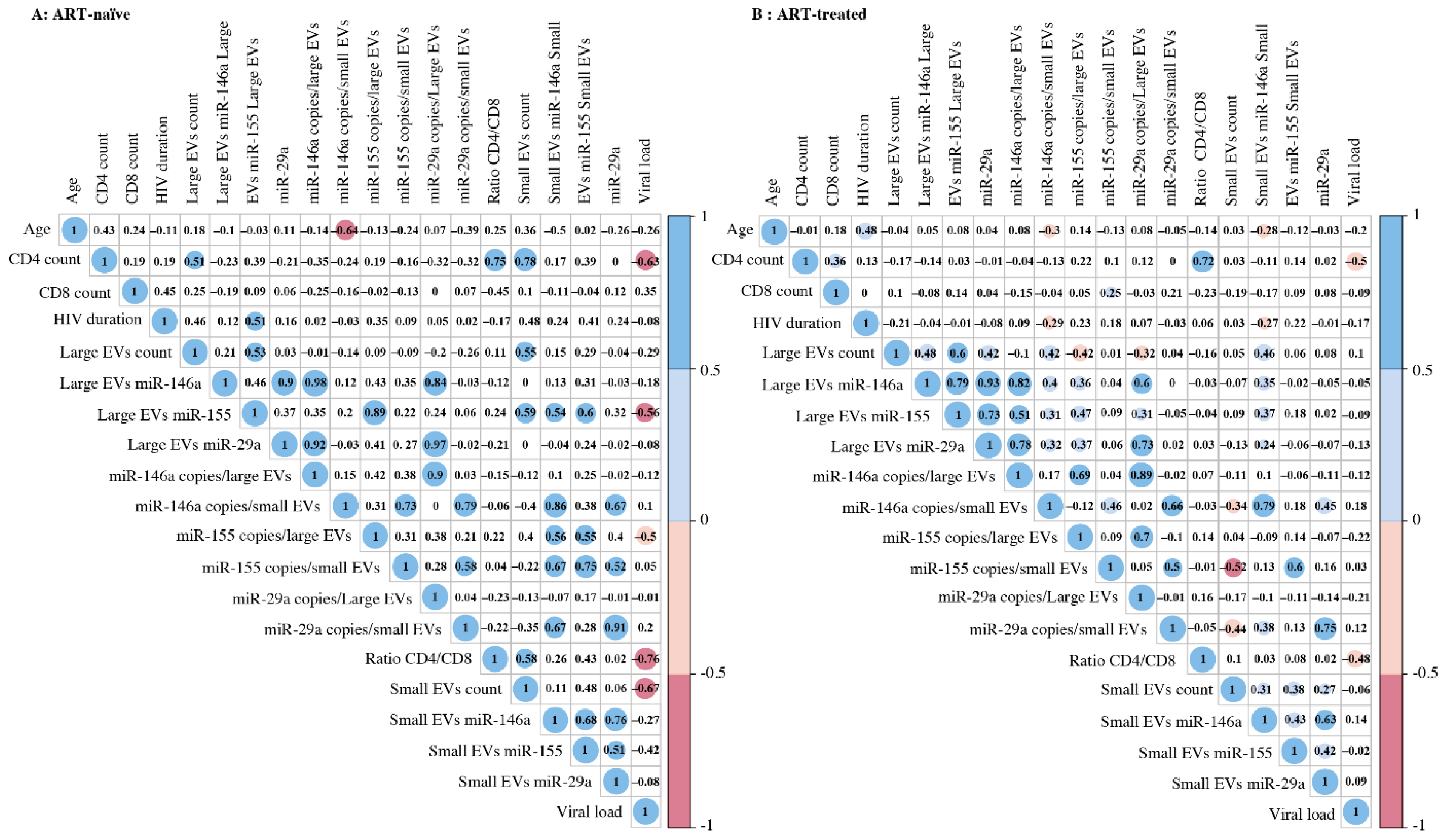
3.3. Correlation between EV MicroRNA Content and Demographic/Clinical Parameters
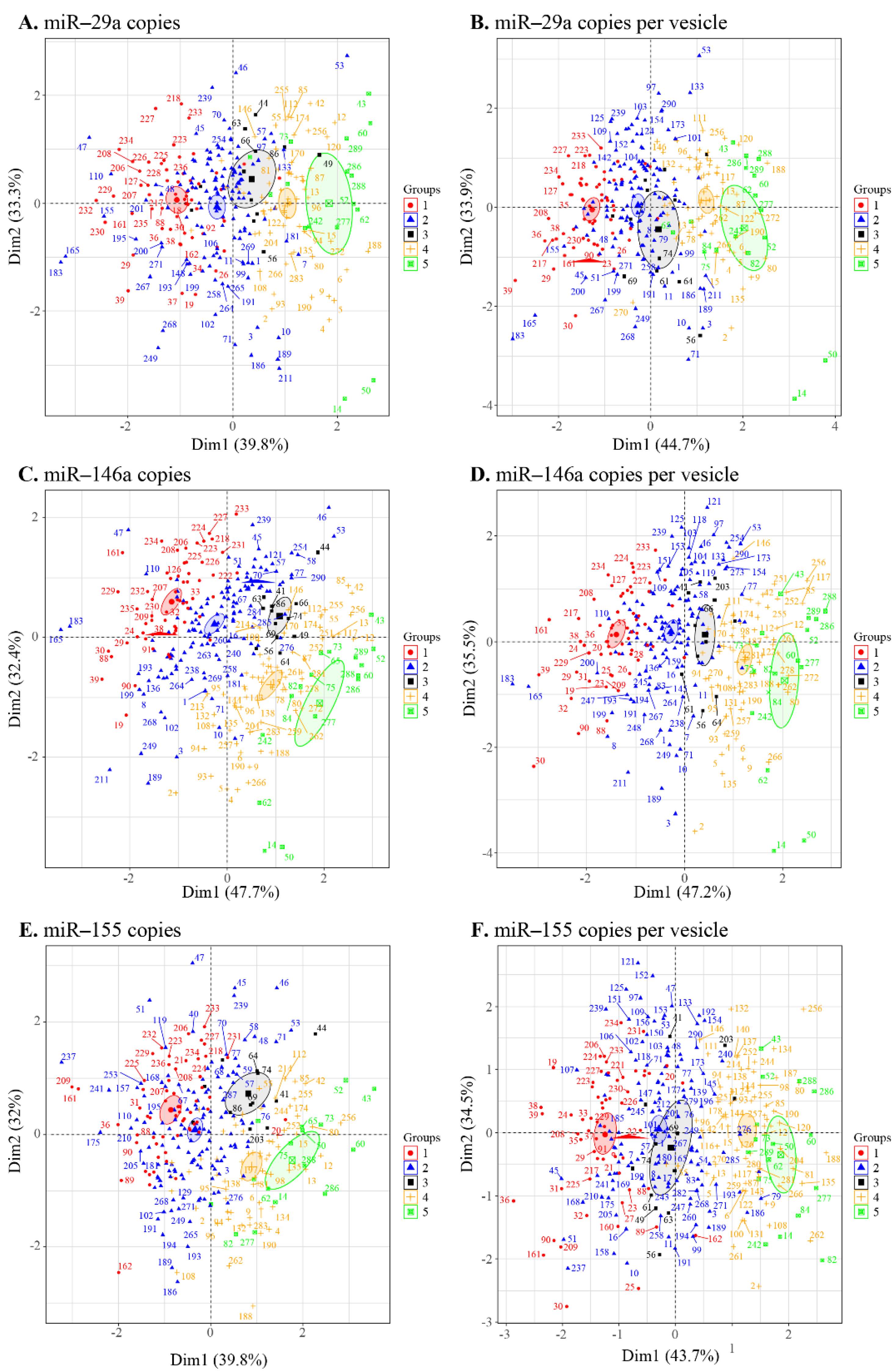
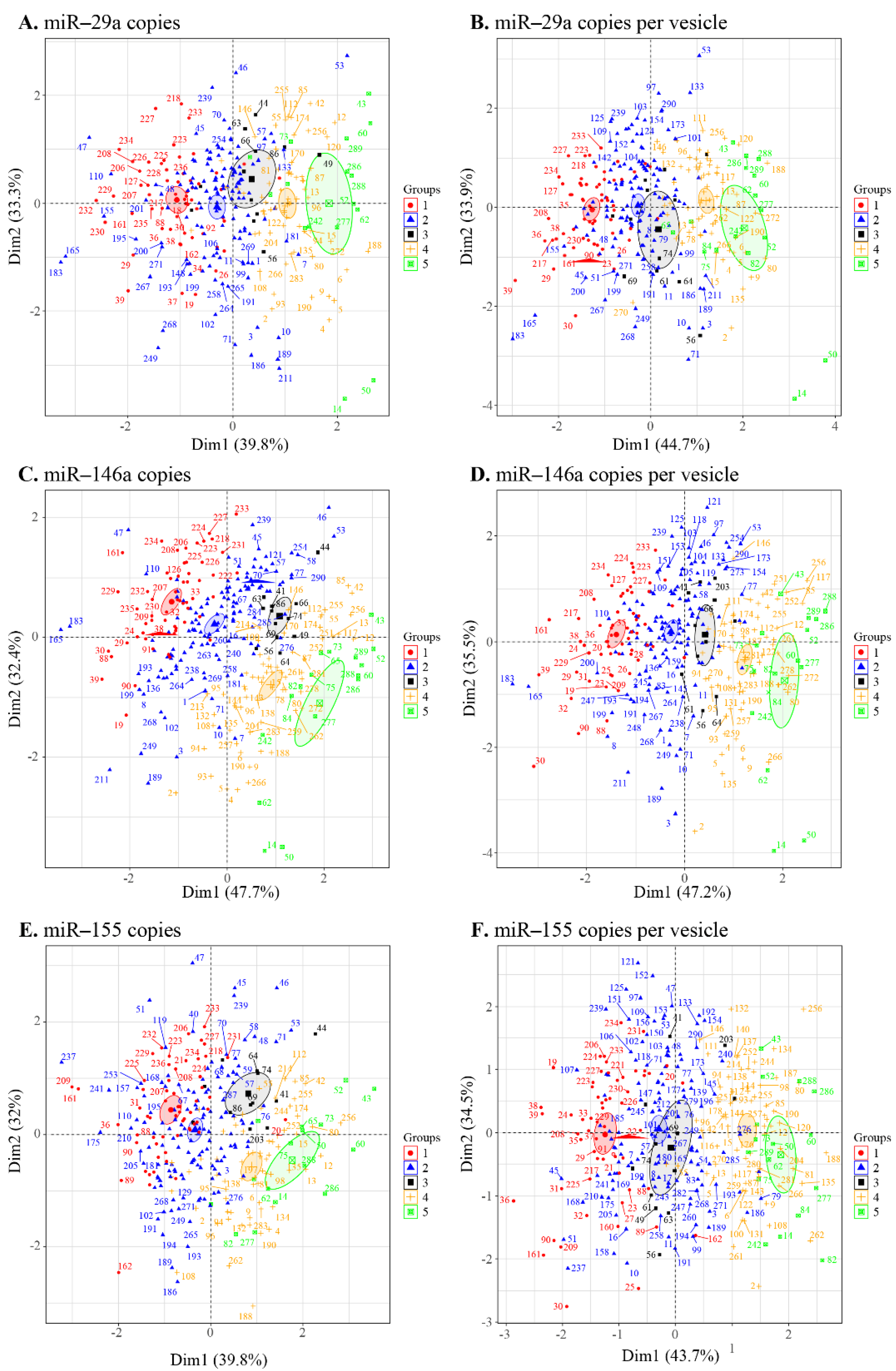
3.4. Principal Component Analysis of the EV Measurement/Viral Replication Association
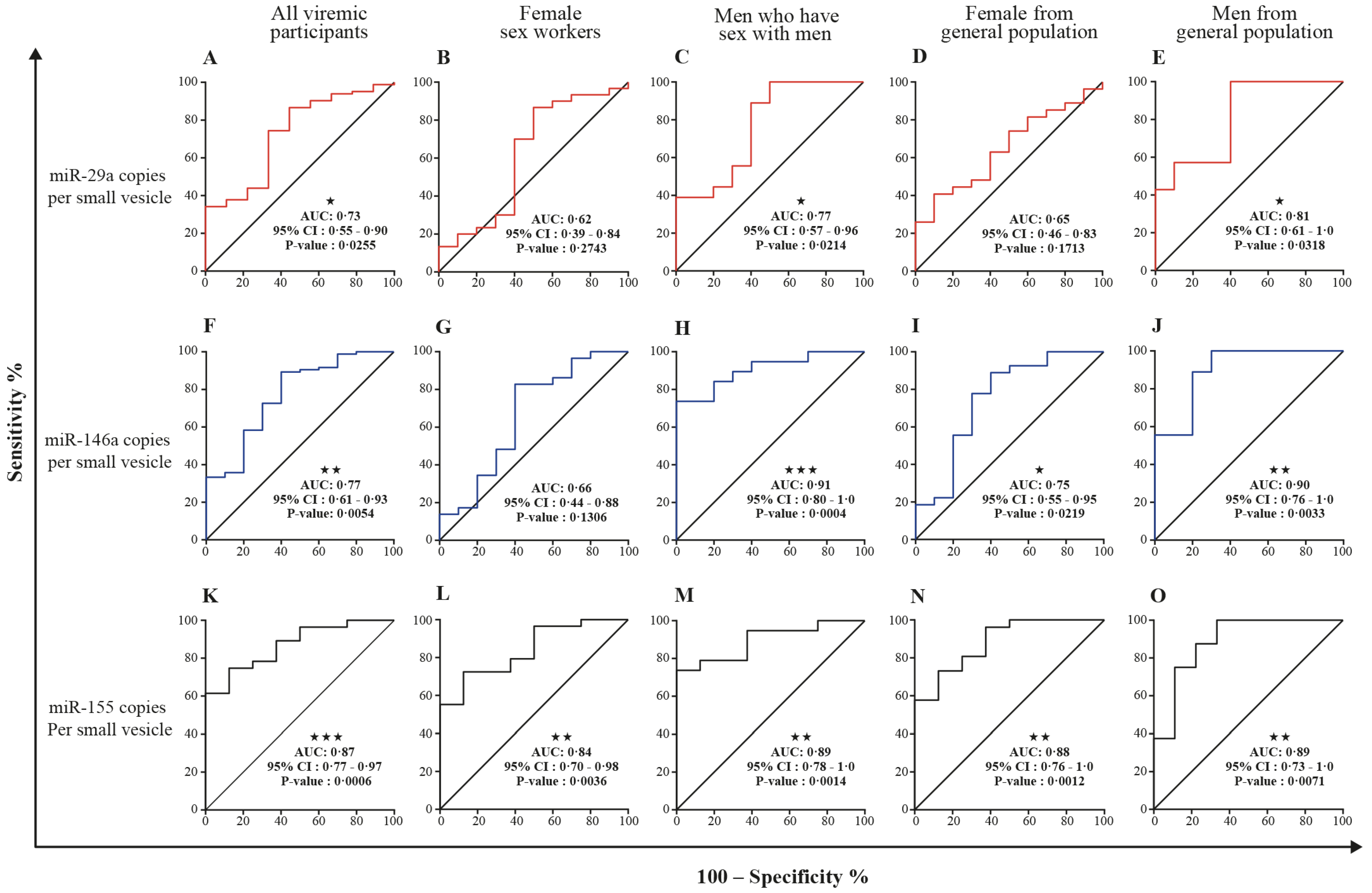
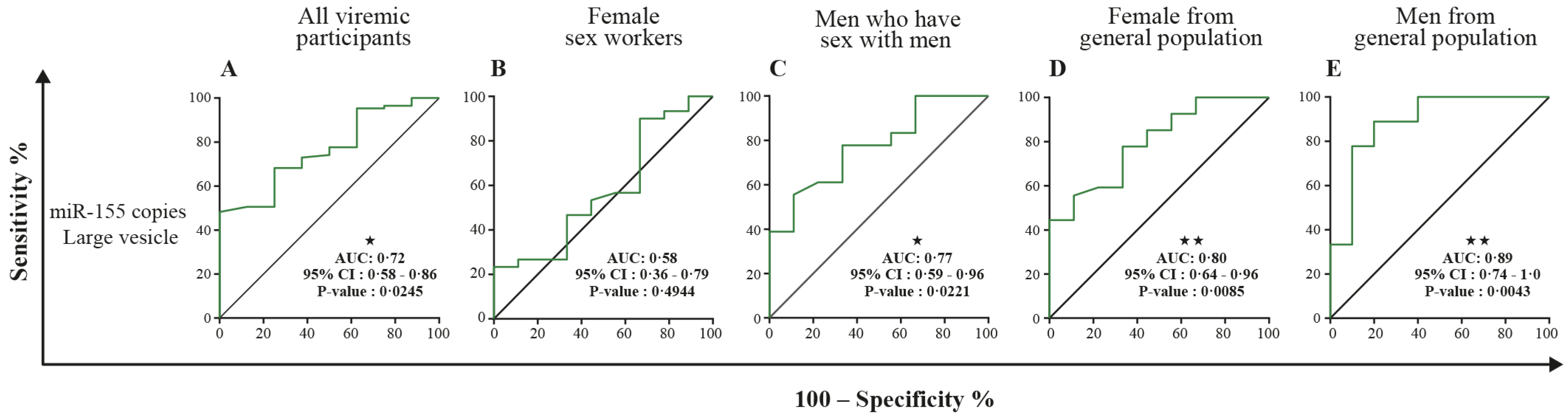
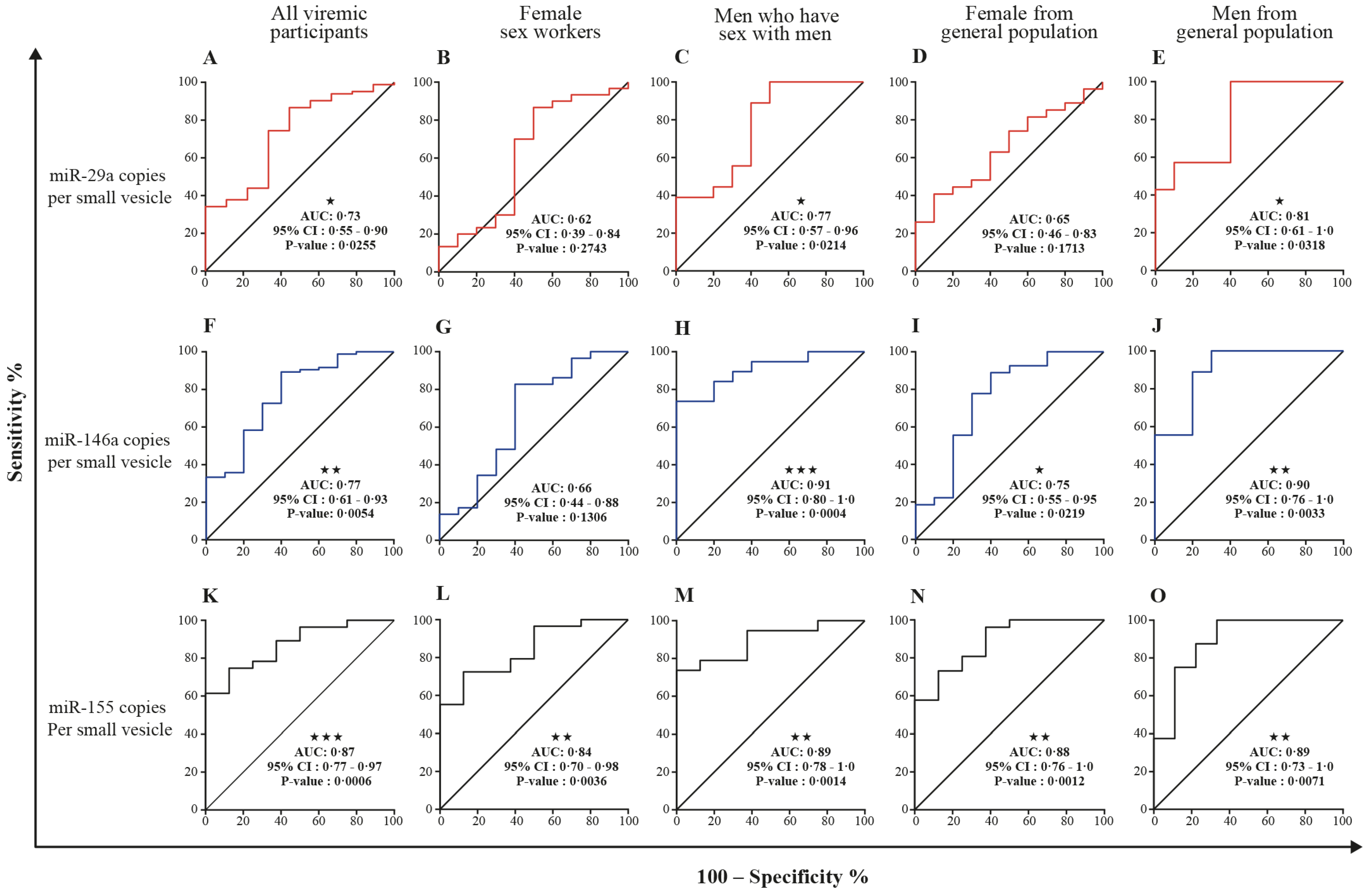
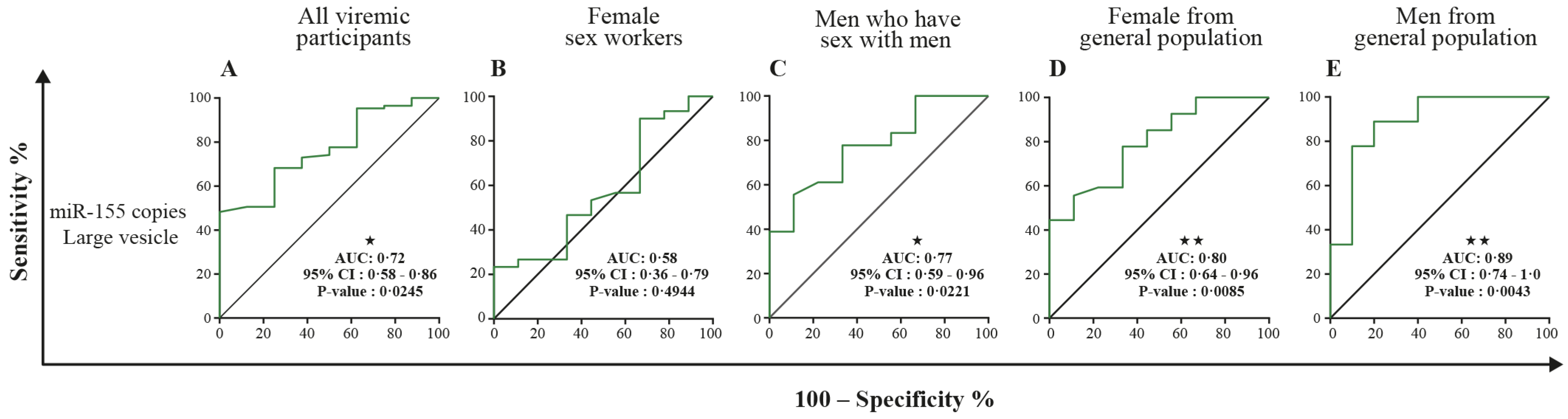
3.5. MicroRNA 29a, 146a, and 155 Copies per Small EV and Mir-155 Copies in Large EV as Potential Biomarkers of Viral Rebound
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierson, T.; McArthur, J.; Siliciano, R.F. Reservoirs for HIV-1: Mechanisms for viral persistence in the presence of antiviral immune responses and antiretroviral therapy. Annu. Rev. Immunol. 2000, 18, 665–708. [Google Scholar] [CrossRef]
- Aamer, H.A.; McClure, J.; Ko, D.; Maenza, J.; Collier, A.C.; Coombs, R.W.; Mullins, J.I.; Frenkel, L.M. Cells producing residual viremia during antiretroviral treatment appear to contribute to rebound viremia following interruption of treatment. PLoS Pathog. 2020, 16, e1008791. [Google Scholar] [CrossRef]
- Dinoso, J.B.; Kim, S.Y.; Wiegand, A.M.; Palmer, S.E.; Gange, S.J.; Cranmer, L.; O’Shea, A.; Callender, M.; Spivak, A.; Brennan, T.; et al. Treatment intensification does not reduce residual HIV-1 viremia in patients on highly active antiretroviral therapy. Proc. Natl. Acad. Sci. USA 2009, 106, 9403–9408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenhende, M.A.; Perrier, A.; Bonnet, F.; Lazaro, E.; Cazanave, C.; Reigadas, S.; Chêne, G.; Morlat, P. Risk of virological failure in HIV-1-infected patients experiencing low-level viraemia under active antiretroviral therapy (ANRS C03 cohort study). Antivir. Ther. 2015, 20, 655–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craw, J.A.; Beer, L.; Tie, Y.; Jaenicke, T.; Shouse, R.L.; Prejean, J. Viral Rebound Among Persons With Diagnosed HIV Who Achieved Viral Suppression, United States. J. Acquir. Immune Defic. Syndr. 2020, 84, 133–140. [Google Scholar] [CrossRef]
- Bernal, E.; Gómez, J.M.; Jarrín, I.; Cano, A.; Muñoz, A.; Alcaraz, A.; Imaz, A.; Iribarren, J.A.; Rivero, M.; Arazo, P.; et al. Low-Level Viremia Is Associated With Clinical Progression in HIV-Infected Patients Receiving Antiretroviral Treatment. J. Acquir. Immune Defic. Syndr. 2018, 78, 329–337. [Google Scholar] [CrossRef]
- Bing, A.; Hu, Y.; Prague, M.; Hill, A.L.; Li, J.Z.; Bosch, R.J.; De Gruttola, V.; Wang, R. Comparison of empirical and dynamic models for HIV viral load rebound after treatment interruption. Stat. Commun. Infect. Dis. 2020, 12, 20190021. [Google Scholar] [CrossRef] [PubMed]
- Conway, J.M.; Perelson, A.S.; Li, J.Z. Predictions of time to HIV viral rebound following ART suspension that incorporate personal biomarkers. PLoS Comput. Biol. 2019, 15, e1007229. [Google Scholar] [CrossRef]
- Hatano, H.; Jain, V.; Hunt, P.W.; Lee, T.H.; Sinclair, E.; Do, T.D.; Hoh, R.; Martin, J.N.; McCune, J.M.; Hecht, F.; et al. Cell-based measures of viral persistence are associated with immune activation and programmed cell death protein 1 (PD-1)-expressing CD4+ T cells. J. Infect. Dis. 2013, 208, 50–56. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatischeff, I. Current Search through Liquid Biopsy of Effective Biomarkers for Early Cancer Diagnosis into the Rich Cargoes of Extracellular Vesicles. Int. J. Mol. Sci. 2021, 22, 5674. [Google Scholar] [CrossRef] [PubMed]
- Poulet, G.; Massias, J.; Taly, V. Liquid Biopsy: General Concepts. Acta Cytol. 2019, 63, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Mathai, R.A.; Vidya, R.V.S.; Reddy, B.S.; Thomas, L.; Udupa, K.; Kolesar, J.; Rao, M. Potential Utility of Liquid Biopsy as a Diagnostic and Prognostic Tool for the Assessment of Solid Tumors: Implications in the Precision Oncology. J. Clin. Med. 2019, 8, 373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazié, W.W.; Boucher, J.; Vitry, J.; Goyer, B.; Routy, J.P.; Tremblay, C.; Trottier, S.; Jenabian, M.A.; Provost, P.; Alary, M.; et al. Plasma Extracellular Vesicle Subtypes May be Useful as Potential Biomarkers of Immune Activation in People with HIV. Pathog. Immun. 2021, 6, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Bazié, W.W.; Goyer, B.; Boucher, J.; Zhang, Y.; Planas, D.; Chatterjee, D.; Routy, J.P.; Alary, M.; Ancuta, P.; Gilbert, C. Diurnal Variation of Plasma Extracellular Vesicle Is Disrupted in People Living with HIV. Pathogens 2021, 10, 518. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Schierer, S.; Blume, K.; Dindorf, J.; Wittki, S.; Xiang, W.; Ostalecki, C.; Koliha, N.; Wild, S.; Schuler, G.; et al. HIV-Nef and ADAM17-Containing Plasma Extracellular Vesicles Induce and Correlate with Immune Pathogenesis in Chronic HIV Infection. EBioMedicine 2016, 6, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Mensah, G.A.; Al Sharif, S.; Pinto, D.O.; Branscome, H.; Yelamanchili, S.V.; Cowen, M.; Erickson, J.; Khatkar, P.; Mahieux, R.; et al. Extracellular Vesicles from Infected Cells Are Released Prior to Virion Release. Cells 2021, 10, 781. [Google Scholar] [CrossRef]
- Sadri Nahand, J.; Bokharaei-Salim, F.; Karimzadeh, M.; Moghoofei, M.; Karampoor, S.; Mirzaei, H.R.; Tabibzadeh, A.; Jafari, A.; Ghaderi, A.; Asemi, Z.; et al. MicroRNAs and exosomes: Key players in HIV pathogenesis. HIV Med. 2020, 21, 246–278. [Google Scholar] [CrossRef] [PubMed]
- Olivetta, E.; Chiozzini, C.; Arenaccio, C.; Manfredi, F.; Ferrantelli, F.; Federico, M. Extracellular vesicle-mediated intercellular communication in HIV-1 infection and its role in the reservoir maintenance. Cytokine Growth Factor Rev. 2020, 51, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Egaña-Gorroño, L.; Escribà, T.; Boulanger, N.; Guardo, A.C.; León, A.; Bargalló, M.E.; Garcia, F.; Gatell, J.M.; Plana, M.; Arnedo, M. Differential microRNA expression profile between stimulated PBMCs from HIV-1 infected elite controllers and viremic progressors. PLoS ONE 2014, 9, e106360. [Google Scholar] [CrossRef]
- Klase, Z.; Houzet, L.; Jeang, K.T. MicroRNAs and HIV-1: Complex interactions. J. Biol. Chem. 2012, 287, 40884–40890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nathans, R.; Chu, C.Y.; Serquina, A.K.; Lu, C.C.; Cao, H.; Rana, T.M. Cellular microRNA and P bodies modulate host-HIV-1 interactions. Mol. Cell 2009, 34, 696–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, P.; Ansari, M.Y.; Bapat, S.; Thakar, M.; Gangakhedkar, R.; Jameel, S. The microRNA miR-29a is associated with human immunodeficiency virus latency. Retrovirology 2014, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Ruelas, D.S.; Chan, J.K.; Oh, E.; Heidersbach, A.J.; Hebbeler, A.M.; Chavez, L.; Verdin, E.; Rape, M.; Greene, W.C. MicroRNA-155 Reinforces HIV Latency. J. Biol. Chem. 2015, 290, 13736–13748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroi, T.; Shibayama, M. Measurement of Particle Size Distribution in Turbid Solutions by Dynamic Light Scattering Microscopy. J. Vis. Exp. 2017, 9, e54885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Deun, J.; Mestdagh, P.; Agostinis, P.; Akay, Ö.; Anand, S.; Anckaert, J.; Martinez, Z.A.; Baetens, T.; Beghein, E.; Bertier, L.; et al. EV-TRACK: Transparent reporting and centralizing knowledge in extracellular vesicle research. Nat. Methods. 2017, 14, 228–232. [Google Scholar]
- Mfunyi, C.M.; Vaillancourt, M.; Vitry, J.; Nsimba Batomene, T.R.; Posvandzic, A.; Lambert, A.A.; Gilbert, C. Exosome release following activation of the dendritic cell immunoreceptor: A potential role in HIV-1 pathogenesis. Virology 2015, 484, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Hubert, A.; Subra, C.; Jenabian, M.A.; Tremblay Labrecque, P.F.; Tremblay, C.; Laffont, B.; Provost, P.; Routy, J.P.; Gilbert, C. Elevated Abundance, Size, and MicroRNA Content of Plasma Extracellular Vesicles in Viremic HIV-1+ Patients: Correlations With Known Markers of Disease Progression. J. Acquir. Immune Defic. Syndr. 2015, 70, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahand, J.S.; Karimzadeh, M.R.; Nezamnia, M.; Fatemipour, M.; Khatami, A.; Jamshidi, S.; Moghoofei, M.; Taghizadieh, M.; Hajighadimi, S.; Shafiee, A.; et al. The role of miR-146a in viral infection. IUBMB Life 2020, 72, 343–360. [Google Scholar] [CrossRef] [PubMed]
- DeMarino, C.; Pleet, M.L.; Cowen, M.; Barclay, R.A.; Akpamagbo, Y.; Erickson, J.; Ndembi, N.; Charurat, M.; Jumare, J.; Bwala, S.; et al. Antiretroviral Drugs Alter the Content of Extracellular Vesicles from HIV-1-Infected Cells. Sci. Rep. 2018, 8, 7653. [Google Scholar] [CrossRef] [PubMed]
- Hileman, C.O.; Funderburg, N.T. Inflammation, Immune Activation, and Antiretroviral Therapy in HIV. Curr. HIV/AIDS Rep. 2017, 14, 93–100. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Baixauli, F.; López-Otín, C.; Mittelbrunn, M. Exosomes and autophagy: Coordinated mechanisms for the maintenance of cellular fitness. Front. Immunol. 2014, 5, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahluwalia, J.K.; Khan, S.Z.; Soni, K.; Rawat, P.; Gupta, A.; Hariharan, M.; Scaria, V.; Lalwani, M.; Pillai, B.; Mitra, D.; et al. Human cellular microRNA hsa-miR-29a interferes with viral nef protein expression and HIV-1 replication. Retrovirology. 2008, 5, 117. [Google Scholar] [CrossRef] [Green Version]
- Balducci, E.; Leroyer, A.S.; Lacroix, R.; Robert, S.; Todorova, D.; Simoncini, S.; Lyonnet, L.; Chareyre, C.; Zaegel-Faucher, O.; Micallef, J.; et al. Extracellular vesicles from T cells overexpress miR-146b-5p in HIV-1 infection and repress endothelial activation. Sci Rep. 2019, 9, 10299. [Google Scholar] [CrossRef] [Green Version]
- Swaminathan, G.; Rossi, F.; Sierra, L.J.; Gupta, A.; Navas-Martín, S.; Martín-García, J. A role for microRNA-155 modulation in the anti-HIV-1 effects of Toll-like receptor 3 stimulation in macrophages. PLoS Pathog. 2012, 8, e1002937. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W.; Watson, A.K.; Blankson, J.N.; Clements, J.E. Relationships of PBMC microRNA expression, plasma viral load, and CD4+ T-cell count in HIV-1-infected elite suppressors and viremic patients. Retrovirology 2012, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Elton, T.S.; Selemon, H.; Elton, S.M.; Parinandi, N.L. Regulation of the MIR155 host gene in physiological and pathological processes. Gene 2013, 532, 1–12. [Google Scholar] [CrossRef]
- Jin, C.; Cheng, L.; Lu, X.; Xie, T.; Wu, H.; Wu, N. Elevated expression of miR-155 is associated with the differentiation of CD8+ T cells in patients with HIV-1. Mol. Med. Rep. 2017, 16, 1584–1589. [Google Scholar] [CrossRef] [PubMed]
- Lind, E.F.; Elford, A.R.; Ohashi, P.S. Micro-RNA 155 is required for optimal CD8+ T cell responses to acute viral and intracellular bacterial challenges. J. Immunol. 2013, 190, 1210–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 60) | HIV+ (n = 235) | p-Value | HIV+ | HIV+ Viral Load Detectable | |||||
|---|---|---|---|---|---|---|---|---|---|
| Viremic (n = 85) | ART > 6 Months, Viral Load Undetectable (n = 128) | p-Value | ART-Naïve (n = 17) | ART-Treated (n = 68) | p-Value | ||||
| Male, n (%) | 14 (23.3) | 68 (28.9) | 0.3880 | 28 (32.9) | 31 (24.2) | 0.1636 | 6 (35.3) | 22 (32.4) | 0.8175 |
| Female sex workers: n (%) | 96 (40.9) | 30 (35.3) | 63 (49.2) | 0.1208 | 2 (11.8) | 28 (41.2) | 0.0536 | ||
| Men who have sex with men: n (%) | 40 (17.0) | 19 (22.3) | 20 (15.6) | 4 (23.5) | 15 (22.1) | ||||
| Age (y) | 27 | 36 | <0.0001 | 35 | 38 | 0.0553 | 37 | 35 | 0.9577 |
| (22–32) | (30–44) | (30–42) | (32–45) | (27–4) | (30–42) | ||||
| <20 | 3 (5.0) | 5 (2.1) | <0.0001 | 2 (2.4) | 2 (1.6) | 0.2670 | 1 (5.9) | 1 (1.5) | 0.4993 |
| 20–29 | 34 (56.7) | 37 (15.7) | 17 (20.0) | 16 (12.5) | 4 (23.5) | 13 (19.1) | |||
| 30–39 | 19 (31.7) | 94 (40.0) | 37 (43.5) | 49 (38.3) | 5 (29.4) | 32 (47.0) | |||
| 40–49 | 4 (6.6) | 70 (29.8) | 20 (23.5) | 47 (36.7) | 5 (29.4) | 15 (22.1) | |||
| ≥ 50 | 29 (12.3) | 9 (10.6) | 14 (10.9) | 2 (11.8) | 7 (10.3) | ||||
| HIV duration (month) | NA | 36 (14–117) | 31 (8–72) | 55 (24–120) | 0.0313 | 8 (1–72) | 36 (12–72) | 0.9891 | |
| CD4 T cells/µL | 992 (794–1276) | 466 (330–696) | <0.0001 | 382 (213–561) | 513 (386–755) | <0.0001 | 583 (318–742) | 354 (188–507) | 0.0860 |
| CD8 T cells/µL | 584 (447–715) | 770 (543–1089) | <0.0001 | 841 (573–1049) | 736 (536–1093) | 0.6081 | 794 (570–1036) | 848 (544–1016) | 0.7852 |
| CD4/CD8 | 1.8 (1.4–2.1) | 0.6 (0.4–1.0) | <0.0001 | 0.5 (0.3–0.8) | 0.7 (0.5–1.0) | <0.0001 | 0.6 (0.4–1.0) | 0.4 (0.3–0.7) | 0.0185 |
| ART, n (%) | NA | 206 (87.7%) | 68 (80.0%) | NA | NA | NA | |||
| ART duration (month) | NA | 34 (13–85) | 24 (8–62) | 38 (21–96) | 0.0231 | NA | 24 (8–62) | ||
| Undetectable HIV viral load (<20 copies/mL), n (%) | NA | 150 (63.8) | NA | NA | NA | NA | |||
| HIV-1 viral load (copies/mL) | NA | 30,032 | 30,032 | NA | 17,330 | 1446 | |||
| NA | (108–33,978) | (108–33,978) | NA | (3,032–39,416) | (82–24,578) | 0.2958 | |||
| Group Used as Control | ||||||
|---|---|---|---|---|---|---|
| HIV-Negative | ART-Naïve, Undetectable Viral Load | Reference Patients | ||||
| AUC (95% CI) | p-Value | AUC (95% CI) | p-Value | AUC (95% CI) | p-Value | |
| miR-29a copies, large EVs | 0.52 (0.43–0.62) | 0.6006 | 0.63 (0.47–0.78) | 0.1548 | 0.58 (0.38–0.77) | 0.4274 |
| miR-29a copies, small EVs | 0.71 * (0.63–0.80) | <0.0001 | 0.60 (0.45–0.76) | 0.2686 | 0.50 (0.35–0.65) | 0.9600 |
| miR-29a copies per large EV | 0.65 (0.56–0.74) | 0.0016 | 0.50 (0.31–0.69) | 0.9911 | 0.54 (0.31–0.76) | 0.7099 |
| miR-29a copies per small EV | 0.80 (0.73–0.88) | <0.0001 | 0.88 (0.49–0.97) | <0.0001 | 0.73 (0.55–0.90) | 0.0255 |
| miR-146a copies, large EVs | 0.54 (0.44–0.63) | 0.4420 | 0.72 * (0.61–0.83) | 0.0133 | 0.60 (0.43–0.78) | 0.2914 |
| miR-146a copies, small EVs | 0.81 (0.74–0.88) | <0.0001 | 0.63 (0.52–0.75) | 0.1348 | 0.66 (0.48–0.83) | 0.1082 |
| miR-146a copies per large EV | 0.56 (0.46–0.65) | 0.2285 | 0.62 (0.45–0.80) | 0.1695 | 0.54 (0.34–0.74) | 0.6801 |
| miR-146a copies per small EV | 0.89 (0.83–0.94) | <0.0001 | 0.68 (0.55–0.81) | 0.0502 | 0.77 (0.61–0.93) | 0.0054 |
| miR-155 copies, large EVs | 0.51 (0.41–0.60) | 0.8914 | 0.80 * (0.70–0.90) | 0.0007 | 0.76 (0.63– 0.90) | 0.0084 |
| miR-155 copies, small EVs | 0.63 (0.54–0.72) | 0.0093 | 0.52 (0.35–0.69) | 0.8010 | 0.61 (0.46–0.75) | 0.2891 |
| miR-155 copies per large EV | 0.68 * (0.59–0.77) | 0.0002 | 0.62 (0.45–0.78) | 0.1674 | 0.52 (0.29–0.75) | 0.8367 |
| miR-155 copies per small EV | 0.76 (0.68–0.85) | <0.0001 | 0.97 (0.93–100) | <0.0001 | 0.87 (0.77–0.97) | 0.0006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazié, W.W.; Boucher, J.; Traoré, I.T.; Kania, D.; Somé, D.Y.; Alary, M.; Gilbert, C. Vesicular MicroRNA as Potential Biomarkers of Viral Rebound. Cells 2022, 11, 859. https://doi.org/10.3390/cells11050859
Bazié WW, Boucher J, Traoré IT, Kania D, Somé DY, Alary M, Gilbert C. Vesicular MicroRNA as Potential Biomarkers of Viral Rebound. Cells. 2022; 11(5):859. https://doi.org/10.3390/cells11050859
Chicago/Turabian StyleBazié, Wilfried Wenceslas, Julien Boucher, Isidore Tiandiogo Traoré, Dramane Kania, Diane Yirgnur Somé, Michel Alary, and Caroline Gilbert. 2022. "Vesicular MicroRNA as Potential Biomarkers of Viral Rebound" Cells 11, no. 5: 859. https://doi.org/10.3390/cells11050859
APA StyleBazié, W. W., Boucher, J., Traoré, I. T., Kania, D., Somé, D. Y., Alary, M., & Gilbert, C. (2022). Vesicular MicroRNA as Potential Biomarkers of Viral Rebound. Cells, 11(5), 859. https://doi.org/10.3390/cells11050859