
Free Fatty Acid Receptors (FFARs) in Adipose: Physiological Role and Therapeutic Outlook

 ,
,
Abstract
1. Introduction
2. FFAR2 and FFAR4 Are Highly Expressed in Adipose Tissue
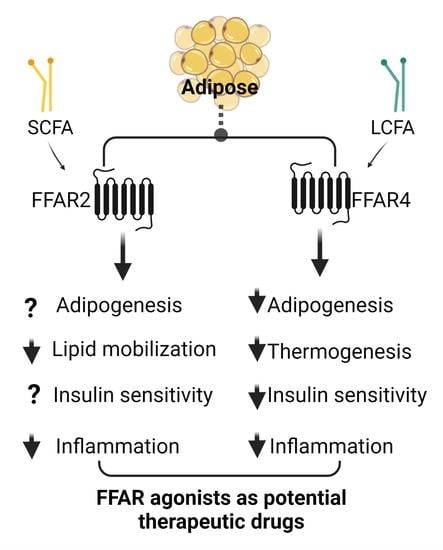
3. Role of FFAR2 in Adipose Metabolism and Energy Homeostasis
4. Role of FFAR4 (GPR120) in Adipogenesis and Adipose Metabolism
5. The Downstream Signal Mediated by FFAR2 and FFAR4 in Adipose Tissue
6. FFAR Agonists in the Treatment of Metabolic Diseases
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, L.; Zhu, L.; Meister, J.; Bone, D.B.; Pydi, S.P.; Rossi, M.; Wess, J. Use of DREADD Technology to Identify Novel Targets for Antidiabetic Drugs. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 421–440. [Google Scholar] [CrossRef]
- Thompson, M.D.; Siminovitch, K.A.; Cole, D.E.C. G Protein-Coupled Receptor Pharmacogenetics. Pharm. Drug Discov. Dev. 2008, 448, 139–185. [Google Scholar] [CrossRef]
- Krishna, S.; Insel, P. GPCRs as targets for approved drugs: How many targets and how many drugs? Mol. Pharmacol. 2018, 93, 251–258. [Google Scholar]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Fredriksson, R.; Lagerström, M.C.; Lundin, L.G.; Schiöth, H.B. The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. Mol. Pharmacol. 2003, 63, 1256–1272. [Google Scholar] [CrossRef]
- Hara, T.; Kimura, I.; Inoue, D.; Ichimura, A.; Hirasawa, A. Free Fatty Acid Receptors and Their Role in Regulation of Energy Metabolism. Rev. Physiol. Biochem. Pharmacol. 2013, 164, 77–116. [Google Scholar] [CrossRef]
- Hirasawa, A.; Hara, T.; Ichimura, A.; Tsujimoto, G. Free fatty acid receptors and their physiological role in metabolic regulation. Yakugaku Zasshi J. Pharm. Soc. Jpn. 2011, 131, 1683–1689. [Google Scholar] [CrossRef][Green Version]
- Ichimura, A.; Hirasawa, A.; Hara, T.; Tsujimoto, G. Free fatty acid receptors act as nutrient sensors to regulate energy homeostasis. Prostaglandins Other Lipid Mediat. 2009, 89, 82–88. [Google Scholar] [CrossRef]
- Lee Kennedy, R.; Vangaveti, V.; Jarrod, G.; Shashidhar, V.; Shashidhar, V.; Baune, B.T. Free fatty acid receptors: Emerging targets for treatment of diabetes and its complications. Ther. Adv. Endocrinol. Metab. 2010, 1, 165–175. [Google Scholar] [CrossRef]
- Hara, T.; Kashihara, D.; Ichimura, A.; Kimura, I.; Tsujimoto, G.; Hirasawa, A. Role of free fatty acid receptors in the regulation of energy metabolism. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2014, 1841, 1292–1300. [Google Scholar] [CrossRef]
- Itoh, Y.; Kawamata, Y.; Harada, M.; Kobayashi, M.; Fujii, R.; Fukusumi, S.; Ogi, K.; Hosoya, M.; Tanaka, Y.; Uejima, H.; et al. Free fatty acids regulate insulin secretion from pancreatic β cells through GPR40. Nature 2003, 422, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, M.M.; Meier, K.E. Free Fatty Acid Receptors and Cancer: From Nutrition to Pharmacology. In Nitric Oxide; Springer: Berlin/Heidelberg, Germany, 2016; Volume 236, pp. 233–251. [Google Scholar]
- Ghislain, J.; Poitout, V. Targeting lipid GPCRs to treat type 2 diabetes mellitus—Progress and challenges. Nat. Rev. Endocrinol. 2021, 17, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Milligan, G.; Shimpukade, B.; Ulven, T.; Hudson, B.D. Complex Pharmacology of Free Fatty Acid Receptors. Chem. Rev. 2017, 117, 67–110. [Google Scholar] [CrossRef] [PubMed]
- Son, S.-E.; Kim, N.-J.; Im, D.-S. Development of Free Fatty Acid Receptor 4 (FFA4/GPR120) Agonists in Health Science. Biomol. Ther. 2021, 29, 22–30. [Google Scholar] [CrossRef]
- Grundmann, M.; Bender, E.; Schamberger, J.; Eitner, F. Pharmacology of free fatty acid receptors and their allosteric modu-lators. Int. J. Mol. Sci. 2021, 22, 1763. [Google Scholar] [CrossRef]
- Hauser, A.S.; Chavali, S.; Masuho, I.; Jahn, L.J.; Martemyanov, K.A.; Gloriam, D.E.; Babu, M.M. Pharmacogenomics of GPCR Drug Targets. Cell 2018, 172, 41–54.e19. [Google Scholar] [CrossRef]
- Tomita, T.; Masuzaki, H.; Iwakura, H.; Fujikura, J.; Noguchi, M.; Tanaka, T.; Ebihara, K.; Kawamura, J.; Komoto, I.; Kawaguchi, Y.; et al. Expression of the gene for a membrane-bound fatty acid receptor in the pancreas and islet cell tumours in humans: Evidence for GPR40 expression in pancreatic beta cells and implications for insulin secretion. Diabetologia 2006, 49, 962–968. [Google Scholar] [CrossRef]
- Mohammad, S. Role of Free Fatty Acid Receptor 2 (FFAR2) in the Regulation of Metabolic Homeostasis. Curr. Drug Targets 2015, 16, 771–775. [Google Scholar] [CrossRef]
- Li, Z.; Xu, X.; Huang, W.; Qian, H. Free Fatty Acid Receptor 1 (FFAR1) as an Emerging Therapeutic Target for Type 2 Diabetes Mellitus: Recent Progress and Prevailing Challenges. Med. Res. Rev. 2018, 38, 381–425. [Google Scholar] [CrossRef]
- Del Guerra, S.; Bugliani, M.; D’Aleo, V.; Del Prato, S.; Boggi, U.; Mosca, F.; Filipponi, F.; Lupi, R. G-protein-coupled receptor 40 (GPR40) expression and its regulation in human pancreatic islets: The role of type 2 diabetes and fatty acids. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 22–25. [Google Scholar] [CrossRef]
- Hauge, M.; Vestmar, M.A.; Husted, A.S.; Ekberg, J.H.; Wright, M.J.; Di Salvo, J.; Weinglass, A.B.; Engelstoft, M.S.; Madsen, A.N.; Lückmann, M.; et al. GPR40 (FFAR1)—Combined Gs and Gq signaling in vitro is associated with robust incretin secretagogue action ex vivo and in vivo. Mol. Metab. 2015, 4, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Edfalk, S.; Steneberg, P.; Edlund, H. Gpr40 Is Expressed in Enteroendocrine Cells and Mediates Free Fatty Acid Stimulation of Incretin Secretion. Diabetes 2008, 57, 2280–2287. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, S. GPR40 Agonists for the Treatment of Type 2 Diabetes Mellitus: Benefits and Challenges. Curr. Drug Targets 2016, 17, 1292–1300. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Inoue, D.; Hirano, K.; Tsujimoto, G. The SCFA receptor GPR43 and energy metabolism. Front. Endocrinol. 2014, 5, 85. [Google Scholar] [CrossRef]
- Bindels, L.B.; Dewulf, E.M.; Delzenne, N.M. GPR43/FFA2: Physiopathological relevance and therapeutic prospects. Trends Pharmacol. Sci. 2013, 34, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ichimura, A.; Ohue-Kitano, R.; Igarashi, M. Free Fatty Acid Receptors in Health and Disease. Physiol. Rev. 2020, 100, 171–210. [Google Scholar] [CrossRef] [PubMed]
- Inoue, D.; Tsujimoto, G.; Kimura, I. Regulation of Energy Homeostasis by GPR41. Front. Endocrinol. 2014, 5, 81. [Google Scholar] [CrossRef]
- Mishra, S.P.; Karunakar, P.; Taraphder, S.; Yadav, H. Free Fatty Acid Receptors 2 and 3 as Microbial Metabolite Sensors to Shape Host Health: Pharmacophysiological View. Biomedicines 2020, 8, 154. [Google Scholar] [CrossRef]
- Halder, S.; Kumar, S.; Sharma, R. The therapeutic potential of GPR120: A patent review. Expert Opin. Ther. Pat. 2013, 23, 1581–1590. [Google Scholar] [CrossRef]
- Leung, P.S.; Zhang, D. Potential roles of GPR120 and its agonists in the management of diabetes. Drug Des. Dev. Ther. 2014, 8, 1013–1027. [Google Scholar] [CrossRef]
- Karakuła-Juchnowicz, H.; Róg, J.; Juchnowicz, D.; Morylowska-Topolska, J. GPR120: Mechanism of action, role and potential for medical applications. Postępy Hig. Med. Doświadczalnej 2017, 71, 942–953. [Google Scholar] [CrossRef]
- Ge, H.; Li, X.; Weiszmann, J.; Wang, P.; Baribault, H.; Chen, J.-L.; Tian, H.; Li, Y. Activation of G Protein-Coupled Receptor 43 in Adipocytes Leads to Inhibition of Lipolysis and Suppression of Plasma Free Fatty Acids. Endocrinology 2008, 149, 4519–4526. [Google Scholar] [CrossRef]
- Hong, Y.-H.; Nishimura, Y.; Hishikawa, D.; Tsuzuki, H.; Miyahara, H.; Gotoh, C.; Choi, K.-C.; Feng, D.D.; Chen, C.; Lee, H.-G.; et al. Acetate and Propionate Short Chain Fatty Acids Stimulate Adipogenesis via GPCR43. Endocrinology 2005, 146, 5092–5099. [Google Scholar] [CrossRef] [PubMed]
- Zaibi, M.S.; Stocker, C.J.; O’Dowd, J.; Davies, A.; Bellahcene, M.; Cawthorne, M.A.; Brown, A.J.; Smith, D.M.; Arch, J.R. Roles of GPR41 and GPR43 in leptin secretory responses of murine adipocytes to short chain fatty acids. FEBS Lett. 2010, 584, 2381–2386. [Google Scholar] [CrossRef]
- Dewulf, E.M.; Ge, Q.; Bindels, L.B.; Sohet, F.M.; Cani, P.D.; Brichard, S.M.; Delzenne, N.M. Evaluation of the relationship between GPR43 and adiposity in human. Nutr. Metab. 2013, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Cai, Z.; Raven, M.; Otway, D.T.; Mushtaq, R.; Johnston, J.D. Effect of short chain fatty acids on the expression of free fatty acid receptor 2 (Ffar2), Ffar3 and early-stage adipogenesis. Nutr. Diabetes 2014, 4, e128. [Google Scholar] [CrossRef] [PubMed]
- Iván, J.; Major, E.; Sipos, A.; Kovács, K.; Horváth, D.; Tamás, I.; Bay, P.; Dombrádi, V.; Lontay, B. The Short-Chain Fatty Acid Propionate Inhibits Adipogenic Differentiation of Human Chorion-Derived Mesenchymal Stem Cells Through the Free Fatty Acid Receptor 2. Stem Cells Dev. 2017, 26, 1724–1733. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Schwandner, R.; Swaminath, G.; Weiszmann, J.; Cardozo, M.; Greenberg, J.; Jaeckel, P.; Ge, H.; Wang, Y.; Jiao, X.; et al. Identification and Functional Characterization of Allosteric Agonists for the G Protein-Coupled Receptor FFA2. Mol. Pharmacol. 2008, 74, 1599–1609. [Google Scholar] [CrossRef]
- Wang, Y.; Jiao, X.; Kayser, F.; Liu, J.; Wang, Z.; Wanska, M.; Greenberg, J.; Weiszmann, J.; Ge, H.; Tian, H.; et al. The first synthetic agonists of FFA2: Discovery and SAR of phenylacetamides as allosteric modulators. Bioorgan. Med. Chem. Lett. 2010, 20, 493–498. [Google Scholar] [CrossRef]
- Ang, Z.; Ding, J.L. GPR41 and GPR43 in Obesity and Inflammation—Protective or Causative? Front. Immunol. 2016, 7, 28. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-Chain Fatty Acids Stimulate Glucagon-Like Peptide-1 Secretion via the G-Protein-Coupled Receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef] [PubMed]
- Bjursell, M.; Admyre, T.; Göransson, M.; Marley, A.E.; Smith, D.M.; Oscarsson, J.; Bohlooly, Y.M. Improved glucose control and reduced body fat mass in free fatty acid receptor 2-deficient mice fed a high-fat diet. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E211–E220. [Google Scholar] [CrossRef] [PubMed]
- Ichimura, A.; Hirasawa, A.; Poulain-Godefroy, O.; Bonnefond, A.; Hara, T.; Yengo, L.; Kimura, I.; Leloire, A.; Liu, N.; Iida, K.; et al. Dysfunction of lipid sensor GPR120 leads to obesity in both mouse and human. Nature 2012, 483, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Bonnefond, A.; Lamri, A.; Leloire, A.; Vaillant, E.; Roussel, R.; Lévy-Marchal, C.; Weill, J.; Galan, P.; Hercberg, S.; Ragot, S.; et al. Contribution of the low-frequency, loss-of-function p.R270H mutation inFFAR4(GPR120) to increased fasting plasma glucose levels. J. Med. Genet. 2015, 52, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Lamri, A.; Bonnefond, A.; Meyre, D.; Balkau, B.; Roussel, R.; Marre, M.; Froguel, P.; Fumeron, F. Interaction between GPR120 p.R270H loss-of-function variant and dietary fat intake on incident type 2 diabetes risk in the D.E.S.I.R. study. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 931–936. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Wang, L.; Meng, Q.; Kuang, H.; Liu, X. G-protein coupled receptor 120 is involved in glucose metabolism in fat cells. Cell. Mol. Biol. 2012, 58, 1757–1762. [Google Scholar]
- Song, T.; Yang, Y.; Zhou, Y.; Wei, H.; Peng, J. GPR120: A critical role in adipogenesis, inflammation, and energy metabolism in adipose tissue. Cell. Mol. Life Sci. 2017, 74, 2723–2733. [Google Scholar] [CrossRef]
- Oh, D.Y.; Talukdar, S.; Bae, E.J.; Imamura, T.; Morinaga, H.; Fan, W.Q.; Li, P.; Lu, W.J.; Watkins, S.M.; Olefsky, J.M. GPR120 Is an Omega-3 Fatty Acid Receptor Mediating Potent Anti-inflammatory and Insulin-Sensitizing Effects. Cell 2010, 142, 687–698. [Google Scholar] [CrossRef]
- Wang, Y.-M.; Liu, H.-X.; Fang, N.-Y. 9-PAHSA promotes browning of white fat via activating G-protein-coupled receptor 120 and inhibiting lipopolysaccharide/NF-kappa B pathway. Biochem. Biophys. Res. Commun. 2018, 506, 153–160. [Google Scholar] [CrossRef]
- Quesada-López, T.; Gavaldà-Navarro, A.; Morón-Ros, S.; Campderros, L.; Iglesias, R.; Giralt, M.; Villarroya, F. GPR120 controls neonatal brown adipose tissue thermogenic induction. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E742–E750. [Google Scholar] [CrossRef] [PubMed]
- Schilperoort, M.; Van Dam, A.D.; Hoeke, G.; Shabalina, I.G.; Okolo, A.; Hanyaloglu, A.C.; Dib, L.H.; Mol, I.M.; Caengprasath, N.; Chan, Y.; et al. The GPR 120 agonist TUG -891 promotes metabolic health by stimulating mitochondrial respiration in brown fat. EMBO Mol. Med. 2018, 10. [Google Scholar] [CrossRef]
- Christian, M. Elucidation of the roles of brown and brite fat genes: GPR120 is a modulator of brown adipose tissue function. Exp. Physiol. 2020, 105, 1201–1205. [Google Scholar] [CrossRef] [PubMed]
- López, T.P.Q.; Cereijo, R.; Turatsinze, J.-V.; Planavila, A.A.; Cairó, M.M.; Gavaldà-Navarro, A.A.; Peyrou, M.; Moure, R.; Iglesias, R.R.; Giralt, M.M.; et al. The lipid sensor GPR120 promotes brown fat activation and FGF21 release from adipocytes. Nat. Commun. 2016, 7, 13479. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, L.A.; Smith, N.J.; Milligan, G. International Union of Pharmacology. LXXI. Free Fatty Acid Receptors FFA1, -2, and -3: Pharmacology and Pathophysiological Functions. Pharmacol. Rev. 2008, 60, 405–417. [Google Scholar] [CrossRef]
- Hirasawa, A.; Hara, T.; Katsuma, S.; Adachi, T.; Tsujimoto, G. Free Fatty Acid Receptors and Drug Discovery. Biol. Pharm. Bull. 2008, 31, 1847–1851. [Google Scholar] [CrossRef]
- Carullo, G.; Mazzotta, S.; Vega-Holm, M.; Iglesias-Guerra, F.; Vega-Pérez, J.M.; Aiello, F.; Brizzi, A. GPR120/FFAR4 Pharmacology: Focus on Agonists in Type 2 Diabetes Mellitus Drug Discovery. J. Med. Chem. 2021, 64, 4312–4332. [Google Scholar] [CrossRef]
- Burant, C.F.; Viswanathan, P.; Marcinak, J.; Cao, C.; Vakilynejad, M.; Xie, B.; Leifke, E. TAK-875 versus placebo or glimepiride in type 2 diabetes mellitus: A phase 2, randomised, double-blind, placebo-controlled trial. Lancet 2012, 379, 1403–1411. [Google Scholar] [CrossRef]
- Arora, A.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Chigurupati, S.; Kaur, R.; Bhatia, S.; Al-Harrasi, A.; Vargas-De-La-Cruz, C.; et al. Free fatty acid receptor 1: A ray of hope in the therapy of type 2 diabetes mellitus. Inflammopharmacology 2021, 29, 1625–1639. [Google Scholar] [CrossRef] [PubMed]
- Tsujihata, Y.; Ito, R.; Suzuki, M.; Harada, A.; Negoro, N.; Yasuma, T.; Momose, Y.; Takeuchi, K. TAK-875, an Orally Available G Protein-Coupled Receptor 40/Free Fatty Acid Receptor 1 Agonist, Enhances Glucose-Dependent Insulin Secretion and Improves Both Postprandial and Fasting Hyperglycemia in Type 2 Diabetic Rats. J. Pharmacol. Exp. Ther. 2011, 339, 228–237. [Google Scholar] [CrossRef]
- Houze, J.B.; Zhu, L.; Sun, Y.; Akerman, M.; Qiu, W.; Zhang, A.J.; Sharma, R.; Schmitt, M.; Wang, Y.; Liu, J.; et al. AMG 837: A potent, orally bioavailable GPR40 agonist. Bioorgan. Med. Chem. Lett. 2012, 22, 1267–1270. [Google Scholar] [CrossRef] [PubMed]
- Ou, H.-Y.; Wu, H.-T.; Hung, H.-C.; Yang, Y.-C.; Wu, J.-S.; Chang, C.-J. Multiple mechanisms of GW-9508, a selective G protein-coupled receptor 40 agonist, in the regulation of glucose homeostasis and insulin sensitivity. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E668–E676. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.J.; Ward, R.J.; Stoddart, L.A.; Hudson, B.D.; Kostenis, E.; Ulven, T.; Morris, J.C.; Tränkle, C.; Tikhonova, I.G.; Adams, D.R.; et al. Extracellular Loop 2 of the Free Fatty Acid Receptor 2 Mediates Allosterism of a Phenylacetamide Ago-Allosteric Modulator. Mol. Pharmacol. 2011, 80, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Hudson, B.; Due-Hansen, M.E.; Christiansen, E.; Hansen, A.M.; Mackenzie, A.E.; Murdoch, H.; Pandey, S.K.; Ward, R.J.; Marquez, R.; Tikhonova, I.G.; et al. Defining the Molecular Basis for the First Potent and Selective Orthosteric Agonists of the FFA2 Free Fatty Acid Receptor. J. Biol. Chem. 2013, 288, 17296–17312. [Google Scholar] [CrossRef]
- Hansen, A.H.; Sergeev, E.; Bolognini, D.; Sprenger, R.R.; Ekberg, J.H.; Ejsing, C.S.; McKenzie, C.J.; Ulven, E.R.; Milligan, G.; Ulven, T. Discovery of a Potent Thiazolidine Free Fatty Acid Receptor 2 Agonist with Favorable Pharmacokinetic Properties. J. Med. Chem. 2018, 61, 9534–9550. [Google Scholar] [CrossRef]
- Hudson, B.D.; Shimpukade, B.; Mackenzie, A.E.; Butcher, A.J.; Pediani, J.D.; Christiansen, E.; Heathcote, H.; Tobin, A.B.; Ulven, T.; Milligan, G. The Pharmacology of a Potent and Selective Agonist, TUG-891, Demonstrates Both Potential Opportunity and Possible Challenges to Therapeutic Agonism of FFA4 (GPR120). Mol. Pharmacol. 2013, 101, mol.113.087783. [Google Scholar]
- Song, T.; Zhou, Y.; Peng, J.; Tao, Y.-X.; Yang, Y.; Xu, T.; Peng, J.; Ren, J.; Xiang, Q.; Wei, H. GPR120 promotes adipogenesis through intracellular calcium and extracellular signal-regulated kinase 1/2 signal pathway. Mol. Cell. Endocrinol. 2016, 434, 1–13. [Google Scholar] [CrossRef]
- Zhao, Y.-F.; Li, X.-C.; Liang, X.-Y.; Zhao, Y.-Y.; Xie, R.; Zhang, L.-J.; Zhang, X.-C.; Chen, C. GPR120 regulates pancreatic polypeptide secretion from male mouse islets via PLC-mediated calcium mobilization. Endocrinology 2020, 161, bqaa157. [Google Scholar] [CrossRef]
- Bianchini, G.; Nigro, C.; Sirico, A.; Novelli, R.; Prevenzano, I.; Miele, C.; Beguinot, F.; Aramini, A. A new synthetic dual agonist of GPR120/GPR40 induces GLP-1 secretion and improves glucose homeostasis in mice. Biomed. Pharmacother. 2021, 139, 111613. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Name | Major Expression Sites | References |
|---|---|---|
| FFAR1 | Pancreatic β-cells, enteroendocrine cells, nerves, immune cells | [7,11,12,13,14] |
| FFAR2 | Adipose, enteroendocrine cells, pancreatic b-cells, immune cells | [25,26,27,28] |
| FFAR3 | Pancreatic β-cells, enteroendocrine cells, nerves | [28,29] |
| FFAR4 | Adipose, enteroendocrine cells, liver, bone, lung, immune cells | [15,16,17] |
| Agonist Name | Target | Physiological Functions | Reference |
|---|---|---|---|
| TAK-875 | FFAR1 | Stimulates glucose-dependent Insulin secretion and Improves glycemic control in T2DM patients | [2] |
| AMG837 | FFAR1 | Increases insulin secretion and lowers blood glucose levels in mice | [3] |
| GW-9508 | FFAR1 | Enhances insulin sensitivity and regulates glucose homeostasis | [4] |
| TUG-424 | FFAR1 | Improves glucose tolerance in mice | [5] |
| AM-1638 | FFAR1 | Increases insulin secretion and lowers blood glucose levels in mice | [6] |
| AM-5262 | FFAR1 | Enhances glucose-stimulated insulin secretion (mouse and human islets) and improves glucose homeostasis in mice | [7,8] |
| LY2881835 | FFAR1 | Stimulates insulin secretion from pancreatic β-cells | [9] |
| MK-2305 | FFAR1 | Increases glucose-stimulated insulin secretion, resulting in improvement of glucose homeostasis in the diabetic mice | [10] |
| CMTB | FFAR2 | Inhibits lipolysis in murine adipocytes | [11] |
| TUG-1375 | FFAR2 | Induces migration of human neutrophils and inhibits lipolysis in murine adipocytes | [12] |
| Compound 1 | FFAR2 | Inhibits lipolysis in murine adipocytes | [13] |
| TUG-891 | FFAR4 | Stimulates GLP-1 secretion from enteroendocrine cells, enhances glucose uptake in 3T3-L1 adipocytes | [14] |
| AZ13581837 | FFAR4 | Increases insulin secretion and reduces blood glucose levels in mice | [15] |
| CpdA | FFAR4 | Increases insulin sensitivity and improves glucose tolerance in mice | [16] |
| Metabolex-36 | FFAR4 | Insulin secretagogue with glucose-lowering properties | [17] |
| GSK137647 | FFAR4 | Improves glucose tolerance | [18] |
| TUG-1197 | FFAR4 | Enhances insulin sensitivity and reducesbody weightt | [19] |
| NCG21 | FFAR4 | Increases GLP-1 secretion | [20] |
| GW9508 | FFAR4 | Enhances insulin sensitivity and thermogenic activity of adipocytes | [21] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Mahri, S.; Malik, S.S.; Al Ibrahim, M.; Haji, E.; Dairi, G.; Mohammad, S. Free Fatty Acid Receptors (FFARs) in Adipose: Physiological Role and Therapeutic Outlook. Cells 2022, 11, 750. https://doi.org/10.3390/cells11040750
Al Mahri S, Malik SS, Al Ibrahim M, Haji E, Dairi G, Mohammad S. Free Fatty Acid Receptors (FFARs) in Adipose: Physiological Role and Therapeutic Outlook. Cells. 2022; 11(4):750. https://doi.org/10.3390/cells11040750
Chicago/Turabian StyleAl Mahri, Saeed, Shuja Shafi Malik, Maria Al Ibrahim, Esraa Haji, Ghida Dairi, and Sameer Mohammad. 2022. "Free Fatty Acid Receptors (FFARs) in Adipose: Physiological Role and Therapeutic Outlook" Cells 11, no. 4: 750. https://doi.org/10.3390/cells11040750
APA StyleAl Mahri, S., Malik, S. S., Al Ibrahim, M., Haji, E., Dairi, G., & Mohammad, S. (2022). Free Fatty Acid Receptors (FFARs) in Adipose: Physiological Role and Therapeutic Outlook. Cells, 11(4), 750. https://doi.org/10.3390/cells11040750