
The Osteocyte Stimulated by Wnt Agonist SKL2001 Is a Safe Osteogenic Niche Improving Bioactivities in a Polycaprolactone and Cell Integrated 3D Module

 ,
,  ,
, 
Abstract
:
1. Introduction
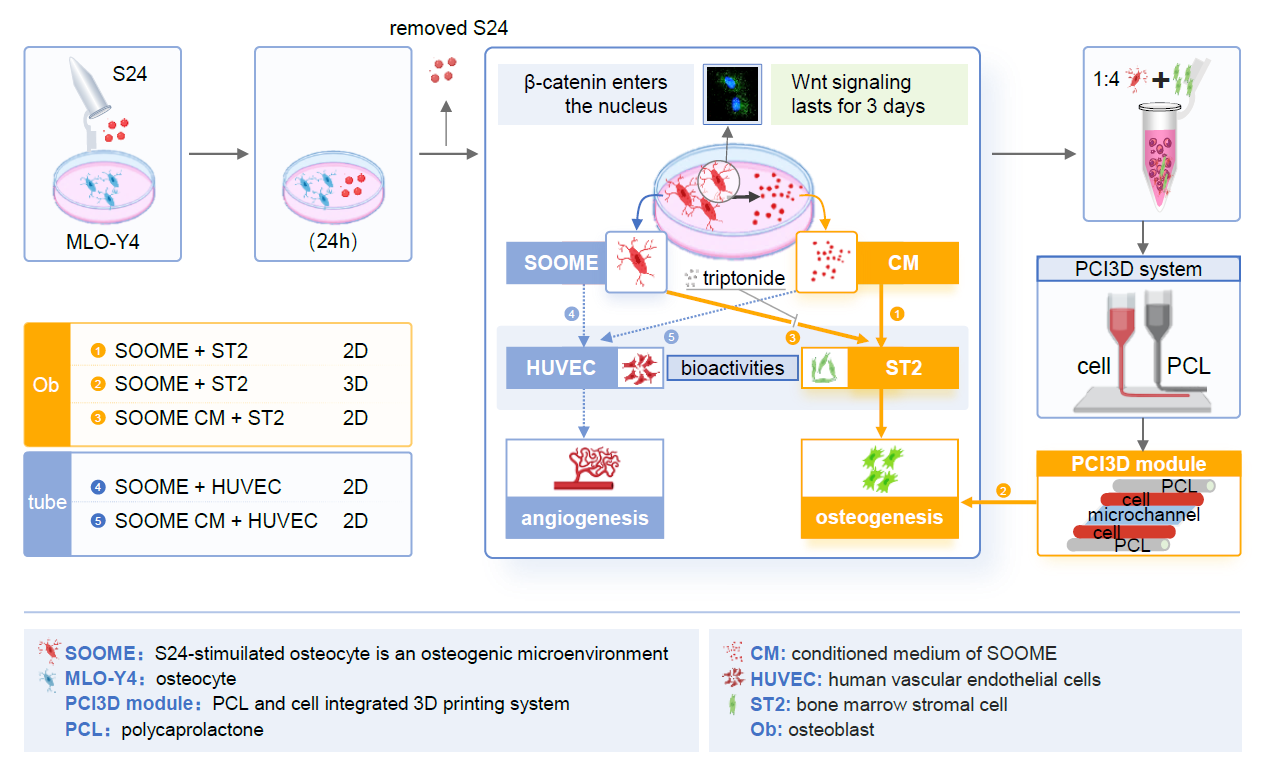
2. Materials and Methods
2.1. Reagents and Cells
2.2. Cell Culture
2.3. Activation and Inhibition of Wnt Signaling in Osteocytic MLO-Y4
2.4. PCL and Cell Integrated 3D Printing
2.5. Cell Viability Assay
2.6. Cell Proliferative Activity
2.7. RNA Extraction and Gene Expression Analysis
2.8. Alkaline Phosphatase Staining
2.9. AP Biochemical Activity Assay
2.10. Detection of Translocation of β-Catenin into the Nucleus
2.11. Mineralization Assay (Alizarin Red S Staining)
2.12. In Vitro HUVEC Tube Formation Assay
2.13. Conditioned Medium Preparation
2.14. Cell Migration Assay
2.15. Statistical Analysis
3. Results
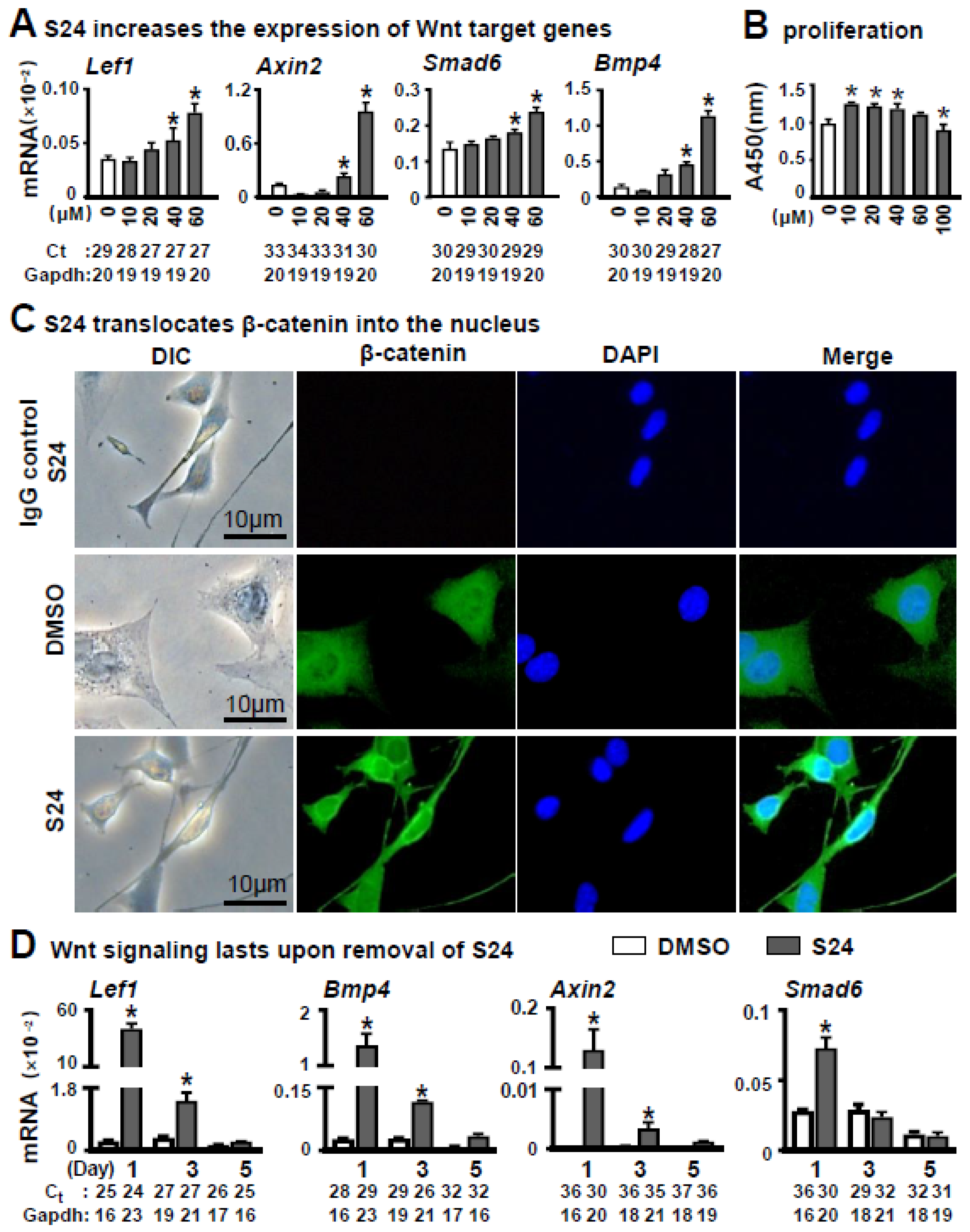
3.1. Effects of S24 on Activation of Wnt/β-Catenin Signaling in Osteocytic MLO-Y4
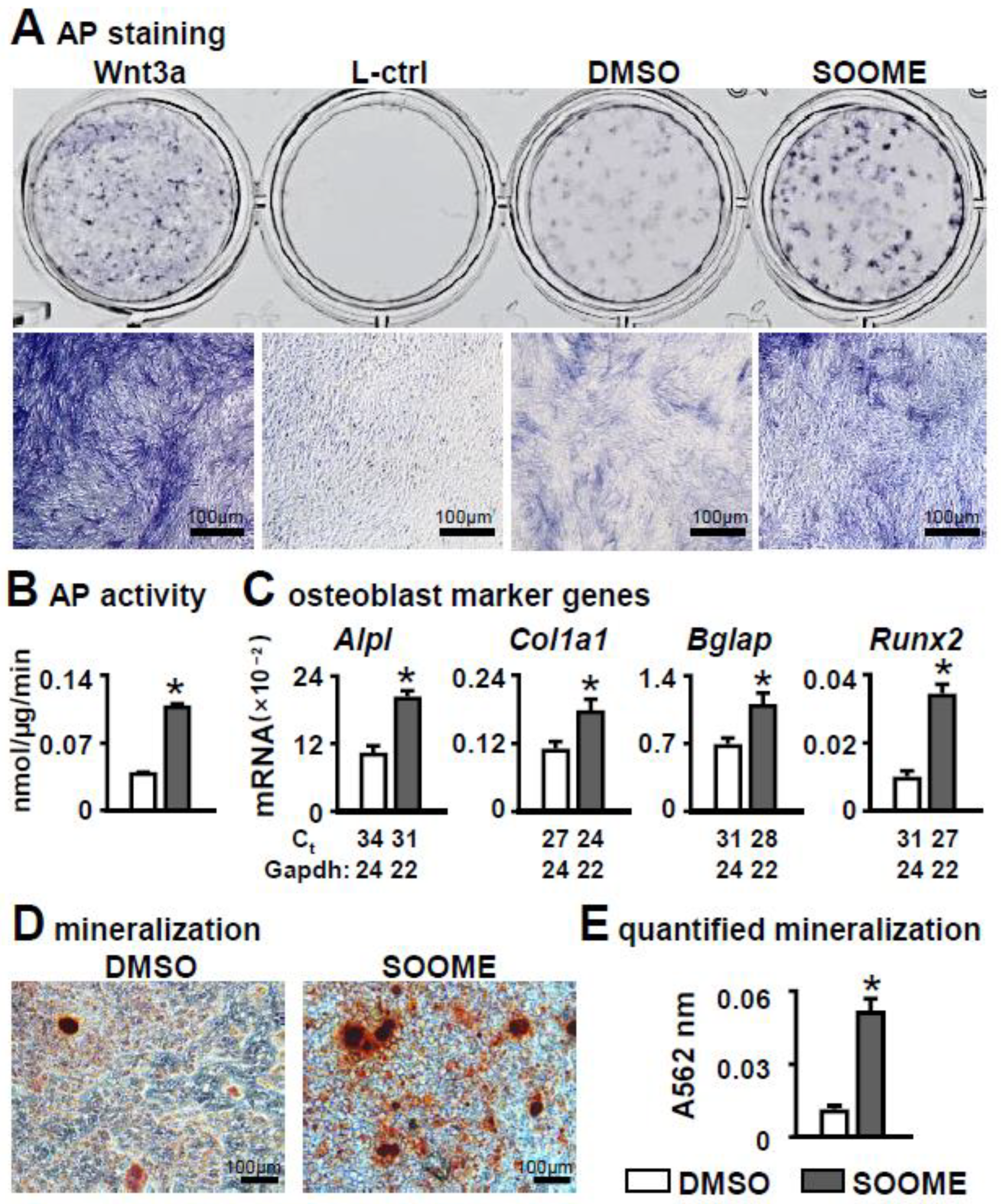
3.2. Effects of S24-Stimulated Osteocytes (SOOME) on Osteoblast Differentiation
3.3. Specificity of Wnt/β-Catenin Signaling in SOOME on Osteoblast Differentiation
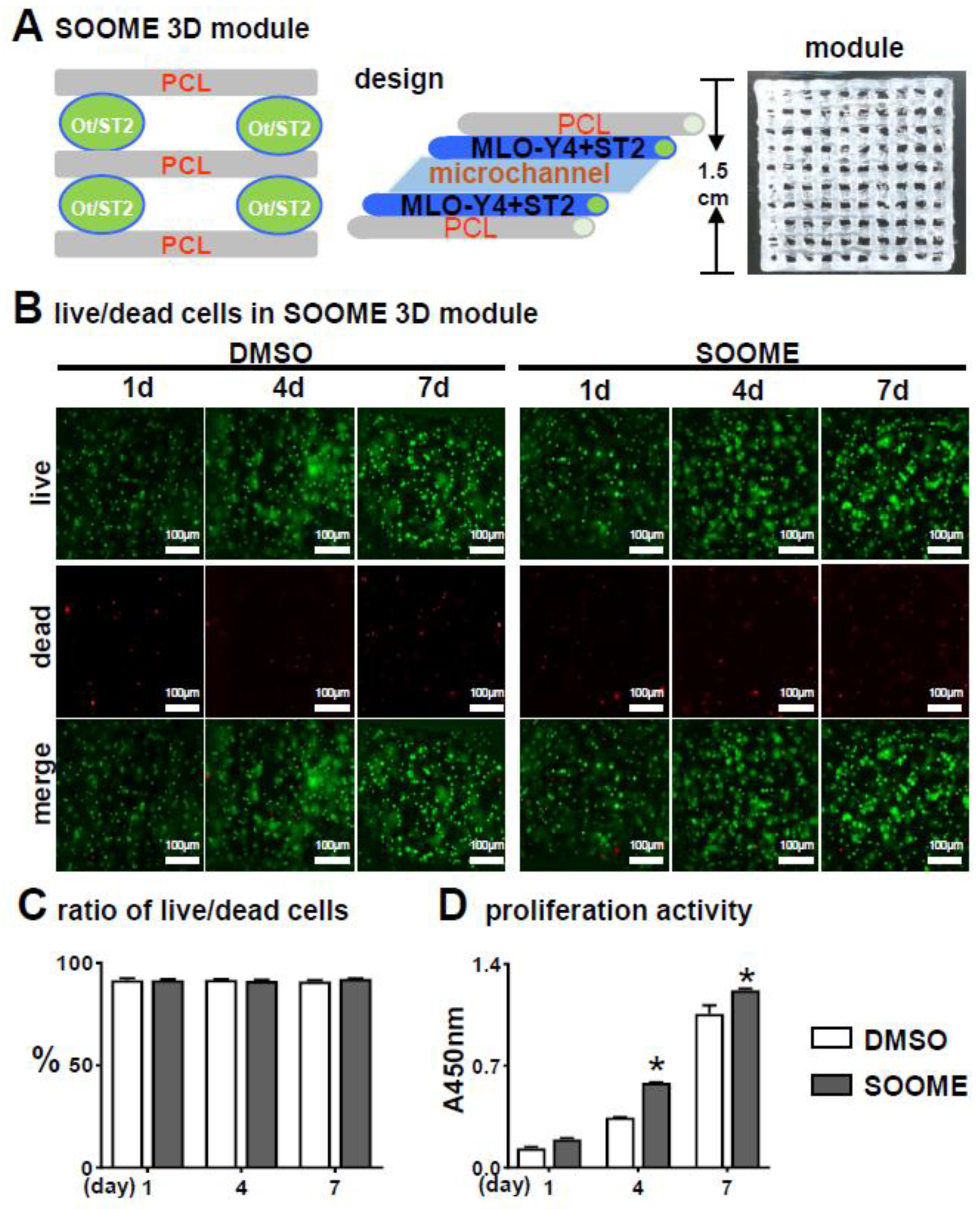
3.4. Effects of SOOME on Cell Viability and Proliferation in PCI3D Modules
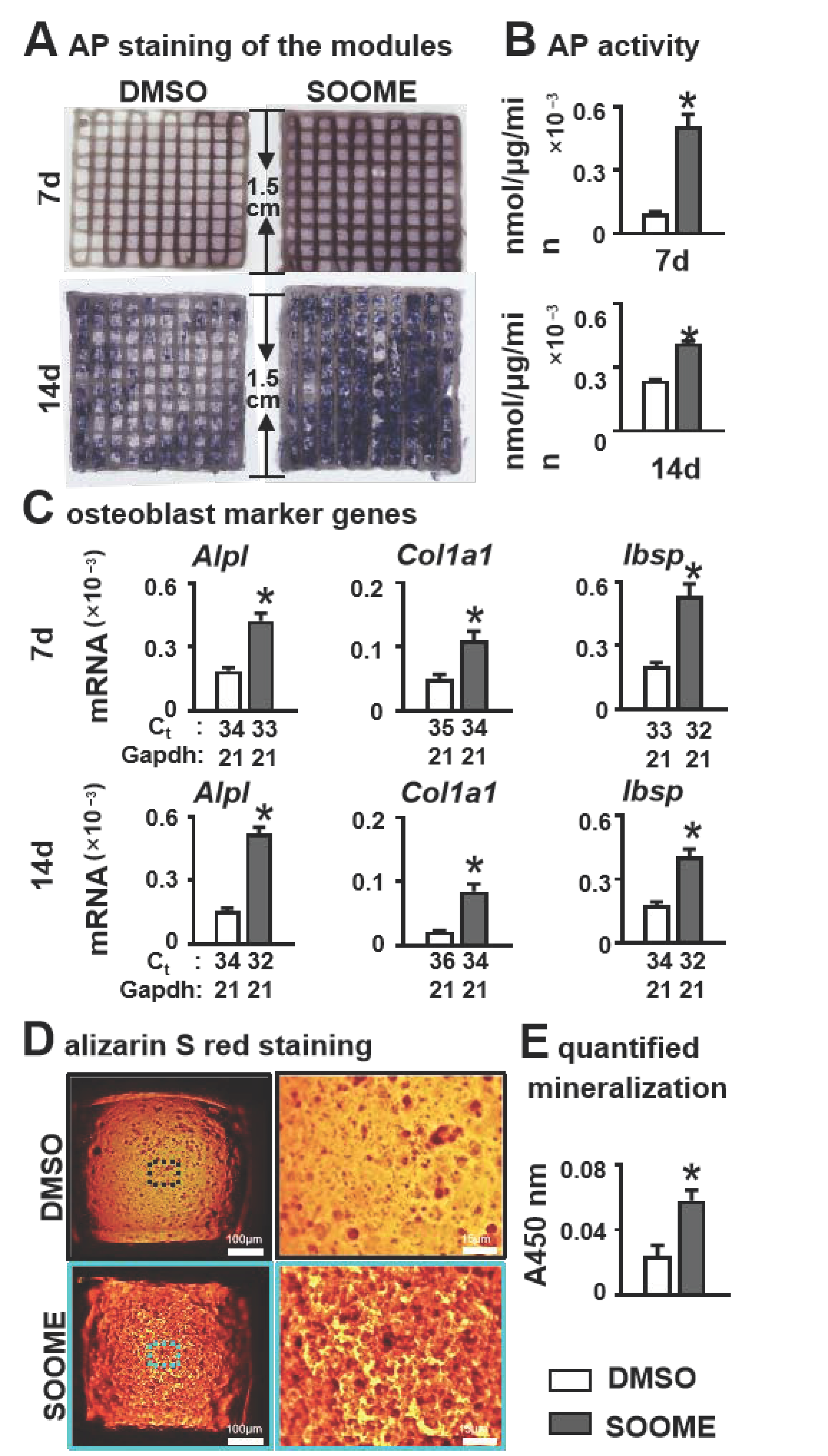
3.5. Effects of SOOME on Osteogenesis and Mineralization in PCI3D Modules
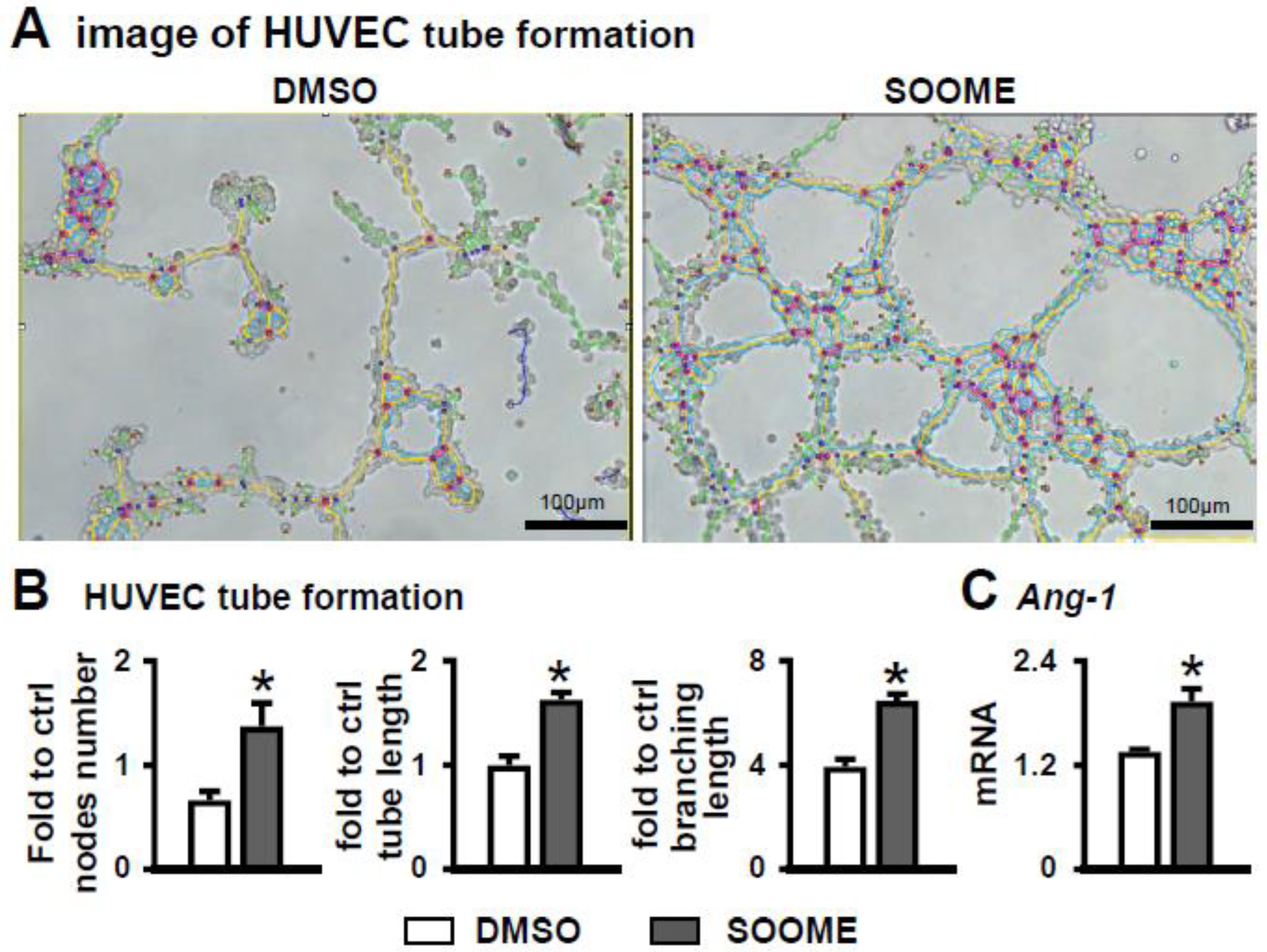
3.6. Effect of SOOME on Angiogenesis
3.7. Effect of Conditioned Medium of SOOME on Osteoblast Differentiation and Angiogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, M.; Zhao, S.C.; Chen, X.; Zhu, Y.F.; Zhang, C.Q. 3D-printed dimethyloxallyl glycine delivery scaffolds to improve angiogenesis and osteogenesis. Biomater. Sci. 2015, 3, 1236–1244. [Google Scholar]
- Perez, J.R.; Kouroupis, D.; Li, D.J.; Best, T.M.; Kaplan, L.; Correa, D. Tissue Engineering and Cell-Based Therapies for Fractures and Bone Defects. Front. Bioeng. Biotechnol. 2018, 6, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumant, O.; Shinde, S. Orthopedic Implants Market by Product Type (Reconstructive Joint Replacements, Spinal Implants, Dental Implants, Trauma, Orthobiologics, and Others), Biomaterial (Metallic, Ceramic, Polymeric, and Others), and Type (Knee, Hip, Wrist & Shoulder, Dental, Spine, Ankle, and Others). Available online: https://www.alliedmarketresearch.com/press-release/orthopedic-implants-market.html (accessed on 24 January 2022).
- Li, J.; Cui, X.L.; Hooper, G.J.; Lim, K.S.; Woodfield, T.B.F. Rational design, bio-functionalization and biological performance of hybrid additive manufactured titanium implants for orthopaedic applications: A review. J. Mech. Behav. Biomed. Mater. 2020, 105, 103671. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.L.; Li, J.; Hartanto, Y.; Durham, M.; Tang, J.N.; Zhang, H.; Hooper, G.; Lim, K.; Woodfield, T. Advances in Extrusion 3D Bioprinting: A Focus on Multicomponent Hydrogel-Based Bioinks. Adv. Healthc. Mater. 2020, 9, 1901648. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.F.; Huang, Y.; Schilling, A.F.; Hubbell, K.; Cui, X.F. Organ Bioprinting: Are We There Yet? Adv. Healthc. Mater. 2018, 7, 1701018. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Hu, N.; Liu, J.; Zhai, X.; Wu, M.; Hu, C.; Li, L.; Lai, Y.; Pan, H.; Lu, W.W.; et al. Three-Dimensional Printing of Biodegradable Piperazine-Based Polyurethane-Urea Scaffolds with Enhanced Osteogenesis for Bone Regeneration. ACS Appl. Mater. Interfaces 2019, 11, 9415–9424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, G.J.; Johnson, B.N.; Jia, X.F. Three-dimensional (3D) printed scaffold and material selection for bone repair. Acta Biomater. 2019, 84, 16–33. [Google Scholar] [CrossRef]
- Hao, Z.C.; Song, Z.H.; Huang, J.; Huang, K.; Panetta, A.; Gu, Z.P.; Wu, J. The scaffold microenvironment for stem cell based bone tissue engineering. Biomater. Sci. 2017, 5, 1382–1392. [Google Scholar] [CrossRef]
- Zhou, F.F.; Hong, Y.; Liang, R.J.; Zhang, X.Z.; Liao, Y.G.; Jiang, D.M.; Zhang, J.Y.; Sheng, Z.X.; Xie, C.; Peng, Z.; et al. Rapid printing of bio-inspired 3D tissue constructs for skin regeneration. Biomaterials 2020, 258, 120287. [Google Scholar] [CrossRef]
- Hann, S.Y.; Cui, H.T.; Esworthy, T.; Zhou, X.; Lee, S.-j.; Plesniak, M.W.; Zhang, L.J.G. Dual 3D printing for vascularized bone tissue regeneration. Acta Biomater. 2021, 123, 263–274. [Google Scholar] [CrossRef]
- Murphy, S.V.; Atala, A. 3D bioprinting of tissues and organs. Nat. Biotechnol. 2014, 32, 773–785. [Google Scholar] [CrossRef]
- Zhuang, P.; Ng, W.L.; An, J.; Chua, C.K.; Tan, L.P. Layer-by-layer ultraviolet assisted extrusion-based (UAE) bioprinting of hydrogel constructs with high aspect ratio for soft tissue engineering applications. PLoS ONE 2019, 14, e0216776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.S.; Jang, J.; Chae, S.H.; Gao, G.; Kong, J.-S.; Ahn, M.J.; Cho, D.-W. Three-dimensional bioprinting of cell-laden constructs with polycaprolactone protective layers for using various thermoplastic polymers. Biofabrication 2016, 8, 035013. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.H.; Shi, W.; Wu, S.H.; Kuss, M.; Jiang, X.P.; Untrauer, J.B.; Reid, S.P.; Duan, B. 3D printed composite scaffolds with dual small molecule delivery for mandibular bone regeneration. Biofabrication 2020, 12, 035020. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Shi, Y.F.; Zhang, X.; Ma, J. Evaluation of BMP-2 and VEGF loaded 3D printed hydroxyapatite composite scaffolds with enhanced osteogenic capacity in vitro and in vivo. Mater. Sci. Eng. C. Mate.R Biol. Appl. 2020, 112, 110893. [Google Scholar] [CrossRef]
- Chimene, D.; Kaunas, R.; Gaharwar, A.K. Hydrogel Bioink Reinforcement for Additive Manufacturing: A Focused Review of Emerging Strategies. Adv. Mater. (Weinh. Ger.) 2020, 32, 1902026. [Google Scholar] [CrossRef]
- Masaeli, R.; Zandsalimi, K.; Rasoulianboroujeni, M.; Tayebi, L. Challenges in Three-Dimensional Printing of Bone Substitutes. Tissue. Eng. Part B Rev. 2019, 25, 387–397. [Google Scholar] [CrossRef]
- Zhu, G.Y.; Zhang, T.X.; Chen, M.; Yao, K.; Huang, X.Q.; Zhang, B.; Li, Y.Z.; Liu, J.; Wang, Y.B.; Zhao, Z.H. Bone physiological microenvironment and healing mechanism: Basis for future bone-tissue engineering scaffolds. Bioact. Mater. 2021, 6, 4110–4140. [Google Scholar] [CrossRef]
- Riquelme, M.A.; Cardenas, E.R.; Jiang, J.X. Osteocytes and Bone Metastasis. Front. Endocrinol. 2020, 11, 567844. [Google Scholar] [CrossRef]
- Kitaura, H.; Marahleh, A.; Ohori, F.; Noguchi, T.; Shen, W.R.; Qi, J.; Nara, Y.; Pramusita, A.; Kinjo, R.; Mizoguchi, I. Osteocyte-Related Cytokines Regulate Osteoclast Formation and Bone Resorption. Int. J. Mol. Sci. 2020, 21, 5169. [Google Scholar] [CrossRef]
- Zhang, J.; Wehrle, E.; Rubert, M.; Muller, R. 3D Bioprinting of Human Tissues: Biofabrication, Bioinks, and Bioreactors. Int. J. Mol. Sci. 2021, 22, 3971. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.M.; Panadero Pérez, J.A.; Kumar, V.; Verbruggen, A.S.K.; McNamara, L.M. A Novel 3D Osteoblast and Osteocyte Model Revealing Changes in Mineralization and Pro-osteoclastogenic Paracrine Signaling During Estrogen Deficiency. Front. Bioeng. Biotechnol. 2020, 8, 601. [Google Scholar] [CrossRef] [PubMed]
- Weivoda, M.M.; Youssef, S.J.; Oursler, M.J. Sclerostin expression and functions beyond the osteocyte. Bone 2017, 96, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonewald, L.F. The amazing osteocyte. J. Bone Miner. Res. 2011, 26, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Sapir-Koren, R.; Livshits, G. Osteocyte control of bone remodeling: Is sclerostin a key molecular coordinator of the balanced bone resorption–formation cycles? Osteoporos Int. 2014, 25, 2685–2700. [Google Scholar] [CrossRef]
- Wodarz, A.; Nusse, R. MECHANISMS OF WNT SIGNALING IN DEVELOPMENT. Annu. Rev. Cell Dev. Biol. 1998, 14, 59–88. [Google Scholar] [CrossRef] [Green Version]
- Osorio, J. Bone. Osteocyte-specific activation of the canonical Wnt-beta catenin pathway stimulates bone formation. Nat. Rev. Endocrinol. 2015, 11, 192. [Google Scholar] [CrossRef]
- Tu, X.L.; Delgado-Calle, J.; Condon, K.W.; Maycas, M.; Zhang, H.J.; Carlesso, N.; Taketo, M.M.; Burr, D.B.; Plotkin, L.I.; Bellido, T. Osteocytes mediate the anabolic actions of canonical Wnt/β-catenin signaling in bone. Proc. Natl. Acad. Sci. USA 2015, 112, E478. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.Y.; Liu, F.Y.; Liu, Y.Y.; Liu, X.L.; Ai, Z.Y.; Guo, Z.K.; Zhang, Y. GSK3 inhibitors CHIR99021 and 6-bromoindirubin-3′-oxime inhibit microRNA maturation in mouse embryonic stem cells. Sci. Rep. 2015, 5, 8666. [Google Scholar] [CrossRef] [Green Version]
- Gwak, J.; Hwang, S.G.; Park, H.-S.; Choi, S.R.; Park, S.-H.; Kim, H.; Ha, N.-C.; Bae, S.J.; Han, J.-K.; Kim, D.-E.; et al. Small molecule-based disruption of the Axin/β-catenin protein complex regulates mesenchymal stem cell differentiation. Cell Res. 2012, 22, 237–247. [Google Scholar] [CrossRef] [Green Version]
- Narcisi, R.; Arikan, O.H.; Lehmann, J.; ten Berge, D.; van Osch, G.J.V.M. Differential Effects of Small Molecule WNT Agonists on the Multilineage Differentiation Capacity of Human Mesenchymal Stem Cells. Tissue Eng. Part. A 2016, 22, 1264–1273. [Google Scholar] [CrossRef] [PubMed]
- Stern, A.R.; Stern, M.M.; Van Dyke, M.E.; Jähn, K.; Prideaux, M.; Bonewald, L.F. Isolation and culture of primary osteocytes from the long bones of skeletally mature and aged mice. BioTechniques 2012, 52, 361–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryynänen, J.; Kriebitzsch, C.; Meyer, M.B.; Janssens, I.; Pike, J.W.; Verlinden, L.; Verstuyf, A. Class 3 semaphorins are transcriptionally regulated by 1,25(OH)2D3 in osteoblasts. J. Steroid Biochem. Mol. Biol. 2017, 173, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Tu, X.L.; Joeng, K.S.; Nakayama, K.I.; Nakayama, K.; Rajagopal, J.; Carroll, T.J.; McMahon, A.P.; Long, F.X. Noncanonical Wnt Signaling through G Protein-Linked PKCδ Activation Promotes Bone Formation. Dev. Cell. 2007, 12, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xu, M.; Ren, Q.; Liu, G.; Meng, S.L.; Xiahou, K.; Zhang, Y.X.; Jiang, N.; Zhou, W.X. Human Induced Pluripotent Stem Cell-Derived Neural Cells from Alzheimer’s Disease Patients Exhibited Different Susceptibility to Oxidative Stress. Stem Cells Dev. 2020, 29, 1444–1456. [Google Scholar] [CrossRef]
- Chiang, C.C.; Hsieh, M.K.; Wang, C.Y.; Tuan, W.H.; Lai, P.L. Cytotoxicity and cell response of preosteoblast in calcium sulfate-augmented PMMA bone cement. Biomed. Mater. 2021, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-Y.; Sun, H. Up-regulation of Foxp3 inhibits cell proliferation, migration and invasion in epithelial ovarian cancer. Cancer Lett. 2010, 287, 91–97. [Google Scholar] [CrossRef]
- Tu, X.; Rhee, Y.; Condon, K.W.; Bivi, N.; Allen, M.R.; Dwyer, D.; Stolina, M.; Turner, C.H.; Robling, A.G.; Plotkin, L.I. Sost downregulation and local Wnt signaling are required for the osteogenic response to mechanical loading. Bone 2012, 50, 209–217. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Jiao, G.; Dong, M.; Chi, H.; Wang, H.; Wu, W.; Liu, H.; Ren, S.; Kong, M.; Li, C.; et al. Orthosilicic Acid Accelerates Bone Formation in Human Osteoblast-Like Cells Through the PI3K–Akt–mTOR Pathway. Biol. Trace Elem. Res. 2019, 190, 327–335. [Google Scholar] [CrossRef]
- Griffin, J.N.; del Viso, F.; Duncan, A.R.; Robson, A.; Hwang, W.; Kulkarni, S.; Liu, K.J.; Khokha, M.K. RAPGEF5 Regulates Nuclear Translocation of β-Catenin. Dev. Cell 2018, 44, 248–260.e244. [Google Scholar] [CrossRef] [Green Version]
- Venugopal, J.R.; Dev, V.R.G.; Senthilram, T.; Sathiskumar, D.; Gupta, D.; Ramakrishna, S. Osteoblast mineralization with composite nanofibrous substrate for bone tissue regeneration. Cell Biol. Int. 2011, 35, 73–80. [Google Scholar] [PubMed]
- Jia, L.Y.; Zhou, X.Y.; Huang, X.J.; Xu, X.H.; Jia, Y.H.; Wu, Y.T.; Yao, J.L.; Wu, Y.M.; Wang, K. Maternal and umbilical cord serum-derived exosomes enhance endothelial cell proliferation and migration. FASEB J. 2018, 32, 4534–4543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertacchini, J.; Magarò, M.S.; Potì, F.; Palumbo, C. Osteocytes Specific GSK3 Inhibition Affects In Vitro Osteogenic Differentiation. Biomedicines 2018, 6, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinyoles, M.; Del Valle-Pérez, B.; Curto, J.; Viñas-Castells, R.; Alba-Castellón, L.; García de Herreros, A.; Duñach, M. Multivesicular GSK3 Sequestration upon Wnt Signaling Is Controlled by p120-Catenin/Cadherin Interaction with LRP5/6. Mol. Cell. 2014, 53, 444–457. [Google Scholar] [CrossRef] [Green Version]
- Yuste, I.; Luciano, F.C.; González-Burgos, E.; Lalatsa, A.; Serrano, D.R. Mimicking bone microenvironment: 2D and 3D in vitro models of human osteoblasts. Pharmacol. Res. 2021, 169, 105626. [Google Scholar] [CrossRef]
- Kang, H.-W.; Lee, S.J.; Ko, I.K.; Kengla, C.; Yoo, J.J.; Atala, A. A 3D bioprinting system to produce human-scale tissue constructs with structural integrity. Nat. Biotechnol. 2016, 34, 312–319. [Google Scholar] [CrossRef]
- Lieben, L. The future of 3D printing of human tissues is taking shape. Nat. Rev. Rheumatol. 2016, 12, 191. [Google Scholar] [CrossRef]
- Pashuck, E.T.; Stevens, M. From clinical imaging to implantation of 3D printed tissues. Nat. Biotechnol. 2016, 34, 295–296. [Google Scholar] [CrossRef]
- Alcala-Orozco, C.R.; Mutreja, I.; Cui, X.; Hooper, G.J.; Lim, K.S.; Woodfield, T.B. Hybrid biofabrication of 3D osteoconductive constructs comprising Mg-based nanocomposites and cell-laden bioinks for bone repair. Bone 2022, 154, 116198. [Google Scholar] [CrossRef]
- Marsell, R.; Sisask, G.; Nilsson, Y.; Sundgren-Andersson, A.K.; Andersson, U.; Larsson, S.; Nilsson, O.; Ljunggren, O.; Jonsson, K.B. GSK-3 inhibition by an orally active small molecule increases bone mass in rats. Bone 2012, 50, 619–627. [Google Scholar] [CrossRef]
- Yan, D.-Y.; Tang, J.H.; Chen, L.; Wang, B.Z.; Weng, S.J.; Xie, Z.J.; Wu, Z.-Y.; Shen, Z.J.; Bai, B.L.; Yang, L. Imperatorin promotes osteogenesis and suppresses osteoclast by activating AKT/GSK3 β/β-catenin pathways. J. Cell. Mol. Med. 2020, 24, 2330–2341. [Google Scholar] [CrossRef] [PubMed]
- Antika, L.D.; Lee, E.-J.; Kim, Y.-H.; Kang, M.-K.; Park, S.-H.; Kim, D.Y.; Oh, H.; Choi, Y.-J.; Kang, Y.-H. Dietary phlorizin enhances osteoblastogenic bone formation through enhancing β-catenin activity via GSK-3β inhibition in a model of senile osteoporosis. J. Nutr. Biochem. 2017, 49, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Atala, A.; Bauer, S.B.; Saker, S.; Yoo, J.J.; Retik, A.B. 196: Tissue-Engineered Autologous Bladders for Patients Needing Cystoplasty. J. Urol. 2007, 177, 66. [Google Scholar] [CrossRef]
- Zerlin, M.; Julius, M.A.; Kitajewski, J. Wnt/Frizzled signaling in angiogenesis. Angiogenesis 2008, 11, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Reis, M.; Liebner, S. Wnt signaling in the vasculature. Exp. Cell Res. 2013, 319, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.F.; Chen, H.; Zhang, H.B.; Guo, C.J.; Yang, K.; Chen, K.Z.; Cheng, R.Y.; Qian, N.D.; Sandler, N.; Zhang, Y.S.; et al. Vascularized 3D printed scaffolds for promoting bone regeneration. Biomaterials 2019, 190, 97–110. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward | Reverse |
|---|---|---|
| Gapdh | GCACAGTCAAGGCCGAGAAT | GCCTTCTCCATGGTGGTGAA |
| beta-actin | AGAGGGAAATCGTGCGTGAC | CCATACCCAAGAAGGAAGGCT |
| Lef1 | TACCCCAGCCAGTGTCAACA | TCCATGATAGGCTTTGATGACTTTC |
| Axin2 | TGCAGGAGGCGGTACAGTTC | GCTGGAAGTGGTAAAGCAGCTT |
| Bmp4 | GAGGAGTTTCCATCACGAAGA | GCTCTGCCGAGGAGATCA |
| Smad6 | AAGATGCTGAAGCCGTTGGT | CGAACTCCAGTATCTCCGCTTT |
| Alpl | CACGGCGTCCATGAGCAGAAC | CAGGCACAGTGGTCAAGGTTGG |
| Runx2 | CCGGTCTCCTTCCAGGAT | GGGAACTGCTGTGGCTTC |
| Osx | CCCTTCTCAAGCACCAATGG | AAGGGTGGGTAGTCA TTTGCA TA |
| Bglap | CAGCGGCCCTGAGTCTGA | GCCGGAGTCTGTTCACTACCTTA |
| Col1a1 | GACAGGCGAACAAGGTGACAGAG | CAGGAGAACCAGGAGAACCAGGAG |
| Ibsp | CAGAGGAGGCAAGCGTCACT | GCTGTCTGGGTGCCAACACT |
| Ang-1 | ACTAGTAGTACAATGACAGTTTTCCTTTCC | AGATCTTCAAAAGTCCAAGGGCCGGATCAT |
| EP2 | GATGAAGCAACCAGAGCAGAC | CAGAGAGGACTCCCACATGAA |
| EP4 | GCCCTCTCCTGCCAATATAAC | TTTCAACACTTTGGCCTGAAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Ruan, X.; Li, J.; Wang, B.; Chen, J.; Wang, X.; Wang, P.; Tu, X. The Osteocyte Stimulated by Wnt Agonist SKL2001 Is a Safe Osteogenic Niche Improving Bioactivities in a Polycaprolactone and Cell Integrated 3D Module. Cells 2022, 11, 831. https://doi.org/10.3390/cells11050831
Liu Y, Ruan X, Li J, Wang B, Chen J, Wang X, Wang P, Tu X. The Osteocyte Stimulated by Wnt Agonist SKL2001 Is a Safe Osteogenic Niche Improving Bioactivities in a Polycaprolactone and Cell Integrated 3D Module. Cells. 2022; 11(5):831. https://doi.org/10.3390/cells11050831
Chicago/Turabian StyleLiu, Yangxi, Xiaojie Ruan, Jun Li, Bo Wang, Jie Chen, Xiaofang Wang, Pengtao Wang, and Xiaolin Tu. 2022. "The Osteocyte Stimulated by Wnt Agonist SKL2001 Is a Safe Osteogenic Niche Improving Bioactivities in a Polycaprolactone and Cell Integrated 3D Module" Cells 11, no. 5: 831. https://doi.org/10.3390/cells11050831
APA StyleLiu, Y., Ruan, X., Li, J., Wang, B., Chen, J., Wang, X., Wang, P., & Tu, X. (2022). The Osteocyte Stimulated by Wnt Agonist SKL2001 Is a Safe Osteogenic Niche Improving Bioactivities in a Polycaprolactone and Cell Integrated 3D Module. Cells, 11(5), 831. https://doi.org/10.3390/cells11050831