
Insulitis in Human Type 1 Diabetic Pancreas: From Stem Cell Grafting to Islet Organoids for a Successful Cell-Based Therapy

 ,
,  , and
, and 
{kind=link}
Abstract
1. Introduction
2. Insulitis Characterisation
3. Insulitis in Human Type 1 Diabetes
4. Attempts to Counteract or Prevent Insulitis
5. Stem Cell Grafting
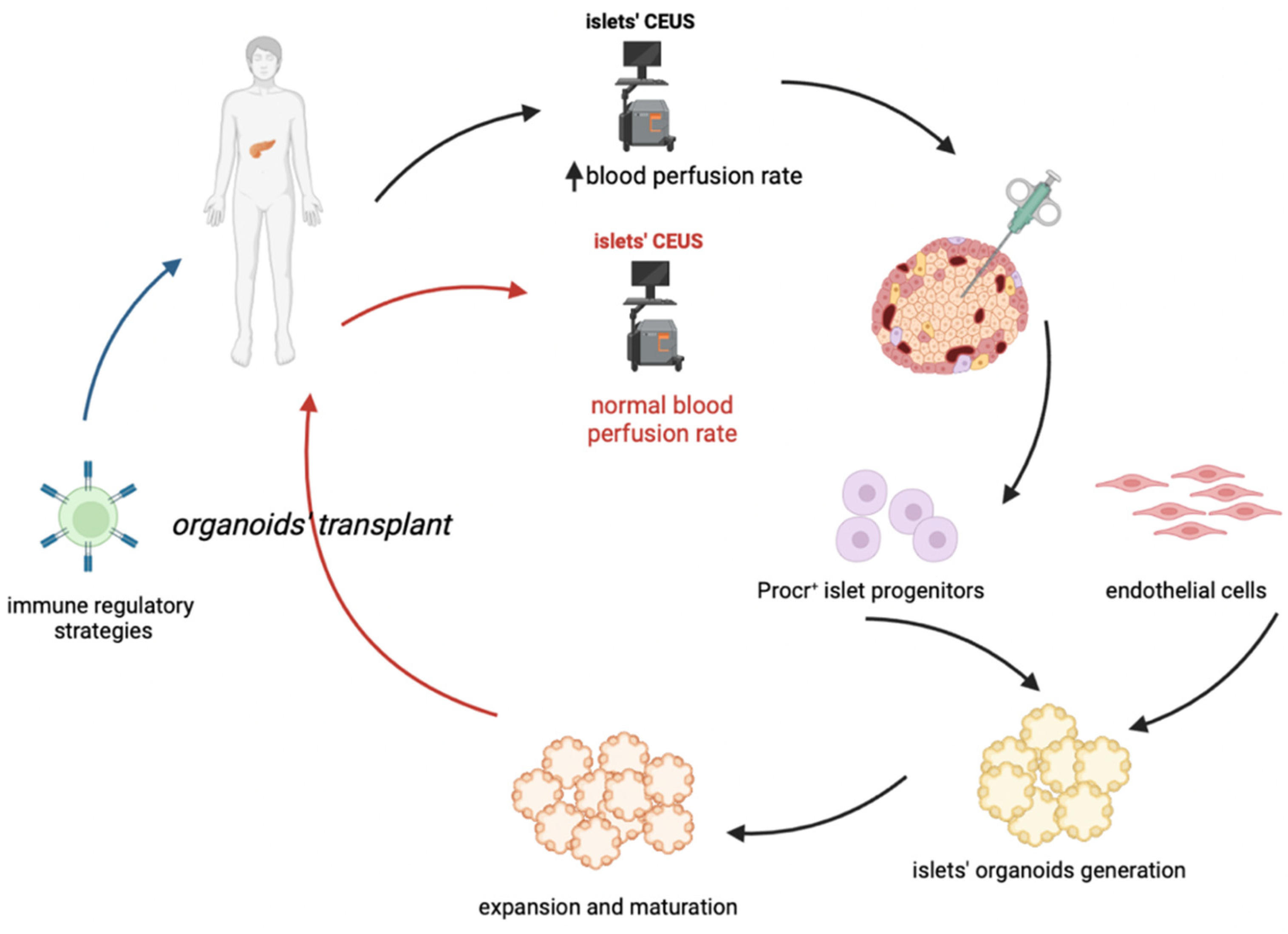
6. Cell Therapy and the Organoid Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pugliese, A. Advances in the etiology and mechanisms of type 1 diabetes. Discov. Med. 2014, 18, 141–150. [Google Scholar] [PubMed]
- Zanone, M.M.; Favaro, E.; Camussi, G. From endothelial to beta cells: Insights into pancreatic islet microendothelium. Curr. Diabetes Rev. 2008, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- St Clair, J.R.; Ramirez, D.; Passman, S.; Benninger, R.K.P. Contrast-enhanced ultrasound measurement of pancreatic blood flow dynamics predicts type 1 diabetes progression in preclinical models. Nat. Commun. 2018, 9, 1742. [Google Scholar] [CrossRef]
- Mizuno, A.; Noma, Y.; Kuwajima, M.; Murakami, T.; Zhu, M.; Shima, K. Changes in islet capillary angioarchitecture coincide with impaired B-cell function but not with insulin resistance in male Otsuka-Long-Evans-Tokushima fatty rats: Dimorphism of the diabetic phenotype at an advanced age. Metabolism 1999, 48, 477–483. [Google Scholar] [CrossRef]
- Papaccio, G. Insulitis and islet microvasculature in type 1 diabetes. Histol. Histopathol. 1993, 8, 751–759. [Google Scholar] [PubMed]
- Papaccio, G.; Chieffi-Baccari, G.; Mezzogiorno, V.; Esposito, V. Capillary area in early low-dose streptozocin-treated mice. Histochemistry 1990, 95, 19–21. [Google Scholar] [CrossRef]
- Papaccio, G.; Latronico, M. Diabetes incidence and histopathological lesions in animal models. Diabetes Res. Clin. Pract. 1992, 18, 137. [Google Scholar] [CrossRef] [PubMed]
- Papaccio, G.; Pisanti, F.A.; Frascatore, S. Acetyl-Homocysteine-Thiolactone-Induced Increase of Superoxide Dismutase Counteracts the Effect of Subdiabetogenic Doses of Streptozocin. Diabetes 1986, 35, 470–474. [Google Scholar] [CrossRef]
- Papaccio, G.; Pisanti, F.A.; Di Montefiano, R.; Graziano, A.; Latronico, M.V. Th1 and Th2 cytokines exert regulatory effects upon islet microvascular areas in the NOD mouse. J. Cell. Biochem. 2002, 86, 651–664. [Google Scholar] [CrossRef]
- Gepts, W. Pathologic Anatomy of the Pancreas in Juvenile Diabetes Mellitus. Diabetes 1965, 14, 619–633. [Google Scholar] [CrossRef]
- Foulis, A.K.; Liddle, C.N.; Farquharson, M.A.; Richmond, J.A.; Weir, R.S. The histopathology of the pancreas in Type I (insulin-dependent) diabetes mellitus: A 25-year review of deaths in patients under 20 years of age in the United Kingdom. Diabetologia 1986, 29, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Krogvold, L.; Edwin, B.; Buanes, T.; Ludvigsson, J.; Korsgren, O.; Hyöty, H.; Frisk, G.; Hanssen, K.F.; Dahl-Jørgensen, K. Pancreatic biopsy by minimal tail resection in live adult patients at the onset of type 1 diabetes: Experiences from the DiViD study. Diabetologia 2014, 57, 841–843. [Google Scholar] [CrossRef]
- Imagawa, A.; Hanafusa, T.; Tamura, S.; Moriwaki, M.; Itoh, N.; Yamamoto, K.; Iwahashi, H.; Yamagata, K.; Waguri, M.; Nanmo, T.; et al. Pancreatic Biopsy as a Procedure for Detecting In Situ Autoimmune Phenomena in Type 1 Diabetes. Diabetes 2001, 50, 1269–1273. [Google Scholar] [CrossRef]
- Campbell-Thompson, M.; Fu, A.; Kaddis, J.S.; Wasserfall, C.; Schatz, D.A.; Pugliese, A.; Atkinson, M.A. Insulitis and β-Cell Mass in the Natural History of Type 1 Diabetes. Diabetes 2016, 65, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Thompson, M.L.; Atkinson, M.A.; Butler, A.E.; Chapman, N.M.; Frisk, G.; Gianani, R.; Giepmans, B.N.; von Herrath, M.G.; Hyöty, H.; Kay, T.W.; et al. The diagnosis of insulitis in human type 1 diabetes. Diabetologia 2013, 56, 2541–2543. [Google Scholar] [CrossRef]
- Krogvold, L.; Wiberg, A.; Edwin, B.; Buanes, T.; Jahnsen, F.L.; Hanssen, K.F.; Larsson, E.; Korsgren, O.; Skog, O.; Dahl-Jørgensen, K. Insulitis and characterisation of infiltrating T cells in surgical pancreatic tail resections from patients at onset of type 1 diabetes. Diabetologia 2016, 59, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Zeng, N.; Al-Diery, H.; Jung, D.; Yeu, C.; Joret, M.O.; Merrilees, M.J.; Wu, F. Analysis of peri-islet CD45-positive leucocytic infiltrates in long-standing type 1 diabetic patients. Diabetologia 2015, 58, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Foulis, A.K.; Farquharson, M.A.; Hardman, R. Aberrant expression of Class II major histocompatibility complex molecules by B cells and hyperexpression of Class I major histocompatibility complex molecules by insulin containing islets in Type 1 (insulin-dependent) diabetes mellitus. Diabetologia 1987, 30, 333–343. [Google Scholar] [CrossRef]
- Foulis, A.K.; Farquharson, M.A. Aberrant Expression of HLA-DR Antigens by Insulin-Containing β-Cells in Recent-Onset Type I Diabetes MelIitus. Diabetes 1986, 35, 1215–1224. [Google Scholar] [CrossRef]
- Bottazzo, G.F.; Dean, B.M.; McNally, J.M.; MacKay, E.H.; Swift, P.G.F.; Gamble, D.R. In Situ Characterization of Autoimmune Phenomena and Expression of HLA Molecules in the Pancreas in Diabetic Insulitis. N. Engl. J. Med. 1985, 313, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.S.; Bluestone, J.A. The NOD mouse: A model of immune dysregulation. Annu. Rev. Immunol. 2005, 23, 447–485. [Google Scholar] [CrossRef]
- Lee, Y.; Chin, R.K.; Christiansen, P.; Sun, Y.; Tumanov, A.V.; Wang, J.; Chervonsky, A.V.; Fu, Y.X. Recruitment and activation of naive T cells in the islets by lymphotoxin beta receptor-dependent tertiary lymphoid structure. Immunity 2006, 25, 499–509. [Google Scholar] [CrossRef]
- Magnuson, A.M.; Thurber, G.M.; Kohler, R.H.; Weissleder, R.; Mathis, D.; Benoist, C. Population dynamics of islet-infiltrating cells in autoimmune diabetes. Proc. Natl. Acad. Sci. USA 2015, 112, 1511–1516. [Google Scholar] [CrossRef]
- Willcox, A.; Richardson, S.J.; Bone, A.J.; Foulis, A.K.; Morgan, N.G. Analysis of islet inflammation in human type 1 diabetes. Clin. Exp. Immunol. 2009, 155, 173–181. [Google Scholar] [CrossRef]
- Coppieters, K.T.; Dotta, F.; Amirian, N.; Campbell, P.D.; Kay, T.W.; Atkinson, M.A.; Roep, B.O.; von Herrath, M.G. Demonstration of islet-autoreactive CD8 T cells in insulitic lesions from recent onset and long-term type 1 diabetes patients. J. Exp. Med. 2012, 209, 51–60. [Google Scholar] [CrossRef]
- Arif, S.; Leete, P.; Nguyen, V.; Marks, K.; Nor, N.M.; Estorninho, M.; Kronenberg-Versteeg, D.; Bingley, P.J.; Todd, J.A.; Guy, C.; et al. Blood and islet phenotypes indicate immunological heterogeneity in type 1 diabetes. Diabetes 2014, 63, 3835–3845. [Google Scholar] [CrossRef]
- Leete, P.; Morgan, N.G. Footprints of Immune Cells in the Pancreas in Type 1 Diabetes; to “B” or Not to “B”: Is That Still the Question? Front. Endocrinol. (Lausanne) 2021, 25, 617437. [Google Scholar] [CrossRef]
- Leete, P.; Willcox, A.; Krogvold, L.; Dahl-Jørgensen, K.; Foulis, A.K.; Richardson, S.J.; Morgan, N.G. Differential Insulitic Profiles Determine the Extent of β-Cell Destruction and the Age at Onset of Type 1 Diabetes. Diabetes 2016, 65, 1362–1369. [Google Scholar] [CrossRef]
- Bogdani, M.; Johnson, P.Y.; Potter-Perigo, S.; Nagy, N.; Day, A.J.; Bollyky, P.L.; Wight, T.N. Hyaluronan and hyaluronan-binding proteins accumulate in both human type 1 diabetic islets and lymphoid tissues and associate with inflammatory cells in insulitis. Diabetes 2014, 63, 2727–2743. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, H.F.; Rieck, M.; Gurevich, I.; Nagy, N.; Butte, M.J.; Negrin, R.S.; Wight, T.N.; Steinman, L.; Bollyky, P.L. Hyaluronan synthesis is necessary for autoreactive T-cell trafficking, activation, and Th1 polarization. Proc. Natl. Acad. Sci. USA 2016, 113, 1339–1344. [Google Scholar] [CrossRef]
- Bogdani, M.; Korpos, E.; Simeonovic, C.J.; Parish, C.R.; Sorokin, L.; Wight, T.N. Extracellular Matrix Components in the Pathogenesis of Type 1 Diabetes. Curr. Diab. Rep. 2014, 14, 552. [Google Scholar] [CrossRef]
- Krogvold, L.; Skog, O.; Sundström, G.; Edwin, B.; Buanes, T.; Hanssen, K.F.; Ludvigsson, J.; Grabherr, M.; Korsgren, O.; Dahl-Jørgensen, K. Function of Isolated Pancreatic Islets From Patients at Onset of Type 1 Diabetes: Insulin Secretion Can Be Restored After Some Days in a Nondiabetogenic Environment In Vitro. Diabetes 2015, 64, 2506–2512. [Google Scholar] [CrossRef]
- Keenan, H.A.; Sun, J.K.; Levine, J.; Doria, A.; Aiello, L.P.; Eisenbarth, G.; Bonner-Weir, S.; King, G.L. Residual Insulin Production and Pancreatic β-Cell Turnover After 50 Years of Diabetes: Joslin Medalist Study. Diabetes 2010, 59, 2846–2853. [Google Scholar] [CrossRef]
- Coppieters, K.T.; Wiberg, A.; Amirian, N.; Kay, T.W.; von Herrath, M.G. Persistent glucose transporter expression on pancreatic beta cells from longstanding type 1 diabetic individuals. Diabetes Metab. Res. Rev. 2011, 27, 746–754. [Google Scholar] [CrossRef]
- Gianani, R.; Campbell-Thompson, M.; Sarkar, S.A.; Wasserfall, C.; Pugliese, A.; Solis, J.M.; Kent, S.C.; Hering, B.J.; West, E.; Steck, A.; et al. Dimorphic histopathology of long-standing childhood-onset diabetes. Diabetologia 2010, 53, 690–698. [Google Scholar] [CrossRef] [PubMed]
- Morgan, N.G.; Leete, P.; Foulis, A.K.; Richardson, S.J. Islet inflammation in human type 1 diabetes mellitus. IUBMB Life 2014, 66, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, A.; Matsumura, K.; Kikuno, S.; Nagasawa, K.; Okubo, M.; Mori, Y.; Kobayashi, T. Slowly Progressive Type 1 Diabetes Mellitus: Current Knowledge And Future Perspectives. Diabetes Metab. Syndr. Obes. 2019, 12, 2461–2477. [Google Scholar] [CrossRef]
- Muralidharan, C.; Linnemann, A.K. β-Cell autophagy in the pathogenesis of type 1 diabetes. Am. J. Physiol. Metab. 2021, 321, E410–E416. [Google Scholar] [CrossRef] [PubMed]
- Like, A.A.; Rossini, A.A. Streptozotocin-Induced Pancreatic Insulitis: New Model of Diabetes Mellitus. Science 1976, 193, 415–417. [Google Scholar] [CrossRef]
- Hayakawa, M.; Yokono, K.; Nagata, M.; Hatamori, N.; Ogawa, W.; Miki, A.; Mizoguti, H.; Baba, S. Morphological Analysis of Selective Destruction of Pancreatic β-cells by Cytotoxic T Lymphocytes in NOD Mice. Diabetes 1991, 40, 1210–1217. [Google Scholar] [CrossRef]
- Stiller, C.R.; Dupré, J.; Gent, M.; Jenner, M.R.; Keown, P.A.; Laupacis, A.; Martell, R.; Rodger, N.W.; von Graffenried, B.; Wolfe, B.M. Effects of cyclosporine immunosuppression in insulin-dependent diabetes mellitus of recent onset. Science 1984, 223, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- Parving, H.H.; Tarnow, L.; Nielsen, F.S.; Rossing, P.; Mandrup-Poulsen, T.; Osterby, R.; Nerup, J. Cyclosporine nephrotoxicity in type 1 diabetic patients. A 7-year follow-up study. Diabetes Care. 1999, 22, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.J.; Hudson, I.; Harrison, L.C.; Dean, B.; Colman, P.G.; Werther, G.A.; Warne, G.L.; Court, J.M. Double-blind controlled trial of azathioprine in children with newly diagnosed type I diabetes. Diabetes 1989, 38, 779–783. [Google Scholar] [CrossRef]
- Böhmer, K.P.; Kolb, H.; Kuglin, B.; Zielasek, J.; Hübinger, A.; Lampeter, E.F.; Weber, B.; Kolb-Bachofen, V.; Jastram, H.U.; Bertrams, J.; et al. Linear loss of insulin secretory capacity during the last six months preceding IDDM. No effect of antiedematous therapy with ketotifen. Diabetes Care 1994, 17, 138–141. [Google Scholar] [CrossRef]
- Lampeter, E.F.; Klinghammer, A.; Scherbaum, W.A.; Heinze, E.; Haastert, B.; Giani, G.; Kolb, H. The Deutsche Nicotinamide Intervention Study: An attempt to prevent type 1 diabetes. DENIS Group. Diabetes 1998, 47, 980–984. [Google Scholar] [CrossRef]
- Eisenbarth, G.S.; Srikanta, S.; Jackson, R.; Rabinowe, S.; Dolinar, R.; Aoki, T.; Morris, M.A. Anti-thymocyte globulin and prednisone immunotherapy of recent onset type 1 diabetes mellitus. Diabetes Res. 1985, 2, 271–276. [Google Scholar]
- Keymeulen, B.; Vandemeulebroucke, E.; Ziegler, A.G.; Mathieu, C.; Kaufman, L.; Hale, G.; Gorus, F.; Goldman, M.; Walter, M.; Candon, S.; et al. Insulin needs after CD3-antibody therapy in new-onset type 1 diabetes. N. Engl. J. Med. 2005, 352, 2598–2608. [Google Scholar] [CrossRef]
- Keymeulen, B.; Walter, M.; Mathieu, C.; Kaufman, L.; Gorus, F.; Hilbrands, R.; Vandemeulebroucke, E.; Van de Velde, U.; Crenier, L.; De Block, C.; et al. Four-year metabolic outcome of a randomised controlled CD3-antibody trial in recent-onset type 1 diabetic patients depends on their age and baseline residual beta cell mass. Diabetologia 2010, 53, 614–623. [Google Scholar] [CrossRef]
- Herold, K.C.; Gitelman, S.; Greenbaum, C.; Puck, J.; Hagopian, W.; Gottlieb, P.; Sayre, P.; Bianchine, P.; Wong, E.; Seyfert-Margolis, V.; et al. Immune Tolerance Network ITN007AI Study Group. Treatment of patients with new onset Type 1 diabetes with a single course of anti-CD3 mAb Teplizumab preserves insulin production for up to 5 years. Clin. Immunol. 2009, 132, 166–173. [Google Scholar] [CrossRef]
- Lernmark, A.; Larsson, H.E. Immune therapy in type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2013, 9, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Rydén, A.K.; Wesley, J.D.; Coppieters, K.T.; Von Herrath, M.G. Non-antigenic and antigenic interventions in type 1 diabetes. Hum. Vaccin. Immunother. 2014, 10, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Pescovitz, M.D.; Greenbaum, C.J.; Krause-Steinrauf, H.; Becker, D.J.; Gitelman, S.E.; Goland, R.; Gottlieb, P.A.; Marks, J.B.; McGee, P.F.; Moran, A.M.; et al. Type 1 Diabetes TrialNet Anti-CD20 Study Group. Rituximab, B-lymphocyte depletion, and preservation of beta-cell function. N. Engl. J. Med. 2009, 361, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Pescovitz, M.D.; Greenbaum, C.J.; Bundy, B.; Becker, D.J.; Gitelman, S.E.; Goland, R.; Gottlieb, P.A.; Marks, J.B.; Moran, A.; Raskin, P.; et al. Type 1 Diabetes TrialNet Anti-CD20 Study Group. B-lymphocyte depletion with rituximab and β-cell function: Two-year results. Diabetes Care 2014, 37, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Orban, T.; Bundy, B.; Becker, D.J.; DiMeglio, L.A.; Gitelman, S.E.; Goland, R.; Gottlieb, P.A.; Greenbaum, C.J.; Marks, J.B.; Monzavi, R.; et al. Type 1 Diabetes TrialNet Abatacept Study Group. Co-stimulation modulation with abatacept in patients with recent-onset type 1 diabetes: A randomised, double-blind, placebo-controlled trial. Lancet 2011, 378, 412–419. [Google Scholar] [CrossRef]
- Achenbach, P.; Warncke, K.; Reiter, J.; Naserke, H.E.; Williams, A.J.; Bingley, P.J.; Bonifacio, E.; Ziegler, A.G. Stratification of type 1 diabetes risk on the basis of islet autoantibody characteristics. Diabetes 2004, 53, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.; Weigmann, B.; Bronson, R.; von Boehmer, H. Prevention of type 1 diabetes in mice by tolerogenic vaccination with a strong agonist insulin mimetope. J. Exp. Med. 2011, 208, 1501–1510. [Google Scholar] [CrossRef]
- Diabetes Prevention Trial--Type 1 Diabetes Study Group. Effects of insulin in relatives of patients with type 1 diabetes mellitus. N. Engl. J. Med. 2002, 346, 1685–1691. [Google Scholar] [CrossRef]
- Skyler, J.S.; Krischer, J.P.; Wolfsdorf, J.; Cowie, C.; Palmer, J.P.; Greenbaum, C.; Cuthbertson, D.; Rafkin-Mervis, L.E.; Chase, H.P.; Leschek, E. Effects of oral insulin in relatives of patients with type 1 diabetes: The Diabetes Prevention Trial--Type 1. Diabetes Care 2005, 28, 1068–1076. [Google Scholar] [CrossRef]
- Elias, D.; Reshef, T.; Birk, O.S.; van der Zee, R.; Walker, M.D.; Cohen, I.R. Vaccination against autoimmune mouse diabetes with a T-cell epitope of the human 65-kDa heat shock protein. Proc. Natl. Acad. Sci. USA 1991, 15, 3088–3091. [Google Scholar] [CrossRef]
- Fischer, B.; Elias, D.; Bretzel, R.G.; Linn, T. Immunomodulation with heat shock protein DiaPep277 to preserve beta cell function in type 1 diabetes—An update. Expert Opin. Biol. Ther. 2010, 10, 265–272. [Google Scholar] [CrossRef]
- Bogdani, M.; Speake, C.; Dufort, M.J.; Johnson, P.Y.; Larmore, M.J.; Day, A.J.; Wight, T.N.; Lernmark, Å.; Greenbaum, C.J. Hyaluronan deposition in islets may precede and direct the location of islet immune-cell infiltrates. Diabetologia 2020, 63, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Hull, R.L.; Bogdani, M.; Nagy, N.; Johnson, P.Y.; Wight, T.N. Hyaluronan: A Mediator of Islet Dysfunction and Destruction in Diabetes? J. Histochem. Cytochem. 2015, 63, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.X.; Zhang, D.Y.; Khan, M.A.; Zheng, S.Y.; Hu, X.M.; Zhang, Q.; Yang, R.H.; Xiong, K. Stem Cell Transplantation in the Treatment of Type 1 Diabetes Mellitus: From Insulin Replacement to Beta-Cell Replacement. Front. Endocrinol. (Lausanne) 2022, 13, 859638. [Google Scholar] [CrossRef] [PubMed]
- Pixley, J.S. Mesenchymal stem cells to treat type 1 diabetes. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165315. [Google Scholar] [CrossRef] [PubMed]
- Cuesta-Gomez, N.; Graham, G.J.; Campbell, J.D.M. Chemokines and their receptors: Predictors of the therapeutic potential of mesenchymal stromal cells. J. Transl. Med. 2021, 19, 156. [Google Scholar] [CrossRef]
- Burke, G.W.; Vendrame, F.; Virdi, S.K.; Ciancio, G.; Chen, L.; Ruiz, P.; Messinger, S.; Reijonen, H.K.; Pugliese, A. Lessons From Pancreas Transplantation in Type 1 Diabetes: Recurrence of Islet Autoimmunity. Curr. Diab. Rep. 2015, 15, 121. [Google Scholar] [CrossRef]
- Chhabra, P.; Brayman, K.L. Stem cell therapy to cure type 1 diabetes: From hype to hope. Stem Cells Transl. Med. 2013, 2, 328–336. [Google Scholar] [CrossRef]
- Wu, S.; Wang, L.; Fang, Y.; Huang, H.; You, X.; Wu, J. Advances in Encapsulation and Delivery Strategies for Islet Transplantation. Adv. Healthc. Mater. 2021, 10, e2100965. [Google Scholar] [CrossRef]
- Yang, S.J.; Singh, A.K.; Drow, T.; Tappen, T.; Honaker, Y.; Barahmand-Pour-Whitman, F.; Linsley, P.S.; Cerosaletti, K.; Mauk, K.; Xiang, Y.; et al. Pancreatic islet-specific engineered Tregs exhibit robust antigen-specific and bystander immune suppression in type 1 diabetes models. Sci. Transl. Med. 2022, 14, eabn1716. [Google Scholar] [CrossRef]
- Marfil-Garza, B.A.; Imes, S.; Verhoeff, K.; Hefler, J.; Lam, A.; Dajani, K.; Anderson, B.; O’Gorman, D.; Kin, T.; Bigam, D.; et al. Pancreatic islet transplantation in type 1 diabetes: 20-year experience from a single-centre cohort in Canada. Lancet Diabetes Endocrinol. 2022, 10, 519–532. [Google Scholar] [CrossRef]
- Loretelli, C.; Assi, E.; Seelam, A.J.; Ben Nasr, M.; Fiorina, P. Cell therapy for type 1 diabetes. Expert Opin. Biol. Ther. 2020, 20, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Hwang, G.; Jeong, H.; Yang, H.K.; Kim, H.S.; Hong, H.; Kim, N.J.; Oh, I.H.; Yim, H.W. Efficacies of Stem Cell Therapies for Functional Improvement of the β Cell in Patients with Diabetes: A Systematic Review of Controlled Clinical Trials. Int. J. Stem Cells 2019, 12, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Cuesta-Gomez, N.; Verhoeff, K.; Jasra, I.T.; Pawlick, R.; Dadheech, N.; Shapiro, A.M.J. Characterization of stem-cell-derived islets during differentiation and after implantation. Cell Rep. 2022, 40, 111238. [Google Scholar] [CrossRef] [PubMed]
- Verhoeff, K.; Cuesta-Gomez, N.; Jasra, I.; Marfil-Garza, B.; Dadheech, N.; Shapiro, A.M.J. Optimizing Generation of Stem Cell-Derived Islet Cells. Stem Cell Rev. Rep. 2022, 18, 2683–2698. [Google Scholar] [CrossRef]
- Dolgin, E. Diabetes cell therapies take evasive action. Nat. Biotechnol. 2022, 40, 291–295. [Google Scholar] [CrossRef]
- From the American Association of Neurological Surgeons (AANS); American Society of Neuroradiology (ASNR); Cardiovascular and Interventional Radiology Society of Europe (CIRSE); Canadian Interventional Radiology Association (CIRA); Congress of Neurological Surgeons (CNS); European Society of Minimally Invasive Neurological Therapy (ESMINT); European Society of Neuroradiology (ESNR); European Stroke Organization (ESO); Society for Cardiovascular Angiography and Interventions (SCAI); Society of Interventional Radiology (SIR); et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef]
- Chellappan, D.K.; Sivam, N.S.; Teoh, K.X.; Leong, W.P.; Fui, T.Z.; Chooi, K.; Khoo, N.; Yi, F.J.; Chellian, J.; Cheng, L.L.; et al. Gene therapy and type 1 diabetes mellitus. Biomed. Pharmacother. 2018, 108, 1188–1200. [Google Scholar] [CrossRef]
- Wang, D.; Wang, J.; Bai, L.; Pan, H.; Feng, H.; Clevers, H.; Zeng, Y.A. Long-Term Expansion of Pancreatic Islet Organoids from Resident Procr+ Progenitors. Cell 2020, 180, 1198–1211. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
La Noce, M.; Nicoletti, G.F.; Papaccio, G.; Del Vecchio, V.; Papaccio, F. Insulitis in Human Type 1 Diabetic Pancreas: From Stem Cell Grafting to Islet Organoids for a Successful Cell-Based Therapy. Cells 2022, 11, 3941. https://doi.org/10.3390/cells11233941
La Noce M, Nicoletti GF, Papaccio G, Del Vecchio V, Papaccio F. Insulitis in Human Type 1 Diabetic Pancreas: From Stem Cell Grafting to Islet Organoids for a Successful Cell-Based Therapy. Cells. 2022; 11(23):3941. https://doi.org/10.3390/cells11233941
Chicago/Turabian StyleLa Noce, Marcella, Giovanni Francesco Nicoletti, Gianpaolo Papaccio, Vitale Del Vecchio, and Federica Papaccio. 2022. "Insulitis in Human Type 1 Diabetic Pancreas: From Stem Cell Grafting to Islet Organoids for a Successful Cell-Based Therapy" Cells 11, no. 23: 3941. https://doi.org/10.3390/cells11233941
APA StyleLa Noce, M., Nicoletti, G. F., Papaccio, G., Del Vecchio, V., & Papaccio, F. (2022). Insulitis in Human Type 1 Diabetic Pancreas: From Stem Cell Grafting to Islet Organoids for a Successful Cell-Based Therapy. Cells, 11(23), 3941. https://doi.org/10.3390/cells11233941