

Polymerized Laminin-521: A Feasible Substrate for Expanding Induced Pluripotent Stem Cells at a Low Protein Concentration
 , ,
, ,
Abstract
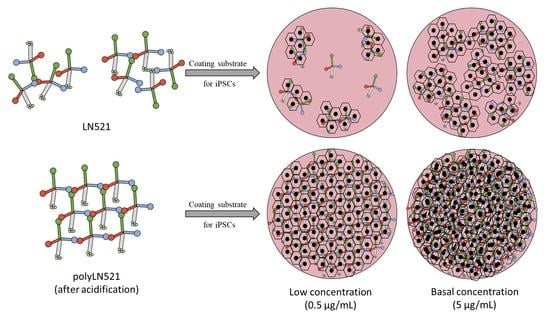
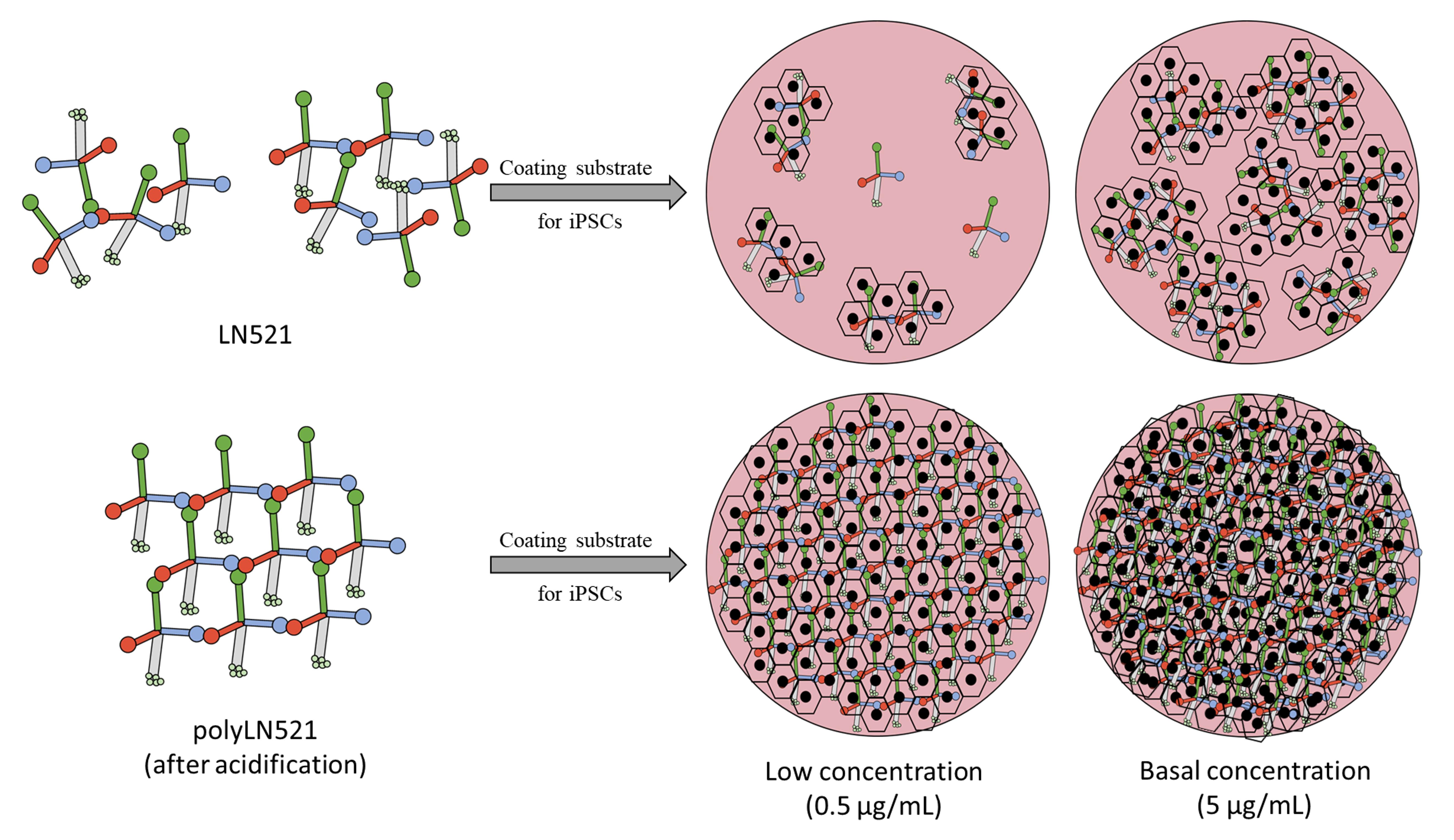{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Microscopy
2.2. Coating Preparation
2.3. Cell Expansion
2.4. Colony Area
2.5. DNA Extraction
2.6. Genetic Analysis of hiPSCs
2.7. Telomere Length
2.8. Gene Expression
2.9. Flow Cytometry
2.10. Differentiation Potential of hiPSCs
2.11. Large-Scale Expansion of hiPSCs
2.12. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, G.; David, B.T.; Trawczynski, M.; Fessler, R.G. Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and Applications. Stem Cell Rev. Rep. 2020, 16, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Goetzke, R.; Keijdener, H.; Franzen, J.; Ostrowska, A.; Nuchtern, S.; Mela, P.; Wagner, W. Differentiation of Induced Pluripotent Stem Cells towards Mesenchymal Stromal Cells is Hampered by Culture in 3D Hydrogels. Sci. Rep. 2019, 9, 15578. [Google Scholar] [CrossRef] [PubMed]
- Colter, J.; Murari, K.; Biernaskie, J.; Kallos, M.S. Induced pluripotency in the context of stem cell expansion bioprocess development, optimization, and manufacturing: A roadmap to the clinic. NPJ Regen. Med. 2021, 6, 72. [Google Scholar] [CrossRef] [PubMed]
- Brehm, J.L.; Ludwig, T.E. Culture, Adaptation, and Expansion of Pluripotent Stem Cells. Methods Mol. Biol. 2017, 1590, 139–150. [Google Scholar] [PubMed]
- Chen, G.; Gulbranson, D.R.; Hou, Z.; Bolin, J.M.; Ruotti, V.; Probasco, M.D.; Smuga-Otto, K.; Howden, S.E.; Diol, N.R.; Propson, N.E.; et al. Chemically defined conditions for human iPSC derivation and culture. Nat. Methods 2011, 8, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Aisenbrey, E.A.; Murphy, W.L. Synthetic alternatives to Matrigel. Nat. Rev. Mater. 2020, 5, 539–551. [Google Scholar] [CrossRef]
- Dayem, A.A.; Won, J.; Goo, H.G.; Yang, G.M.; Seo, D.S.; Jeon, B.M.; Choi, H.Y.; Park, S.E.; Lim, K.M.; Jang, S.H.; et al. The immobilization of fibronectin- and fibroblast growth factor 2-derived peptides on a culture plate supports the attachment and proliferation of human pluripotent stem cells. Stem Cell Res. 2020, 43, 101700. [Google Scholar] [CrossRef]
- Nasir, A.; Thorpe, J.; Burroughs, L.; Meurs, J.; Pijuan-Galito, S.; Irvine, D.J.; Alexander, M.R.; Denning, C. Discovery of a Novel Polymer for Xeno-Free, Long-Term Culture of Human Pluripotent Stem Cell Expansion. Adv. Healthc. Mater. 2021, 10, e2001448. [Google Scholar] [CrossRef]
- Ireland, R.G.; Kibschull, M.; Audet, J.; Ezzo, M.; Hinz, B.; Lye, S.J.; Simmons, C.A. Combinatorial extracellular matrix microarray identifies novel bioengineered substrates for xeno-free culture of human pluripotent stem cells. Biomaterials 2020, 248, 120017. [Google Scholar] [CrossRef]
- Grant, D.S.; Tashiro, K.; Segui-Real, B.; Yamada, Y.; Martin, G.R.; Kleinman, H.K. Two different laminin domains mediate the differentiation of human endothelial cells into capillary-like structures in vitro. Cell 1989, 58, 933–943. [Google Scholar] [CrossRef]
- Li, M.L.; Aggeler, J.; Farson, D.A.; Hatier, C.; Hassell, J.; Bissell, M.J. Influence of a reconstituted basement membrane and its components on casein gene expression and secretion in mouse mammary epithelial cells. Proc. Natl. Acad. Sci. USA 1987, 84, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Dye, J.; Lawrence, L.; Linge, C.; Leach, L.; Firth, J.; Clark, P. Distinct patterns of microvascular endothelial cell morphology are determined by extracellular matrix composition. Endothelium 2004, 11, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y. Laminin: Loss-of-function studies. Cell. Mol. Life Sci. 2017, 74, 1095–1115. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, Y.; Omasa, T. What Kind of Signaling Maintains Pluripotency and Viability in Human-Induced Pluripotent Stem Cells Cultured on Laminin-511 with Serum-Free Medium? Biores Open Access 2016, 5, 84–93. [Google Scholar] [CrossRef]
- Lu, H.F.; Chai, C.; Lim, T.C.; Leong, M.F.; Lim, J.K.; Gao, S.; Lim, K.L.; Wan, A.C. A defined xeno-free and feeder-free culture system for the derivation, expansion and direct differentiation of transgene-free patient-specific induced pluripotent stem cells. Biomaterials 2014, 35, 2816–2826. [Google Scholar] [CrossRef] [PubMed]
- Paccola Mesquita, F.C.; Hochman-Mendez, C.; Morrissey, J.; Sampaio, L.C.; Taylor, D.A. Laminin as a Potent Substrate for Large-Scale Expansion of Human Induced Pluripotent Stem Cells in a Closed Cell Expansion System. Stem Cells Int. 2019, 2019, 9704945. [Google Scholar] [CrossRef]
- Nie, Y.; Wang, W.; Xu, X.; Zou, J.; Bhuvanesh, T.; Schulz, B.; Ma, N.; Lendlein, A. Enhancement of human induced pluripotent stem cells adhesion through multilayer laminin coating. Clin. Hemorheol. Microcirc. 2018, 70, 531–542. [Google Scholar] [CrossRef]
- Dziedzicka, D.; Markouli, C.; Barbe, L.; Spits, C.; Sermon, K.; Geens, M. A High Proliferation Rate is Critical for Reproducible and Standardized Embryoid Body Formation from Laminin-521-Based Human Pluripotent Stem Cell Cultures. Stem Cell Rev. Rep. 2016, 12, 721–730. [Google Scholar] [CrossRef]
- Liu, Y.C.; Ban, L.K.; Lee, H.H.; Lee, H.T.; Chang, Y.T.; Lin, Y.T.; Su, H.Y.; Hsu, S.T.; Higuchi, A. Laminin-511 and recombinant vitronectin supplementation enables human pluripotent stem cell culture and differentiation on conventional tissue culture polystyrene surfaces in xeno-free conditions. J. Mater. Chem. B 2021, 9, 8604–8614. [Google Scholar] [CrossRef]
- Hochman-Mendez, C.; Lacerda de Menezes, J.R.; Sholl-Franco, A.; Coelho-Sampaio, T. Polylaminin recognition by retinal cells. J. Neurosci. Res. 2014, 92, 24–34. [Google Scholar] [CrossRef]
- Freire, E.; Gomes, F.C.; Linden, R.; Neto, V.M.; Coelho-Sampaio, T. Structure of laminin substrate modulates cellular signaling for neuritogenesis. J. Cell Sci. 2002, 115, 4867–4876. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hochman-Mendez, C.; Cantini, M.; Moratal, D.; Salmeron-Sanchez, M.; Coelho-Sampaio, T. A fractal nature for polymerized laminin. PLoS ONE 2014, 9, e109388. [Google Scholar] [CrossRef]
- Coelho-Sampaio, T.; Tenchov, B.; Nascimento, M.A.; Hochman-Mendez, C.; Morandi, V.; Caarls, M.B.; Altankov, G. Type IV collagen conforms to the organization of polylaminin adsorbed on planar substrata. Acta Biomater. 2020, 111, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Freire, E.; Barroso, M.M.; Klier, R.N.; Coelho-Sampaio, T. Biocompatibility and structural stability of a laminin biopolymer. Macromol. Biosci. 2012, 12, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Palmero, C.Y.; Miranda-Alves, L.; Sant’Ana Barroso, M.M.; Souza, E.C.; Machado, D.E.; Palumbo-Junior, A.; Santos, C.A.; Portilho, D.M.; Mermelstein, C.S.; Takiya, C.M.; et al. The follicular thyroid cell line PCCL3 responds differently to laminin and to polylaminin, a polymer of laminin assembled in acidic pH. Mol. Cell Endocrinol. 2013, 376, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Kent, A.J.; Mayer, N.; Inman, J.L.; Hochman-Mendez, C.; Bissell, M.J.; Robertson, C. The microstructure of laminin-111 compensates for dystroglycan loss in mammary epithelial cells in downstream expression of milk proteins. Biomaterials 2019, 218, 119337. [Google Scholar] [CrossRef]
- Hochman-Mendez, C.; Mesquita, F.C.P.; Morrissey, J.; da Costa, E.C.; Hulsmann, J.; Tang-Quan, K.; Xi, Y.; Lee, P.F.; Sampaio, L.C.; Taylor, D.A. Restoring anatomical complexity of a left ventricle wall as a step toward bioengineering a human heart with human induced pluripotent stem cell-derived cardiac cells. Acta Biomater. 2022, 141, 48–58. [Google Scholar] [CrossRef]
- Haack-Sorensen, M.; Follin, B.; Juhl, M.; Brorsen, S.K.; Sondergaard, R.H.; Kastrup, J.; Ekblond, A. Culture expansion of adipose derived stromal cells. A closed automated Quantum Cell Expansion System compared with manual flask-based culture. J. Transl. Med. 2016, 14, 319. [Google Scholar] [CrossRef]
- Barroso, M.M.; Freire, E.; Limaverde, G.S.; Rocha, G.M.; Batista, E.J.; Weissmuller, G.; Andrade, L.R.; Coelho-Sampaio, T. Artificial laminin polymers assembled in acidic pH mimic basement membrane organization. J. Biol. Chem. 2008, 283, 11714–11720. [Google Scholar] [CrossRef]
- Yurchenco, P.D.; Cheng, Y.S.; Colognato, H. Laminin forms an independent network in basement membranes. J. Cell Biol. 1992, 117, 1119–1133. [Google Scholar] [CrossRef]
- Yurchenco, P.D.; Tsilibary, E.C.; Charonis, A.S.; Furthmayr, H. Models for the self-assembly of basement membrane. J. Histochem. Cytochem. 1986, 34, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Solanki, A.; Hamilos, A.; Levy, O.; Wen, K.; Yin, X.; Karp, J.M. Application of biomaterials to advance induced pluripotent stem cell research and therapy. EMBO J. 2015, 34, 987–1008. [Google Scholar] [CrossRef] [PubMed]
- Nesterenko, Y.; Hill, C.J.; Fleming, J.R.; Murray, P.; Mayans, O. The ZT Biopolymer: A Self-Assembling Protein Scaffold for Stem Cell Applications. Int. J. Mol. Sci. 2019, 20, 4299. [Google Scholar] [CrossRef] [PubMed]
- Rodin, S.; Antonsson, L.; Niaudet, C.; Simonson, O.E.; Salmela, E.; Hansson, E.M.; Domogatskaya, A.; Xiao, Z.; Damdimopoulou, P.; Sheikhi, M.; et al. Clonal culturing of human embryonic stem cells on laminin-521/E-cadherin matrix in defined and xeno-free environment. Nat. Commun. 2014, 5, 3195. [Google Scholar] [CrossRef]
- Albalushi, H.; Kurek, M.; Karlsson, L.; Landreh, L.; Kjartansdottir, K.R.; Soder, O.; Hovatta, O.; Stukenborg, J.B. Laminin 521 Stabilizes the Pluripotency Expression Pattern of Human Embryonic Stem Cells Initially Derived on Feeder Cells. Stem Cells Int. 2018, 2018, 7127042. [Google Scholar] [CrossRef]
- Hongisto, H.; Ilmarinen, T.; Vattulainen, M.; Mikhailova, A.; Skottman, H. Xeno- and feeder-free differentiation of human pluripotent stem cells to two distinct ocular epithelial cell types using simple modifications of one method. Stem Cell Res. Ther. 2017, 8, 291. [Google Scholar] [CrossRef]
- Lambshead, J.W.; Meagher, L.; Goodwin, J.; Labonne, T.; Ng, E.; Elefanty, A.; Stanley, E.; O’Brien, C.M.; Laslett, A.L. Long-Term Maintenance of Human Pluripotent Stem Cells on cRGDfK-Presenting Synthetic Surfaces. Sci. Rep. 2018, 8, 701. [Google Scholar] [CrossRef]
- Shimizu, E.; Iguchi, H.; Le, M.N.T.; Nakamura, Y.; Kobayashi, D.; Arai, Y.; Takakura, K.; Benno, S.; Yoshida, N.; Tsukahara, M.; et al. A chemically-defined plastic scaffold for the xeno-free production of human pluripotent stem cells. Sci. Rep. 2022, 12, 2516. [Google Scholar] [CrossRef]
- Leino, M.; Astrand, C.; Hughes-Brittain, N.; Robb, B.; McKean, R.; Chotteau, V. Human embryonic stem cell dispersion in electrospun PCL fiber scaffolds by coating with laminin-521 and E-cadherin-Fc. J. Biomed. Mater. Res. B Appl. Biomater. 2018, 106, 1226–1236. [Google Scholar] [CrossRef]
- Rivera-Ordaz, A.; Peli, V.; Manzini, P.; Barilani, M.; Lazzari, L. Critical Analysis of cGMP Large-Scale Expansion Process in Bioreactors of Human Induced Pluripotent Stem Cells in the Framework of Quality by Design. BioDrugs 2021, 35, 693–714. [Google Scholar] [CrossRef]
- Vitillo, L.; Kimber, S.J. Integrin and FAK Regulation of Human Pluripotent Stem Cells. Curr. Stem Cell Rep. 2017, 3, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Rowland, T.J.; Miller, L.M.; Blaschke, A.J.; Doss, E.L.; Bonham, A.J.; Hikita, S.T.; Johnson, L.V.; Clegg, D.O. Roles of integrins in human induced pluripotent stem cell growth on Matrigel and vitronectin. Stem Cells Dev. 2010, 19, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Rodin, S.; Domogatskaya, A.; Strom, S.; Hansson, E.M.; Chien, K.R.; Inzunza, J.; Hovatta, O.; Tryggvason, K. Long-term self-renewal of human pluripotent stem cells on human recombinant laminin-511. Nat. Biotechnol. 2010, 28, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Marti, M.; Mulero, L.; Pardo, C.; Morera, C.; Carrio, M.; Laricchia-Robbio, L.; Esteban, C.R.; Izpisua Belmonte, J.C. Characterization of pluripotent stem cells. Nat. Protoc. 2013, 8, 223–253. [Google Scholar] [CrossRef]
- Huang, Y.; Liang, P.; Liu, D.; Huang, J.; Songyang, Z. Telomere regulation in pluripotent stem cells. Protein Cell 2014, 5, 194–202. [Google Scholar] [CrossRef]
- Rivera, T.; Haggblom, C.; Cosconati, S.; Karlseder, J. A balance between elongation and trimming regulates telomere stability in stem cells. Nat. Struct. Mol. Biol. 2017, 24, 30–39. [Google Scholar] [CrossRef]
- Boyle, J.M.; Hennick, K.M.; Regalado, S.G.; Vogan, J.M.; Zhang, X.; Collins, K.; Hockemeyer, D. Telomere length set point regulation in human pluripotent stem cells critically depends on the shelterin protein TPP1. Mol. Biol. Cell 2020, 31, 2583–2596. [Google Scholar] [CrossRef]
- Yap, L.; Tay, H.G.; Nguyen, M.T.X.; Tjin, M.S.; Tryggvason, K. Laminins in cellular differentiation. Trends Cell Biol. 2019, 29, 987–1000. [Google Scholar] [CrossRef]
- Nascimento, M.A.; Sorokin, L.; Coelho-Sampaio, T. Fractone bulbs derive from ependymal cells and their laminin composition influence the stem cell niche in the subventricular zone. J. Neurosci. 2018, 38, 3880–3889. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesquita, F.C.P.; Leite, E.S.; Morrissey, J.; Freitas, C.; Coelho-Sampaio, T.; Hochman-Mendez, C. Polymerized Laminin-521: A Feasible Substrate for Expanding Induced Pluripotent Stem Cells at a Low Protein Concentration. Cells 2022, 11, 3955. https://doi.org/10.3390/cells11243955
Mesquita FCP, Leite ES, Morrissey J, Freitas C, Coelho-Sampaio T, Hochman-Mendez C. Polymerized Laminin-521: A Feasible Substrate for Expanding Induced Pluripotent Stem Cells at a Low Protein Concentration. Cells. 2022; 11(24):3955. https://doi.org/10.3390/cells11243955
Chicago/Turabian StyleMesquita, Fernanda C. P., Eliel S. Leite, Jacquelynn Morrissey, Catarina Freitas, Tatiana Coelho-Sampaio, and Camila Hochman-Mendez. 2022. "Polymerized Laminin-521: A Feasible Substrate for Expanding Induced Pluripotent Stem Cells at a Low Protein Concentration" Cells 11, no. 24: 3955. https://doi.org/10.3390/cells11243955
APA StyleMesquita, F. C. P., Leite, E. S., Morrissey, J., Freitas, C., Coelho-Sampaio, T., & Hochman-Mendez, C. (2022). Polymerized Laminin-521: A Feasible Substrate for Expanding Induced Pluripotent Stem Cells at a Low Protein Concentration. Cells, 11(24), 3955. https://doi.org/10.3390/cells11243955