1. Introduction
Although it is known that stress can influence female reproduction, how stress affects embryo development is not very clear. Because there are reports that the early stages of pregnancy are more sensitive to stress than the later stages are [
1,
2], the preimplantation period is envisioned as one of the most stress-vulnerable phases [
3,
4]. However, reports on the effects of female stress on preimplantation embryos are very limited. Furthermore, although limited studies have demonstrated that restraint stress on pregnant mice significantly impaired preimplantation embryo development [
5,
6], how the stress effect is delivered to the preimplantation embryo is rarely reported [
7,
8]. In addition, whether the female stress would affect fertilization within the oviduct is not known.
Stresses are commonly associated with the enhanced activity of the hypothalamo-pituitary-adrenal (HPA) axis. The activation of the HPA axis leads to secretion of corticotrophin-releasing hormone (CRH), adrenocorticotropic hormone (ACTH) and glucocorticoids. For instance, restraint of female mice increased their CRH concentration in both serum and ovaries [
9]. An immediate and constant increase in CRH secretion was observed following the transportation of ewes [
10]. A significant elevation in serum corticosterone and cortisol was observed after mice were exposed to various stresses [
11,
12]. Moreover, sows exposed to stresses showed significant increases in cortisol concentrations [
13]. However, the mechanisms by which the HPA hormones influence embryo development remain to be explored.
It is known that reproduction in female pigs is sensitive to various stressors, which can activate the HPA axis and facilitate the secretion of CRH, ACTH and cortisol [
14,
15]. Furthermore, repeated ACTH stimulation [
16] or food deprivation [
17] after ovulation in sows significantly impaired cleavage of the preimplantation embryos while inducing a significant elevation in the level of blood cortisol. However, although these data suggest that stresses might compromise the preimplantation embryo development by facilitating production of the HPA hormones in pigs, the mechanisms by which HPA hormones impair embryo development are largely unclear. Because of the high incidence of polyspermy during pig in vitro fertilization [
18], the effects of oviducts on polyspermic fertilization should be studied in this species to find out the etiology for the pathological condition of polyspermy. Furthermore, although by using an in vitro model, Tan et al. [
8] have observed that CRH and corticosterone affected mouse preimplantation embryo development indirectly by triggering apoptosis of the oviductal epithelial cells (OECs), their conclusions remain to be verified in other species.
Thus, the major objectives of this study were to specify the effects of CRH, ACTH and cortisol on pig embryo development; to clarify whether these hormones would damage pig embryos directly or indirectly by changing the oviductal environment; and to explore whether they would affect fertilization in the oviducts.
2. Methods
The procedures for animal care and handling were performed according to guidelines approved by the Animal Care and Use Committee of the Shandong Agricultural University P. R. China (Permit number: SDAUA-2001-001). Unless pointed out otherwise, all chemicals and reagents used in this study were bought from Sigma-Aldrich Corp. (St. Louis, MO, USA).
2.1. Collection of Ovaries and Oviducts
Pig ovaries and oviducts were obtained from the Feicheng slaughterhouse of Yinbao Food Corporation Ltd. (Taian, China). Only ovaries and oviducts at the follicular stage were collected from gilts, approximately 6 months after birth (approximately 100 kg bodyweight). While the ovaries were transported in a thermos bottle with sterile saline containing 100 IU/mL penicillin and 0.05 mg/mL streptomycin, maintained at 30–35 °C, the oviducts were transported in physiological saline solution, maintained at 4 °C, to the laboratory within 3 h after slaughtering.
2.2. Collection, In Vitro Maturation and Activation of Oocytes
To collect cumulus-oocyte complexes (COCs), 3–6 mm follicles were aspirated using a syringe containing Dulbecco’s phosphate-buffered saline (D-PBS, HyClone, Logan, UT, USA) supplemented with 0.88 mM CaCl2·2H2O, 0.49 mM MgCl2·6H2O, 0.1% polyvinyl alcohol, 0.03 mM phenol red, 50 IU/mL penicillin and 50 µg/mL streptomycin. The COCs recovered were washed three times in D-PBS, and those showing a uniform ooplasm and compact cumulus were selected for in vitro maturation. Approximately 80 ovaries were obtained on each experimental day, and the oocytes recovered from them were pooled and divided into different treatments.
The culture medium used for oocyte maturation was TCM-199 (Gibco, Grand Island, NY, USA) containing 10% porcine follicle fluid, 0.1% PVP, 0.91 mM sodium pyruvate, 3.05 mM glucose, 0.05 IU/mL FSH, 10 ng/mL EGF, 0.05 IU/mL LH, 0.57 mM cysteine, 50 μg/mL streptomycin and 100 IU/mL penicillin. We aspirated porcine follicular fluid from 3–6 mm follicles of the follicular stage ovaries. After centrifugation at 1600× g for 30 min, we collected the supernatants and filtered them sequentially through 0.22 μm syringe filters. The prepared follicular fluid was stored at −20 °C until use. We placed the maturation medium in culture wells of a 96-well plate (150 μL per well), and performed a pre-equilibration at 38.5 °C in an atmosphere of 5% CO2 in humidified air for 3 h before the introduction of the oocytes. We then washed the COCs three times in D-PBS and once in the maturation medium, and then, placed them in the wells (approximately 25 per well) covered with mineral oil before culture for 44 h at 38.5 °C under 5% CO2 in humidified air.
After the maturation culture, the cumulus cells were mechanically removed from oocytes by repeatedly pipetting in D-PBS with 0.1% hyaluronidase using a small-bore pipette. For activation treatment, the cumulus-free oocytes were first cultured for 5 min in D-PBS containing 5 µM ionomycin, and then, they were incubated for 5 h in PZM-3 medium with 2 mM 6-DMAP after being washed three times in PZM-3 medium.
2.3. Preparation of Oviduct Epithelium Cells (OECs) and Conditioned Medium (CM)
One oviduct was used on each experimental day. We dissected the oviducts free from surrounding tissues, ligated them at both ends, and washed them three times in Hanks’ balanced salt solution at 4 °C. After being closed off at one end with a clip, the oviduct was filled with 0.25% trypsin solution and closed off at the other end. Then, the oviduct was incubated at 37 °C for 45 min before being squeezed with tweezers to obtain the contents. We resuspended the recovered pellets in Dulbecco’s Modified Eagles Medium/Ham’s F12 (DMEM/F12, Gibco, Beijing, China) containing 10% (
v/
v) fetal calf serum (Gibco) and 0.5% (
v/
v) penicillin/streptomycin solution (Gibco) and washed them twice by centrifugation (200×
g, 5 min). We added the final suspension (1~2 × 10
5 cells/mL) to wells (500 μL per well) of a 24-well culture plate and cultured them at 38.5 °C in a humidified atmosphere of 5% CO
2 in air. At 24 h after seeding, when most OECs had attached, the medium was renewed for the first time, and then, the medium was renewed every 48 h. Mouse OECs were prepared as reported previously [
8].
Immunohistochemical detection of the cytokeratin filaments was conducted to check the purity of pig OECs we cultured. Pig OECs were cultured on glass coverslips in a 24-well dish. When cells grew to 20% or 90% of confluence, coverslips were rinsed with PBS and fixed in pure ethanol. Cytokeratin-18 were labeled using the cytokeratin-18 rabbit polyclonal antibody (1:100; 18708-1-AP, Proteintech, Chicago, IL, USA) and Cy3-conjugated goat anti-rabbit IgG (1:200; AP132C, Jackson ImmunoResearch, West Grove, PA, USA), and nuclei were stained with Hoechst 33324. Then, the sample was mounted, sealed and visualized under a fluorescent microscope. Each treatment contained three coverslips and three fields were observed on each coverslip. Approximately 200 cells were counted in each field to calculate rates of positive cells. The results showed that the cytokeratin 18-positive rates were 96.4 ± 2.0% and 98.2 ± 1.2% in OECs cultured to 20% and 90% of confluence, respectively.
To prepare CM, when the cultured OECs reached 60–70% of confluence, we replaced the spent DMEM/F12 with fresh serum-free DMEM/F12 without (control) or with CRH, ACTH or cortisol, and we cultured the growing OECs further for 48 h. Then, we replaced the medium with fresh PZM-3 medium without hormones and cultured cells for 24 h. After the culture, we collected the supernatants and centrifuged them for 10 min at 1000× g to collect CM. We froze and stored the CM in aliquots at −20 °C before use.
According to the experimental design, CRH, ACTH and cortisol and their antagonists were added to PZM-3 medium or to serum-free DMEM/F12 to observe their effects on the embryos and OECs, respectively. To prepare stock solutions, we dissolved CRH (10 mM) and ACTH (10 mM) in water, cortisol (10 mM) and RU486 (10 mM) in ethanol and antalarmin (10 mM) in dimethylsulfoxide (DMSO). We stored the stock solutions at −20 °C before use. When the OECs or embryos were treated with cortisol, RU486 or antalarmin, the control OECs/embryos were cultured with the same amount of ethanol or DMSO. The final concentrations of ethanol and DMSO in culture medium were 0.02–0.1% (v/v) and 0.02% (v/v), respectively.
2.4. Embryo Culture
Culture in PZM-3 alone: After the activation treatment, the oocytes were washed in D-PBS to remove 6-DMAP and were cultured in PZM-3 with different concentrations of CRH, ACTH or cortisol. Approximately 25 oocytes were cultured in 200 μL medium covered with paraffin oil in a well of a 96-well plate at 38.5 °C in 5% CO
2 in air [
19]. The medium was renewed every 72 h of culture. Culture wells containing fresh medium were precultured for 6 h under the same temperature and atmosphere conditions for equilibration. Then, embryos were transferred from the old wells to the equilibrated wells.
Coculture with OECs: When OECs grew to 70–80% confluent, the spent DMEM/F12 in wells of the 96-well plate was replaced with 200 μL of PZM-3 with or without CRH, ACTH or cortisol, and the OECs were cultured for 24 h. After the culture, approximately 25 activated oocytes were introduced into each well containing the precultured OECs, covered with mineral oil and cultured for 48 h. Then, the embryos were changed into the new precultured OECs every 48 h.
Culture in CM: Approximately 25 activated oocytes were cultured in 200 μL CM covered with mineral oil in wells of a 96-well plate. Then, the CM was renewed at 72 h intervals.
The cleavage rate was observed at 48 h of embryo culture, and the blastocyst rate was observed at 144 h of embryo culture under a phase contrast microscope. For cell number counting, some of the blastocysts were stained with Hoechst 33342 (10 μg/mL) and observed under a fluorescence microscope. For embryo culture, each treatment was repeated four times on four different experimental days with each replicate containing approximately 25 oocytes.
2.5. In Vitro Fertilization
Modified Tris-buffered medium (mTBM) was used for the fertilization medium, which contained 113.1 mM NaCl, 3.0 mM KCl, 20.0 mM Tris, 11.0 mM glucose, 5.0 mM Na-pyruvate, 7.5 mM CaCl2, 2 mM caffeine and 0.6% BSA. To prepare mTBM-CM, when the cultured OECs grew to 60–70% confluence, they were changed into serum-free DMEM/F12 without (control) or with CRH or cortisol and cultured further for 48 h. Then, the cells were cultured for 24 h in mTBM without hormones. At the end of the culture, we collected supernatants and centrifuged them for 10 min at 1000× g to obtain mTBM-CM. We froze and stored the mTBM-CM in aliquots at −20 °C before use. The freshly prepared mTBM or mTBM-CM was kept in 5% CO2 incubator for 10 h to stabilize its pH at 7.2 to 7.4.
After the maturation culture, we removed cumulus cells as described above, and oocytes showing the first polar bodies were washed three times in mTBM or mTBM-CM. After washing, 15 oocytes were placed in 50 μL mTBM or mTBM-CM under mineral oil in a well of a 96-well plate. The plate was kept in an incubator until spermatozoa were added for fertilization. Freshly-collected semen (1 mL) was washed once in D-PBS and once in mTBM or mTBM-CM by centrifugation at 200× g for 4 min. After the washing procedure, we resuspended the sperm pellets in 1 mL of mTBM or mTBM-CM and incubated them in a CO2 incubator at 38.5 °C for 1 h for capacitation. After capacitation and appropriate dilution with mTBM or mTBM-CM, we added 30 μL of the sperm suspension to 50 μL of the medium containing oocytes, producing a final concentration of 2.5 × 105 sperm/mL. Then, the oocytes and spermatozoa were co-incubated for 6 h at 38.5 °C in an atmosphere of 5% CO2 in air.
After the co-incubation, oocytes were washed twice with pre-equilibrated PZM-3 medium and cultured for 10 h in PZM-3 (15 oocytes/150 μL medium) at 38.5 °C in an atmosphere of 5% CO2 in air. Then, the zona pellucida was removed by treating oocytes with 0.5% pronase for 2 min. The zona-free oocytes were fixed in 4% paraformaldehyde for 2 min and stained with 10 μg/mL Hoechst 33342 for 5 min before being mounted on a slide and examined for fertilization under a fluorescence microscope at a magnification of ×400. Oocytes with one or no pronucleus, oocytes with two pronuclei and those with three or more pronuclei were considered unfertilized, normally fertilized and polyspermic, respectively. Each treatment was repeated three times on three experimental days with each replicate including approximately 15 oocytes.
2.6. Hoechst Staining of OECs
Cultured OECs were stained with Hoechst 33342 in situ in wells of a 24-well culture plate. Briefly, at the end of culture, OECs were washed three times with PBS, and then, stained for 5 min in situ using 10 μg/mL Hoechst in the dark. Then, the stained cells were observed under a Leica DMLB fluorescence microscope at a magnification of 200×. The heterochromatin was heavily stained and was identified by its characteristic bright fluorescence. While the apoptotic cells showed pyknotic nuclei full of heterochromatin, healthy cells showed normal nuclei with sparse heterochromatin spots. Three to five fields were randomly observed in each well, and percentages of apoptotic cells were calculated from 200–250 cells observed in each field. For each experimental series, all images were acquired with identical settings. Cell numbers were counted on raw images by using the Image J software (1.51 K, National Institutes of Health, Bethesda, MD, USA). Briefly, the image was first converted into an 8-bit black and white format (image-Type-8-bit). The brightness threshold was then adjusted to eliminate the background brightness. Then, the threshold value for all the cells and for positive cells were set to 90–255 and 145–255, respectively. Finally, the range of nuclear area detected was set to 100 pixels. Each treatment was repeated three times on three different experimental days with each replicate including cells from one well of a 12-well culture plate.
2.7. Flow Cytometry Assay
The OECs were first stained with Annexin V-FITC/propidium iodide (PI) staining (556547; BD, Franklin Lakes, NJ, USA) and then measured by flow cytometry. Briefly, the OECs were washed in PBS and digested at 37 °C with 0.25% trypsin for 3–5 min. After that, a DMEM/F12 medium containing 10% fetal bovine serum was added to neutralize the residual trypsin. We resuspended the cells of each sample in 100 μL binding buffer, containing 5 μL Annexin V-FITC, and incubated them for 10 min at room temperature in the dark. Finally, we added 5 μL PI to the OECs and analyzed the fluorescence of 10,000 cells per sample using a LSRFortessa (BD). Each treatment was repeated three times on three different experimental days with each replicate including cells from one well of a 24-well culture plate.
2.8. Western Blot Analysis
We first prepared a radio immuno-precipitation assay (RIPA) buffer containing 150 mM NaCl, 1 mM phenylmethyl sulfonyl fluoride, 50 mM Tris (pH 8), 1.0% Triton X-100, 0.5% sodium deoxycholate, and 0.1% SDS. We then washed the OECs in cooled PBS, and lysed them in 100 μL RIPA buffer. After that, the total protein concentration was determined using a BCA Protein Assay Kit (P0012; Beyotime Institute of Biotechnology, Beijing, China) and was adjusted to 1 μg/μL. Then, 20 μL of total protein was placed in a 0.5-mL microfuge tube and frozen at −80 °C before use. To extract proteins, we added 5 μL of 5× sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer to each tube and heated the tubes to 100 °C for 5 min. We ran SDS-PAGE on a 10% polyacrylamide gel. We transferred the proteins electrophoretically onto polyvinylidene fluoride membranes. After that, we washed the membranes in TBST (150 mM NaCl, 2 mM KCl, 25 mM Tris and 0.05% Tween 20; pH 7.4), blocked them with TBST containing 3% BSA at 37 °C for 2 h and incubated them at 4 °C overnight with primary antibodies. We then washed the membranes in TBST, and incubated them for 1.5 h at 37 °C with secondary antibodies. Finally, we detected signals using a BeyoECL Plus kit (P0018S, Beyotime). Each treatment was repeated three times on three different experimental days with each replicate including OECs from one well of a 6-well plate.
The primary antibodies we used are as follows: rabbit anti-activated caspase-3 antibodies (1:1000, ab2302, Abcam Co., Ltd., Beijing, China), goat anti-CRHR1 polyclonal antibodies (1:500, ab59023, Abcam Co., Ltd.), rabbit anti-GR antibodies (1:1000, ab196944, Abcam Co., Ltd.), rabbit anti-Fas (1:500, ab82419, Abcam Co., Ltd.), rabbit anti-TNFR (1:1000, AF8196, Beyotime Co., Ltd., Beijing, China), mouse anti-ACTB monoclonal antibodies (1:1000, CW0096, CWBio Co., Ltd., Beijing, China), and mouse anti-GAPDH monoclonal antibody (1:1000, CW0100, CWBio Co., Ltd.). The secondary antibodies are: HRP-conjugated rabbit anti-goat IgG (1:1000, A0181, Beyotime Co., Ltd.), goat anti-rabbit IgG (1:1000, CW0103, CWBio Co., Ltd.) and goat anti-mouse IgG (1:4000, CW0102, CWBio Co., Ltd.).
2.9. Real-Time PCR and Temperature Gradient PCR for OECs
RNA was isolated by treating the OECs with 1 mL TRIzol reagent, resuspended in diethyl pyrocarbonate-treated MilliQ water (DEPC-dH2O) and digested with RNase-free DNase I (Takara Biotechniques, Dalian, China). Spectroscopical quantification of RNA was performed at 260 nm. The RNA purity and integrity were assessed by determining the A260:A280 ratio (1.8–2.0) and performing electrophoresis in 1% agarose.
We conducted reverse transcription using a total volume of 20 μL and PrimeScript RT Reagent Kit (Takara RR047A): (a) we mixed 2 μL RNA sample, 2 μL 5× gDNA Eraser Buffer, 1 μL gDNA Eraser and 5 μL DEPC-dH
2O in a 0.2-mL reaction tube; (b) we incubated the mixture at 42 °C for 2 min in a PCR instrument (Thermo Scientific, Hudson, NH, USA); (c) we added 4 μL 5× PrimeScript Buffer 2, 1 μL PrimeScript RT Enzyme Mix I, 1 μL RT Primer Mix and 4 μL RNase Free dH
2O to the reaction tube; and (d) we incubated the mixture at 37 °C for 15 min, at 85 °C for 5 s before store at −20 °C until use.
Table 1 shows gene-specific primers for real-time RT-PCR.
A Mx3005P Real-Time PCR System (Stratagene, Valencia, CA, USA) was used to carry out the mRNA quantification, which was performed in a 10-μL reaction volume, containing 1 μL of cDNA, 5 μL of 2× TB Green Premix Ex Taq (Takara, RR420A), 0.2 μL ROX Reference Dye II, 3.4 μL of RNase-free water and 0.2 μL each of forward and reverse gene-specific primers (10 μM, the total, final concentration). The following cycle amplification conditions were used: (a) an initial denaturation step at 95 °C for 5 min; (b) 40 cycles at 95 °C for 5 s; and (c) 59 °C for 34 s for IGF1, 60 °C for 34 s for BDNF, 58 °C for 34 s for TGFB1, 59 °C for 34 s for CRHBP, 63 °C for 34 s for 11-beta hydroxysteroid dehydrogenase 1 (HSD11B1) and HSD11B2. To determine reaction specificity, PCR products were analyzed by sequencing, dissociation-curve analysis and gel electrophoresis. Gene expression was normalized to internal control (GAPDH), and the values were expressed relative to calibrator samples by using the 2−(ΔΔCT) method. For the OECs, each treatment was repeated three times on three different experimental days with each replicate including cells from one well of a 24-well plate, and for embryos, each treatment was repeated three times with each replicate including 250 2- or 4-cell embryos.
We used the Normfinder (
https://www.moma.dk/normfinder-software/, accessed on 30 May 2022), an algorithm-based tool, to identify the most stable genes among the six commonly used reference genes in samples derived from 10 different primary pOECs cultures. The ranking of expression stability calculated by Normfinder in the genes analyzed was
GAPDH =
HMBS >
ACTB >
HPRT1 >
RPL13A >
YWHAZ, and thus, Gapdh could be used as a stable internal reference gene in our experiments.
We conducted the temperature gradient PCR using the GS00482 Multi Block Thermal Cycler (GSTORM, Somerton, UK). Total RNA was extracted from OECs as described above. Approximately 300 2-cell and 4-cell stage embryos or 50 blastocysts were used to isolate total RNA using a RNAqueous-Micro Total RNA Isolation Kit (AM1931, 202Ambion, Austin, TX, USA). Reverse transcription was conducted as mentioned above. Amplification reactions were performed in a 15 μL reaction volume containing 0.6 μL of cDNA, 1.5 μL of 10× PCR buffer, 0.97 μL dNTP, 0.06 μL Taq (Takara, R006A), 11.27 μL RNase-free water and 0.3 μL each of forward and reverse gene-specific primers (10 μM,
Table 1). Amplification started at 95 °C for 3 min followed by 40 cycles at 95 °C for 30 s, gradient temperature 55 °C to 65 °C for 30 s and 72 °C for 30 s; and finally ended with 72 °C for 10 min. We ran the PCR product in 1% agarose gel and isolated desired bands and photographed them under ultraviolet light (Azure c300, Azure biosystems, Dublin, CA, USA). Each treatment was repeated three times with each replicate containing 300 2- or 4-cell embryos or 50 blastocysts.
2.10. One-Step PCR for Blastocysts
To extract RNA, 10 blastocysts were lysed with a commercial cell lysis kit (CellAmp Direct Prep Kit for RT-PCR & Protein Analysis, Takara, 3733Q). The RNA obtained was analyzed by real-time PCR using One Step TB Green PrimeScript PLUS RT-PCR Kit (Takara, RR096A) and primers for Bcl2, Bax and Actb (
Table 1). We used a 10-μL reaction volume for the amplification (0.4 μL each of forward and reverse primers, 2.2 μL RNase Free d H
2O, 1 μL template, 5 μL 2× One Step TB Green RT-PCR Buffer, 0.2 μL PrimeScript PLUS RTase Mix, 0.6 μL Takara Ex Taq HS Mix, and 0.2 μL ROX Reference Dye II). We analyzed the relative gene expression using the 2
−∆∆CT method following normalization against sus scrofa actin beta (
ACTB). We conducted mRNA quantification with the Mx3005P real-time PCR instrument (Stratagene, Valencia, CA, USA). Each treatment was repeated three times with each replicate including 10 blastocysts.
2.11. Enzyme-Linked Immunosorbent Assay (ELISA)
We carried out ELISA for FASL and TNFα in CM using Porcine Factor Related Apoptosis Ligand (FASL) Elisa kit (BlueGene Co., Shanghai, China) and Porcine tumor necrosis factor α (TNF-α) Elisa kit (Mlbio Co., Shanghai, China), respectively. We centrifuged CM for 10 min at 1000× g, and added 100 μL of supernatant or standards in the coated wells. We then added 50 μL of conjugate to each well before incubation for 1 h at 37 °C. After we washed the coated wells with washing solution and dried them with paper towels, we added 50 μL of substrates A and B to each well before incubation for 15 min at 37 °C. Lastly, we terminated the reaction with 50 μL of stop solution, and measured the optical density within 15 min at 450 nm using a plate reader (Infinite50, TECAN, Mannedorf, Switzerland). We calculated the concentrations of FASL or TNFα against respective standard curves. Each treatment was repeated three times on three different experimental days with each replicate containing CM from one well of a 24-well plate.
2.12. Immunofluorescence
Immunofluorescence microscopy was performed to localize CRHR1, glucocorticoid receptor (GR), TNFR and FAS in embryos. We removed zona pellucida from the 2-cell, 4-cell and blastocyst embryos with 0.5% pronase in PBS at 37 °C. Then, the embryos were treated as follows: (a) fixation for 30 min in 4% paraformaldehyde in PHEM buffer (60 mM Pipes, 25 mM Hepes, 10 mM EGTA and 4 mM MgSO4, pH 7); (b) permeabilization with 0.25% TritonX-100 to observe GR; (c) block for 1 h in PBS with 3% bovine serum albumin (BSA); (d) overnight incubation at 4 °C with primary antibodies in PBS with 3% BSA; (e) 1-h incubation with secondary antibodies in PBS with 3% BSA; (f) 10-min incubation in PBS with 10 μg/mL of Hoechst 33342; and (g) observation under a Leica laser-scanning confocal microscope (TCS SP2; Leica Microsystems, GmbH, Wetzlar, Germany). We washed the embryos in PBS between treatments. We detected fluorescence using band pass emission filters (Hoechst 33342, 420–480 nm; Cy3, 560–605 nm). We also processed negative control samples with the primary antibody omitted to verify the specificity of the secondary antibodies.
The primary antibodies used are as follows: goat anti-CRHR1 polyclonal antibodies (1:500, ab59023; Abcam Co., Ltd., Cambridge, MA, USA) for CRHR1; rabbit anti-GR polyclonal antibodies (1:1000, ab196944; Abcam Co., Ltd.) for GR; rabbit anti-TNFR1 polyclonal antibodies (1:200, AF8196, Beyotime Co., Ltd.) for TNFR; and rabbit anti-FAS (1:100, ab82419, Abcam Co., Ltd.) for FAS. Cy3-conjugated AffiniPure goat anti-rabbit IgG (1:500; Jackson ImmunoResearch, West Grove, PA, USA) were used as secondary antibodies for GR, TNFR1 and FAS and Cy3-conjugated donkey anti-goat IgG (1:500; Beyotime Co., Ltd.) were used for CRHR1 secondary antibodies.
2.13. RNA Interference
The RiboBio (Guangzhou, China) designed and synthesized the siRNAs targeting mRNAs and the negative control siRNA. The sense strands of siRNAs targeting the FASL gene were FL siRNA-1 (5-CAA TCT ACC CTC TGA GAAA-3), FL siRNA-2 (5-GTC AGT ACT GCA ACA ACCA-3), and FL siRNA-3 (5-GAT GAA CTA TTG CAC TACT-3); those targeting the TNFα gene were TN siRNA-1 (5-CTC AGA TCA TCG TCT CAAA-3), TN siRNA-2 (5-CGA CTA TCT GGA CTT TGCT-3) and TN siRNA-3 (5-CCT ACC AGA CCA AGG TCAA-3); and we used siR-RiboTM for negative control. We performed transfection with 100 nM siRNAs using the lipofectamine RNAiMAX reagent (Invitrogen/Life Technologies, Grand Island, NY, USA). When OECs grew to approximately 60% of confluence, we replaced the spent medium in the wells with 450 μL fresh DMEM/F-12 medium and transfected the cells using the forward transfection method. To prepare the transfection complex, we diluted 1 μL of a 20 μM solution of each siRNA in 50-μL Opti-MEM medium (Invitrogen), and mixed it with 3 μL of Lipofectamine RNAiMAX reagent (Invitrogen) diluted in 50 μL Opti-MEM medium. Following incubation for 5 min at room temperature, we added 50 μL of the transfection complex to the wells before incubation for 6 h at 38.5 °C in a humidified 5% CO2 atmosphere. The transfected cells were further cultured for 48 h in serum-free DMEM/F12 with or without CRH or cortisol before recovery of CM or cells for ELISA measurement of FASL or TNFα or for flow cytometry analysis of apoptosis, respectively.
2.14. Data Analysis
Each treatment contained at least three replicates, unless otherwise specified. We used the software of Statistics Package for Social Sciences (SPSS 11.5, IBM, Chicago, IL, USA) to perform data analysis. We arc sine-transformed the percentage data before analysis. We used independent t-test to analyze data when each measure contained only two groups, and used ANOVA when each measure had more than two groups. We carried out a Duncan multiple comparison test to locate differences during ANOVA. We expressed data as means ± SEM, and considered differences significant when p < 0.05.
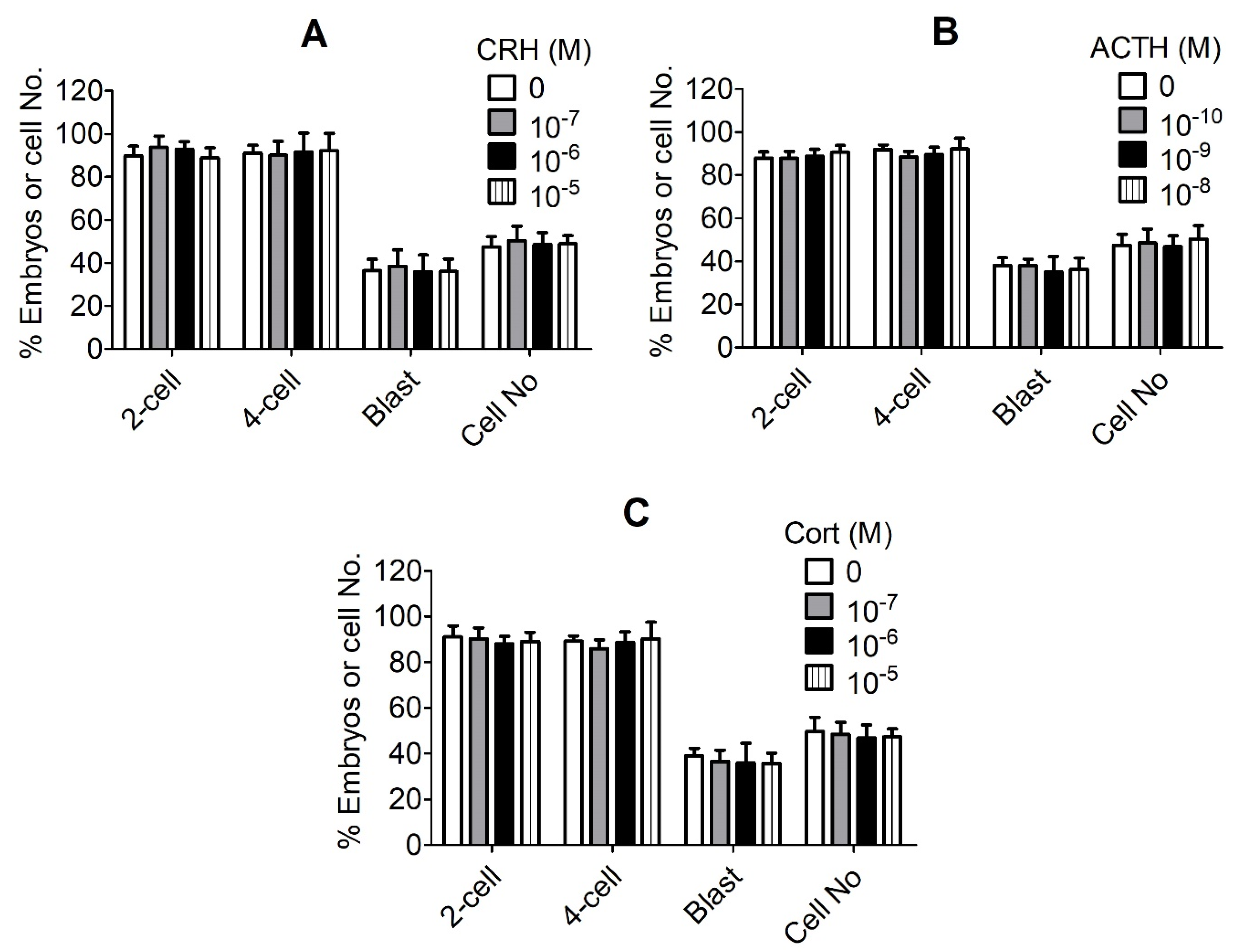
4. Discussion
Although it was reported that repeated ACTH stimulation [
16] or food deprivation [
20] after ovulation in sows significantly impaired the cleavage of the preimplantation embryos while increasing the level of serum cortisol, the mechanisms by which the stress hormones affect embryo development are largely unknown. In this study, we first observed that the preimplantation development of pig embryos was unaffected when the parthenotes were cultured with various concentrations of CRH, ACTH or cortisol, ruling out the possibility that these stress hormones affect the pig embryos directly. However, Razdan et al. [
16] observed that the stimulation of sows with ACTH impaired the cleavage of the preimplantation embryos in vivo. The present results suggest that in the study by Razdan et al. [
16], it might be the ACTH-induced elevation of cortisol that triggered OECs apoptosis and impaired embryo cleavage. To find out why CRH and cortisol did not affect the pig embryos directly, we found that pig embryos did not express CRHR1 until the blastocyst stage, and that they expressed CRHBP at all stages of the preimplantation development. It is known that CRHBP can bind CRH with an affinity greater than the CRHR to modulate the CRHR activity [
25], and it inhibits the CRH-induced ACTH secretion from pituitary corticotropes [
26]. Although pig embryos expressed GR at all the preimplantation stages observed, their mRNA level of
HSD11B2 was 45 times as much as that of
HSD11B1. According to Michael et al. [
24], whereas HSD11B1 can activate (reduce) cortisone to cortisol or corticosterone, and HSD11B2 can inactivate (oxidize) cortisol or corticosterone to cortisone. Michael et al. [
27] observed that human female conception rates following in vitro fertilization were associated with an increased ratio of cortisol:cortisone in follicular fluid, due to a low level of cortisol oxidation by HSD11B2. Furthermore, Gong et al. [
28] reported that the species difference in glucocorticoid sensitivity between pig and mouse oocytes can be attributed to their different contents/ratios of HSD11B1 and HSD11B2, which maintain different concentrations of active glucocorticoids.
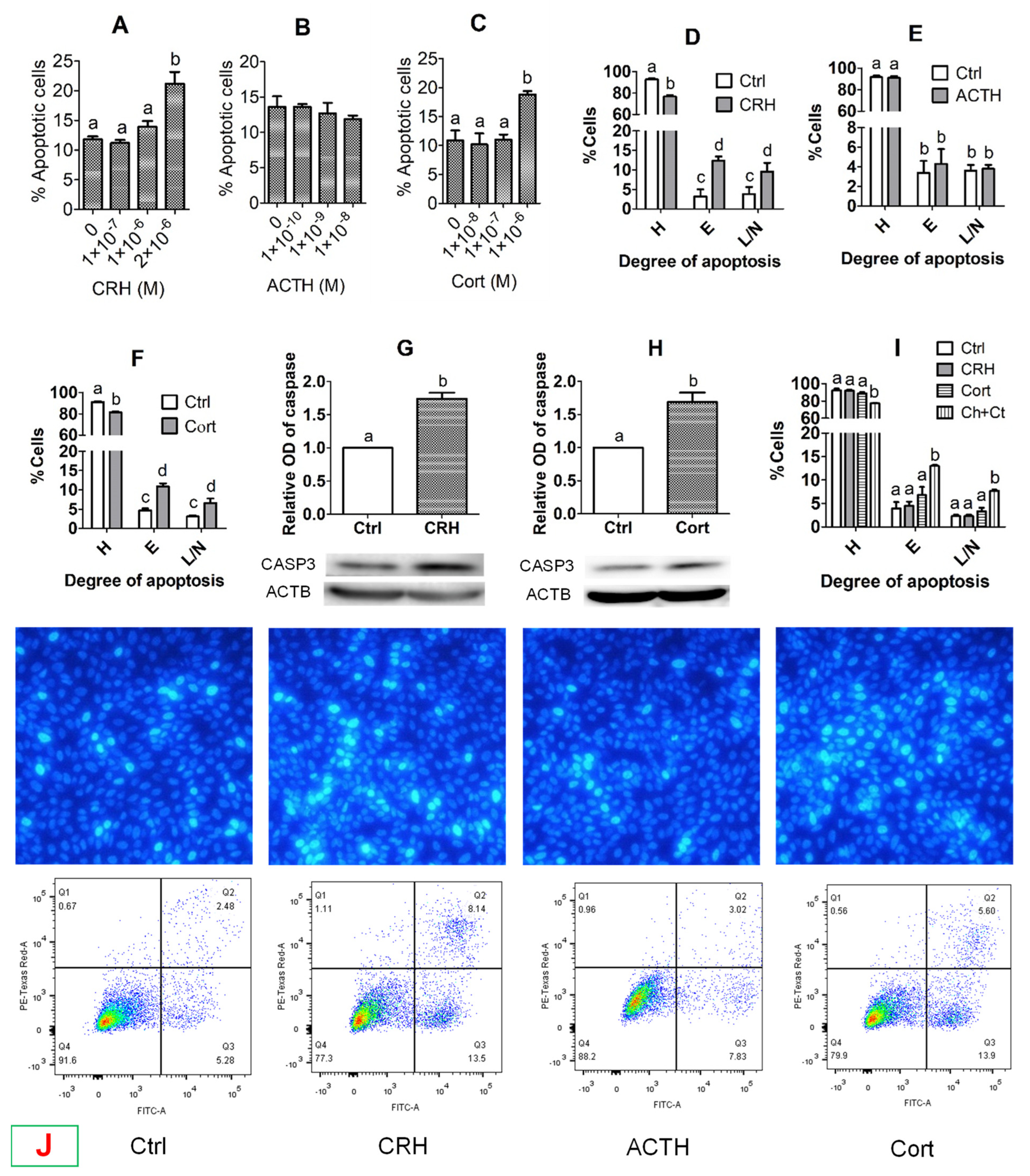
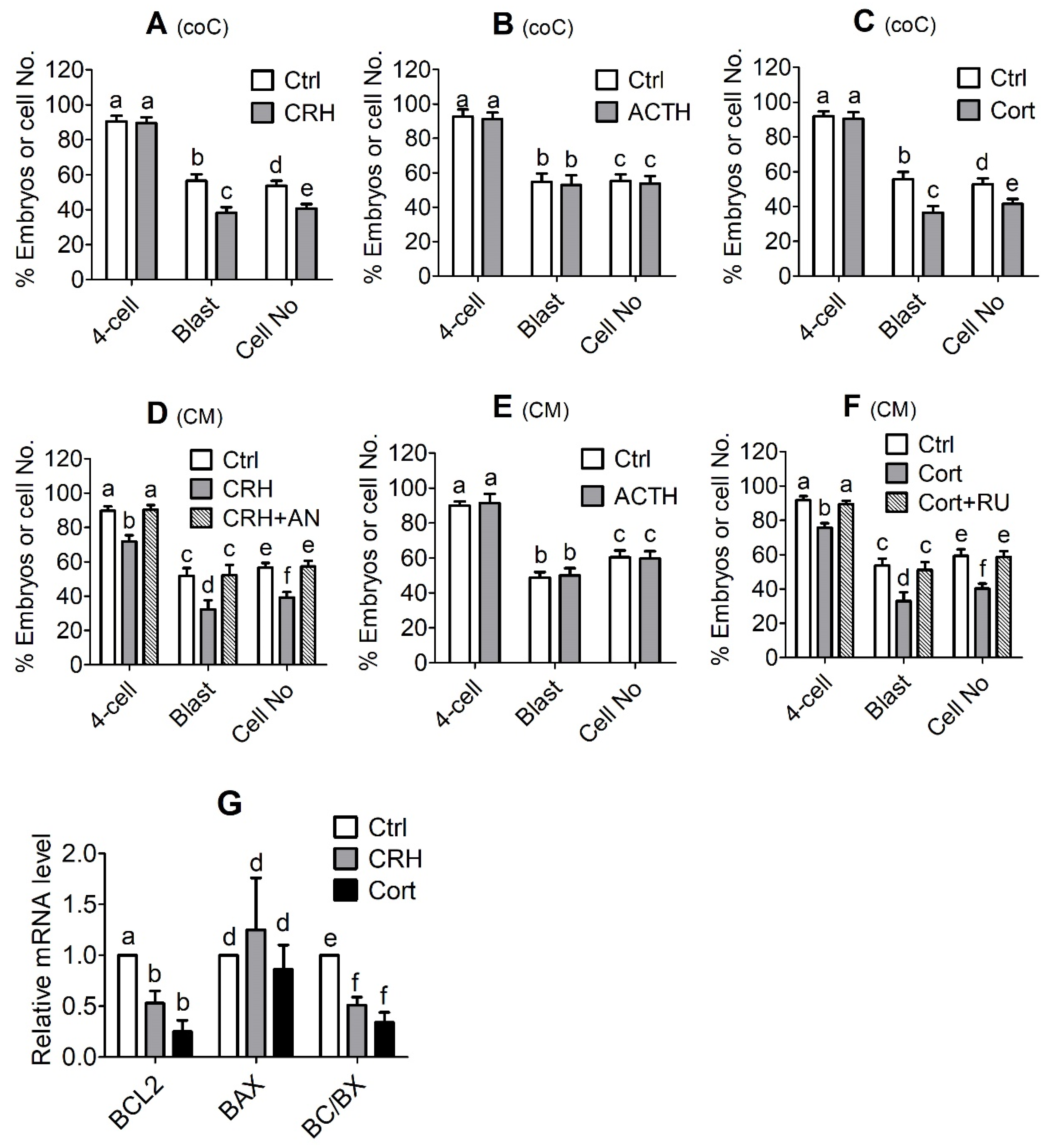
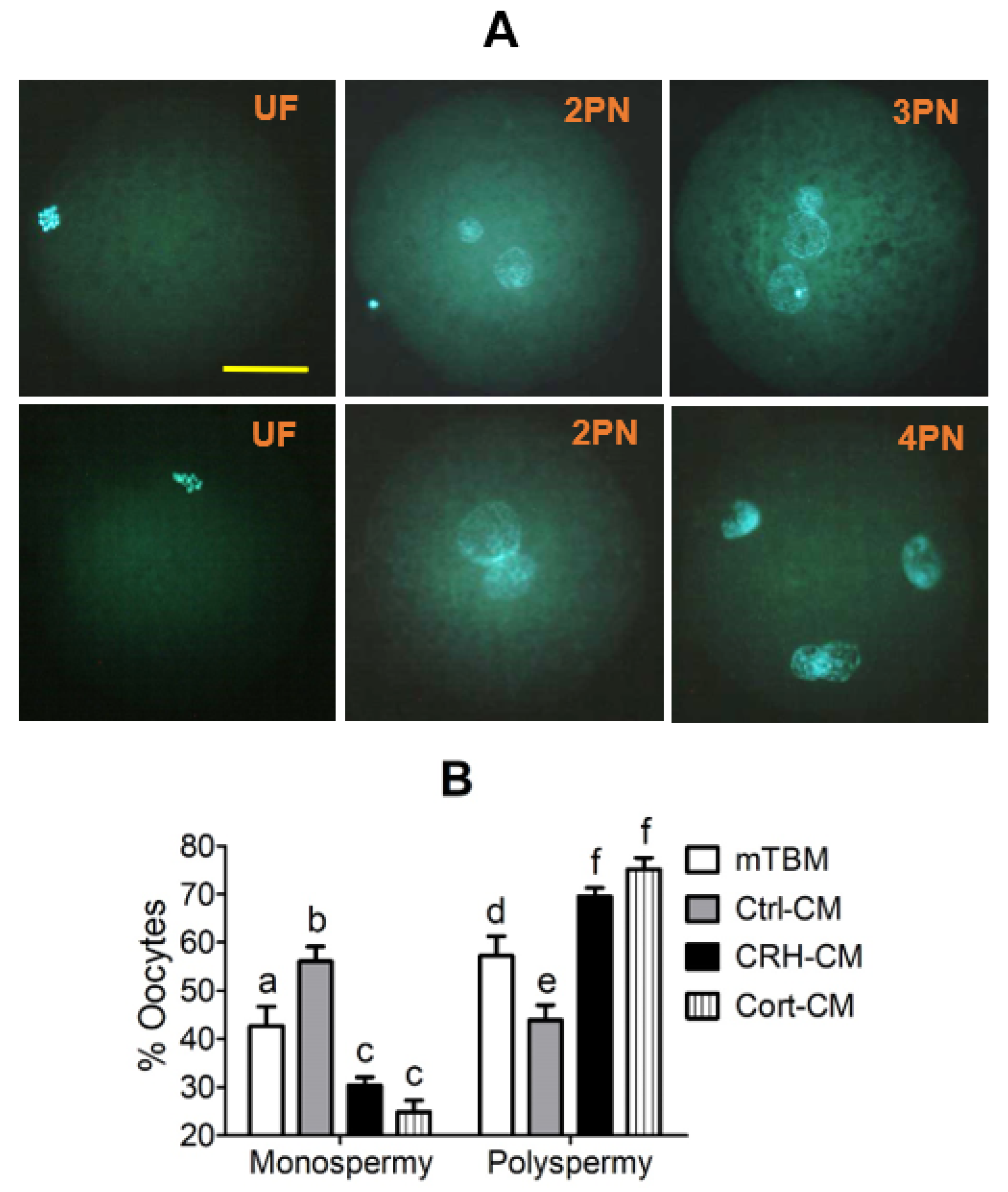
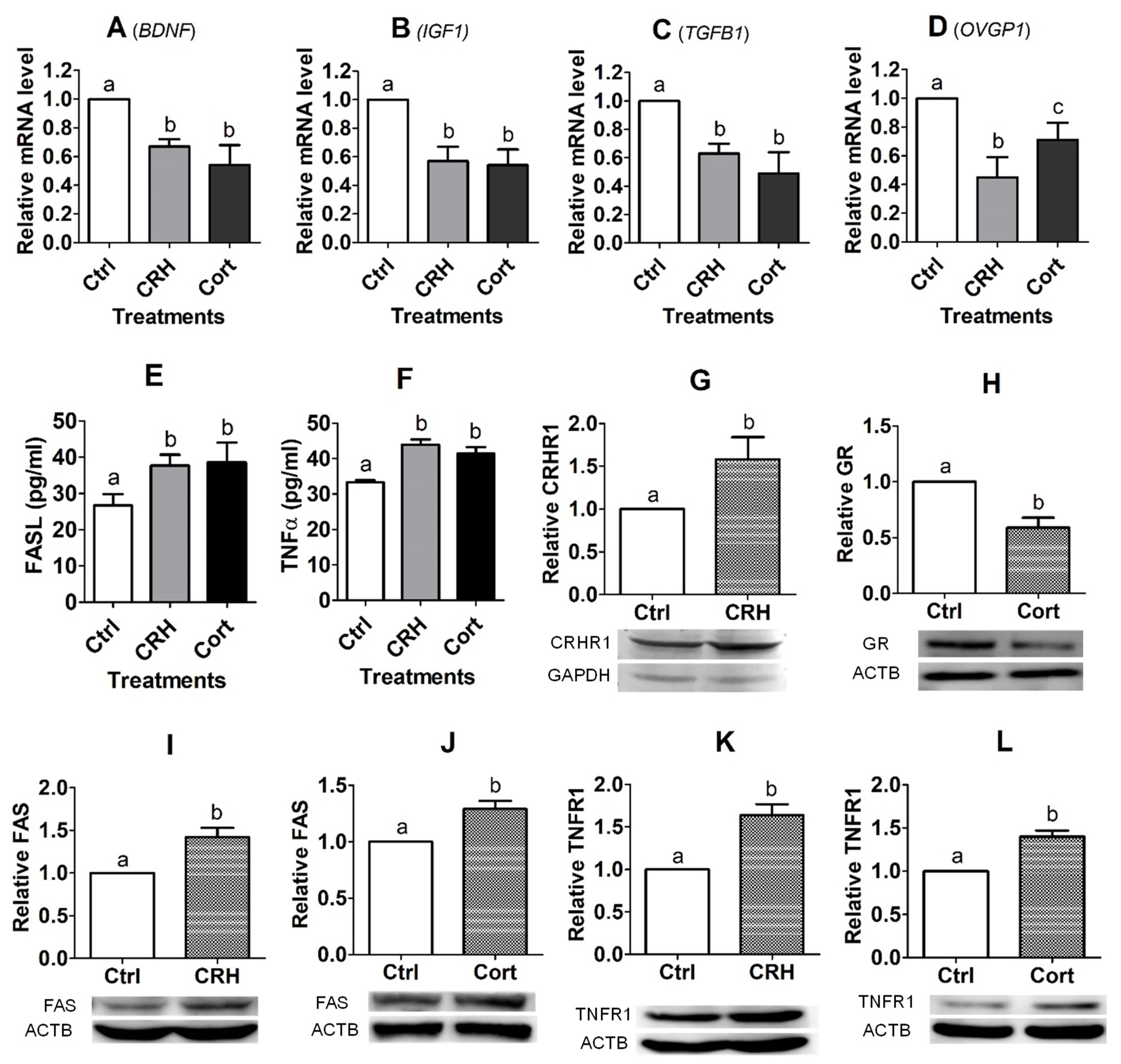
However, preimplantation development was significantly impaired when the pig parthenotes were cocultured with the OECs in the presence of CRH or cortisol or when they were cultured in CM conditioned with CRH- or cortisol-pretreated OECs. Furthermore, fertilization in CM conditioned with CRH- or cortisol-pretreated OECs significantly increased rates of polyspermy. Further investigation indicated that pig OECs underwent significant apoptosis and produced significantly less growth factors and OVGP1 after culture with CRH or cortisol. In addition, pig OECs also produced more FASL/TNFα when cultured in the presence of CRH or cortisol, and embryo culture in CM conditioned by OECs co-transfected with
FASL and
TNFα siRNAs significantly increased blastocyst rates and the cell number per blastocyst, compared with those following transfection with NC siRNA. Our immunofluorescence microscopy confirmed that FAS and TNFR1 were expressed in pig embryos at the 2-cell, 4-cell and blastocyst stages. There are many reports that OVGP1 secreted by OECs around ovulation time causes pre-fertilization zona hardening which contributes to polyspermy block in mammals, including pigs [
29,
30]. Furthermore, it has been reported that a higher incidence of polyspermy was associated with a lower level of plasma IGF1 in sows with a lactational negative energy balance [
31]. Taken together, the current results suggest that CRH and cortisol impaired fertilization and the development of pig preimplantation embryos by inducing the apoptosis of OECs and the apoptotic OECs damaged embryos by producing less antiapoptotic growth factors and OVGP1 but more proapoptotic FASL and TNFα. Because our previous study, using mouse zygotes, obtained similar results that CRH/cortisol impaired development of fertilized embryos indirectly by inducing OECs apoptosis [
8], the present results suggest that parthenotes might have the same sensitivity to CRH/cortisol as fertilized embryos do.
Many papers have documented that CRH or cortisol trigger apoptosis in various somatic cells. For instance, CRH has been reported inducing apoptosis in neurons [
32], in PC12 rat pheochromocytoma cell line [
33], in microglial cells [
34], in prostate cancer cell RM-1 [
35] and in the mural granulosa cells (MGCs) of mouse ovarian follicles [
36,
37]. It is widely known that glucocorticoids can cause apoptosis in T lymphocytes [
38]. Treatment of mice with dexamethasone significantly increased apoptosis in testicular germ cells [
39]. Culture with dexamethasone induced apoptosis in monocytes [
40] and osteocytes [
41]. Furthermore, injection of mice with cortisol [
42] or culture with corticosterone [
43] induced apoptosis in mouse MGCs.
The present results show that treatment with either CRH or cortisol significantly increased the expression of both FASL and TNFα and their receptors in pig OECs. Furthermore, knocking down either
FASL or
TNFα by RNA interference significantly reduced the apoptotic percentages of pig OECs following culture in the presence of either CRH or cortisol. Taken together, the results suggest that both CRH and cortisol induced the apoptosis of pig OECs by activating both the FASL/FAS system and the TNFα/TNFR1 signaling. Both in vitro and in vivo studies have demonstrated that CRH increased expression of FASL and/or FAS in various somatic cells [
36,
44,
45,
46,
47], and silencing the
FASL gene by RNA interference significantly alleviated the proapoptotic effect of CRH on mouse MGCs [
36]. Song et al. [
48] reported that CRH promoted TNFα production by CD14+ cells. Zhao et al. [
37] observed that CRH treatment of mouse MGCs significantly increased their apoptotic percentages and levels of TNFα and TNFR1 expression, and in vitro knockdown by interfering RNA or in vivo knockout of the
TNFα gene significantly alleviated the proapoptotic effect of CRH on MGCs.
Culture with glucocorticoids activated the Fas/FasL system in osteocytes and monocytes [
40,
41]. Treatment of mice with dexamethasone significantly increased FASL expression in testicular germ cells [
39]. The injection of mice with cortisol increased FASL secretion in ovaries and FAS expression in MGCs, cumulus cells (CCs) and oocytes, and the proapoptotic effect of cortisol injection on MGCs and CCs were significantly relieved when the
gld mice harboring
FASL mutations were observed [
42]. While Dinkel et al. [
49] reported that glucocorticoids upregulated the
TNFα signaling in rat CNS, Messmer et al. [
50] and Zhang et al. [
51] documented that the glucocorticoids inhibited TNFα expression in human mammary carcinoma cell line MCF-7 and human subcutaneous adipocytes and preadipocytes, respectively. Yuan et al. [
43] demonstrated that in mice, corticosterone treatment increased the TNFα expression in MGCs but decreased it in OECs. Furthermore, Zhao et al. [
23] found that in mice, while the CRH-induced OEC apoptosis involved both FAS signaling and TNFα signaling, corticosterone-induced OEC apoptosis involved only the FAS, but not the TNFα, signaling. Thus, the effect of glucocorticoids on TNFα expression may vary between species and/or cell types.
The present study demonstrated that treatment with ACTH did not have any detrimental effect on pig embryos, nor OECs, although pig embryos expressed the ACTH receptor. Gong et al. [
52] observed that culture with ACTH did not affect oocyte competence and embryo development in the pig, nor that in the mouse. Furthermore, they found that the ACTH receptor was expressed and ACTH-activated protein kinase A in both cumulus-denuded oocytes and CCs. Thus far, we could not find any documentation for the detrimental effects of ACTH on any type of cells.
In this study, the effective concentrations of CRH and cortisol that caused the significant apoptosis of pig OECs were higher than peak serum concentrations reported in stressed pigs. Du et al. [
53,
54] observed no effect on pig OEC apoptosis by in vitro treatment with cortisol at a stress-induced concentration (2.5 × 10
−7 M). Our previous study [
52] demonstrated that although culture with either 1 × 10
−8 M CRH or 1 mg/mL cortisol did not affect pig oocyte development into blastocysts, culture with both significantly decreased blastocyst rates. The present results showed that although culture with neither stress-induced concentration of CRH (1 × 10
−8 M) nor that of cortisol (4 × 10
−7 M) alone increased apoptosis, culture with both 1 × 10
−8 M CRH and 4 × 10
−7 M cortisol significantly increased early and late apoptosis of pig OECs. Thus, the results suggest that in vivo, stresses might increase the release of multiple hormones that work together to induce a symptom at lower concentrations than in vitro in the presence of only a single hormone.
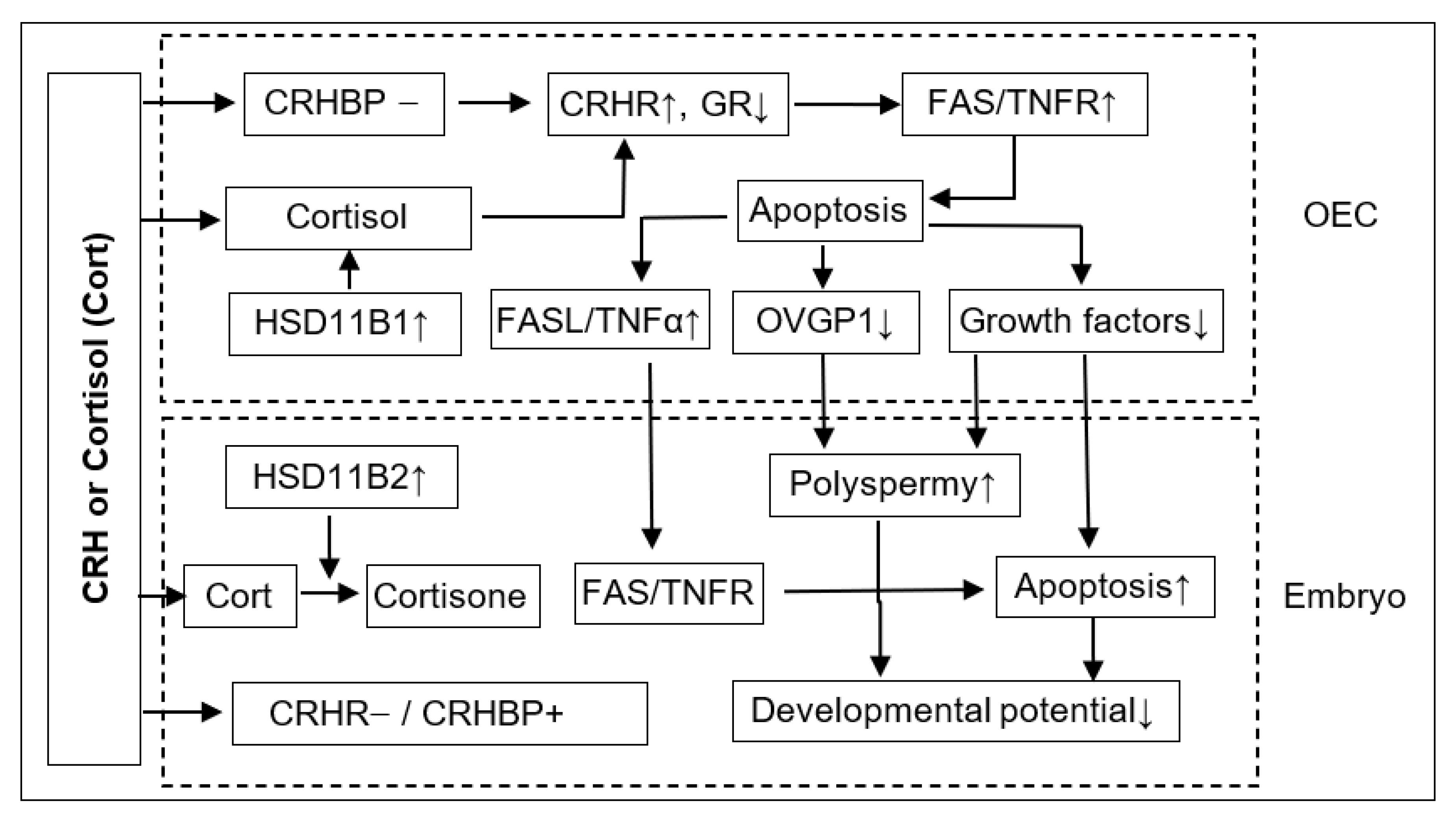
In summary, this study shows that although ACTH showed no effect on pig embryo development, CRH and cortisol impaired preimplantation embryo development and increased polyspermy indirectly by inducing the apoptosis of OECs. Thus, after the treatment of OECs with CRH or cortisol, while the expression level of GR decreased, that of CRHR1, FAS and TNFR1 increased significantly, which would lead to apoptosis (
Figure 8). The apoptotic OECs produced more FASL and TNFα but less growth factors and OVGP1. The FASL and TNFα would interact with FAS and TNFR1, respectively, on the embryo, which, together with a desperate shortage of growth factors, would trigger the apoptosis of embryos. The increased polyspermy was associated with a decrease in OVGP1 and growth factors. Both apoptosis and polyspermy would contribute to the decreased developmental potential of embryos. Pig preimplantation embryos did not express CRHR1 but did express CRHBP, which might lead to their insensitivity to CRH. Pig preimplantation embryos expressed more HSD11B2 than HSD11B1, which converts cortisol to inactive cortisone and might contribute to their tolerance to cortisol. Furthermore, when used at a stress-induced physiological concentration, while culture with either CRH or cortisol alone showed no effects, culture with both significantly increased OECs apoptosis, suggesting that in vivo, both hormones may work together to induce apoptosis of pig OECs.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}