
Enhanced Loss of Retinoic Acid Network Genes in Xenopus laevis Achieves a Tighter Signal Regulation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Embryo Culture and Treatment
2.2. Quantitative Reverse Transcription Real-Time PCR (qPCR)
2.3. Generation of CRISPant Embryos
2.4. Statistical Analysis
3. Results and Discussion
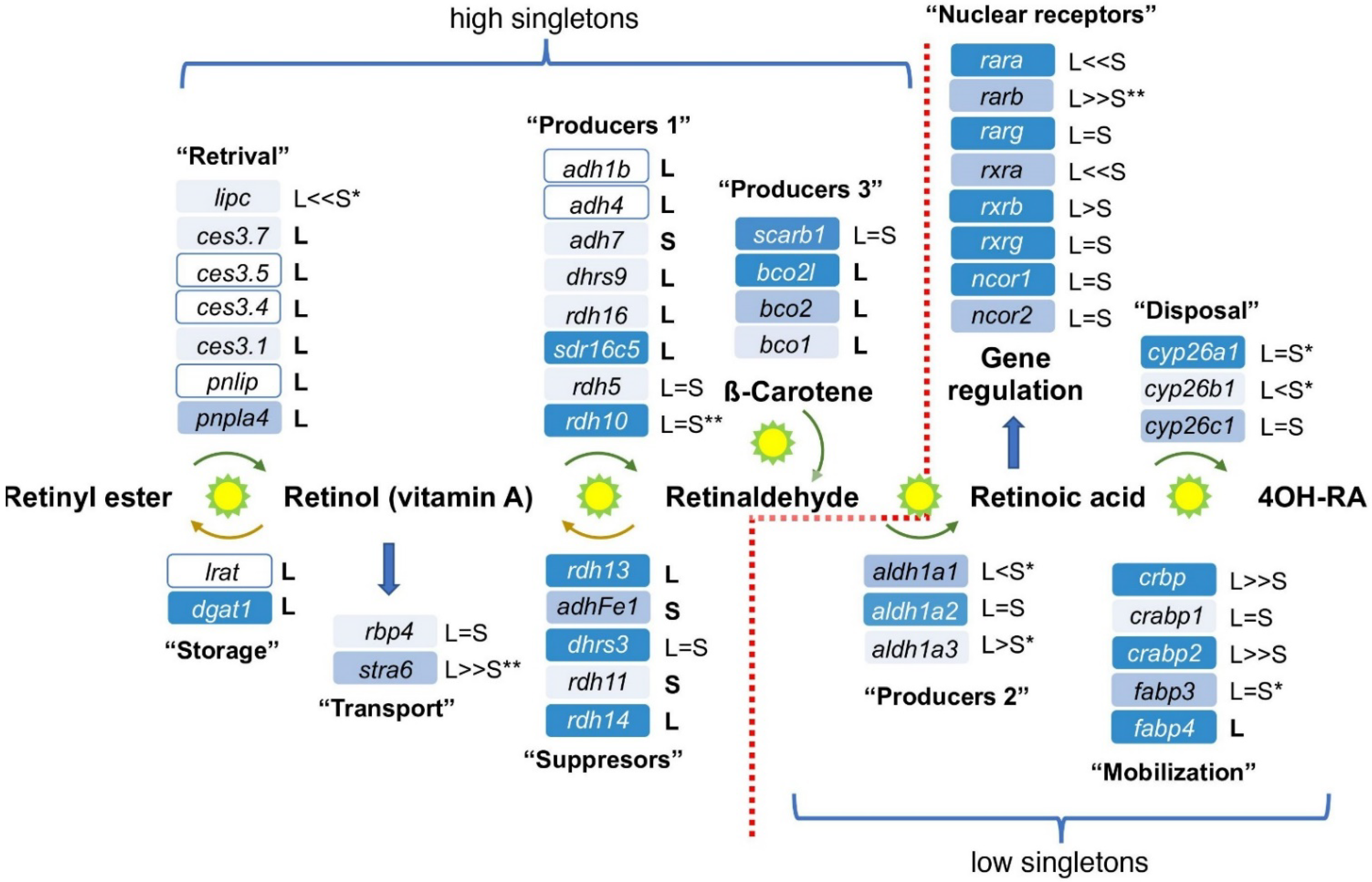
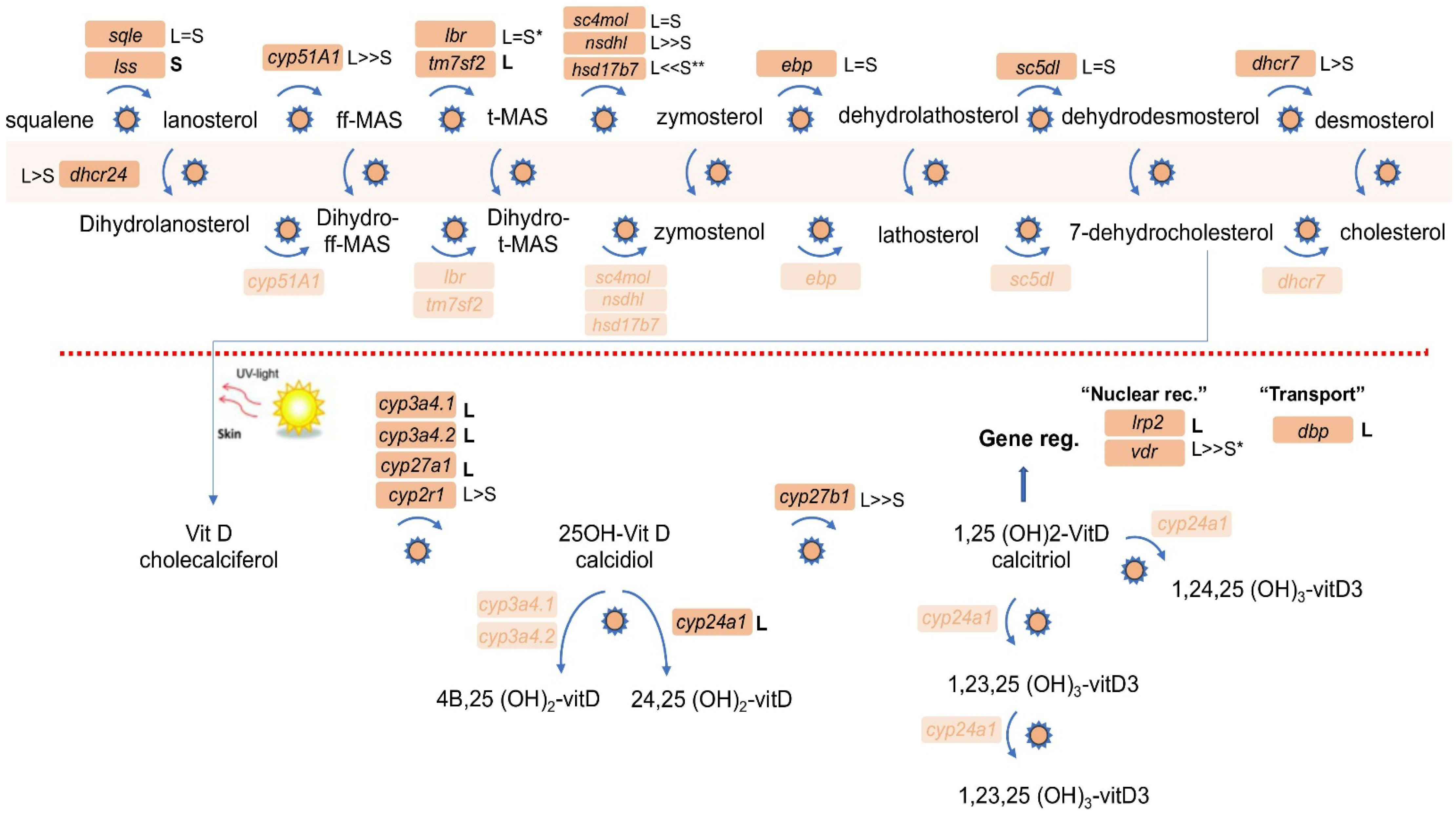
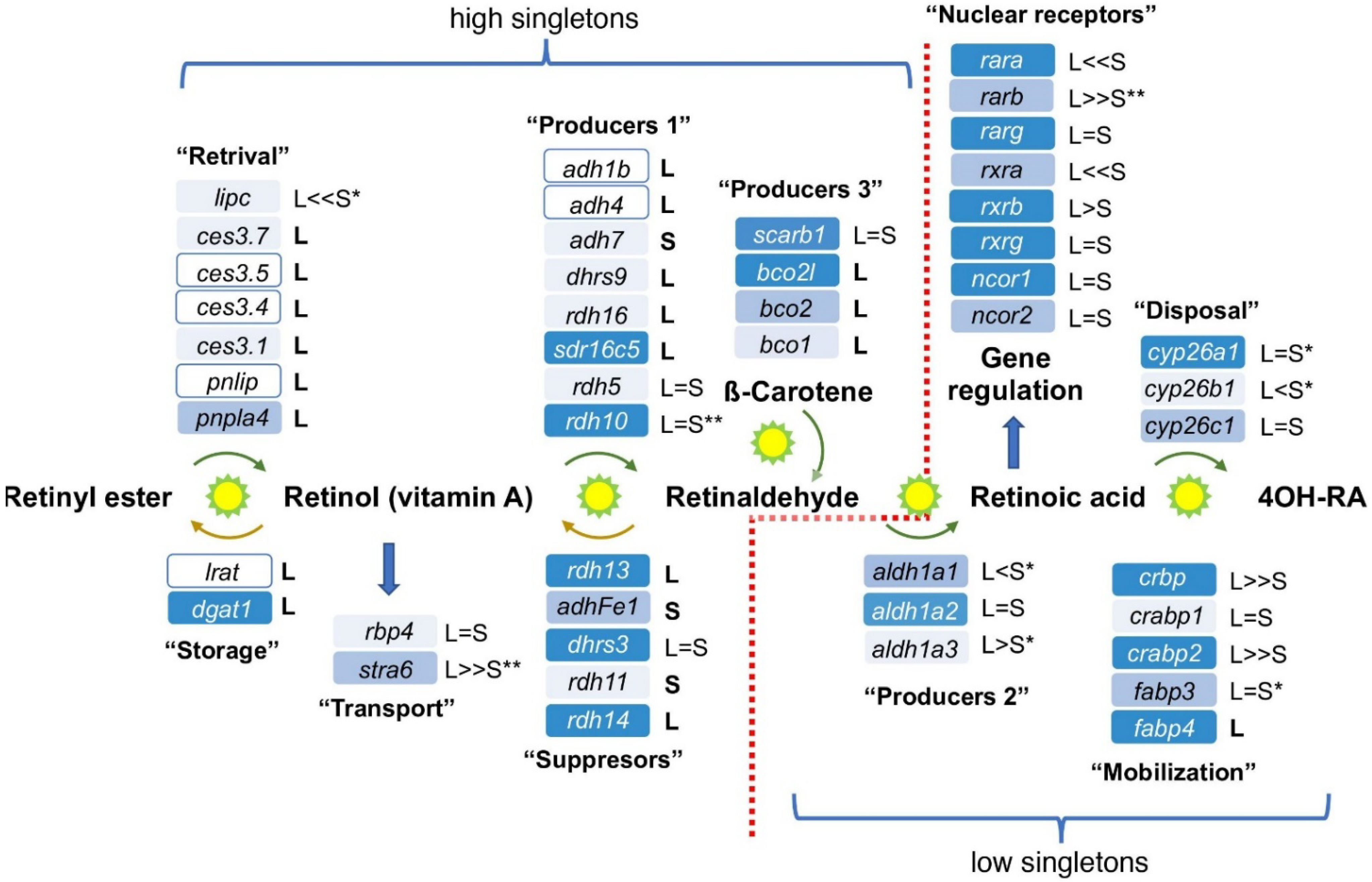
3.1. Conservation of the RA Network
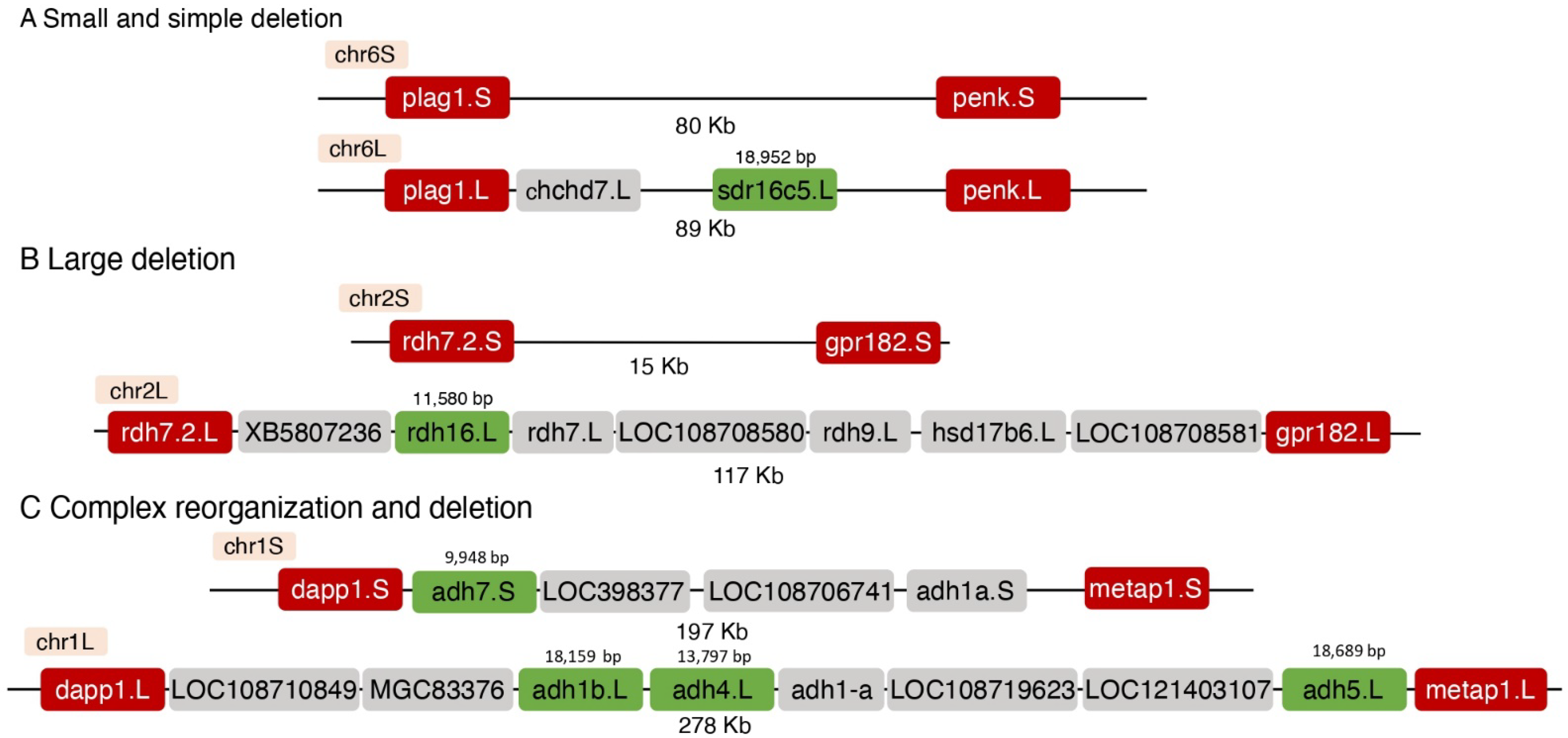
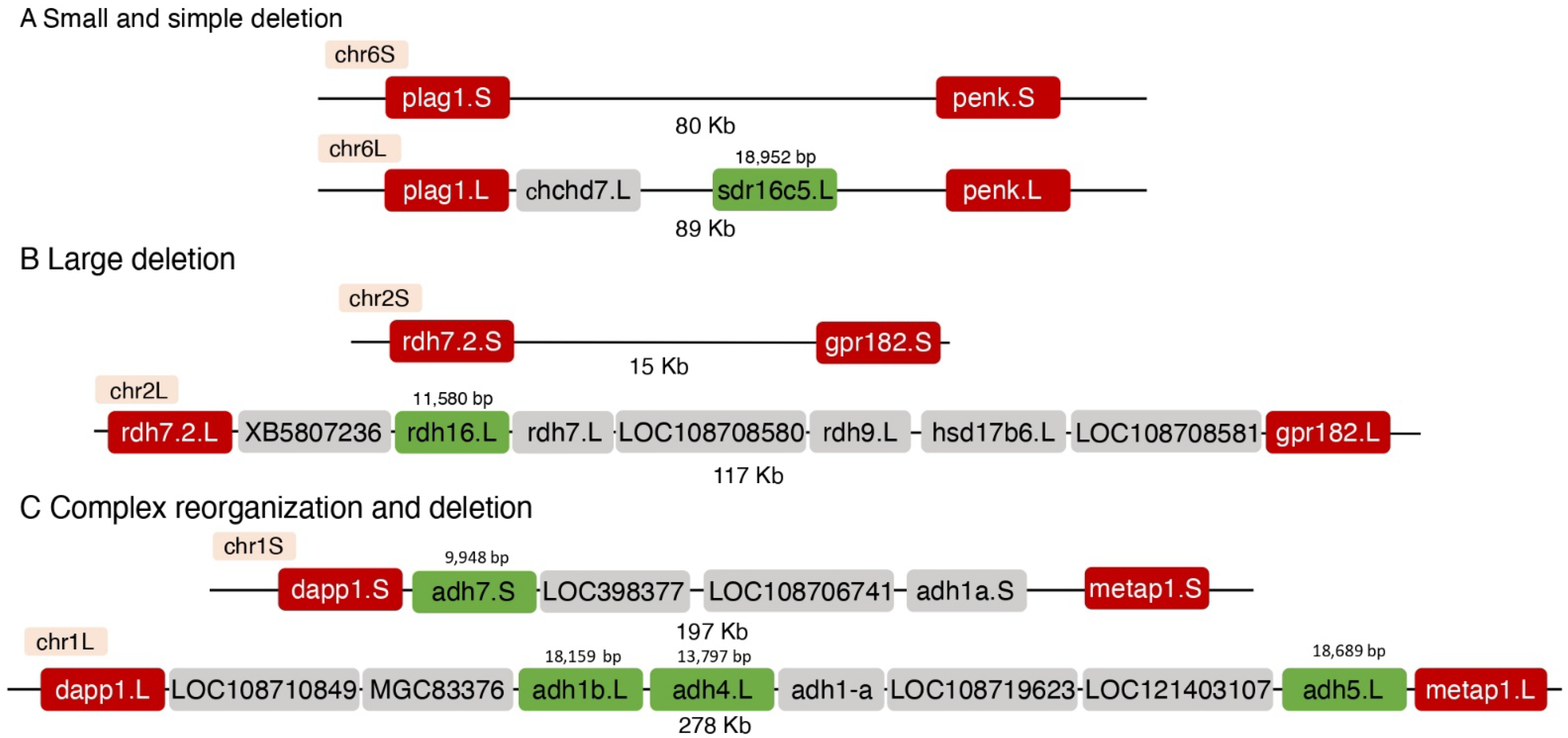
3.2. Genomic Changes in the Loss of a Homoeolog
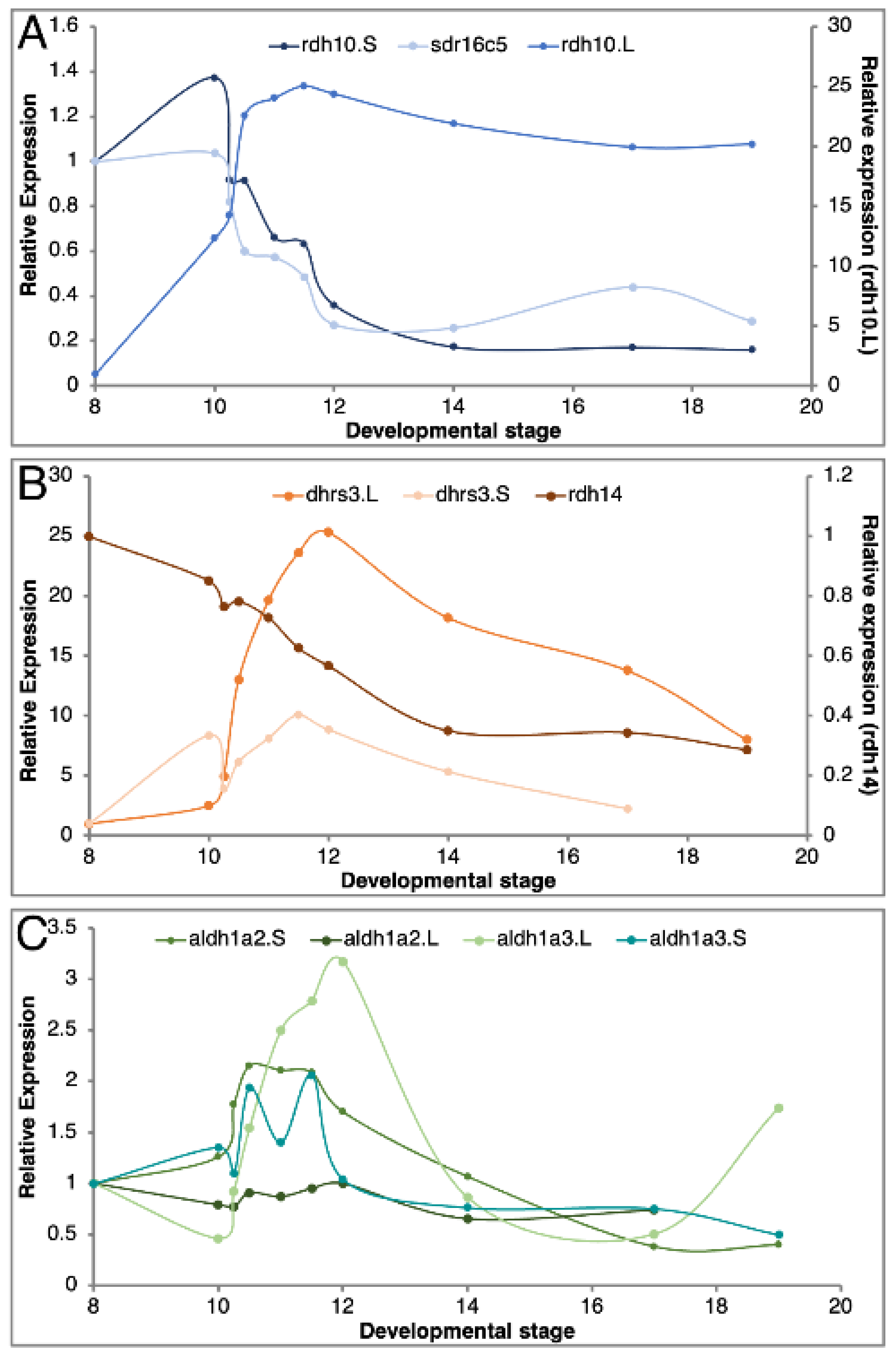
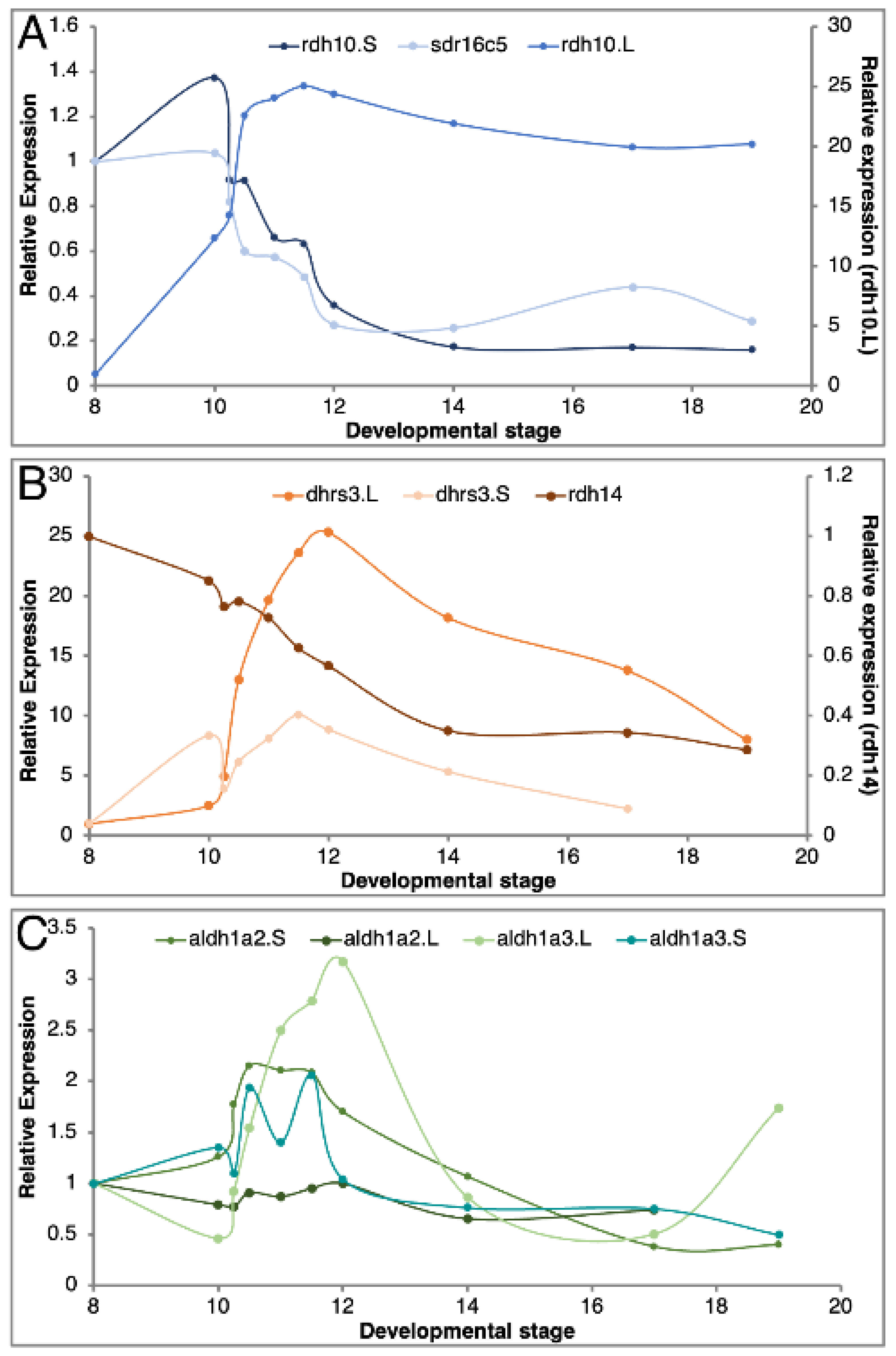
3.3. Expression Overlaps and Responsiveness of RA Network Components
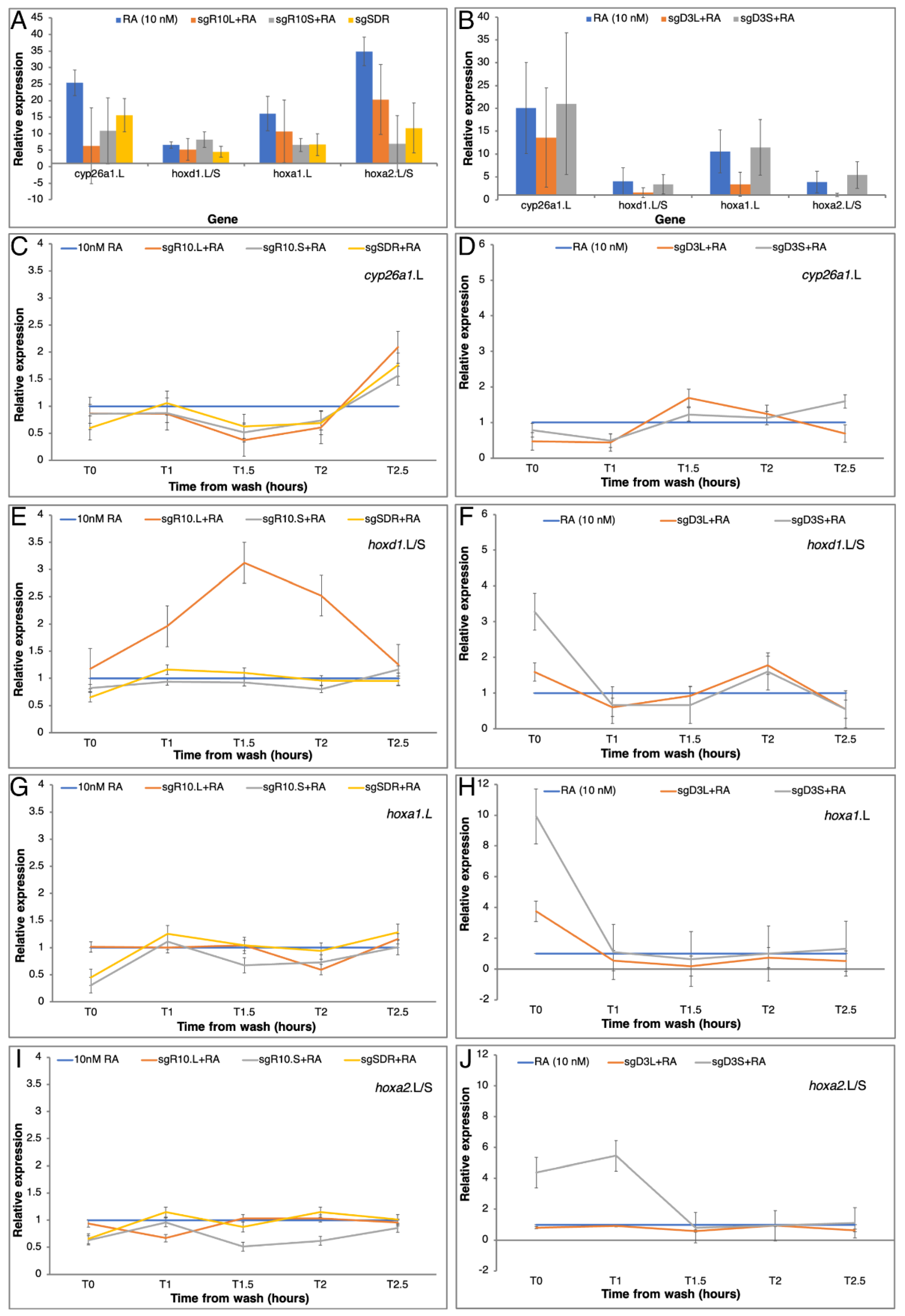
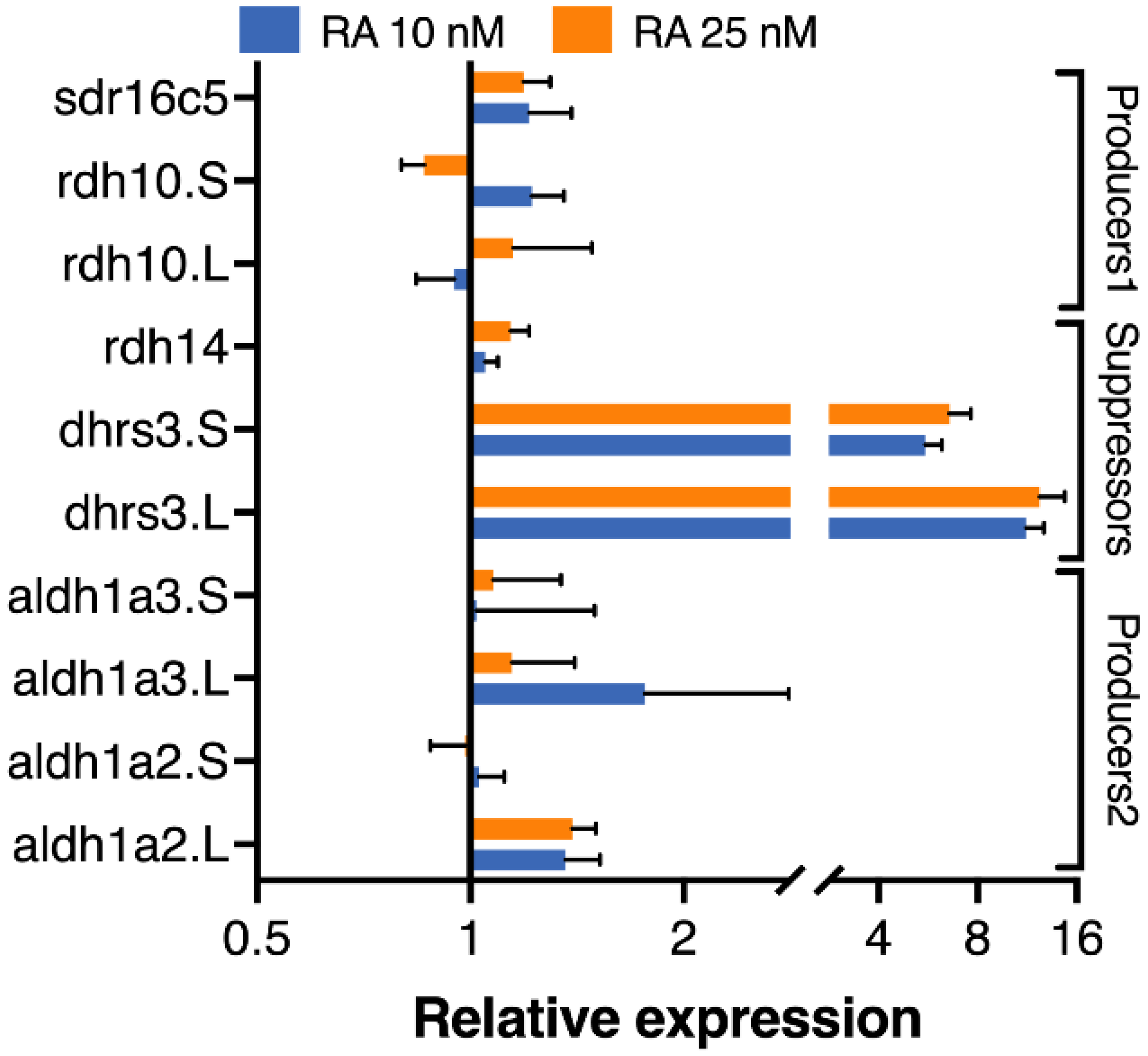
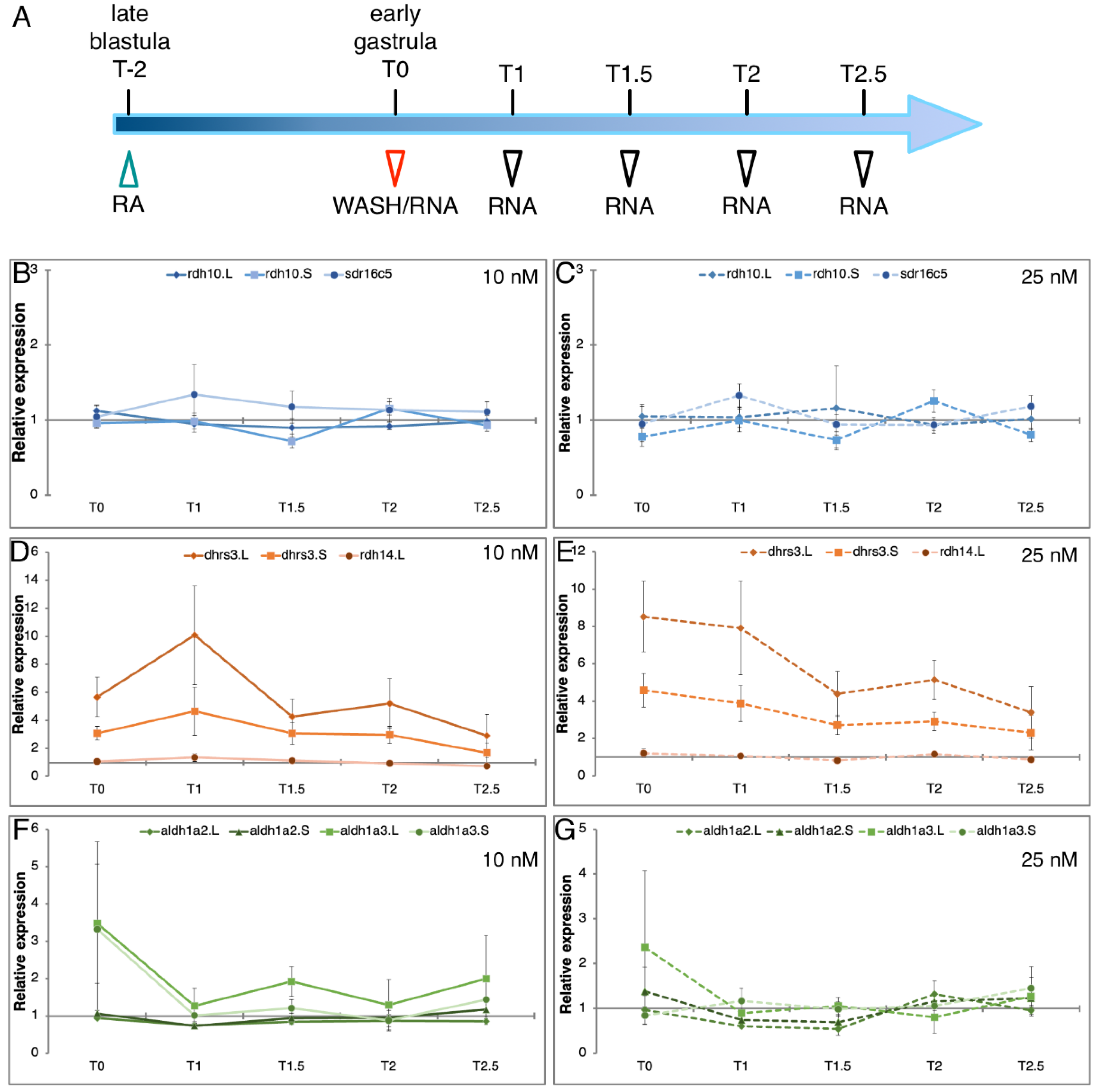
3.4. Homoeolog Response to Transient RA Manipulation
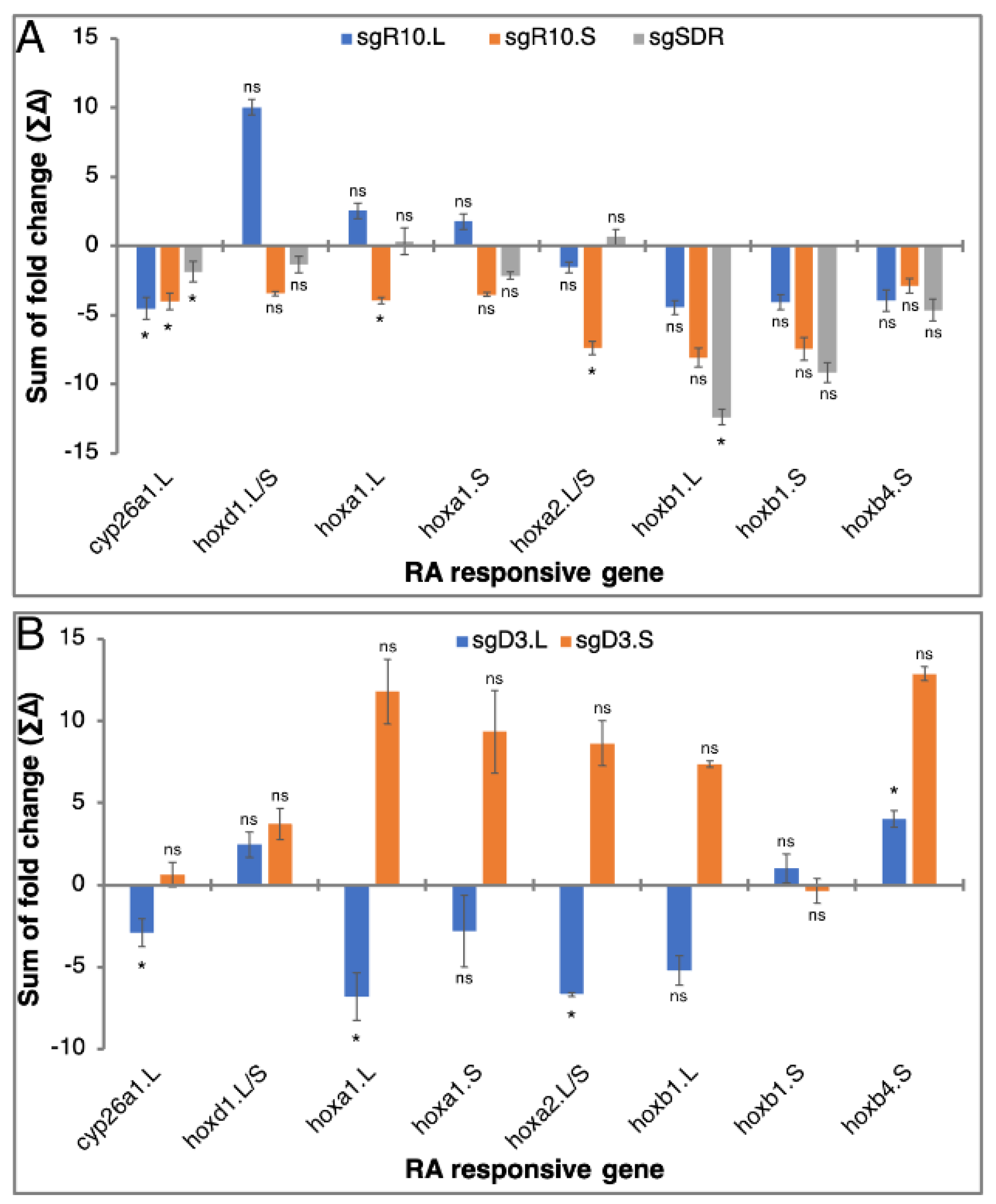
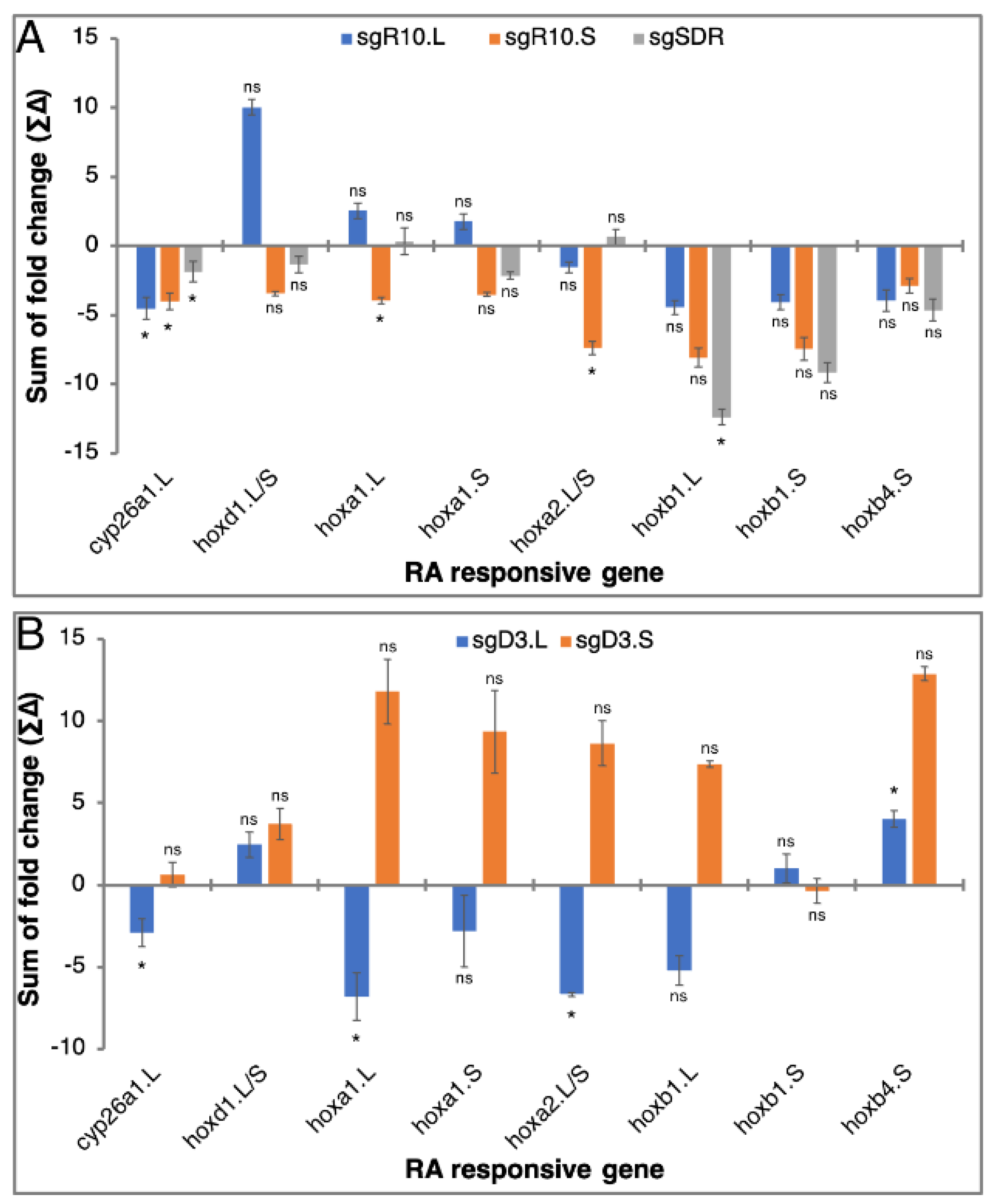
3.5. RA Responsiveness in Homoeolog CRISPant Embryos
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Metzler, M.A.; Sandell, L.L. Enzymatic metabolism of vitamin A in developing vertebrate embryos. Nutrients 2016, 8, 812. [Google Scholar] [CrossRef]
- le Maire, A.; Bourguet, W. Retinoic acid receptors: Structural basis for coregulator interaction and exchange. Subcell. Biochem. 2014, 70, 37–54. [Google Scholar] [CrossRef]
- Nolte, C.; De Kumar, B.; Krumlauf, R. Hox genes: Downstream “effectors” of retinoic acid signaling in vertebrate embryogenesis. Genesis 2019, 57, e23306. [Google Scholar] [CrossRef]
- Draut, H.; Liebenstein, T.; Begemann, G. New Insights into the Control of Cell Fate Choices and Differentiation by Retinoic Acid in Cranial, Axial and Caudal Structures. Biomolecules 2019, 9, 860. [Google Scholar] [CrossRef] [Green Version]
- Blaner, W.S. Vitamin A signaling and homeostasis in obesity, diabetes, and metabolic disorders. Pharmacol. Ther. 2019, 197, 153–178. [Google Scholar] [CrossRef]
- Napoli, J.L. Post-natal all-trans-retinoic acid biosynthesis. Meth. Enzymol. 2020, 637, 27–54. [Google Scholar] [CrossRef]
- Blaner, W.S.; Li, Y.; Brun, P.-J.; Yuen, J.J.; Lee, S.-A.; Clugston, R.D. Vitamin A absorption, storage and mobilization. Subcell. Biochem. 2016, 81, 95–125. [Google Scholar] [CrossRef]
- Ghyselinck, N.B.; Duester, G. Retinoic acid signaling pathways. Development 2019, 146. [Google Scholar] [CrossRef] [Green Version]
- Kedishvili, N.Y. Retinoic acid synthesis and degradation. Subcell. Biochem. 2016, 81, 127–161. [Google Scholar] [CrossRef] [Green Version]
- Shabtai, Y.; Fainsod, A. Competition between ethanol clearance and retinoic acid biosynthesis in the induction of fetal alcohol syndrome. Biochem. Cell Biol. 2018, 96, 148–160. [Google Scholar] [CrossRef]
- Collins, M.D.; Mao, G.E. Teratology of retinoids. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 399–430. [Google Scholar] [CrossRef]
- Mark, M.; Ghyselinck, N.B.; Chambon, P. Function of retinoid nuclear receptors: Lessons from genetic and pharmacological dissections of the retinoic acid signaling pathway during mouse embryogenesis. Annu. Rev. Pharmacol. Toxicol. 2006, 46, 451–480. [Google Scholar] [CrossRef]
- Fainsod, A.; Bendelac-Kapon, L.; Shabtai, Y. Fetal alcohol spectrum disorder: Embryogenesis under reduced retinoic acid signaling conditions. Subcell. Biochem. 2020, 95, 197–225. [Google Scholar] [CrossRef] [PubMed]
- Fainsod, A.; Kot-Leibovich, H. Xenopus embryos to study fetal alcohol syndrome, a model for environmental teratogenesis. Biochem. Cell Biol. 2018, 96, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Shabtai, Y.; Bendelac, L.; Jubran, H.; Hirschberg, J.; Fainsod, A. Acetaldehyde inhibits retinoic acid biosynthesis to mediate alcohol teratogenicity. Sci. Rep. 2018, 8, 347. [Google Scholar] [CrossRef] [PubMed]
- Paganelli, A.; Gnazzo, V.; Acosta, H.; López, S.L.; Carrasco, A.E. Glyphosate-based herbicides produce teratogenic effects on vertebrates by impairing retinoic acid signaling. Chem. Res. Toxicol. 2010, 23, 1586–1595. [Google Scholar] [CrossRef]
- Dickinson, A.J.G.; Turner, S.D.; Wahl, S.; Kennedy, A.E.; Wyatt, B.H.; Howton, D.A. E-liquids and vanillin flavoring disrupts retinoic acid signaling and causes craniofacial defects in Xenopus embryos. Dev. Biol. 2022, 481, 14–29. [Google Scholar] [CrossRef]
- Niederreither, K.; McCaffery, P.; Dräger, U.C.; Chambon, P.; Dollé, P. Restricted expression and retinoic acid-induced downregulation of the retinaldehyde dehydrogenase type 2 (RALDH-2) gene during mouse development. Mech. Dev. 1997, 62, 67–78. [Google Scholar] [CrossRef]
- Pavez Loriè, E.; Chamcheu, J.C.; Vahlquist, A.; Törmä, H. Both all-trans retinoic acid and cytochrome P450 (CYP26) inhibitors affect the expression of vitamin A metabolizing enzymes and retinoid biomarkers in organotypic epidermis. Arch. Dermatol. Res. 2009, 301, 475–485. [Google Scholar] [CrossRef]
- Moss, J.B.; Xavier-Neto, J.; Shapiro, M.D.; Nayeem, S.M.; McCaffery, P.; Dräger, U.C.; Rosenthal, N. Dynamic patterns of retinoic acid synthesis and response in the developing mammalian heart. Dev. Biol. 1998, 199, 55–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollemann, T.; Chen, Y.; Grunz, H.; Pieler, T. Regionalized metabolic activity establishes boundaries of retinoic acid signalling. EMBO J. 1998, 17, 7361–7372. [Google Scholar] [CrossRef] [Green Version]
- Topletz, A.R.; Tripathy, S.; Foti, R.S.; Shimshoni, J.A.; Nelson, W.L.; Isoherranen, N. Induction of CYP26A1 by metabolites of retinoic acid: Evidence that CYP26A1 is an important enzyme in the elimination of active retinoids. Mol. Pharmacol. 2015, 87, 430–441. [Google Scholar] [CrossRef] [Green Version]
- Reijntjes, S.; Zile, M.H.; Maden, M. The expression of Stra6 and Rdh10 in the avian embryo and their contribution to the generation of retinoid signatures. Int. J. Dev. Biol. 2010, 54, 1267–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strate, I.; Min, T.H.; Iliev, D.; Pera, E.M. Retinol dehydrogenase 10 is a feedback regulator of retinoic acid signalling during axis formation and patterning of the central nervous system. Development 2009, 136, 461–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.K.; Belyaeva, O.V.; Wu, L.; Kedishvili, N.Y. The retinaldehyde reductase activity of DHRS3 is reciprocally activated by retinol dehydrogenase 10 to control retinoid homeostasis. J. Biol. Chem. 2014, 289, 14868–14880. [Google Scholar] [CrossRef] [Green Version]
- Kam, R.K.T.; Chen, Y.; Chan, S.-O.; Chan, W.-Y.; Dawid, I.B.; Zhao, H. Developmental expression of Xenopus short-chain dehydrogenase/reductase 3. Int. J. Dev. Biol. 2010, 54, 1355–1360. [Google Scholar] [CrossRef] [Green Version]
- Parihar, M.; Bendelac-Kapon, L.; Gur, M.; Abbou, T.; Belorkar, A.; Achanta, S.; Kinberg, K.; Vadigepalli, R.; Fainsod, A. Retinoic Acid Fluctuation Activates an Uneven, Direction-Dependent Network-Wide Robustness Response in Early Embryogenesis. Front. Cell Dev. Biol. 2021, 9, 747969. [Google Scholar] [CrossRef]
- Eldar, A.; Shilo, B.-Z.; Barkai, N. Elucidating mechanisms underlying robustness of morphogen gradients. Curr. Opin. Genet. Dev. 2004, 14, 435–439. [Google Scholar] [CrossRef]
- Ohno, S.; Wolf, U.; Atkin, N.B. Evolution from fish to mammals by gene duplication. Hereditas 1968, 59, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S. Gene duplication and the uniqueness of vertebrate genomes circa 1970-1999. Semin. Cell Dev. Biol. 1999, 10, 517–522. [Google Scholar] [CrossRef] [PubMed]
- MacKintosh, C.; Ferrier, D.E.K. Recent advances in understanding the roles of whole genome duplications in evolution. [version 2; peer review: 2 approved]. F1000Research 2017, 6, 1623. [Google Scholar] [CrossRef] [Green Version]
- Session, A.M.; Uno, Y.; Kwon, T.; Chapman, J.A.; Toyoda, A.; Takahashi, S.; Fukui, A.; Hikosaka, A.; Suzuki, A.; Kondo, M.; et al. Genome evolution in the allotetraploid frog Xenopus laevis. Nature 2016, 538, 336–343. [Google Scholar] [CrossRef] [Green Version]
- Wolf, Y.I.; Koonin, E.V. Genome reduction as the dominant mode of evolution. Bioessays 2013, 35, 829–837. [Google Scholar] [CrossRef] [Green Version]
- Newton, M.S.; Arcus, V.L.; Gerth, M.L.; Patrick, W.M. Enzyme evolution: Innovation is easy, optimization is complicated. Curr. Opin. Struct. Biol. 2018, 48, 110–116. [Google Scholar] [CrossRef]
- Albalat, R.; Cañestro, C. Evolution by gene loss. Nat. Rev. Genet. 2016, 17, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Morin, R.D.; Chang, E.; Petrescu, A.; Liao, N.; Griffith, M.; Chow, W.; Kirkpatrick, R.; Butterfield, Y.S.; Young, A.C.; Stott, J.; et al. Sequencing and analysis of 10,967 full-length cDNA clones from Xenopus laevis and Xenopus tropicalis reveals post-tetraploidization transcriptome remodeling. Genome Res. 2006, 16, 796–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellsten, U.; Harland, R.M.; Gilchrist, M.J.; Hendrix, D.; Jurka, J.; Kapitonov, V.; Ovcharenko, I.; Putnam, N.H.; Shu, S.; Taher, L.; et al. The genome of the Western clawed frog Xenopus tropicalis. Science 2010, 328, 633–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sémon, M.; Wolfe, K.H. Preferential subfunctionalization of slow-evolving genes after allopolyploidization in Xenopus laevis. Proc. Natl. Acad. Sci. USA 2008, 105, 8333–8338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chain, F.J.J.; Dushoff, J.; Evans, B.J. The odds of duplicate gene persistence after polyploidization. BMC Genom. 2011, 12, 599. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Yasuoka, Y.; Mawaribuchi, S.; Kuretani, A.; Ito, M.; Kondo, M.; Ochi, H.; Ogino, H.; Fukui, A.; Taira, M.; et al. Conservatism and variability of gene expression profiles among homeologous transcription factors in Xenopus laevis. Dev. Biol. 2017, 426, 301–324. [Google Scholar] [CrossRef]
- Michiue, T.; Yamamoto, T.; Yasuoka, Y.; Goto, T.; Ikeda, T.; Nagura, K.; Nakayama, T.; Taira, M.; Kinoshita, T. High variability of expression profiles of homeologous genes for Wnt, Hh, Notch, and Hippo signaling pathways in Xenopus laevis. Dev. Biol. 2017, 426, 270–290. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, A.; Yoshida, H.; van Heeringen, S.J.; Takebayashi-Suzuki, K.; Veenstra, G.J.C.; Taira, M. Genomic organization and modulation of gene expression of the TGF-β and FGF pathways in the allotetraploid frog Xenopus laevis. Dev. Biol. 2017, 426, 336–359. [Google Scholar] [CrossRef]
- Nieuwkoop, P.D.; Faber, J. Normal Table of Xenopus laevis (Daudin): A Systematical and Chronological Survey of the Development from the Fertilized Egg till the End of Metamorphosis; North-Holland Publishing Company: Amsterdam, The Netherland, 1967; p. 260. [Google Scholar]
- Nenni, M.J.; Fisher, M.E.; James-Zorn, C.; Pells, T.J.; Ponferrada, V.; Chu, S.; Fortriede, J.D.; Burns, K.A.; Wang, Y.; Lotay, V.S.; et al. Xenbase: Facilitating the use of xenopus to model human disease. Front. Physiol. 2019, 10, 154. [Google Scholar] [CrossRef] [Green Version]
- Naito, Y.; Hino, K.; Bono, H.; Ui-Tei, K. CRISPRdirect: Software for designing CRISPR/Cas guide RNA with reduced off-target sites. Bioinformatics 2015, 31, 1120–1123. [Google Scholar] [CrossRef]
- Moreno-Mateos, M.A.; Vejnar, C.E.; Beaudoin, J.-D.; Fernandez, J.P.; Mis, E.K.; Khokha, M.K.; Giraldez, A.J. CRISPRscan: Designing highly efficient sgRNAs for CRISPR-Cas9 targeting in vivo. Nat. Methods 2015, 12, 982–988. [Google Scholar] [CrossRef] [Green Version]
- Naert, T.; Tulkens, D.; Edwards, N.A.; Carron, M.; Shaidani, N.-I.; Wlizla, M.; Boel, A.; Demuynck, S.; Horb, M.E.; Coucke, P.; et al. Maximizing CRISPR/Cas9 phenotype penetrance applying predictive modeling of editing outcomes in Xenopus and zebrafish embryos. Sci. Rep. 2020, 10, 14662. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.W.; Arbab, M.; Hsu, J.Y.; Worstell, D.; Culbertson, S.J.; Krabbe, O.; Cassa, C.A.; Liu, D.R.; Gifford, D.K.; Sherwood, R.I. Predictable and precise template-free CRISPR editing of pathogenic variants. Nature 2018, 563, 646–651. [Google Scholar] [CrossRef]
- Hoshijima, K.; Jurynec, M.J.; Klatt Shaw, D.; Jacobi, A.M.; Behlke, M.A.; Grunwald, D.J. Highly Efficient CRISPR-Cas9-Based Methods for Generating Deletion Mutations and F0 Embryos that Lack Gene Function in Zebrafish. Dev. Cell 2019, 51, 645–657.e4. [Google Scholar] [CrossRef]
- Brinkman, E.K.; Chen, T.; Amendola, M.; van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Res. 2014, 42, e168. [Google Scholar] [CrossRef]
- Hsiau, T.; Maures, T.; Waite, K.; Yang, J.; Kelso, R.; Holden, K.; Stoner, R. Inference of CRISPR Edits from Sanger Trace Data. BioRxiv 2018. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2021. [Google Scholar] [CrossRef]
- Yanai, I.; Peshkin, L.; Jorgensen, P.; Kirschner, M.W. Mapping gene expression in two Xenopus species: Evolutionary constraints and developmental flexibility. Dev. Cell 2011, 20, 483–496. [Google Scholar] [CrossRef] [Green Version]
- Savova, V.; Pearl, E.J.; Boke, E.; Nag, A.; Adzhubei, I.; Horb, M.E.; Peshkin, L. Transcriptomic insights into genetic diversity of protein-coding genes in X. laevis. Dev. Biol. 2017, 424, 181–188. [Google Scholar] [CrossRef]
- Shannon, S.R.; Moise, A.R.; Trainor, P.A. New insights and changing paradigms in the regulation of vitamin A metabolism in development. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Belyaeva, O.V.; Adams, M.K.; Wu, L.; Kedishvili, N.Y. The antagonistically bifunctional retinoid oxidoreductase complex is required for maintenance of all-trans-retinoic acid homeostasis. J. Biol. Chem. 2017, 292, 5884–5897. [Google Scholar] [CrossRef] [Green Version]
- Shabtai, Y.; Shukrun, N.; Fainsod, A. ADHFe1: A novel enzyme involved in retinoic acid-dependent Hox activation. Int. J. Dev. Biol. 2017, 61, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Billings, S.E.; Pierzchalski, K.; Butler Tjaden, N.E.; Pang, X.-Y.; Trainor, P.A.; Kane, M.A.; Moise, A.R. The retinaldehyde reductase DHRS3 is essential for preventing the formation of excess retinoic acid during embryonic development. FASEB J. 2013, 27, 4877–4889. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.; Hernandez, R.E.; Waxman, J.S.; Yelon, D.; Moens, C.B. Dhrs3a regulates retinoic acid biosynthesis through a feedback inhibition mechanism. Dev. Biol. 2010, 338, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Porté, S.; Xavier Ruiz, F.; Giménez, J.; Molist, I.; Alvarez, S.; Domínguez, M.; Alvarez, R.; de Lera, A.R.; Parés, X.; Farrés, J. Aldo-keto reductases in retinoid metabolism: Search for substrate specificity and inhibitor selectivity. Chem. Biol. Interact. 2013, 202, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Michaille, J.-J.; Jiang, W.; Zile, M.H. Retinoid receptors and vitamin A deficiency: Differential patterns of transcription during early avian development and the rapid induction of RARs by retinoic acid. Dev. Biol. 2003, 260, 496–511. [Google Scholar] [CrossRef]
- Lohnes, D.; Mark, M.; Mendelsohn, C.; Dollé, P.; Decimo, D.; LeMeur, M.; Dierich, A.; Gorry, P.; Chambon, P. Developmental roles of the retinoic acid receptors. J. Steroid Biochem. Mol. Biol. 1995, 53, 475–486. [Google Scholar] [CrossRef]
- Janesick, A.; Wu, S.C.; Blumberg, B. Retinoic acid signaling and neuronal differentiation. Cell. Mol. Life Sci. 2015, 72, 1559–1576. [Google Scholar] [CrossRef] [Green Version]
- De Bosscher, K.; Desmet, S.J.; Clarisse, D.; Estébanez-Perpiña, E.; Brunsveld, L. Nuclear receptor crosstalk-defining the mechanisms for therapeutic innovation. Nat. Rev. Endocrinol. 2020, 16, 363–377. [Google Scholar] [CrossRef]
- Evans, R.M.; Mangelsdorf, D.J. Nuclear receptors, RXR, and the big bang. Cell 2014, 157, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Niederreither, K.; Subbarayan, V.; Dollé, P.; Chambon, P. Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nat. Genet. 1999, 21, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Begemann, G.; Schilling, T.F.; Rauch, G.J.; Geisler, R.; Ingham, P.W. The zebrafish neckless mutation reveals a requirement for raldh2 in mesodermal signals that pattern the hindbrain. Development 2001, 128, 3081–3094. [Google Scholar] [CrossRef]
- Grandel, H.; Lun, K.; Rauch, G.-J.; Rhinn, M.; Piotrowski, T.; Houart, C.; Sordino, P.; Küchler, A.M.; Schulte-Merker, S.; Geisler, R.; et al. Retinoic acid signalling in the zebrafish embryo is necessary during pre-segmentation stages to pattern the anterior-posterior axis of the CNS and to induce a pectoral fin bud. Development 2002, 129, 2851–2865. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Pollet, N.; Niehrs, C.; Pieler, T. Increased XRALDH2 activity has a posteriorizing effect on the central nervous system of Xenopus embryos. Mech. Dev. 2001, 101, 91–103. [Google Scholar] [CrossRef]
- Durston, A.J.; Timmermans, J.P.; Hage, W.J.; Hendriks, H.F.; de Vries, N.J.; Heideveld, M.; Nieuwkoop, P.D. Retinoic acid causes an anteroposterior transformation in the developing central nervous system. Nature 1989, 340, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Schuh, T.J.; Hall, B.L.; Kraft, J.C.; Privalsky, M.L.; Kimelman, D. v-erbA and citral reduce the teratogenic effects of all-trans retinoic acid and retinol, respectively, in Xenopus embryogenesis. Development 1993, 119, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, L.; Solursh, M. A concentration gradient of retinoids in the early Xenopus laevis embryo. Dev. Biol. 1994, 161, 70–76. [Google Scholar] [CrossRef]
- Kraft, J.C.; Schuh, T.; Juchau, M.; Kimelman, D. The retinoid X receptor ligand, 9-cis-retinoic acid, is a potential regulator of early Xenopus development. Proc. Natl. Acad. Sci. USA 1994, 91, 3067–3071. [Google Scholar] [CrossRef] [Green Version]
- Creech Kraft, J.; Schuh, T.; Juchau, M.R.; Kimelman, D. Temporal distribution, localization and metabolism of all-trans-retinol, didehydroretinol and all-trans-retinal during Xenopus development. Biochem. J. 1994, 301 Pt 1, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Creech Kraft, J.; Kimelman, D.; Juchau, M.R. Xenopus laevis: A model system for the study of embryonic retinoid metabolism. II. Embryonic metabolism of all-trans-3,4-didehydroretinol to all-trans-3,4-didehydroretinoic acid. Drug Metab. Dispos. 1995, 23, 83–89. [Google Scholar] [PubMed]
- Kraft, J.C.; Kimelman, D.; Juchau, M.R. Xenopus laevis: A model system for the study of embryonic retinoid metabolism. I. Embryonic metabolism of 9-cis- and all-trans-retinals and retinols to their corresponding acid forms. Drug Metab. Dispos. 1995, 23, 72–82. [Google Scholar]
- Dobbs-McAuliffe, B.; Zhao, Q.; Linney, E. Feedback mechanisms regulate retinoic acid production and degradation in the zebrafish embryo. Mech. Dev. 2004, 121, 339–350. [Google Scholar] [CrossRef]
- Sive, H.L.; Draper, B.W.; Harland, R.M.; Weintraub, H. Identification of a retinoic acid-sensitive period during primary axis formation in Xenopus laevis. Genes Dev. 1990, 4, 932–942. [Google Scholar] [CrossRef] [Green Version]
- Kessel, M.; Gruss, P. Homeotic transformations of murine vertebrae and concomitant alteration of Hox codes induced by retinoic acid. Cell 1991, 67, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Papalopulu, N.; Clarke, J.D.; Bradley, L.; Wilkinson, D.; Krumlauf, R.; Holder, N. Retinoic acid causes abnormal development and segmental patterning of the anterior hindbrain in Xenopus embryos. Development 1991, 113, 1145–1158. [Google Scholar] [CrossRef]
- Corcoran, J.; So, P.L.; Maden, M. Absence of retinoids can induce motoneuron disease in the adult rat and a retinoid defect is present in motoneuron disease patients. J. Cell Sci. 2002, 115, 4735–4741. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.K.; Lee, S.-A.; Belyaeva, O.V.; Wu, L.; Kedishvili, N.Y. Characterization of human short chain dehydrogenase/reductase SDR16C family members related to retinol dehydrogenase 10. Chem. Biol. Interact. 2017, 276, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Belyaeva, O.V.; Lee, S.-A.; Adams, M.K.; Chang, C.; Kedishvili, N.Y. Short chain dehydrogenase/reductase rdhe2 is a novel retinol dehydrogenase essential for frog embryonic development. J. Biol. Chem. 2012, 287, 9061–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belyaeva, O.V.; Kedishvili, N.Y. Human pancreas protein 2 (PAN2) has a retinal reductase activity and is ubiquitously expressed in human tissues. FEBS Lett. 2002, 531, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Kam, R.K.T.; Shi, W.; Chan, S.O.; Chen, Y.; Xu, G.; Lau, C.B.-S.; Fung, K.P.; Chan, W.Y.; Zhao, H. Dhrs3 protein attenuates retinoic acid signaling and is required for early embryonic patterning. J. Biol. Chem. 2013, 288, 31477–31487. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| qPCR Analysis | ||
| cyp26a1.L | TCGAGGTTCGGCTTCATC | CGGCACAATTCCACAACA |
| hoxb1.L | TTGCCCCAGTGCCAATGAC | TCCCCCTCCAACAACAAACC |
| hoxb1.S | CCAACTTCACGACCAAACAA | GTGGCTGCGATCTCTACTCTC |
| hoxd1.L/S | TTCTTGCGGGGATGTTTT | CCGACTGGCATAAAGGAATG |
| hoxa1.L | CCGCTCACTATATCCACCATTC | TGGCAGGAGAACGACAAAC |
| hoxa1.S | AATTATGAGATGATGGAATGGTAAA | TGACTGTAAACACCTAGTAAATGAGAG |
| hoxb4.S | CCAAGGATCTGTGCGTCAA | GCAGGATGGAGGCGAACT |
| hoxa2.L/S | GGAGATTGCAGCCCTGTT | GGGTTTGCCTCTTGTGTTTC |
| sdr16c5.L | TTTGTGGTTCCTTCCCTCTC | GTGCCATCAGTCTCCCTATACC |
| rdh14.L | TGCCCGTACACAAAGACAGA | GAGACCAAGGAGGTGGTGAG |
| dhrs3.L | CAGGCGCAAGAAATCCTAAG | CAAAGGCCACGTTCAAGGAT |
| dhrs3.S | TGCAGATGGTATTGTCCCTTC | TCCTTAGCGAGGTGTCGG |
| rdh10.L | CGTCTCTTTGCCCTGGAGTTT | CACCATCTCCGCCGTCTC |
| rdh10.S | TTGCTTGGCCTGTAGAAGAGA | TGCATGGCGAAATAGGAGTAG |
| slc35b1.L | CGCATTTCCAAACAGGCTCC | CAAGAAGTCCCAGAGCTCGC |
| Genomic Nested PCR | ||
| Outer Genomic PCR | ||
| dhrs3.L | TGACTGTAAGAATAGCCGCGT | AGCGGGCAGACAAGACAAAT |
| dhrs3.S | ACCGCTATAGAACCACAGTCG | GAAACACTTCATTCCTTTTAGTGGA |
| rdh10.L | TAAGTTGGCAGCGGTTTGGG | GAGAGACCCCACATAACTCAGC |
| rdh10.S | TCCAGAGCGAAAATCTGACGA | TCCCATGGTCATGAAACTCCTCAG |
| sdr16c5 | ACTGTCTTCATAGTCGAGCCC | TGGTCCGAATAGAAAATCTGGG |
| rdh14 | CACACCAAACATGGCGACTT | TGAAGGGCGTTGACTGTGAC |
| Inner Genomic PCR | ||
| sdr16c5 | GTTCCACCTTTCTGTCAATGCTC | AGCATTTGTCCTCAGCGTTTT |
| rdh14 | GAGTAACAGCGTCAGAGCCG | TGAAGGGCGTTGACTGTGAC |
| dhrs3.L | GAGTCTCAGCACAGGGCAAA | TCAAAGGGTGACAGGGAACG |
| dhrs3.S | CACAGTCGTTGGCTTGAGTG | CCTTTCACTTTTTGCAGGATTCA |
| rdh10.L | CGCACGGAACTTACTGTCCA | TTGGACCCTGGAGCTGTACT |
| rdh10.S | TCCAGAGCGAAAATCTGACGA | GAGTGGCAGTAGAGTGAAGTCAG |
| sgRNA (crRNA) | ||
| sdr16c5 | AAACAAAGAGACCTGTAGAC | |
| rdh14 | GTCTGTCCGGCGATTCTGTC | |
| dhrs3.L | CTTATTGGGCATCAGCAAGT | |
| dhrs3.S | GTCCTAGTGTGTTAATGTGT | |
| rdh10.L | GCGCAGCAGCCACTTGCCCG | |
| rdh10.S | AGGCGGAGGACTCTGCGCGG | |
| Pathway | Total Genes | Singletons | % Singletons |
|---|---|---|---|
| High Singleton | |||
| Vitamin D from 7-dehydrocholesterol 1 | 9 | 6 | 66.7% |
| RA up to RAL 1 | 28 | 21 | 75% |
| Folic acid metabolism 1 | 27 | 19 | 70.4% |
| DNA repair | 57 | 45 | 78.9% |
| Average Genome Singletons | |||
| De novo Purine biosynthesis 1 | 15 | 7 | 46.7% |
| Thyroid hormone synthesis 1 | 52 | 23 | 44.2% |
| Protein-coding 2 | >13,781 | >6008 | 43.6% |
| RA signaling metabolism and signaling | 47 | 22 | 46.8% |
| Vitamin D incl. 7-dehydrocholesterol 1 | 21 | 8 | 38.1% |
| Suppressed Singletons | |||
| Glycolysis/Gluconeogenesis/Krebs cycle 1 | 44 | 9 | 20.5% |
| Notch 3 | 48 | 8 | 16.7% |
| MicroRNAs 2 | 180 | 24 | 13.3% |
| Transcription Factors 4 | 218 | 28 | 12.8% |
| Wnt 3 | 108 | 13 | 12% |
| Hippo 3 | 48 | 5 | 10.4% |
| BMP/TGFβ 5 | 126 | 13 | 10.3% |
| RA from RAL 1 | 19 | 1 | 5.2% |
| FGF 5 | 60 | 3 | 5% |
| cis-regulatory elements (non-coding) 2 | 550 | 9 | 1.6% |
| Hh 3 | 18 | 0 | 0% |
| HSPG 3 | 16 | 0 | 0% |
| TLE 3 | 4 | 0 | 0% |
| Name | L/S 1 | Transcribed Region Length (bp) | Flanking Genes 2 | Genes in the L Genomic Region | Genes in the S Genomic Region | Genomic Length (bp) 3 | Length Difference (Kb) | Deletion Type | |
|---|---|---|---|---|---|---|---|---|---|
| L | S | ||||||||
| Producers 1 | |||||||||
| sdr16c5 | 6L | 18,952 | plag1-penk | chchd7, sdr16c5 | - | 89,214 | 79,69 | 9.5 | Restricted |
| adh7 adh4 adh1b | 1S 1L 1L | 9948 13,797 18,159 | dapp1-metap1 | LOC108710849, MGC83376, adh1b, adh4, adh1-a, LOC108719623, adh5 | LOC398377, LOC108706741, adh1a, adh7 | 277,881 | 196,627 | 81.3 | Complex |
| rdh16 | 2L | 11,580 | rdh7.2-gpr182 | rdh16, XB5807236, rdh7, LOC108708580, rdh9, hsd17b6, LOC10870858 | - | 116,911 | 14,959 | 102.0 | Large |
| dhrs9 | 9_10L | 9073 | stk39-klhl41 | dhrs9, LOC108701361, nostrin, LOC108701845, XB5957220, lrp2, LOC108701363, LOC108701365, LOC108701364, bbs5 | LOC108702983, abcb11.2, LOC108702985, LOC108702267 | 521,928 | 283,492 | 238.4 | Complex |
| Suppressors | |||||||||
| adhFe1 | 6S | 19,503 | rrs1-mybl1 | LOC108718816, vxn | LOC121395160, adhFe1 | 63.4 | 63.3 | 0.1 | Restricted |
| rdh13 | 5L | 10,266 | slc30a6-sos1 | rdh13 | - | 42,225 | 5878 | 36.3 | Restricted |
| rdh14 | 5L | 4946 | kcns3- osr1 | LOC121393845, LOC108717194, LOC121393566, rdh14 | - | 609,595 | 488,124 | 121.5 | Large |
| Producers 3 | |||||||||
| bco1 | 4L | 20,949 | gan- cenpn | bco1, LOC108714031, LOC108714032, gcsh, LOC108714033, LOC108714034, LOC108714035, cdk10, LOC121403143, LOC121403144, LOC108714036, LOC108714037, atmin | LOC108708941, LOC108715382, LOC108715380, LOC108705941 | 306,957 | 114,993 | 192.0 | Large |
| bco2 bco2l | 2L 2L | 14,883 38,972 | urpta-1-zbtb16 | sfxn4, bco2, bco2l, LOC121395554, LOC108697004, nnmt, LOC108695837, LOC108696501, LOC108697006, LOC108696502, LOC121395555, LOC108696503, LOC108697007, LOC121395556, LOC108696506 | LOC108697615 | 611,546 | 200,702 | 410.8 | Large |
| Gene | ∑∆ Score 1 | |
|---|---|---|
| 10 nM RA | 25 nM RA | |
| Suppressors | ||
| dhrs3.L | 28.1 | 29.4 |
| dhrs3.S | 15.5 | 16.4 |
| rdh14.L | 5.2 | 5.1 |
| Producers 1 | ||
| rdh10.L | 4.9 | 5.2 |
| rdh10.S | 4.8 | 4.6 |
| sdr16c5 | 5.8 | 5.3 |
| Producers 2 | ||
| aldh1a2.L | 4.3 | 4.4 |
| aldh1a2.S | 4.9 | 5.2 |
| aldh1a3.L | 10.0 | 6.4 |
| aldh1a3.S | 7.8 | 5.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbou, T.; Bendelac-Kapon, L.; Sebag, A.; Fainsod, A. Enhanced Loss of Retinoic Acid Network Genes in Xenopus laevis Achieves a Tighter Signal Regulation. Cells 2022, 11, 327. https://doi.org/10.3390/cells11030327
Abbou T, Bendelac-Kapon L, Sebag A, Fainsod A. Enhanced Loss of Retinoic Acid Network Genes in Xenopus laevis Achieves a Tighter Signal Regulation. Cells. 2022; 11(3):327. https://doi.org/10.3390/cells11030327
Chicago/Turabian StyleAbbou, Tali, Liat Bendelac-Kapon, Audeliah Sebag, and Abraham Fainsod. 2022. "Enhanced Loss of Retinoic Acid Network Genes in Xenopus laevis Achieves a Tighter Signal Regulation" Cells 11, no. 3: 327. https://doi.org/10.3390/cells11030327
APA StyleAbbou, T., Bendelac-Kapon, L., Sebag, A., & Fainsod, A. (2022). Enhanced Loss of Retinoic Acid Network Genes in Xenopus laevis Achieves a Tighter Signal Regulation. Cells, 11(3), 327. https://doi.org/10.3390/cells11030327