
Role of Polo-like Kinases Plk1 and Plk4 in the Initiation of Centriole Duplication—Impact on Cancer


Abstract
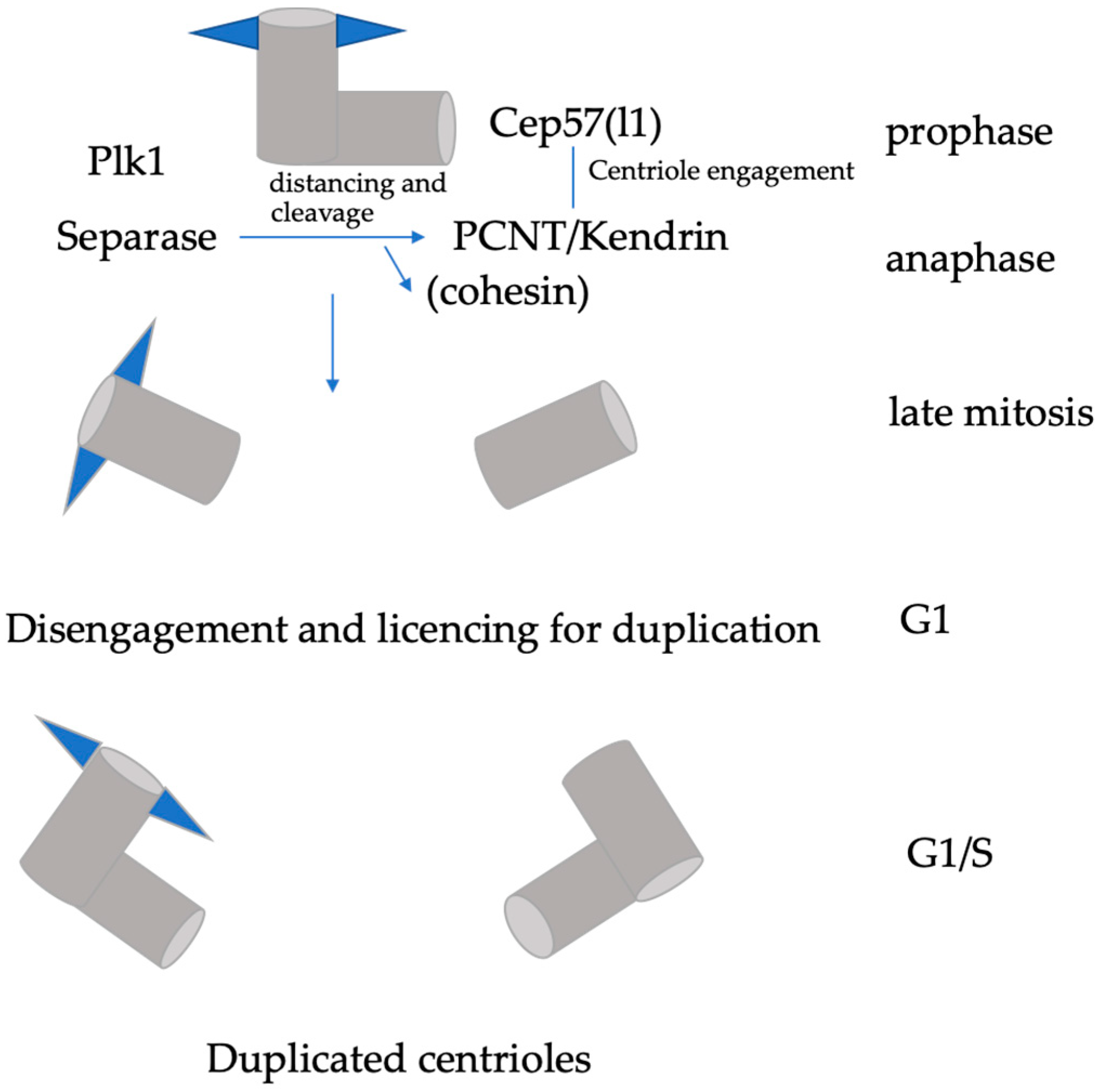
:1. Introduction
2. “Licencing” Centrioles for a New Round of Centriole Duplication
3. Recruitment of Plk4 by Cep152 and Cep192 to the Centrosome
4. Regulators of Cep192 and Cep152 Recruitment to the Centrosome
5. Formation of a New Centriole
6. Centrosome Defects and Cancer
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Bettencourt-Dias, M.; Rodrigues-Martins, A.; Carpenter, L.; Riparbelli, M.; Lehmann, L.; Gatt, M.; Carmo, N.; Balloux, F.; Callaini, G.; Glover, D. SAK/PLK4 Is Required for Centriole Duplication and Flagella Development. Curr. Biol. 2005, 15, 2199–2207. [Google Scholar] [CrossRef] [Green Version]
- Bornens, M. The Centrosome in Cells and Organisms. Science 2012, 335, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, J.B.; Gomes, B.F.; Widlund, P.O.; Mahamid, J.; Honigmann, A.; Hyman, A.A. The Centrosome Is a Selective Condensate that Nucleates Microtubules by Concentrating Tubulin. Cell 2017, 169, 1066–1077.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollman, J.M.; Merdes, A.; Mourey, L.; Agard, D.A. Microtubule nucleation by γ-tubulin complexes. Nat. Rev. Mol. Cell Biol. 2011, 12, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Fry, A.M.; Sampson, J.; Shak, C.; Shackleton, S. Recent advances in pericentriolar material organization: Ordered layers and scaffolding gels. F1000Research 2017, 6, 1622. [Google Scholar] [CrossRef]
- Nigg, E.A.; Holland, A.J. Once and only once: Mechanisms of centriole duplication and their deregulation in disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 297–312. [Google Scholar] [CrossRef]
- O’Connell, K.F.; Caron, C.; Kopish, K.R.; Hurd, D.D.; Kemphues, K.J.; Li, Y.; White, J.G. The C. elegans zyg-1 Gene Encodes a Regulator of Centrosome Duplication with Distinct Maternal and Paternal Roles in the Embryo. Cell 2001, 105, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Arquint, C.; Sonnen, K.; Stierhof, Y.-D.; Nigg, E. Cell-cycle-regulated expression of STIL controls centriole number in human cells. J. Cell Sci. 2012, 125, 1342–1352. [Google Scholar] [CrossRef] [Green Version]
- Delattre, M.; Leidel, S.; Wani, K.; Baumer, K.; Bamat, J.; Schnabel, H.; Feichtinger, R.; Schnabel, R.; Gönczy, P. Centriolar SAS-5 is required for centrosome duplication in C. elegans. Nat. Cell Biol. 2004, 6, 656–664. [Google Scholar] [CrossRef]
- Wong, C.; Stearns, T. Centrosome number is controlled by a centrosome-intrinsic block to reduplication. Nat. Cell Biol. 2003, 5, 539–544. [Google Scholar] [CrossRef]
- Kuriyama, R.; Borisy, G.G. Centriole cycle in Chinese hamster ovary cells as determined by whole-mount electron microscopy. J. Cell Biol. 1981, 91, 814–821. [Google Scholar] [CrossRef]
- Barrera, J.A.; Kao, L.-R.; Hammer, R.E.; Seemann, J.; Fuchs, J.L.; Megraw, T.L. CDK5RAP2 Regulates Centriole Engagement and Cohesion in Mice. Dev. Cell 2010, 18, 913–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsou, M.-F.B.; Wang, W.-J.; George, K.A.; Uryu, K.; Stearns, T.; Jallepalli, P.V. Polo Kinase and Separase Regulate the Mitotic Licensing of Centriole Duplication in Human Cells. Dev. Cell 2009, 17, 344–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schöckel, L.; Möckel, M.; Mayer, B.; Boos, D.; Stemmann, O. Cleavage of cohesin rings coordinates the separation of centrioles and chromatids. Nat. Cell Biol. 2011, 13, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.; Nasmyth, K. Cohesin cleavage is insufficient for centriole disengagement in Drosophila. Curr. Biol. 2013, 23, R601–R603. [Google Scholar] [CrossRef] [Green Version]
- Laan, L.; Pavin, N.; Husson, J.; Romet-Lemonne, G.; van Duijn, M.; López, M.P.; Vale, R.D.; Jülicher, F.; Reck-Peterson, S.L.; Dogterom, M. Cortical Dynein Controls Microtubule Dynamics to Generate Pulling Forces that Position Microtubule Asters. Cell 2012, 148, 502–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, K.; Ohsumi, K.; Iwabuchi, M.; Kawamata, T.; Ono, Y.; Takahashi, M. Kendrin Is a Novel Substrate for Separase Involved in the Licensing of Centriole Duplication. Curr. Biol. 2012, 22, 915–921. [Google Scholar] [CrossRef] [Green Version]
- Pagan, J.K.; Marzio, A.; Jones, M.J.; Saraf, A.; Jallepalli, P.V.; Florens, L.; Washburn, M.P.; Pagano, M. Degradation of Cep68 and PCNT cleavage mediate Cep215 removal from the PCM to allow centriole separation, disengagement and licensing. Nat. Cell Biol. 2015, 17, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Kong, D.; Sharma, M.; Magidson, V.; Loncarek, J. Plk1 relieves centriole block to reduplication by promoting daughter centriole maturation. Nat. Commun. 2015, 6, 8077. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Takao, D.; Ito, K.K.; Takahashi, M.; Kitagawa, D. The Cep57-pericentrin module organizes PCM expansion and centriole engagement. Nat. Commun. 2019, 10, 931. [Google Scholar] [CrossRef] [Green Version]
- Ito, K.K.; Watanabe, K.; Ishida, H.; Matsuhashi, K.; Chinen, T.; Hata, S.; Kitagawa, D. Cep57 and Cep57L1 maintain centriole engagement in interphase to ensure centriole duplication cycle. J. Cell Biol. 2021, 220, e202005153. [Google Scholar] [CrossRef] [PubMed]
- Lončarek, J.; Hergert, P.; Khodjakov, A. Centriole Reduplication during Prolonged Interphase Requires Procentriole Maturation Governed by Plk1. Curr. Biol. 2010, 20, 1277–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karki, M.; Keyhaninejad, N.; Shuster, C.B. Precocious centriole disengagement and centrosome fragmentation induced by mitotic delay. Nat. Commun. 2017, 8, 15803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cizmecioglu, O.; Arnold, M.; Bahtz, R.; Settele, F.; Ehret, L.; Haselmann-Weiss, U.; Antony, C.; Hoffmann, I. Cep152 acts as a scaffold for recruitment of Plk4 and CPAP to the centrosome. J. Cell Biol. 2010, 191, 731–739. [Google Scholar] [CrossRef] [Green Version]
- Sonnen, K.; Gabryjonczyk, A.-M.; Anselm, E.; Stierhof, Y.-D.; Nigg, E.A. Human Cep192 and Cep152 cooperate in Plk4 recruitment and centriole duplication. J. Cell Sci. 2013, 126, 3223–3233. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-S.; Park, J.-E.; Shukla, A.; Choi, S.; Murugan, R.N.; Lee, J.H.; Ahn, M.; Rhee, K.; Bang, J.K.; Kim, B.Y.; et al. Hierarchical recruitment of Plk4 and regulation of centriole biogenesis by two centrosomal scaffolds, Cep192 and Cep152. Proc. Natl. Acad. Sci. USA 2013, 110, E4849–E4857. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-Y.; Park, J.-E.; Kim, T.-S.; Kim, J.H.; Kwak, M.-J.; Ku, B.; Tian, L.; Murugan, R.N.; Ahn, M.; Komiya, S.; et al. Molecular basis for unidirectional scaffold switching of human Plk4 in centriole biogenesis. Nat. Struct. Mol. Biol. 2014, 21, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Zitouni, S.; Nabais, C.; Jana, S.; Guerrero, A.; Bettencourt-Dias, M. Polo-like kinases: Structural variations lead to multiple functions. Nat. Rev. Mol. Cell Biol. 2014, 15, 433–452. [Google Scholar] [CrossRef]
- Gomez-Ferreria, M.A.; Rath, U.; Buster, D.W.; Chanda, S.K.; Caldwell, J.S.; Rines, D.; Sharp, D.J. Human Cep192 Is Required for Mitotic Centrosome and Spindle Assembly. Curr. Biol. 2007, 17, 1960–1966. [Google Scholar] [CrossRef] [Green Version]
- Lawo, S.; Hasegan, M.; Gupta, G.D.; Pelletier, L. Subdiffraction imaging of centrosomes reveals higher-order organizational features of pericentriolar material. Nat. Cell Biol. 2012, 14, 1148–1158. [Google Scholar] [CrossRef]
- Dzhindzhev, N.S.; Yu, Q.D.; Weiskopf, K.; Tzolovsky, G.; Cunha-Ferreira, I.; Riparbelli, M.; Rodrigues-Martins, A.; Bettencourt-Dias, M.; Callaini, G.; Glover, D.M. Asterless is a scaffold for the onset of centriole assembly. Nature 2010, 467, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.A.; Kopish, K.R.; Zipperlen, P.; Ahringer, J.; O’Connell, K.F. Centrosome Maturation and Duplication in C. elegans Require the Coiled-Coil Protein SPD-2. Dev. Cell 2004, 6, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, L.; Ozlu, N.; Hannak, E.; Cowan, C.; Habermann, B.; Ruer, M.; Müller-Reichert, T.; Hyman, A. The Caenorhabditis elegans Centrosomal Protein SPD-2 Is Required for both Pericentriolar Material Recruitment and Centriole Duplication. Curr. Biol. 2004, 14, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habedanck, R.; Stierhof, Y.-D.; Wilkinson, C.; Nigg, E. The Polo kinase Plk4 functions in centriole duplication. Nat. Cell Biol. 2005, 7, 1140–1146. [Google Scholar] [CrossRef] [PubMed]
- Hatch, E.; Kulukian, A.; Holland, A.J.; Cleveland, D.; Stearns, T. Cep152 interacts with Plk4 and is required for centriole duplication. J. Cell Biol. 2010, 191, 721–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Lawo, S.; Bird, A.; Pinchev, D.; Ralph, A.; Richter, C.; Müller-Reichert, T.; Kittler, R.; Hyman, A.A.; Pelletier, L. The Mammalian SPD-2 Ortholog Cep192 Regulates Centrosome Biogenesis. Curr. Biol. 2008, 18, 136–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vulprecht, J.; David, A.; Tibelius, A.; Castiel, A.; Konotop, G.; Liu, F.; Bestvater, F.; Raab, M.S.; Zentgraf, H.; Izraeli, S.; et al. STIL is required for centriole duplication in human cells. J. Cell Sci. 2012, 125, 1353–1362. [Google Scholar] [CrossRef] [Green Version]
- Stevens, N.R.; Dobbelaere, J.; Brunk, K.; Franz, A.; Raff, J.W. Drosophila Ana2 is a conserved centriole duplication factor. J. Cell Biol. 2010, 188, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Conduit, P.T.; Brunk, K.; Dobbelaere, J.; Dix, C.I.; Lucas, E.P.; Raff, J.W. Centrioles Regulate Centrosome Size by Controlling the Rate of Cnn Incorporation into the PCM. Curr. Biol. 2010, 20, 2178–2186. [Google Scholar] [CrossRef] [Green Version]
- Hamill, D.R.; Severson, A.F.; Carter, J.C.; Bowerman, B. Centrosome maturation and mitotic spindle assembly in C. elegans require SPD-5, a protein with multiple coiled-coil domains. Dev. Cell 2002, 3, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Strnad, P.; Leidel, S.; Vinogradova, T.; Euteneuer, U.; Khodjakov, A.; Gönczy, P. Regulated HsSAS-6 Levels Ensure Formation of a Single Procentriole per Centriole during the Centrosome Duplication Cycle. Dev. Cell 2007, 13, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues-Martins, A.; Bettencourt-Dias, M.; Riparbelli, M.; Ferreira, C.; Ferreira, I.; Callaini, G.; Glover, D. DSAS-6 Organizes a Tube-like Centriole Precursor, and Its Absence Suggests Modularity in Centriole Assembly. Curr. Biol. 2007, 17, 1465–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leidel, S.; Delattre, M.; Cerutti, L.; Baumer, K.; Gönczy, P. SAS-6 defines a protein family required for centrosome duplication in C. elegans and in human cells. Nat. Cell Biol. 2005, 7, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Leidel, S.; Gönczy, P. SAS-4 Is Essential for Centrosome Duplication in C. elegans and Is Recruited to Daughter Centrioles Once per Cell Cycle. Dev. Cell 2003, 4, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.I.; Kleylein-Sohn, J.; Westendorf, J.; Le Clech, M.; Lavoie, S.B.; Stierhof, Y.-D.; Nigg, E.A. Control of Centriole Length by CPAP and CP110. Curr. Biol. 2009, 19, 1005–1011. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, J.; Mennella, V.; Blachon, S.; Zhai, B.; Smith, A.H.; Megraw, T.L.; Nicastro, D.; Gygi, S.P.; Agard, D.A.; Avidor-Reiss, T. Sas-4 provides a scaffold for cytoplasmic complexes and tethers them in a centrosome. Nat. Commun. 2011, 2, 359. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, Y.; Yoshiba, S.; Gupta, A.; Watanabe, K.; Kitagawa, D. Cep295 is a conserved scaffold protein required for generation of a bona fide mother centriole. Nat. Commun. 2016, 7, 12567. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-W.; Hsu, W.-B.; Tsai, J.-J.; Tang, C.-J.C.; Tang, T.K. CEP295 interacts with microtubules and is required for centriole elongation. J. Cell Sci. 2016, 129, 2501–2513. [Google Scholar] [CrossRef] [Green Version]
- Saurya, S.; Roque, H.; Novak, Z.A.; Wainman, A.; Aydogan, M.G.; Volanakis, A.; Sieber, B.; Pinto, D.M.S.; Raff, J.W. Drosophila Ana1 is required for centrosome assembly and centriole elongation. J. Cell Sci. 2016, 129, 2514–2525. [Google Scholar] [CrossRef] [Green Version]
- Chinen, T.; Yamazaki, K.; Hashimoto, K.; Fujii, K.; Watanabe, K.; Takeda, Y.; Yamamoto, S.; Nozaki, Y.; Tsuchiya, Y.; Takao, D.; et al. Centriole and PCM cooperatively recruit CEP192 to spindle poles to promote bipolar spindle assembly. J. Cell Biol. 2021, 220, e202006085. [Google Scholar] [CrossRef]
- Brown, N.J.; Marjanović, M.; Luders, J.; Stracker, T.H.; Costanzo, V. Cep63 and Cep152 Cooperate to Ensure Centriole Duplication. PLoS ONE 2013, 8, e69986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sir, J.-H.; Barr, A.; Nicholas, A.K.; Carvalho, O.; Khurshid, M.; Sossick, A.; Reichelt, S.; D’Santos, C.; Woods, C.G.; Gergely, F. A primary microcephaly protein complex forms a ring around parental centrioles. Nat. Genet. 2011, 43, 1147–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Zhu, L.; Zhu, Y.; Cao, J.; Li, S.; Huang, Q.; Xu, T.; Huang, X.; Yan, X.; Zhu, X. The Cep63 paralogue Deup1 enables massive de novo centriole biogenesis for vertebrate multiciliogenesis. Nat. Cell Biol. 2013, 15, 1434–1444. [Google Scholar] [CrossRef]
- Zhao, H.; Yang, S.; Chen, Q.; Duan, X.; Li, G.; Huang, Q.; Zhu, X.; Yan, X. Cep57 and Cep57l1 function redundantly to recruit the Cep63-Cep152 complex for centriole biogenesis. J. Cell Sci. 2020, 133, jcs241836. [Google Scholar] [CrossRef] [PubMed]
- Bossard, C.; Laurell, H.; Berghe, L.V.D.; Meunier, S.; Zanibellato, C.; Prats, H. Translokin is an intracellular mediator of FGF-2 trafficking. Nat. Cell Biol. 2003, 5, 433–439. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, T.; Zheng, T.; Teng, J.; Chen, J. Cep57 is a Mis12-interacting kinetochore protein involved in kinetochore targeting of Mad1–Mad2. Nat. Commun. 2016, 7, 10151. [Google Scholar] [CrossRef] [Green Version]
- Snape, K.; Hanks, S.; Ruark, E.; Barros-Núñez, P.; Elliott, A.; Murray, A.; Lane, A.H.; Shannon, N.; Callier, P.; Chitayat, D.; et al. Mutations in CEP57 cause mosaic variegated aneuploidy syndrome. Nat. Genet. 2011, 43, 527–529. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; He, R.; Zhou, H.; Yu, A.C.; Zhang, B.; Teng, J.; Chen, J. Cep57, a NEDD1-binding pericentriolar material component, is essential for spindle pole integrity. Cell Res. 2012, 22, 1390–1401. [Google Scholar] [CrossRef] [Green Version]
- Cuevas, R.; Korzeniewski, N.; Tolstov, Y.; Hohenfellner, M.; Duensing, S. FGF-2 Disrupts Mitotic Stability in Prostate Cancer through the Intracellular Trafficking Protein CEP57. Cancer Res. 2012, 73, 1400–1410. [Google Scholar] [CrossRef] [Green Version]
- Lukinavicius, G.; Lavogina, D.; Orpinell, M.; Umezawa, K.; Reymond, L.; Garin, N.; Gönczy, P.; Johnsson, K. Selective Chemical Crosslinking Reveals a Cep57-Cep63-Cep152 Centrosomal Complex. Curr. Biol. 2013, 23, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Lipinszki, Z.; Rangone, H.; Min, M.; Mykura, C.; Chao-Chu, J.; Schneider, S.; Dzhindzhev, N.; Gottardo, M.; Riparbelli, M.G.; et al. Conserved molecular interactions in centriole-to-centrosome conversion. Nat. Cell Biol. 2015, 18, 87–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, Z.A.; Conduit, P.T.; Wainman, A.; Raff, J.W. Asterless Licenses Daughter Centrioles to Duplicate for the First Time in Drosophila Embryos. Curr. Biol. 2014, 24, 1276–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodani, A.; Yu, T.W.; Johnson, J.R.; Jayaraman, D.; Johnson, T.L.; Al-Gazali, L.; Sztriha, L.; Partlow, J.N.; Kim, H.; Krup, A.L.; et al. Centriolar satellites assemble centrosomal microcephaly proteins to recruit CDK2 and promote centriole duplication. eLife 2015, 4, e07519. [Google Scholar] [CrossRef] [Green Version]
- Slevin, L.K.; Nye, J.; Pinkerton, D.C.; Buster, D.W.; Rogers, G.C.; Slep, K.C. The Structure of the Plk4 Cryptic Polo Box Reveals Two Tandem Polo Boxes Required for Centriole Duplication. Structure 2012, 20, 1905–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha-Ferreira, I.; Bento, I.; Pimenta-Marques, A.; Jana, S.; Lince-Faria, M.; Duarte, P.; Borrego-Pinto, J.; Gilberto, S.; Amado, T.; Brito, D.; et al. Regulation of Autophosphorylation Controls PLK4 Self-Destruction and Centriole Number. Curr. Biol. 2013, 23, 2245–2254. [Google Scholar] [CrossRef] [Green Version]
- Holland, A.J.; Lan, W.; Niessen, S.; Hoover, H.; Cleveland, D.W. Polo-like kinase 4 kinase activity limits centrosome overduplication by autoregulating its own stability. J. Cell Biol. 2010, 188, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klebba, J.E.; Buster, D.W.; Nguyen, A.L.; Swatkoski, S.; Gucek, M.; Rusan, N.M.; Rogers, G.C. Polo-like Kinase 4 Autodestructs by Generating Its Slimb-Binding Phosphodegron. Curr. Biol. 2013, 23, 2255–2261. [Google Scholar] [CrossRef] [Green Version]
- Guderian, G.; Westendorf, J.; Uldschmid, A.; Nigg, E.A. Plk4 trans-autophosphorylation regulates centriole number by controlling βTrCP-mediated degradation. J. Cell Sci. 2010, 123, 2163–2169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holland, A.J.; Fachinetti, D.; Zhu, Q.; Bauer, M.; Verma, I.M.; Nigg, E.A.; Cleveland, D.W. The autoregulated instability of Polo-like kinase 4 limits centrosome duplication to once per cell cycle. Genes Dev. 2012, 26, 2684–2689. [Google Scholar] [CrossRef] [Green Version]
- Cunha-Ferreira, I.; Rodrigues-Martins, A.; Bento, I.; Riparbelli, M.; Zhang, W.; Laue, E.; Callaini, G.; Glover, D.; Bettencourt-Dias, M. The SCF/Slimb Ubiquitin Ligase Limits Centrosome Amplification through Degradation of SAK/PLK4. Curr. Biol. 2009, 19, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Rogers, G.C.; Rusan, N.M.; Roberts, D.M.; Peifer, M.; Rogers, S.L. The SCFSlimb ubiquitin ligase regulates Plk4/Sak levels to block centriole reduplication. J. Cell Biol. 2009, 184, 225–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, M.; Ashikawa, T.; Nozaki, Y.; Kozuka-Hata, H.; Goto, H.; Inagaki, M.; Oyama, M.; Kitagawa, D. Direct interaction of Plk4 with STIL ensures formation of a single procentriole per parental centriole. Nat. Commun. 2014, 5, 5267. [Google Scholar] [CrossRef] [Green Version]
- Arquint, C.; Gabryjonczyk, A.-M.; Imseng, S.; Böhm, R.; Sauer, E.; Hiller, S.; Nigg, E.; Maier, T. STIL binding to Polo-box 3 of PLK4 regulates centriole duplication. eLife 2015, 4, e07888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyer, T.C.; Clutario, K.M.; Lambrus, B.G.; Daggubati, V.; Holland, A.J. Binding of STIL to Plk4 activates kinase activity to promote centriole assembly. J. Cell Biol. 2015, 209, 863–878. [Google Scholar] [CrossRef] [Green Version]
- Lopes, C.A.M.; Jana, S.; Cunha-Ferreira, I.; Zitouni, S.; Bento, I.; Duarte, P.; Gilberto, S.; Freixo, F.; Guerrero, A.; Francia, M.; et al. PLK4 trans-Autoactivation Controls Centriole Biogenesis in Space. Dev. Cell 2015, 35, 222–235. [Google Scholar] [CrossRef] [Green Version]
- Swallow, C.J.; Ko, M.A.; Siddiqui, N.U.; Hudson, J.W.; Dennis, J.W. Sak/Plk4 and mitotic fidelity. Oncogene 2005, 24, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Saito, H.; Takekawa, M. SAPK pathways and p53 cooperatively regulate PLK4 activity and centrosome integrity under stress. Nat. Commun. 2013, 4, 1775. [Google Scholar] [CrossRef] [Green Version]
- Klebba, J.E.; Buster, D.W.; McLamarrah, T.A.; Rusan, N.M.; Rogers, G.C. Autoinhibition and relief mechanism for Polo-like kinase 4. Proc. Natl. Acad. Sci. USA 2015, 112, E657–E666. [Google Scholar] [CrossRef] [Green Version]
- Dzhindzhev, N.S.; Tzolovsky, G.; Lipinszki, Z.; Schneider, S.; Lattao, R.; Fu, J.; Dębski, J.; Dadlez, M.; Glover, D.M. Plk4 Phosphorylates Ana2 to Trigger Sas6 Recruitment and Procentriole Formation. Curr. Biol. 2014, 24, 2526–2532. [Google Scholar] [CrossRef] [Green Version]
- Kratz, A.-S.; Bärenz, F.; Richter, K.; Hoffmann, I. Plk4-dependent phosphorylation of STIL is required for centriole duplication. Biol. Open 2015, 4, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, D.; Vakonakis, I.; Olieric, N.; Hilbert, M.; Keller, D.; Olieric, V.; Bortfeld, M.; Erat, M.C.; Flückiger, I.; Gönczy, P.; et al. Structural Basis of the 9-Fold Symmetry of Centrioles. Cell 2011, 144, 364–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Breugel, M.; Hirono, M.; Andreeva, A.; Yanagisawa, H.-A.; Yamaguchi, S.; Nakazawa, Y.; Morgner, N.; Petrovich, M.; Ebong, I.-O.; Robinson, C.V.; et al. Structures of SAS-6 Suggest Its Organization in Centrioles. Science 2011, 331, 1196–1199. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.-J.C.; Lin, S.-Y.; Hsu, W.-B.; Lin, Y.-N.; Wu, C.-T.; Lin, Y.-C.; Chang, C.-W.; Wu, K.-S.; Tang, T.K. The human microcephaly protein STIL interacts with CPAP and is required for procentriole formation. EMBO J. 2011, 30, 4790–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatzopoulos, G.N.; Erat, M.C.; Cutts, E.; Rogala, K.B.; Slater, L.M.; Stansfeld, P.J.; Vakonakis, I. Structural Analysis of the G-Box Domain of the Microcephaly Protein CPAP Suggests a Role in Centriole Architecture. Structure 2013, 21, 2069–2077. [Google Scholar] [CrossRef] [Green Version]
- Cottee, M.A.; Muschalik, N.; Wong, Y.L.; Johnson, C.M.; Johnson, S.; Andreeva, A.; Oegema, K.; Lea, S.M.; Raff, J.W.; Van Breugel, M. Crystal structures of the CPAP/STIL complex reveal its role in centriole assembly and human microcephaly. eLife 2013, 2, e01071. [Google Scholar] [CrossRef] [PubMed]
- Marteil, G.; Guerrero, A.; Vieira, A.F.; de Almeida, B.P.; Machado, P.; Mendonça, S.; Mesquita, M.; Villarreal, B.; Fonseca, I.; Francia, M.E.; et al. Over-elongation of centrioles in cancer promotes centriole amplification and chromosome missegregation. Nat. Commun. 2018, 9, 1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gönczy, P. Centrosomes and cancer: Revisiting a long-standing relationship. Nat. Cancer 2015, 15, 639–652. [Google Scholar] [CrossRef]
- Godinho, S.A.; Pellman, D. Causes and consequences of centrosome abnormalities in cancer. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130467. [Google Scholar] [CrossRef] [Green Version]
- Godinho, S.A.; Picone, R.; Burute, M.; Dagher, R.; Su, Y.; Leung, C.T.; Polyak, K.; Brugge, J.S.; Théry, M.; Pellman, D. Oncogene-like induction of cellular invasion from centrosome amplification. Nature 2014, 510, 167–171. [Google Scholar] [CrossRef] [Green Version]
- Ganier, O.; Schnerch, D.; Nigg, E.A. Structural centrosome aberrations sensitize polarized epithelia to basal cell extrusion. Open Biol. 2018, 8, 180044. [Google Scholar] [CrossRef] [Green Version]
- Ganem, N.J.; Godinho, S.A.; Pellman, D. A mechanism linking extra centrosomes to chromosomal instability. Nature 2009, 460, 278–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crasta, K.; Ganem, N.; Dagher, R.; Lantermann, A.B.; Ivanova, E.V.; Pan, Y.; Nezi, L.; Protopopov, A.; Chowdhury, D.; Pellman, D. DNA breaks and chromosome pulverization from errors in mitosis. Nature 2012, 482, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-Z.; Spektor, A.; Cornils, H.; Francis, J.M.; Jackson, E.K.; Liu, S.; Meyerson, M.; Pellman, D. Chromothripsis from DNA damage in micronuclei. Nature 2015, 522, 179–184. [Google Scholar] [CrossRef] [Green Version]
- Quintyne, N.J.; Reing, J.E.; Hoffelder, D.R.; Gollin, S.M.; Saunders, W.S.; Augui, S.; Filion, G.J.; Huart, S.; Nora, E.; Guggiari, M.; et al. Spindle Multipolarity Is Prevented by Centrosomal Clustering. Science 2005, 307, 127–129. [Google Scholar] [CrossRef]
- Silkworth, W.T.; Nardi, I.K.; Scholl, L.M.; Cimini, D. Multipolar Spindle Pole Coalescence Is a Major Source of Kinetochore Mis-Attachment and Chromosome Mis-Segregation in Cancer Cells. PLoS ONE 2009, 4, e6564. [Google Scholar] [CrossRef] [Green Version]
- Basto, R.; Brunk, K.; Vinadogrova, T.; Peel, N.; Franz, A.; Khodjakov, A.; Raff, J.W. Centrosome Amplification Can Initiate Tumorigenesis in Flies. Cell 2008, 133, 1032–1042. [Google Scholar] [CrossRef] [Green Version]
- Vitre, B.; Holland, A.J.; Kulukian, A.; Shoshani, O.; Hirai, M.; Wang, Y.; Maldonado, M.; Cho, T.; Boubaker, J.; Swing, D.A.; et al. Chronic centrosome amplification without tumorigenesis. Proc. Natl. Acad. Sci. USA 2015, 112, E6321–E6330. [Google Scholar] [CrossRef] [Green Version]
- Coelho, P.A.; Bury, L.; Shahbazi, M.N.; Liakath-Ali, K.; Tate, P.H.; Wormald, S.; Hindley, C.J.; Huch, M.; Archer, J.; Skarnes, W.C.; et al. Over-expression of Plk4 induces centrosome amplification, loss of primary cilia and associated tissue hyperplasia in the mouse. Open Biol. 2015, 5, 150209. [Google Scholar] [CrossRef] [Green Version]
- Kulukian, A.; Holland, A.J.; Vitre, B.; Naik, S.; Cleveland, D.W.; Fuchs, E. Epidermal development, growth control, and homeostasis in the face of centrosome amplification. Proc. Natl. Acad. Sci. USA 2015, 112, E6311–E6320. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.S.; Bakker, B.; Boeckx, B.; Moyett, J.; Lu, J.; Vitre, B.; Spierings, D.; Lansdorp, P.; Cleveland, D.; Lambrechts, D.; et al. Centrosome Amplification Is Sufficient to Promote Spontaneous Tumorigenesis in Mammals. Dev. Cell 2017, 40, 313–322.e5. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Mammals | D. melanogaster | C. elegans | References |
|---|---|---|---|
| Plk4 | Sak | Zyg-1 | [34] |
| [1] | |||
| [7] | |||
| Cep152 | Asterless | [24] | |
| [35] | |||
| [31] | |||
| Cep192 | Spd-2 | Spd-2 | [33] |
| [32] | |||
| [36] | |||
| STIL | Ana-2 | Sas-5 | [37] |
| [8] | |||
| [38] | |||
| [9] | |||
| Cdk5Rap2 | CNN | Spd-5 | [39] |
| [40] | |||
| Sas-6 | Sas-6 | Sas-6 | [41] |
| [42] | |||
| [43] | |||
| CPAP | Sas-4 | Sas-4 | [44] |
| [45] | |||
| [46] | |||
| Cep295 | Ana-1 | [47] | |
| [48] | |||
| [49] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffmann, I. Role of Polo-like Kinases Plk1 and Plk4 in the Initiation of Centriole Duplication—Impact on Cancer. Cells 2022, 11, 786. https://doi.org/10.3390/cells11050786
Hoffmann I. Role of Polo-like Kinases Plk1 and Plk4 in the Initiation of Centriole Duplication—Impact on Cancer. Cells. 2022; 11(5):786. https://doi.org/10.3390/cells11050786
Chicago/Turabian StyleHoffmann, Ingrid. 2022. "Role of Polo-like Kinases Plk1 and Plk4 in the Initiation of Centriole Duplication—Impact on Cancer" Cells 11, no. 5: 786. https://doi.org/10.3390/cells11050786
APA StyleHoffmann, I. (2022). Role of Polo-like Kinases Plk1 and Plk4 in the Initiation of Centriole Duplication—Impact on Cancer. Cells, 11(5), 786. https://doi.org/10.3390/cells11050786