
FcεRI: A Master Regulator of Mast Cell Functions


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. FcεRI
2.1. Crosslinking of the IgE–FcεRI Complex with Antigen
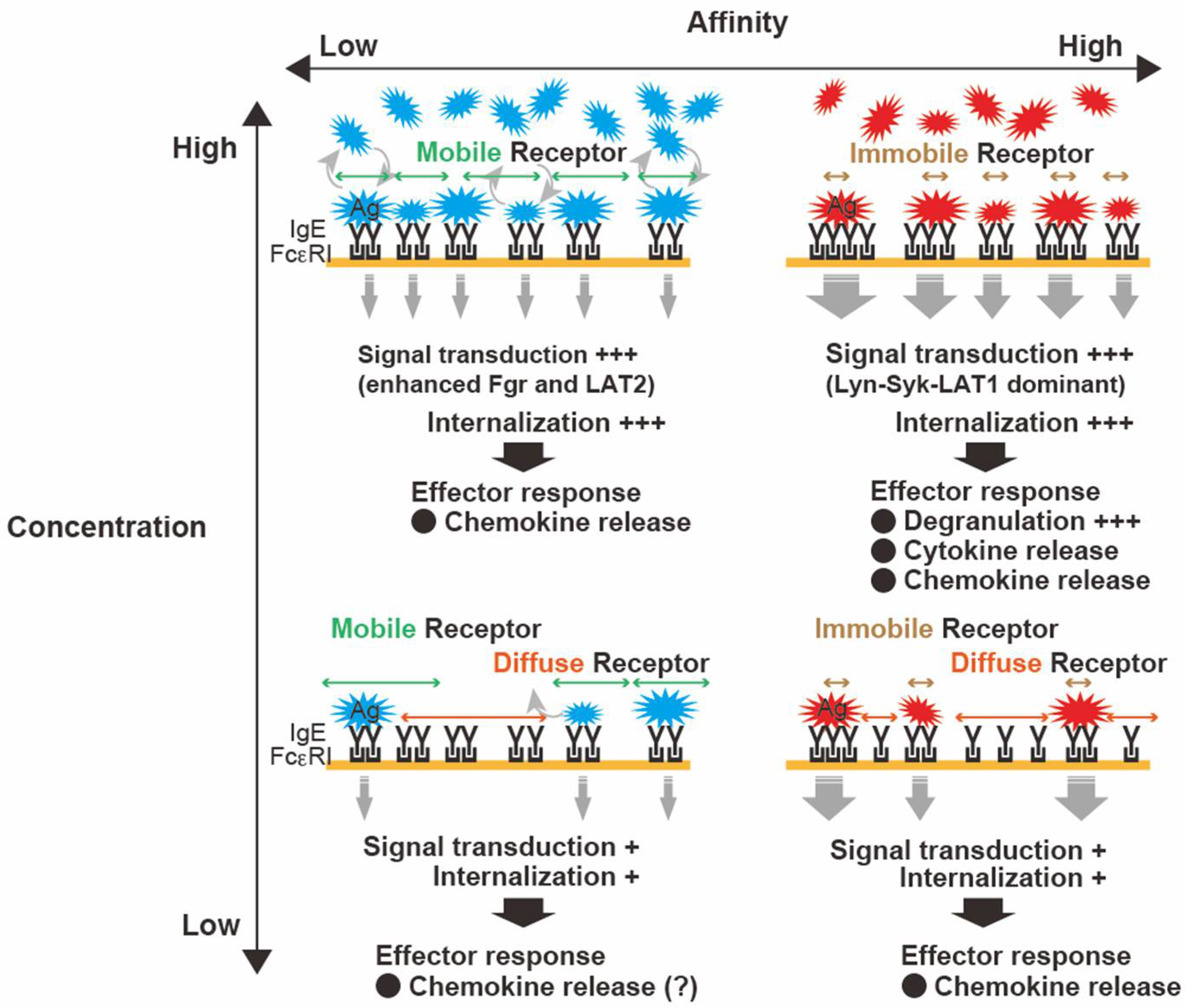
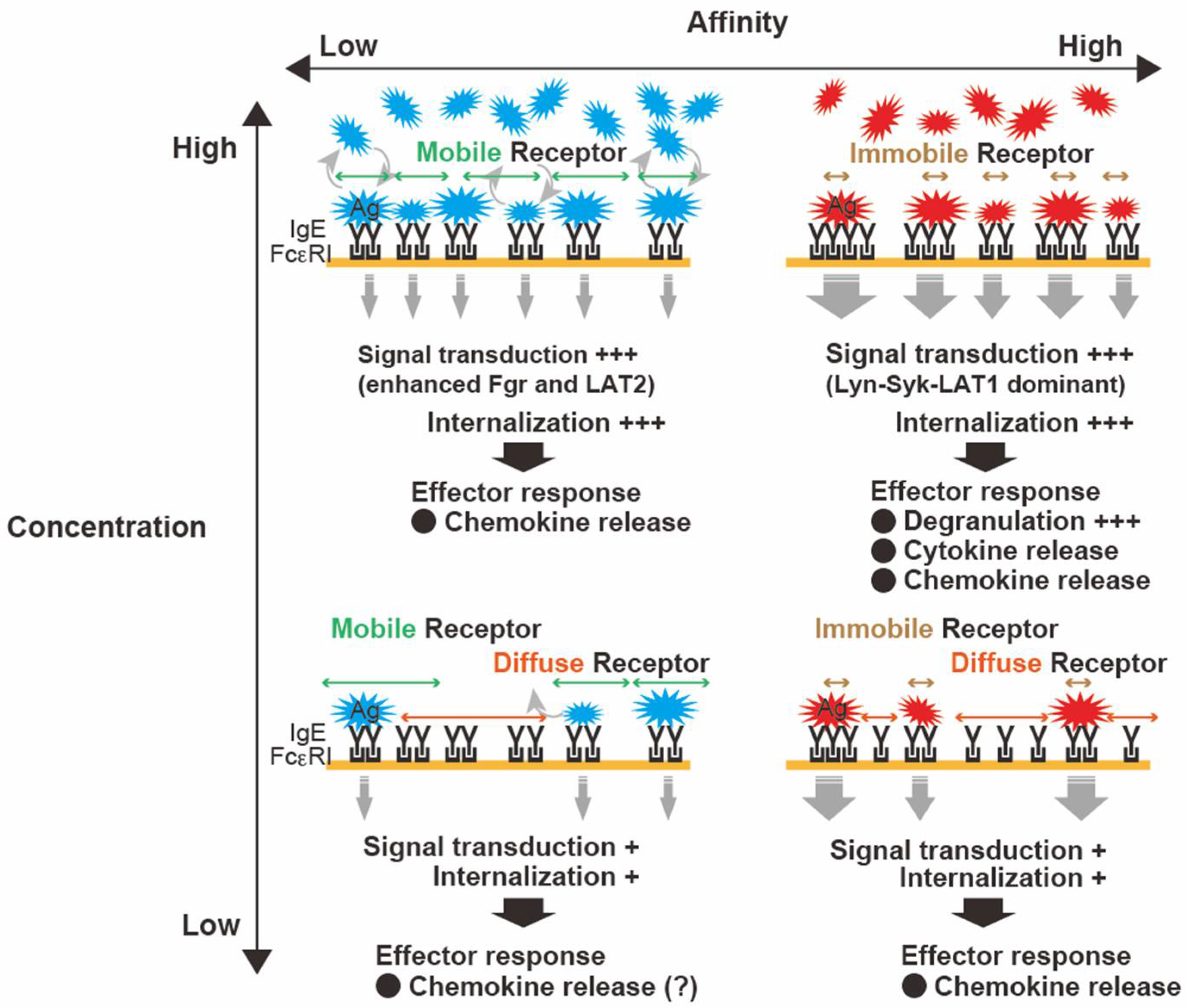
2.2. FcεRI Clustering: Size, Stability, and Mobility Dynamics
2.3. Antigen Properties: Concentration, Valency, and Affinity
2.4. Perspective
3. FcεRI: Therapeutic Potential and Benefits
3.1. Inhibition of IgE–FcεRI Binding
3.2. FcεRI–FcγRIIb Coaggregation and Inhibitory Signal against FcεRI
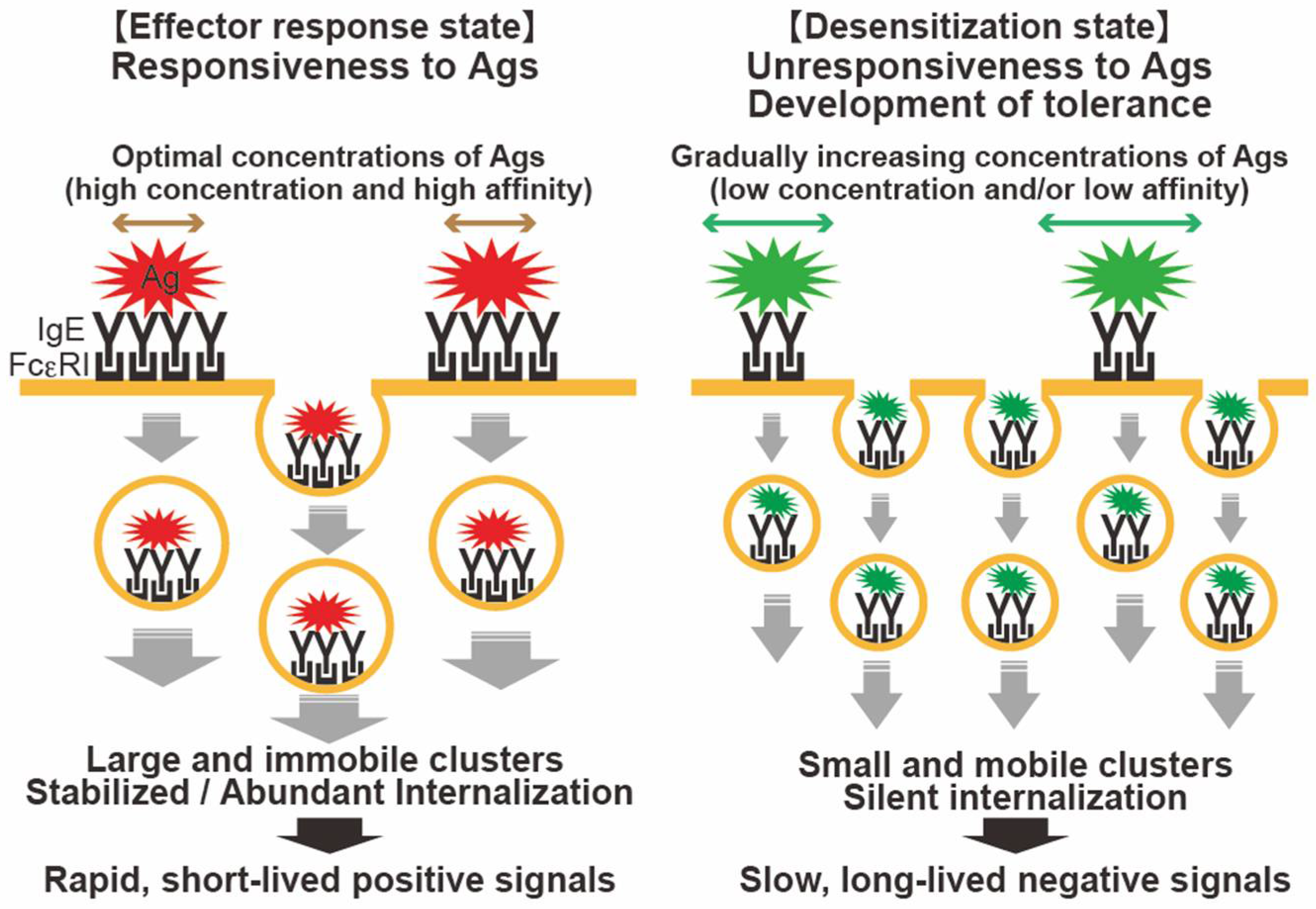
3.3. FcεRI Internalization in Allergen-Induced FcεRI Desensitization
3.4. Perspective
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pawankar, R.; Canonica, G.W.; Holgate, S.T.; Lockey, R.F.; Blaiss, M.S. (Eds.) WAO White Book on Allergy 2011-2012: Exective Summary; World Allergy Organization: Milwaukee, WI, USA, 2011. [Google Scholar]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matricardi, P.M.; Kleine-Tebbe, J.; Hoffmann, H.J.; Valenta, R.; Hilger, C.; Hofmaier, S.; Aalberse, R.C.; Agache, I.; Asero, R.; Ballmer-Weber, B.; et al. EAACI Molecular Allergology User’s Guide. Pediatric Allergy Immunol. Off. Publ. Eur. Soc. Pediatric Allergy Immunol. 2016, 27 (Suppl. S23), 1–250. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J.; Fierro, N.A.; Olivera, A.; Suzuki, R. New insights on mast cell activation via the high affinity receptor for IgE. Adv. Immunol. 2008, 98, 85–120. [Google Scholar] [CrossRef] [Green Version]
- Burton, O.T.; Oettgen, H.C. Beyond immediate hypersensitivity: Evolving roles for IgE antibodies in immune homeostasis and allergic diseases. Immunol. Rev. 2011, 242, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Kalesnikoff, J.; Galli, S.J. New developments in mast cell biology. Nat. Immunol. 2008, 9, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Kawakami, Y.; Kasakura, K.; Kawakami, T. Recent advances in mast cell activation and regulation. F1000Research 2020, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawakami, T.; Galli, S.J. Regulation of mast-cell and basophil function and survival by IgE. Nat. Rev. Immunol. 2002, 2, 773–786. [Google Scholar] [CrossRef]
- Kambayashi, T.; Koretzky, G.A. Proximal signaling events in Fc epsilon RI-mediated mast cell activation. J. Allergy Clin. Immunol. 2007, 119, 544–552; quiz 553–544. [Google Scholar] [CrossRef]
- Oettgen, H.C.; Burton, O.T. IgE receptor signaling in food allergy pathogenesis. Curr. Opin. Immunol. 2015, 36, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Dombrowicz, D.; Flamand, V.; Brigman, K.K.; Koller, B.H.; Kinet, J.-P. Abolition of anaphylaxis by targeted disruption of the high affinity immunoglobulin E receptor α chain gene. Cell 1993, 75, 969–976. [Google Scholar] [CrossRef]
- Blank, U.; Ra, C.; Miller, L.; White, K.; Metzger, H.; Kinet, J.P. Complete structure and expression in transfected cells of high affinity IgE receptor. Nature 1989, 337, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Garman, S.C.; Wurzburg, B.A.; Tarchevskaya, S.S.; Kinet, J.-P.; Jardetzky, T.S. Structure of the Fc fragment of human IgE bound to its high-affinity receptor FcεRIα. Nature 2000, 406, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Gould, H.J.; Sutton, B.J. IgE in allergy and asthma today. Nat. Rev. Immunol. 2008, 8, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J. Molecular adapters in Fc(epsilon)RI signaling and the allergic response. Curr. Opin. Immunol. 2002, 14, 688–693. [Google Scholar] [CrossRef]
- Xiao, W.; Nishimoto, H.; Hong, H.; Kitaura, J.; Nunomura, S.; Maeda-Yamamoto, M.; Kawakami, Y.; Lowell, C.A.; Ra, C.; Kawakami, T. Positive and negative regulation of mast cell activation by Lyn via the FcepsilonRI. J. Immunol. 2005, 175, 6885–6892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibilano, R.; Frossi, B.; Pucillo, C.E. Mast cell activation: A complex interplay of positive and negative signaling pathways. Eur. J. Immunol. 2014, 44, 2558–2566. [Google Scholar] [CrossRef] [PubMed]
- Asai, K.; Kitaura, J.; Kawakami, Y.; Yamagata, N.; Tsai, M.; Carbone, D.P.; Liu, F.-T.; Galli, S.J.; Kawakami, T. Regulation of Mast Cell Survival by IgE. Immunity 2001, 14, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Menon, A.K.; Holowka, D.; Webb, W.W.; Baird, B. Cross-linking of receptor-bound IgE to aggregates larger than dimers leads to rapid immobilization. J. Cell Biol. 1986, 102, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Andrews, N.L.; Lidke, K.A.; Pfeiffer, J.R.; Burns, A.R.; Wilson, B.S.; Oliver, J.M.; Lidke, D.S. Actin restricts FcepsilonRI diffusion and facilitates antigen-induced receptor immobilization. Nat. Cell Biol. 2008, 10, 955–963. [Google Scholar] [CrossRef]
- Andrews, N.L.; Pfeiffer, J.R.; Martinez, A.M.; Haaland, D.M.; Davis, R.W.; Kawakami, T.; Oliver, J.M.; Wilson, B.S.; Lidke, D.S. Small, mobile FcepsilonRI receptor aggregates are signaling competent. Immunity 2009, 31, 469–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll-Portillo, A.; Spendier, K.; Pfeiffer, J.; Griffiths, G.; Li, H.; Lidke, K.A.; Oliver, J.M.; Lidke, D.S.; Thomas, J.L.; Wilson, B.S.; et al. Formation of a mast cell synapse: Fc epsilon RI membrane dynamics upon binding mobile or immobilized ligands on surfaces. J. Immunol. 2010, 184, 1328–1338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, B.S.; Oliver, J.M.; Lidke, D.S. Spatio-temporal signaling in mast cells. Adv. Exp. Med. Biol. 2011, 716, 91–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spendier, K.; Lidke, K.A.; Lidke, D.S.; Thomas, J.L. Single-particle tracking of immunoglobulin E receptors (FcεRI) in micron-sized clusters and receptor patches. FEBS Lett. 2012, 586, 416–421. [Google Scholar] [CrossRef] [Green Version]
- Huber, M.; Helgason, C.D.; Damen, J.E.; Liu, L.; Humphries, R.K.; Krystal, G. The src homology 2-containing inositol phosphatase (SHIP) is the gatekeeper of mast cell degranulation. Proc. Natl. Acad. Sci. USA 1998, 95, 11330–11335. [Google Scholar] [CrossRef] [Green Version]
- Mahajan, A.; Barua, D.; Cutler, P.; Lidke, D.S.; Espinoza, F.A.; Pehlke, C.; Grattan, R.; Kawakami, Y.; Tung, C.-S.; Bradbury, A.R.M.; et al. Optimal aggregation of FcεRI with a structurally defined trivalent ligand overrides negative regulation driven by phosphatases. ACS Chem. Biol. 2014, 9, 1508–1519. [Google Scholar] [CrossRef]
- Harmon, B.; Chylek, L.A.; Liu, Y.; Mitra, E.D.; Mahajan, A.; Saada, E.A.; Schudel, B.R.; Holowka, D.A.; Baird, B.A.; Wilson, B.S.; et al. Timescale Separation of Positive and Negative Signaling Creates History-Dependent Responses to IgE Receptor Stimulation. Sci. Rep. 2017, 7, 15586. [Google Scholar] [CrossRef] [Green Version]
- Bag, N.; Wagenknecht-Wiesner, A.; Lee, A.; Shi, S.M.; Holowka, D.A.; Baird, B.A. Lipid-based and protein-based interactions synergize transmembrane signaling stimulated by antigen clustering of IgE receptors. Proc. Natl. Acad. Sci. USA 2021, 118, e2026583118. [Google Scholar] [CrossRef]
- Deak, P.E.; Vrabel, M.R.; Pizzuti, V.J.; Kiziltepe, T.; Bilgicer, B. Nanoallergens: A multivalent platform for studying and evaluating potency of allergen epitopes in cellular degranulation. Exp. Biol. Med. 2016, 241, 996–1006. [Google Scholar] [CrossRef]
- MacDonald, S.M.; Lichtenstein, L.M.; Proud, D.; Plaut, M.; Naclerio, R.M.; MacGlashan, D.W.; Kagey-Sobotka, A. Studies of IgE-dependent histamine releasing factors: Heterogeneity of IgE. J. Immunol. 1987, 139, 506–512. [Google Scholar]
- Bucaite, G.; Kang-Pettinger, T.; Moreira, J.; Gould, H.J.; James, L.K.; Sutton, B.J.; McDonnell, J.M. Interplay between Affinity and Valency in Effector Cell Degranulation: A Model System with Polcalcin Allergens and Human Patient-Derived IgE Antibodies. J. Immunol. 2019, 203, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Napoli, A.; Aiello, D.; Di Donna, L.; Moschidis, P.; Sindona, G. Vegetable Proteomics: The Detection of Ole e 1 Isoallergens by Peptide Matching of MALDI MS/MS Spectra of Underivatized and Dansylated Glycopeptides. J. Proteome Res. 2008, 7, 2723–2732. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.H.; Riise, E.; Bang, L.; Zhang, C.; Lund, K. Isoallergen Variations Contribute to the Overall Complexity of Effector Cell Degranulation: Effect Mediated through Differentiated IgE Affinity. J. Immunol. 2010, 184, 4966–4972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.W.; Kim, K.S.; Jin, H.S.; Kim, C.W.; Kang, D.B.; Choi, S.Y.; Yong, T.S.; Oh, S.H.; Hong, C.S. Der p 2 isoallergens have different allergenicity, and quantification with 2-site ELISA using monoclonal antibodies is influenced by the isoallergens. Clin. Exp. Allergy 2002, 32, 1042–1047. [Google Scholar] [CrossRef]
- Gimborn, K.; Lessmann, E.; Kuppig, S.; Krystal, G.; Huber, M. SHIP Down-Regulates FcεR1-Induced Degranulation at Supraoptimal IgE or Antigen Levels. J. Immunol. 2005, 174, 507–516. [Google Scholar] [CrossRef] [Green Version]
- Fehrenbach, K.; Lessmann, E.; Zorn, C.N.; Kuhny, M.; Grochowy, G.; Krystal, G.; Leitges, M.; Huber, M. Steel Factor Enhances Supraoptimal Antigen-Induced IL-6 Production from Mast Cells via Activation of Protein Kinase C-β. J. Immunol. 2009, 182, 7897–7905. [Google Scholar] [CrossRef] [Green Version]
- Seagrave, J.; Pfeiffer, J.R.; Wofsy, C.; Oliver, J.M. Relationship of IgE receptor topography to secretion in RBL-2H3 mast cells. J. Cell. Physiol. 1991, 148, 139–151. [Google Scholar] [CrossRef]
- Seagrave, J.; Oliver, J.M. Antigen-Dependent transition of IgE to a detergent-insoluble form is associated with reduced IgE receptor-dependent secretion from RBL-2H3 mast cells. J. Cell. Physiol. 1990, 144, 128–136. [Google Scholar] [CrossRef]
- Huber, M. Activation/Inhibition of mast cells by supra-optimal antigen concentrations. Cell Commun. Signal 2013, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Furumoto, Y.; Nunomura, S.; Terada, T.; Rivera, J.; Ra, C. The FcepsilonRIbeta immunoreceptor tyrosine-based activation motif exerts inhibitory control on MAPK and IkappaB kinase phosphorylation and mast cell cytokine production. J. Biol. Chem. 2004, 279, 49177–49187. [Google Scholar] [CrossRef] [Green Version]
- Gast, M.; Preisinger, C.; Nimmerjahn, F.; Huber, M. IgG-Independent Co-aggregation of FcεRI and FcγRIIB Results in LYN- and SHIP1-Dependent Tyrosine Phosphorylation of FcγRIIB in Murine Bone Marrow-Derived Mast Cells. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, M.; Rotthus, S.; Wendel, S.; Najafi, N.; Källström, E.; Focke-Tejkl, M.; Valenta, R.; Flicker, S.; Ohlin, M. Multiple independent IgE epitopes on the highly allergenic grass pollen allergen Phl p 5. Clin. Exp. Allergy 2014, 44, 1409–1419. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Takasu, Y.; Mikura, S.; Satoh, N.; Ichikawa, A. Antigen-independent Induction of Histamine Synthesis by Immunoglobulin E in Mouse Bone Marrow–derived Mast Cells. J. Exp. Med. 2002, 196, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Oka, T.; Hori, M.; Tanaka, A.; Matsuda, H.; Karaki, H.; Ozaki, H. IgE alone-induced actin assembly modifies calcium signaling and degranulation in RBL-2H3 mast cells. Am. J. Physiol.-Cell Physiol. 2004, 286, C256–C263. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Furuta, K. Roles of IgE and Histamine in Mast Cell Maturation. Cells 2021, 10, 2170. [Google Scholar] [CrossRef] [PubMed]
- Jackola, D.R.; Pierson-Mullany, L.K.; Liebeler, C.L.; Blumenthal, M.N.; Rosenberg, A. Variable binding affinities for allergen suggest a ‘selective competition’ among immunoglobulins in atopic and non-atopic humans. Mol. Immunol. 2002, 39, 367–377. [Google Scholar] [CrossRef]
- Suzuki, R.; Leach, S.; Liu, W.; Ralston, E.; Scheffel, J.; Zhang, W.; Lowell, C.A.; Rivera, J. Molecular editing of cellular responses by the high-affinity receptor for IgE. Science 2014, 343, 1021–1025. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Scheffel, J.; Rivera, J. New insights on the signaling and function of the high-affinity receptor for IgE. Curr. Top. Microbiol. Immunol. 2015, 388, 63–90. [Google Scholar] [CrossRef]
- Mahajan, A.; Youssef, L.A.; Cleyrat, C.; Grattan, R.; Lucero, S.R.; Mattison, C.P.; Erasmus, M.F.; Jacobson, B.; Tapia, L.; Hlavacek, W.S.; et al. Allergen Valency, Dose, and FcεRI Occupancy Set Thresholds for Secretory Responses to Pen a 1 and Motivate Design of Hypoallergens. J. Immunol. 2017, 198, 1034–1046. [Google Scholar] [CrossRef] [Green Version]
- Dispenza, M.C.; Bochner, B.S.; MacGlashan, D.W., Jr. Targeting the FcεRI Pathway as a Potential Strategy to Prevent Food-Induced Anaphylaxis. Front. Immunol. 2020, 11, 3282. [Google Scholar] [CrossRef]
- Gomez, G.; Jogie-Brahim, S.; Shima, M.; Schwartz, L.B. Omalizumab reverses the phenotypic and functional effects of IgE-enhanced Fc epsilonRI on human skin mast cells. J. Immunol. 2007, 179, 1353–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, C.X.; Lv, M.; Guo, L.M.; Yu, M.; Li, Y.; Lin, Z.; Hua, X.L.; Hou, C.M.; Feng, J.N.; Shen, B.F. Inhibition of IgE Activity to Bind its High Affinity Receptor (FcεRIα) by Mouse Anti-IgE Cε3∼4 Monoclonal Antibody (QME5). Int. J. Biomed. Sci. 2009, 5, 336–344. [Google Scholar] [PubMed]
- Pennington, L.F.; Tarchevskaya, S.; Brigger, D.; Sathiyamoorthy, K.; Graham, M.T.; Nadeau, K.C.; Eggel, A.; Jardetzky, T.S. Structural basis of omalizumab therapy and omalizumab-mediated IgE exchange. Nat. Commun. 2016, 7, 11610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonacci, A.; Billeci, L.; Pioggia, G.; Navarra, M.; Gangemi, S. Omalizumab for the Treatment of Chronic Idiopathic Urticaria: Systematic Review of the Literature. Pharmacotherapy 2017, 37, 464–480. [Google Scholar] [CrossRef]
- Beck, L.A.; Marcotte, G.V.; MacGlashan, D.; Togias, A.; Saini, S. Omalizumab-induced reductions in mast cell FcεRI expression and function. J. Allergy Clin. Immunol. 2004, 114, 527–530. [Google Scholar] [CrossRef]
- MacGlashan, D.W.; Bochner, B.S.; Adelman, D.C.; Jardieu, P.M.; Togias, A.; McKenzie-White, J.; Sterbinsky, S.A.; Hamilton, R.G.; Lichtenstein, L.M. Down-regulation of Fc(epsilon)RI expression on human basophils during in vivo treatment of atopic patients with anti-IgE antibody. J. Immunol. 1997, 158, 1438–1445. [Google Scholar]
- Furuichi, K.; Rivera, J.; Isersky, C. The receptor for immunoglobulin E on rat basophilic leukemia cells: Effect of ligand binding on receptor expression. Proc. Natl. Acad. Sci. USA 1985, 82, 1522–1525. [Google Scholar] [CrossRef] [Green Version]
- Lantz, C.S.; Yamaguchi, M.; Oettgen, H.C.; Katona, I.M.; Miyajima, I.; Kinet, J.P.; Galli, S.J. IgE regulates mouse basophil Fc epsilon RI expression in vivo. J. Immunol. 1997, 158, 2517–2521. [Google Scholar]
- Tontini, C.; Bulfone-Paus, S. Novel Approaches in the Inhibition of IgE-Induced Mast Cell Reactivity in Food Allergy. Front. Immunol. 2021, 12, 613461. [Google Scholar] [CrossRef]
- Eggel, A.; Baravalle, G.; Hobi, G.; Kim, B.; Buschor, P.; Forrer, P.; Shin, J.-S.; Vogel, M.; Stadler, B.M.; Dahinden, C.A.; et al. Accelerated dissociation of IgE-FcεRI complexes by disruptive inhibitors actively desensitizes allergic effector cells. J. Allergy Clin. Immunol. 2014, 133, 1709–1719.e8. [Google Scholar] [CrossRef] [Green Version]
- Maggi, L.; Rossettini, B.; Montaini, G.; Matucci, A.; Vultaggio, A.; Mazzoni, A.; Palterer, B.; Parronchi, P.; Maggi, E.; Liotta, F.; et al. Omalizumab dampens type 2 inflammation in a group of long-term treated asthma patients and detaches IgE from FcεRI. Eur. J. Immunol. 2018, 48, 2005–2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.; Eggel, A.; Tarchevskaya, S.S.; Vogel, M.; Prinz, H.; Jardetzky, T.S. Accelerated disassembly of IgE-receptor complexes by a disruptive macromolecular inhibitor. Nature 2012, 491, 613–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, A.M.; Allan, E.G.; Keeble, A.H.; Delgado, J.; Cossins, B.P.; Mitropoulou, A.N.; Pang, M.O.Y.; Ceska, T.; Beavil, A.J.; Craggs, G.; et al. Allosteric mechanism of action of the therapeutic anti-IgE antibody omalizumab. J. Biol. Chem. 2017, 292, 9975–9987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilleminault, L.; Michelet, M.; Reber, L.L. Combining Anti-IgE Monoclonal Antibodies and Oral Immunotherapy for the Treatment of Food Allergy. Clin. Rev. Allergy Immunol. 2021. [Google Scholar] [CrossRef]
- Nadeau, K.C.; Schneider, L.C.; Hoyte, L.; Borras, I.; Umetsu, D.T. Rapid oral desensitization in combination with omalizumab therapy in patients with cow’s milk allergy. J. Allergy Clin. Immunol. 2011, 127, 1622–1624. [Google Scholar] [CrossRef] [Green Version]
- Schneider, L.C.; Rachid, R.; LeBovidge, J.; Blood, E.; Mittal, M.; Umetsu, D.T. A pilot study of omalizumab to facilitate rapid oral desensitization in high-risk peanut-allergic patients. J. Allergy Clin. Immunol. 2013, 132, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Gadir, A.; Schneider, L.; Casini, A.; Charbonnier, L.M.; Little, S.V.; Harrington, T.; Umetsu, D.T.; Rachid, R.; Chatila, T.A. Oral immunotherapy with omalizumab reverses the Th2 cell-like programme of regulatory T cells and restores their function. Clin. Exp. Allergy 2018, 48, 825–836. [Google Scholar] [CrossRef]
- Gasser, P.; Tarchevskaya, S.S.; Guntern, P.; Brigger, D.; Ruppli, R.; Zbären, N.; Kleinboelting, S.; Heusser, C.; Jardetzky, T.S.; Eggel, A. The mechanistic and functional profile of the therapeutic anti-IgE antibody ligelizumab differs from omalizumab. Nat. Commun. 2020, 11, 165. [Google Scholar] [CrossRef] [Green Version]
- Stumpp, M.T.; Binz, H.K.; Amstutz, P. DARPins: A new generation of protein therapeutics. Drug Discovery Today 2008, 13, 695–701. [Google Scholar] [CrossRef]
- Baumann, M.J.; Eggel, A.; Amstutz, P.; Stadler, B.M.; Vogel, M. DARPins against a functional IgE epitope. Immunol. Lett. 2010, 133, 78–84. [Google Scholar] [CrossRef]
- Pennington, L.F.; Gasser, P.; Brigger, D.; Guntern, P.; Eggel, A.; Jardetzky, T.S. Structure-guided design of ultrapotent disruptive IgE inhibitors to rapidly terminate acute allergic reactions. J. Allergy Clin. Immunol. 2021, 148, 1049–1060. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Sugawara, T.; Matsubara, K.; Hirata, T. Inhibitory effect of carotenoids on the degranulation of mast cells via suppression of antigen-induced aggregation of high affinity IgE receptors. J. Biol. Chem. 2009, 284, 28172–28179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokura, T.; Nakano, N.; Ito, T.; Matsuda, H.; Nagasako-Akazome, Y.; Kanda, T.; Ikeda, M.; Okumura, K.; Ogawa, H.; Nishiyama, C. Inhibitory Effect of Polyphenol-Enriched Apple Extracts on Mast Cell Degranulation in Vitro Targeting the Binding between IgE and FcεRI. Biosci. Biotechnol. Biochem. 2005, 69, 1974–1977. [Google Scholar] [CrossRef] [PubMed]
- Nakano, N.; Nishiyama, C.; Tokura, T.; Nagasako-Akazome, Y.; Ohtake, Y.; Okumura, K.; Ogawa, H. Procyanidin C1 from apple extracts inhibits Fc epsilon RI-mediated mast cell activation. Int. Arch Allergy Immunol. 2008, 147, 213–221. [Google Scholar] [CrossRef]
- López-Expósito, I.; Castillo, A.; Yang, N.; Liang, B.; Li, X.-M. Chinese herbal extracts of Rubia cordifolia and Dianthus superbus suppress IgE production and prevent peanut-induced anaphylaxis. Chin. Med. 2011, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Kageyama-Yahara, N.; Suehiro, Y.; Maeda, F.; Kageyama, S.-i.; Fukuoka, J.; Katagiri, T.; Yamamoto, T.; Kadowaki, M. Pentagalloylglucose down-regulates mast cell surface FcεRI expression in vitro and in vivo. FEBS Lett. 2010, 584, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Kageyama-Yahara, N.; Hayashi, S.; Yamamoto, T.; Kadowaki, M. Sphingosine kinase-1-dependent and -independent inhibitory effects of zanthoxyli fructus to attenuate the activation of mucosal mast cells and ameliorate food allergies in mice. Evid Based Complement Alternat Med. 2012, 2012, 862743. [Google Scholar] [CrossRef]
- Nagata, Y.; Ando, H.; Sasaki, Y.; Suzuki, R. Ephedra Herb, Mao, Inhibits Antigen-Induced Mast Cell Degranulation by Induction of the Affinity Receptor for IgE Internalization. Pharm. Res. 2021, 38, 569–581. [Google Scholar] [CrossRef]
- Kepley, C.L.; Taghavi, S.; Mackay, G.; Zhu, D.; Morel, P.A.; Zhang, K.; Ryan, J.J.; Satin, L.S.; Zhang, M.; Pandolfi, P.P.; et al. Co-aggregation of FcγRII with FcϵRI on Human Mast Cells Inhibits Antigen-induced Secretion and Involves SHIP-Grb2-Dok Complexes. J. Biol. Chem. 2004, 279, 35139–35149. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Kepley, C.L.; Morel, P.A.; Okumoto, L.M.; Fukuoka, Y.; Schwartz, L.B. Fc gamma RIIa, not Fc gamma RIIb, is constitutively and functionally expressed on skin-derived human mast cells. J. Immunol. 2006, 177, 694–701. [Google Scholar] [CrossRef]
- Burton, O.T.; Epp, A.; Fanny, M.E.; Miller, S.J.; Stranks, A.J.; Teague, J.E.; Clark, R.A.; van de Rijn, M.; Oettgen, H.C. Tissue-Specific Expression of the Low-Affinity IgG Receptor, FcγRIIb, on Human Mast Cells. Front. Immunol. 2018, 9, 1244. [Google Scholar] [CrossRef] [PubMed]
- Tam, S.W.; Demissie, S.; Thomas, D.; Daëron, M. A bispecific antibody against human IgE and human FcγRII that inhibits antigen-induced histamine release by human mast cells and basophils. Allergy 2004, 59, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Allen, L.C.; Kepley, C.L.; Saxon, A.; Zhang, K. Modifications to an Fcγ-Fcɛ fusion protein alter its effectiveness in the inhibition of FcɛRI-mediated functions. J. Allergy Clin. Immunol. 2007, 120, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Strait, R.T.; Morris, S.C.; Finkelman, F.D. IgG-blocking antibodies inhibit IgE-mediated anaphylaxis in vivo through both antigen interception and Fc gamma RIIb cross-linking. J. Clin. Investig. 2006, 116, 833–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orengo, J.M.; Radin, A.R.; Kamat, V.; Badithe, A.; Ben, L.H.; Bennett, B.L.; Zhong, S.; Birchard, D.; Limnander, A.; Rafique, A.; et al. Treating cat allergy with monoclonal IgG antibodies that bind allergen and prevent IgE engagement. Nat. Commun. 2018, 9, 1421. [Google Scholar] [CrossRef]
- Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy: Multiple suppressor factors at work in immune tolerance to allergens. J. Allergy Clin. Immunol. 2014, 133, 621–631. [Google Scholar] [CrossRef]
- Berings, M.; Karaaslan, C.; Altunbulakli, C.; Gevaert, P.; Akdis, M.; Bachert, C.; Akdis, C.A. Advances and highlights in allergen immunotherapy: On the way to sustained clinical and immunologic tolerance. J. Allergy Clin. Immunol. 2017, 140, 1250–1267. [Google Scholar] [CrossRef] [Green Version]
- Musa, F.; Al-Ahmad, M.; Arifhodzic, N.; Al-Herz, W. Compliance with allergen immunotherapy and factors affecting compliance among patients with respiratory allergies. Hum. Vaccin Immunother 2017, 13, 514–517. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Jones, S.M.; Pongracic, J.A.; Song, Y.; Yang, N.; Sicherer, S.H.; Makhija, M.M.; Robison, R.G.; Moshier, E.; Godbold, J.; et al. Safety, clinical, and immunologic efficacy of a Chinese herbal medicine (Food Allergy Herbal Formula-2) for food allergy. J. Allergy Clin. Immunol. 2015, 136, 962–970.e1. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Fujiwara, K.; Tsubota, Y.; Kageyama-Yahara, N.; Hayashi, S.; Kadowaki, M. Induction of Regulatory T Cells as a Novel Mechanism Underlying the Therapeutic Action of Kakkonto, a Traditional Japanese Herbal Medicine, in a Murine Food Allergy Model. Int. Arch Allergy Immunol. 2016, 169, 146–156. [Google Scholar] [CrossRef]
- Srivastava, K.D.; Song, Y.; Yang, N.; Liu, C.; Goldberg, I.E.; Nowak-Węgrzyn, A.; Sampson, H.A.; Li, X.M. B-FAHF-2 plus oral immunotherapy (OIT) is safer and more effective than OIT alone in a murine model of concurrent peanut/tree nut allergy. Clin. Exp. Allergy 2017, 47, 1038–1049. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Yamamoto, T.; Hayashi, M.; Hayashi, S.; Kadowaki, M. Improvement of Therapeutic Efficacy of Oral Immunotherapy in Combination with Regulatory T Cell-Inducer Kakkonto in a Murine Food Allergy Model. PLoS ONE 2017, 12, e0170577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satitsuksanoa, P.; Głobińska, A.; Jansen, K.; van de Veen, W.; Akdis, M. Modified Allergens for Immunotherapy. Curr. Allergy Asthma Rep. 2018, 18, 9. [Google Scholar] [CrossRef] [PubMed]
- Carnes, J.; Gallego, T.M.; Moya, R.; Iraola, V. Allergoids for Allergy Treatment. Recent Pat. Inflamm. Allergy Drug Discov. 2018, 12, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Klimek, L.; Schmidt-Weber, C.B.; Kramer, M.F.; Skinner, M.A.; Heath, M.D. Clinical use of adjuvants in allergen-immunotherapy. Expert Rev. Clin. Immunol. 2017, 13, 599–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhernov, Y.; Curin, M.; Khaitov, M.; Karaulov, A.; Valenta, R. Recombinant allergens for immunotherapy: State of the art. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 402–414. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Z.; Lin, H. Reducing the Allergenicity of Shrimp Tropomyosin and Allergy Desensitization Based on Glycation Modification. J. Agric. Food Chem. 2021. [Google Scholar] [CrossRef]
- Oka, T.; Rios, E.J.; Tsai, M.; Kalesnikoff, J.; Galli, S.J. Rapid desensitization induces internalization of antigen-specific IgE on mouse mast cells. J. Allergy Clin. Immunol. 2013, 132, 922–932.e16. [Google Scholar] [CrossRef] [Green Version]
- Sancho-Serra, M.d.C.; Simarro, M.; Castells, M. Rapid IgE desensitization is antigen specific and impairs early and late mast cell responses targeting FcεRI internalization. Eur. J. Immunol. 2011, 41, 1004–1013. [Google Scholar] [CrossRef]
- Ang, W.X.G.; Church, A.M.; Kulis, M.; Choi, H.W.; Burks, A.W.; Abraham, S.N. Mast cell desensitization inhibits calcium flux and aberrantly remodels actin. J. Clin. Investig. 2016, 126, 4103–4118. [Google Scholar] [CrossRef] [Green Version]
- Takasato, Y.; Kurashima, Y.; Kiuchi, M.; Hirahara, K.; Murasaki, S.; Arai, F.; Izawa, K.; Kaitani, A.; Shimada, K.; Saito, Y.; et al. Orally desensitized mast cells form a regulatory network with Treg cells for the control of food allergy. Mucosal Immunol. 2021, 14, 640–651. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Suzuki, R. FcεRI Cluster Size Determines Effective Mast Cell Desensitization without Effector Responses in vitro. Int. Arch Allergy Immunol. 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Espinosa, C.; Odom, S.; Olivera, A.; Hobson, J.P.; Martinez, M.E.C.; Oliveira-Dos-Santos, A.; Barra, L.; Spiegel, S.; Penninger, J.M.; Rivera, J. Preferential signaling and induction of allergy-promoting lymphokines upon weak stimulation of the high affinity IgE receptor on mast cells. J. Exp. Med. 2003, 197, 1453–1465. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagata, Y.; Suzuki, R. FcεRI: A Master Regulator of Mast Cell Functions. Cells 2022, 11, 622. https://doi.org/10.3390/cells11040622
Nagata Y, Suzuki R. FcεRI: A Master Regulator of Mast Cell Functions. Cells. 2022; 11(4):622. https://doi.org/10.3390/cells11040622
Chicago/Turabian StyleNagata, Yuka, and Ryo Suzuki. 2022. "FcεRI: A Master Regulator of Mast Cell Functions" Cells 11, no. 4: 622. https://doi.org/10.3390/cells11040622
APA StyleNagata, Y., & Suzuki, R. (2022). FcεRI: A Master Regulator of Mast Cell Functions. Cells, 11(4), 622. https://doi.org/10.3390/cells11040622