
A High Hepatic Uptake of Conjugated Bile Acids Promotes Colorectal Cancer—Associated Liver Metastasis

 , , ,
, , ,  ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Animals and Ethics Statement
2.2. Establishment of CRC–Associated Liver or Lung Metastatic Mouse Models
2.3. Intervention of TCA or NTCP Neutralizing Antibody
2.4. Detection of Serum TC, TG, LDL and HDL
2.5. Quantification of Serum Metabolomic Profile
2.6. Quantitation of BA Metabolites in Serum and Liver
2.7. Hematoxylin–Eosin Staining
2.8. Immunofluorescence
2.9. Flow Cytometry
2.10. RNA Sequencing and Data Processing
2.11. Primary Hepatic Stellate Cells Isolation and Incubation
2.12. Cell Culture and Treatment
2.13. Transwell Assays
2.14. RNA Extraction and Quantitative RT–qPCR
2.15. Western Blot
2.16. Statistical Analysis
3. Results
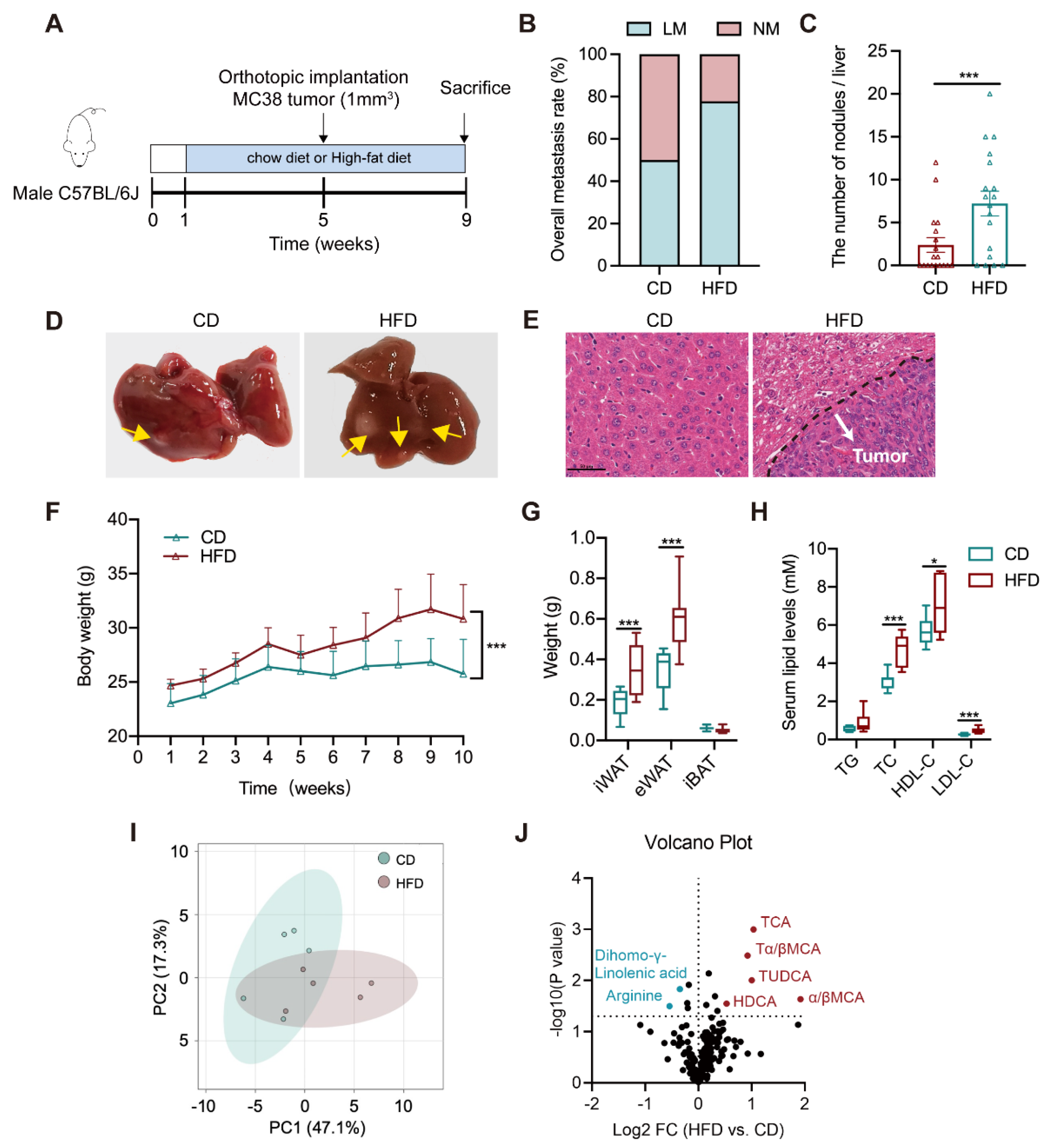
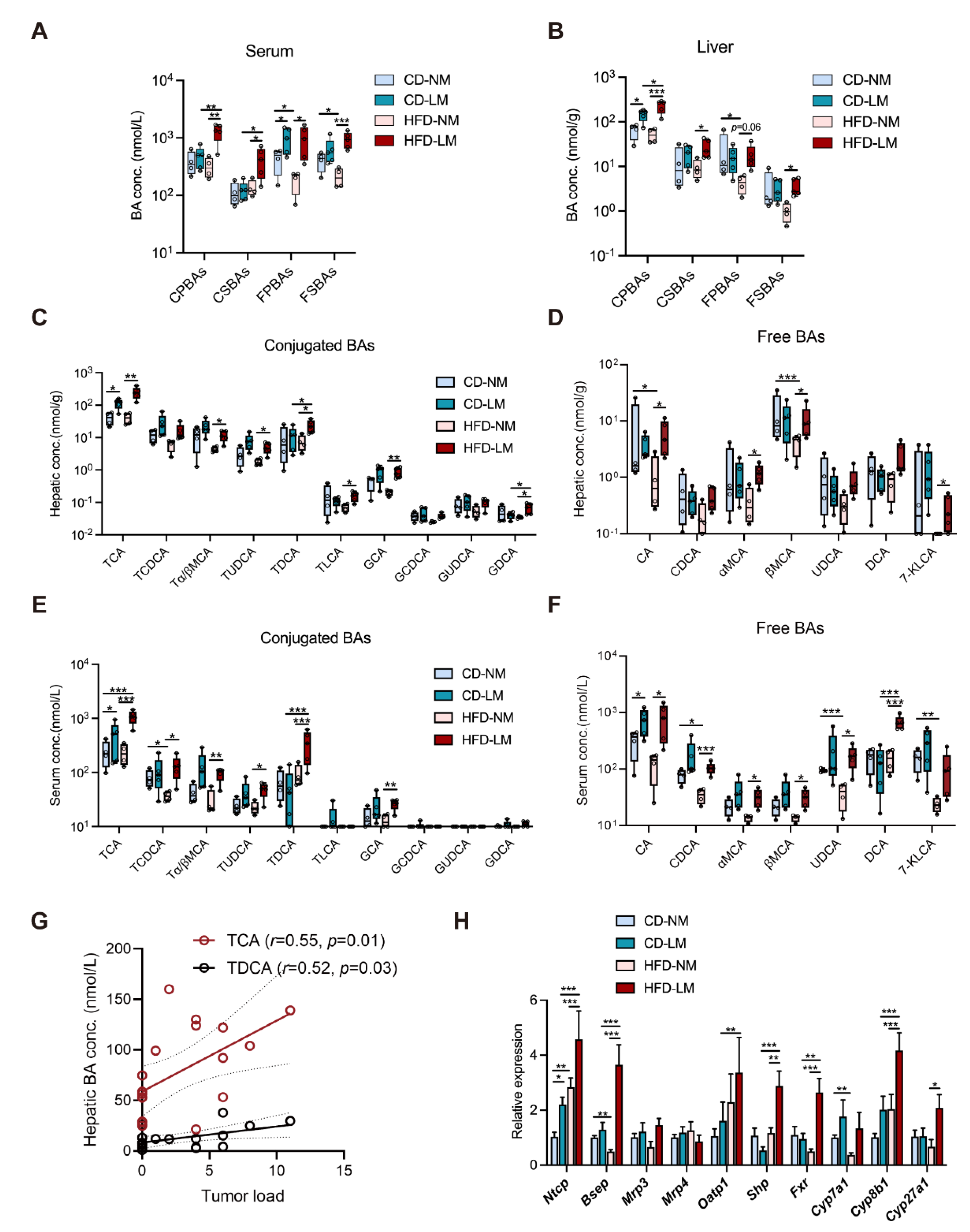
3.1. Altered Bile Acid Profile Is Associated with Liver Metastasis
3.2. Increased Hepatic TCA Level and Ntcp Expression in Liver-Metastatic Mice
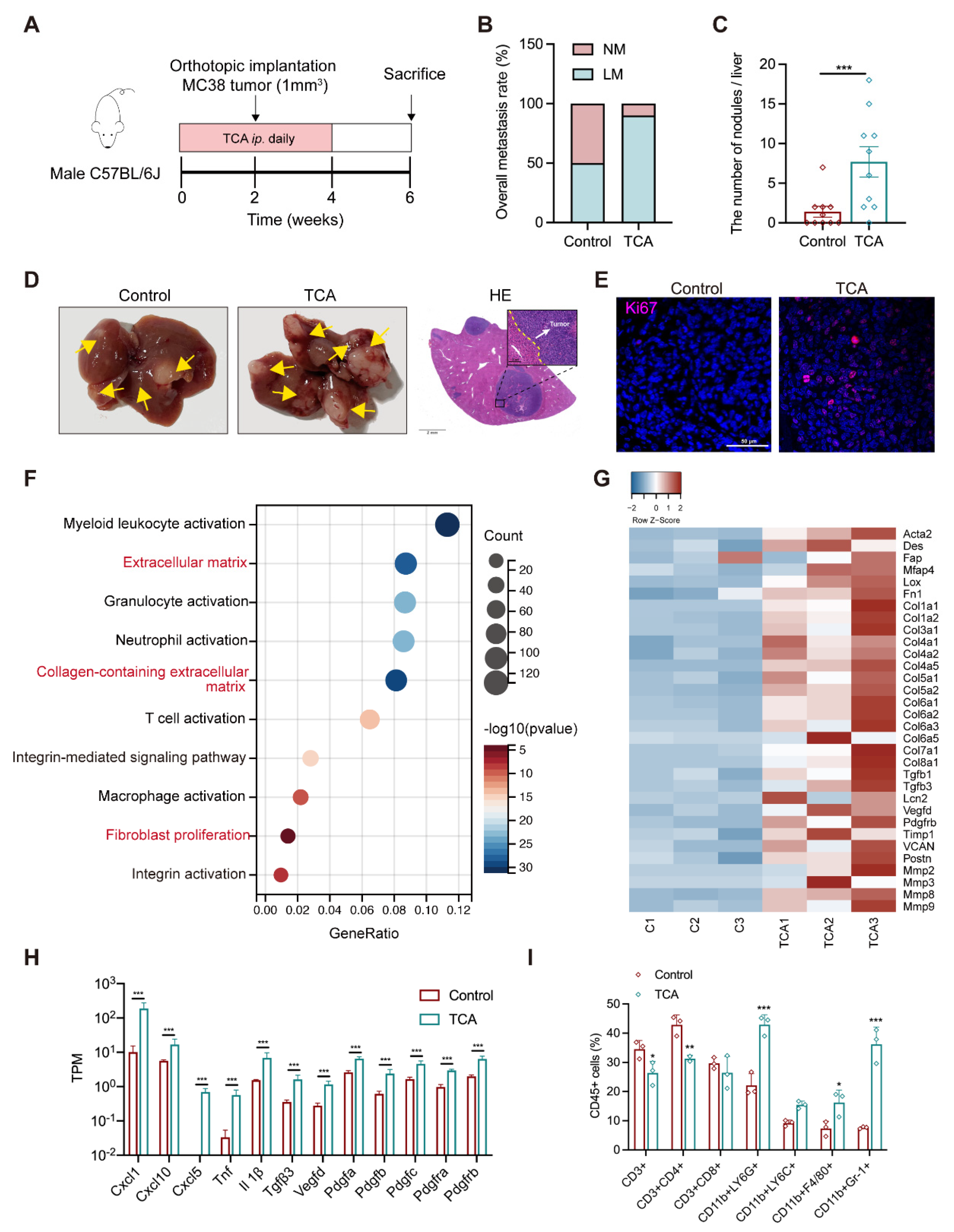
3.3. TCA Promotes Liver Metastases and Induces a Tumor-Favorable Microenvironment
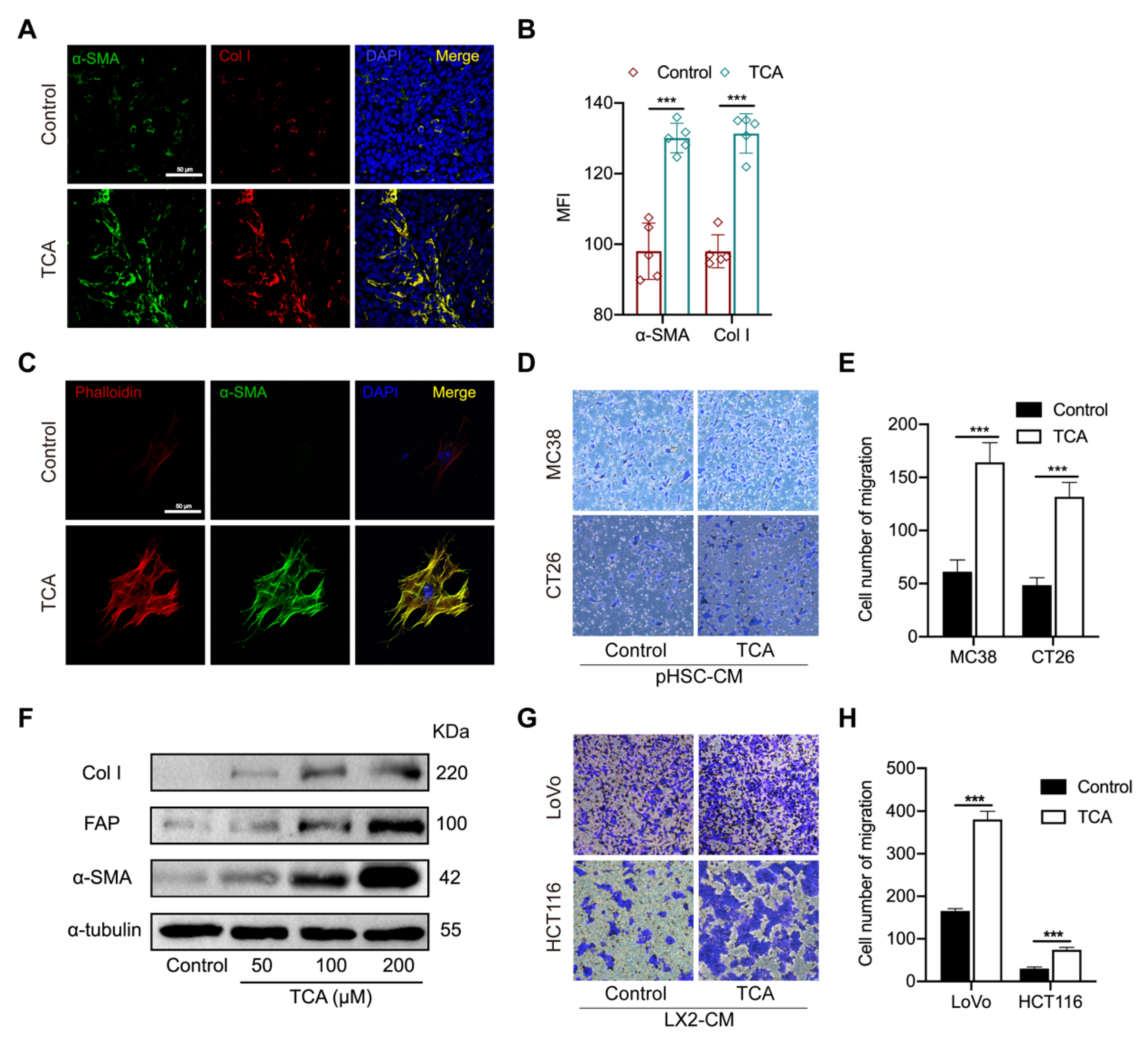
3.4. TCA–Activated HSCs Enhance the Migration Capability of CRC Cells
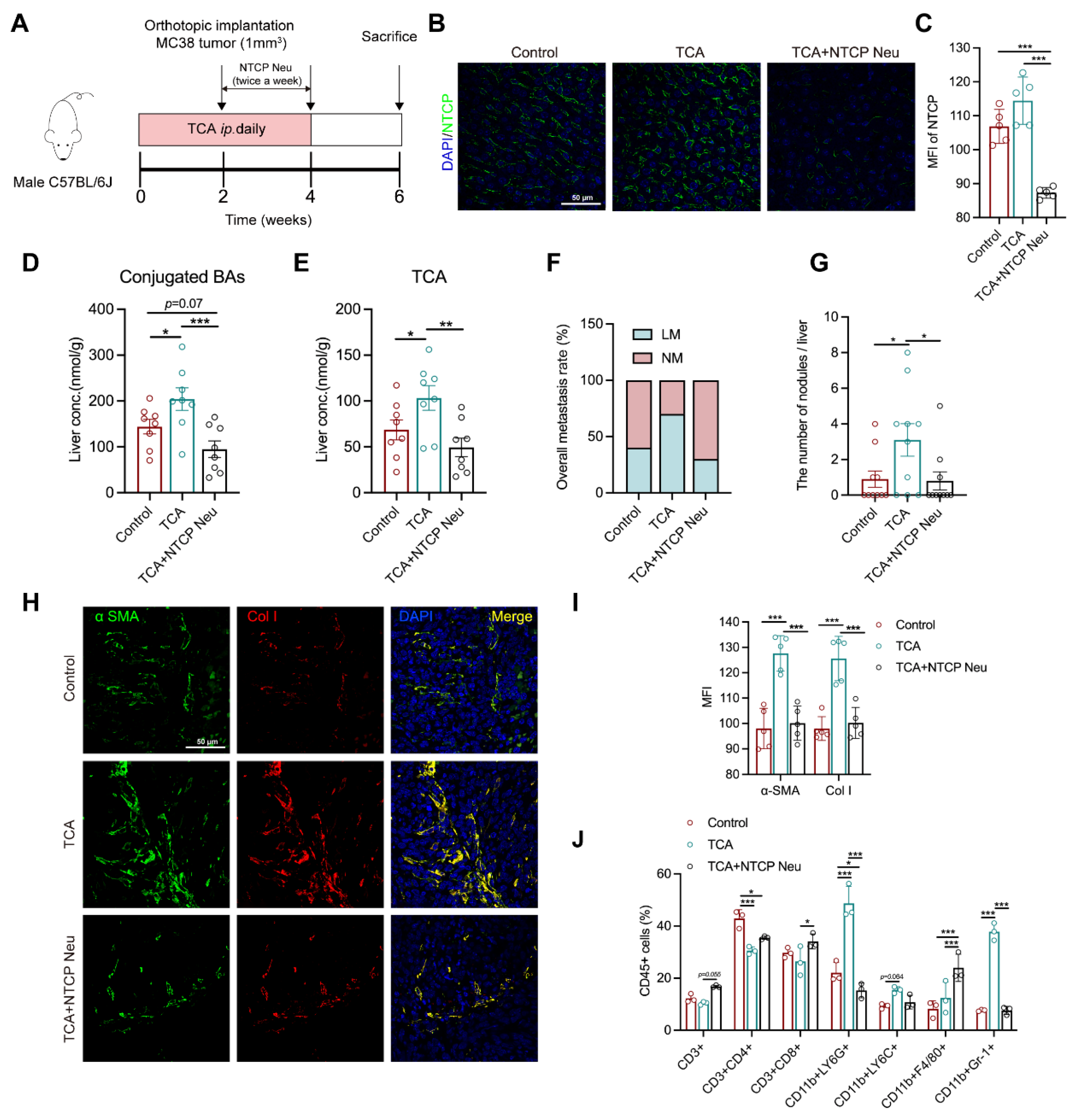
3.5. Blockade of TCA Uptake Effectively Attenuates Liver Metastasis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Yang, Y.; Han, Z.; Li, X.; Huang, A.; Shi, J.; Gu, J. Epidemiology and risk factors of colorectal cancer in China. Chin. J. Cancer Res. 2020, 32, 729–741. [Google Scholar] [CrossRef]
- van der Geest, L.G.M.; Lam-Boer, J.T.; Koopman, M.; Verhoef, C.; Elferink, M.A.G.; de Wilt, J.H.W. Nationwide trends in incidence, treatment and survival of colorectal cancer patients with synchronous metastases. Clin. Exp. Metastasis 2015, 32, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.B.; Bekaii-Saab, T.; Chan, E.; Chen, Y.-J.; Choti, M.A.; Cooper, H.S.; Engstrom, P.F.; Enzinger, P.C.; Fakih, M.G.; Fenton, M.J.; et al. Metastatic colon cancer, version 3.2013: Featured updates to the NCCN Guidelines. J. Natl. Compr. Cancer Netw. 2013, 11, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Nordlinger, B.; Sorbye, H.; Glimelius, B.; Poston, G.J.; Schlag, P.M.; Rougier, P.; Bechstein, W.O.; Primrose, J.N.; Walpole, E.T.; Finch-Jones, M.; et al. Perioperative FOLFOX4 chemotherapy and surgery versus surgery alone for resectable liver metastases from colorectal cancer (EORTC 40983): Long-term results of a randomised, controlled, phase 3 trial. Lancet Oncol. 2013, 14, 1208–1215. [Google Scholar] [CrossRef]
- Viganò, L.; Capussotti, L.; Lapointe, R.; Barroso, E.; Hubert, C.; Giuliante, F.; Ijzermans, J.N.M.; Mirza, D.F.; Elias, D.; Adam, R. Early recurrence after liver resection for colorectal metastases: Risk factors, prognosis, and treatment. A LiverMetSurvey-based study of 6025 patients. Ann. Surg. Oncol. 2014, 21, 1276–1286. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Wang, K.; Ding, Y.; Li, H.; Liu, Y.; Ding, L. Which patients are prone to suffer liver metastasis? A review of risk factors of metachronous liver metastasis of colorectal cancer. Eur. J. Med. Res. 2022, 27, 130. [Google Scholar] [CrossRef]
- Zarour, L.R.; Anand, S.; Billingsley, K.G.; Bisson, W.H.; Cercek, A.; Clarke, M.F.; Coussens, L.M.; Gast, C.E.; Geltzeiler, C.B.; Hansen, L.; et al. Colorectal Cancer Liver Metastasis: Evolving Paradigms and Future Directions. Cell. Mol. Gastroenterol. Hepatol. 2017, 3, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Liu, Z.; Wang, Y.; Wen, X.; Amador, E.H.; Yuan, L.; Ran, X.; Xiong, L.; Ran, Y.; Chen, W.; et al. Colorectal liver metastasis: Molecular mechanism and interventional therapy. Signal Transduct. Target. Ther. 2022, 7, 70. [Google Scholar] [CrossRef]
- Zhang, K.L.; Zhu, W.W.; Wang, S.H.; Gao, C.; Pan, J.J.; Du, Z.G.; Lu, L.; Jia, H.L.; Dong, Q.Z.; Chen, J.H.; et al. Organ-specific cholesterol metabolic aberration fuels liver metastasis of colorectal cancer. Theranostics 2021, 11, 6560–6572. [Google Scholar] [CrossRef]
- Meyerhardt, J.A.; Niedzwiecki, D.; Hollis, D.; Saltz, L.B.; Hu, F.B.; Mayer, R.J.; Nelson, H.; Whittom, R.; Hantel, A.; Thomas, J.; et al. Association of dietary patterns with cancer recurrence and survival in patients with stage III colon cancer. JAMA 2007, 298, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Cai, Y.; Yang, B.; Xie, S.; Shen, W.; Wu, Y.; Sui, Z.; Cai, J.; Ni, C.; Ye, J. High-Fat Diet Enhances the Liver Metastasis Potential of Colorectal Cancer through Microbiota Dysbiosis. Cancers 2022, 14, 2573. [Google Scholar] [CrossRef] [PubMed]
- Tseng, W.; Leong, X.; Engleman, E. Orthotopic mouse model of colorectal cancer. J. Vis. Exp. 2007, 10, e484. [Google Scholar] [CrossRef] [PubMed]
- Salhab, A.; Amer, J.; Lu, Y.; Safadi, R. Sodium(+)/taurocholate cotransporting polypeptide as target therapy for liver fibrosis. Gut 2022, 71, 1373–1385. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Wang, L.; Chen, T.; Zhou, K.; Zhang, Z.; Li, J.; Sun, B.; Guo, Y.; Wang, X.; Wang, Y.; et al. A Metabolite Array Technology for Precision Medicine. Anal. Chem. 2021, 93, 5709–5717. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, W.; Chen, Y.; Huang, F.; Lu, L.; Lin, C.; Huang, T.; Ning, Z.; Zhai, L.; Zhong, L.L.; et al. A Clostridia-rich microbiota enhances bile acid excretion in diarrhea-predominant irritable bowel syndrome. J. Clin. Investig. 2020, 130, 438–450. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA–seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Kühn, T.; Stepien, M.; López-Nogueroles, M.; Damms-Machado, A.; Sookthai, D.; Johnson, T.; Roca, M.; Hüsing, A.; Maldonado, S.G.; Cross, A.J.; et al. Prediagnostic Plasma Bile Acid Levels and Colon Cancer Risk: A Prospective Study. J. Natl. Cancer Inst. 2020, 112, 516–524. [Google Scholar] [CrossRef]
- Zhang, R.; Qi, F.; Zhao, F.; Li, G.; Shao, S.; Zhang, X.; Yuan, L.; Feng, Y. Cancer-associated fibroblasts enhance tumor-associated macrophages enrichment and suppress NK cells function in colorectal cancer. Cell Death Dis. 2019, 10, 273. [Google Scholar] [CrossRef]
- Kobayashi, H.; Gieniec, K.A.; Lannagan, T.R.M.; Wang, T.; Asai, N.; Mizutani, Y.; Iida, T.; Ando, R.; Thomas, E.M.; Sakai, A.; et al. The Origin and Contribution of Cancer-Associated Fibroblasts in Colorectal Carcinogenesis. Gastroenterology 2022, 162, 890–906. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Mederacke, I.; Hsu, C.C.; Troeger, J.S.; Huebener, P.; Mu, X.; Dapito, D.H.; Pradere, J.-P.; Schwabe, R.F. Fate tracing reveals hepatic stellate cells as dominant contributors to liver fibrosis independent of its aetiology. Nat. Commun. 2013, 4, 2823. [Google Scholar] [CrossRef] [PubMed]
- Matsusue, R.; Kubo, H.; Hisamori, S.; Okoshi, K.; Takagi, H.; Hida, K.; Nakano, K.; Itami, A.; Kawada, K.; Nagayama, S.; et al. Hepatic stellate cells promote liver metastasis of colon cancer cells by the action of SDF–1/CXCR4 axis. Ann. Surg. Oncol. 2009, 16, 2645–2653. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Mi, Y.; Zheng, B.; Wei, P.; Gu, Y.; Zhang, Z.; Xu, Y.; Cai, S.; Li, X.; Li, D. Highly-metastatic colorectal cancer cell released miR-181a-5p-rich extracellular vesicles promote liver metastasis by activating hepatic stellate cells and remodelling the tumour microenvironment. J. Extracell. Vesicles 2022, 11, e12186. [Google Scholar] [CrossRef]
- Chiang, J.Y. Bile acids: Regulation of synthesis. J. Lipid Res. 2009, 50, 1955–1966. [Google Scholar] [CrossRef]
- Chiang, J.Y.L. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [CrossRef]
- Sayin, S.I.; Wahlström, A.; Felin, J.; Jäntti, S.; Marschall, H.-U.; Bamberg, K.; Angelin, B.; Hyötyläinen, T.; Orešič, M.; Bäckhed, F. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. 2013, 17, 225–235. [Google Scholar] [CrossRef]
- Islam, K.B.M.S.; Fukiya, S.; Hagio, M.; Fujii, N.; Ishizuka, S.; Ooka, T.; Ogura, Y.; Hayashi, T.; Yokota, A. Bile acid is a host factor that regulates the composition of the cecal microbiota in rats. Gastroenterology 2011, 141, 1773–1781. [Google Scholar] [CrossRef]
- Zheng, X.; Huang, F.; Zhao, A.; Lei, S.; Zhang, Y.; Xie, G.; Chen, T.; Qu, C.; Rajani, C.; Dong, B.; et al. Bile acid is a significant host factor shaping the gut microbiome of diet-induced obese mice. BMC Biol. 2017, 15, 120. [Google Scholar] [CrossRef]
- Nakade, Y.; Kitano, R.; Sakamoto, K.; Kimoto, S.; Yamauchi, T.; Inoue, T.; Kobayashi, Y.; Ohashi, T.; Sumida, Y.; Ito, K.; et al. Characteristics of bile acid composition in high fat diet-induced nonalcoholic fatty liver disease in obese diabetic rats. PLoS ONE 2021, 16, e0247303. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; He, F.; Yan, X.; Xing, Y.; Lei, Y.; Gao, J.; He, M.; Li, D.; Bai, L.; Yuan, Z.; et al. Hepatic Reduction in Cholesterol 25-Hydroxylase Aggravates Diet-induced Steatosis. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1161–1179. [Google Scholar] [CrossRef] [PubMed]
- Anwer, M.S. Cellular regulation of hepatic bile acid transport in health and cholestasis. Hepatology 2004, 39, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Geier, A.; Martin, I.V.; Dietrich, C.G.; Balasubramaniyan, N.; Strauch, S.; Suchy, F.J.; Gartung, C.; Trautwein, C.; Ananthanarayanan, M. Hepatocyte nuclear factor-4alpha is a central transactivator of the mouse Ntcp gene. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G226–G233. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y.L. Hepatocyte nuclear factor 4alpha regulation of bile acid and drug metabolism. Expert Opin. Drug Metab. Toxicol. 2009, 5, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kang, H.S.; Kim, K.-H. Roles of hepatocyte nuclear factors in hepatitis B virus infection. World J. Gastroenterol. 2016, 22, 7017–7029. [Google Scholar] [CrossRef] [PubMed]
- Mani, U.V.; Bhatt, S.; Mehta, N.C.; Pradhan, S.N.; Shah, V.; Mani, I. Glycemic index of traditional Indian carbohydrate foods. J. Am. Coll. Nutr. 1990, 9, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Fujino, S.; Miyoshi, N.; Ito, A.; Yasui, M.; Matsuda, C.; Ohue, M.; Uemura, M.; Mizushima, T.; Doki, Y.; Eguchi, H. HNF1A regulates colorectal cancer progression and drug resistance as a downstream of POU5F1. Sci. Rep. 2021, 11, 10363. [Google Scholar] [CrossRef] [PubMed]
- Sang, L.; Wang, X.; Bai, W.; Shen, J.; Zeng, Y.; Sun, J. The role of hepatocyte nuclear factor 4α (HNF4α) in tumorigenesis. Front. Oncol. 2022, 12, 1011230. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Yu, A.-M.; Yim, S.H.; Ma, X.; Krausz, K.W.; Inoue, J.; Xiang, C.C.; Brownstein, M.J.; Eggertsen, G.; Björkhem, I.; et al. Regulation of bile acid biosynthesis by hepatocyte nuclear factor 4alpha. J. Lipid Res. 2006, 47, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Qu, X.; Zhang, L.; Wang, N.; Chen, M.; Zhao, X.; Wang, J.; Lv, H.; Qi, Y.; Zhang, L.; et al. Serum Total Bile Acids in Relation to Gastrointestinal Cancer Risk: A Retrospective Study. Front. Oncol. 2022, 12, 859716. [Google Scholar] [CrossRef]
- Yachida, S.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Watanabe, H.; Masuda, K.; Nishimoto, Y.; Kubo, M.; et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat. Med. 2019, 25, 968–976. [Google Scholar] [CrossRef]
- Fu, T.; Coulter, S.; Yoshihara, E.; Oh, T.G.; Fang, S.; Cayabyab, F.; Zhu, Q.; Zhang, T.; Leblanc, M.; Liu, S.; et al. FXR Regulates Intestinal Cancer Stem Cell Proliferation. Cell 2019, 176, 1098–1112.e1018. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Ung, T.T.; Kim, N.H.; Jung, Y.D. Role of bile acids in colon carcinogenesis. World J. Clin. Cases 2018, 6, 577–588. [Google Scholar] [CrossRef]
- Liu, L.; Dong, W.; Wang, S.; Zhang, Y.; Liu, T.; Xie, R.; Wang, B.; Cao, H. Deoxycholic acid disrupts the intestinal mucosal barrier and promotes intestinal tumorigenesis. Food Funct. 2018, 9, 5588–5597. [Google Scholar] [CrossRef]
- Ignacio Barrasa, J.; Olmo, N.; Pérez-Ramos, P.; Santiago-Gómez, A.; Lecona, E.; Turnay, J.; Antonia Lizarbe, M. Deoxycholic and chenodeoxycholic bile acids induce apoptosis via oxidative stress in human colon adenocarcinoma cells. Apoptosis 2011, 16, 1054–1067. [Google Scholar] [CrossRef]
- Payne, C.M.; Bernstein, C.; Dvorak, K.; Bernstein, H. Hydrophobic bile acids, genomic instability, Darwinian selection, and colon carcinogenesis. Clin. Exp. Gastroenterol. 2008, 1, 19–47. [Google Scholar] [CrossRef]
- Pai, R.; Tarnawski, A.S.; Tran, T. Deoxycholic acid activates beta-catenin signaling pathway and increases colon cell cancer growth and invasiveness. Mol. Biol. Cell 2004, 15, 2156–2163. [Google Scholar] [CrossRef]
- Arvind, P.; Papavassiliou, E.D.; Tsioulias, G.J.; Duceman, B.W.; Lovelace, C.I.; Geng, W.; Staiano-Coico, L.; Rigas, B. Lithocholic acid inhibits the expression of HLA class I genes in colon adenocarcinoma cells. Differential effect on HLA-A, -B and -C loci. Mol. Immunol. 1994, 31, 607–614. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Lian, S.; Ung, T.T.; Xia, Y.; Han, J.Y.; Jung, Y.D. Lithocholic Acid Stimulates IL-8 Expression in Human Colorectal Cancer Cells Via Activation of Erk1/2 MAPK and Suppression of STAT3 Activity. J. Cell. Biochem. 2017, 118, 2958–2967. [Google Scholar] [CrossRef]
- Liu, R.; Li, X.; Hylemon, P.B.; Zhou, H. Conjugated Bile Acids Promote Invasive Growth of Esophageal Adenocarcinoma Cells and Cancer Stem Cell Expansion via Sphingosine 1-Phosphate Receptor 2-Mediated Yes-Associated Protein Activation. Am. J. Pathol. 2018, 188, 2042–2058. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-K.; Jeong, S.-H.; Jang, C.; Bae, H.; Kim, Y.H.; Park, I.; Kim, S.K.; Koh, G.Y. Tumor metastasis to lymph nodes requires YAP-dependent metabolic adaptation. Science 2019, 363, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Jiang, R.; Wang, X.; Liu, P.; Zhao, A.; Wu, Y.; Huang, F.; Liu, Z.; Rajani, C.; Zheng, X.; et al. Conjugated secondary 12α-hydroxylated bile acids promote liver fibrogenesis. EBioMedicine 2021, 66, 103290. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Mak, K.M.; Ren, C.; Lieber, C.S. Leptin stimulates tissue inhibitor of metalloproteinase-1 in human hepatic stellate cells: Respective roles of the JAK/STAT and JAK-mediated H2O2-dependent MAPK pathways. J. Biol. Chem. 2004, 279, 4292–4304. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.; Gores, G.J.; Shah, V.H. Hepatic stellate cells: Partners in crime for liver metastases? Hepatology 2011, 54, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, T.; Shi, H.; Zhou, Z.; Huang, Z.; Lei, X. The cellular and molecular components involved in pre-metastatic niche formation in colorectal cancer liver metastasis. Expert Rev. Gastroenterol. Hepatol. 2021, 15, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Sugata, K.; Kamimura, S.; Shijo, H. Differential effects between tauroursodeoxycholic and taurochenodeoxycholic acids in hepatic fibrosis: An assessment by primary cultured Ito and Kupffer cells from the rat liver. J. Gastroenterol. Hepatol. 1996, 11, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Williamson, T.; Sultanpuram, N.; Sendi, H. The role of liver microenvironment in hepatic metastasis. Clin. Transl. Med. 2019, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Zhang, Z.; Bao, R.; Guo, X.; Gu, Y.; Yang, W.; Wei, J.; Chen, X.; Tong, L.; Meng, J.; et al. Loss of SIRT5 promotes bile acid-induced immunosuppressive microenvironment and hepatocarcinogenesis. J. Hepatol. 2022, 77, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Bertocchi, A.; Carloni, S.; Ravenda, P.S.; Bertalot, G.; Spadoni, I.; Lo Cascio, A.; Gandini, S.; Lizier, M.; Braga, D.; Asnicar, F.; et al. Gut vascular barrier impairment leads to intestinal bacteria dissemination and colorectal cancer metastasis to liver. Cancer Cell 2021, 39, 708–724.e711. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, R.; Kawada, K.; Itatani, Y.; Ogawa, R.; Kiyasu, Y.; Sakai, Y. The Role of Tumor-Associated Neutrophils in Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 529. [Google Scholar] [CrossRef] [PubMed]
- Grossman, J.G.; Nywening, T.M.; Belt, B.; Pittman, E.; Giorgi, A.; Michael, A.; Hawkins, W.G.; Strasberg, S.M.; Goedegebuure, P.S.; Fields, R.C.; et al. The role of inflammatory monocytes in human metastatic colorectal cancer. J. Clin. Oncol. 2015, 33, 624. [Google Scholar] [CrossRef]
- Rao, A.; Kosters, A.; Mells, J.E.; Zhang, W.; Setchell, K.D.R.; Amanso, A.M.; Wynn, G.M.; Xu, T.; Keller, B.T.; Yin, H.; et al. Inhibition of ileal bile acid uptake protects against nonalcoholic fatty liver disease in high-fat diet-fed mice. Sci. Transl. Med. 2016, 8, 357ra122. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′−3′) |
|---|---|
| Fxr forward | TGGGCTCCGAATCCTCTTAGA |
| Fxr reverse | TGGTCCTCAAATAAGATCCTTGG |
| Shp forward | CACCTGCATCTCACAGCCACT |
| Shp reverse | GCCAACCCAAGCAGGAAGA |
| Ntcp forward | ATGACCACCTGCTCCAGCTT |
| Ntcp reverse | GCCTTTGTAGGGCACCTTGT |
| Bsep forward | CTGCCAAGGATGCTAATGCA |
| Bsep reverse | CGATGGCTACCCTTTGCTTCT |
| Mrp3 forward | CTGGGTCCCCTGCATCTAC |
| Mrp3 reverse | GCCGTCTTGAGCCTGGATAAC |
| Mrp4 forward | CATCGCGGTAACCGTCCTC |
| Mrp4 reverse | CCGCAGTTTTACTCCGCAG |
| Oatp1 forward | GTGCATACCTAGCCAAATCACT |
| Oatp1 reverse | CCAGGCCCATAACCACACATC |
| Cyp7a1 forward | AACAACCTGCCAGTACTAGATAGC |
| Cyp7a1 reverse | TGTAGAGTGAAGTCCTCCTTAGC |
| Cyp8b1 forward | GGCTGGCTTCCTGAGCTTATT |
| Cyp8b1 reverse | ACTTCCTGAACAGCTCATCGG |
| Cyp27a1 forward | GCCTCACCTATGGGATCTTCA |
| Cyp27a1 reverse | TCAAAGCCTGACGCAGATG |
| Asbt forward | GGAACTGGCTCCAATATCCTG |
| Asbt reverse | GTTCCCGAGTCAACCCACAT |
| Ostα forward | GCCAGGCAGGACTCATATCAAA |
| Ostα reverse | GGCAACTGAGCCAGTGGTAAGA |
| Ostβ forward | CAGGAACTGCTGGAAGAAATGC |
| Ostβ reverse | GCAGGTCTTCTGGTGTTTCTTTGT |
| Gapdh forward | CGACTTCAACAGCAACTCCCACTCTTCC |
| Gapdh reverse | TGGGTGGTCCAGGGTTTCTTACTCCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, Z.; Wei, J.; Hou, X.; Jia, F.; Zhang, Z.; Guo, H.; Yuan, F.; He, F.; Ke, Z.; Wang, Y.; et al. A High Hepatic Uptake of Conjugated Bile Acids Promotes Colorectal Cancer—Associated Liver Metastasis. Cells 2022, 11, 3810. https://doi.org/10.3390/cells11233810
Zheng Z, Wei J, Hou X, Jia F, Zhang Z, Guo H, Yuan F, He F, Ke Z, Wang Y, et al. A High Hepatic Uptake of Conjugated Bile Acids Promotes Colorectal Cancer—Associated Liver Metastasis. Cells. 2022; 11(23):3810. https://doi.org/10.3390/cells11233810
Chicago/Turabian StyleZheng, Zongmei, Jiao Wei, Xinxin Hou, Fengjing Jia, Zhaozhou Zhang, Haidong Guo, Fuwen Yuan, Feng He, Zunji Ke, Yan Wang, and et al. 2022. "A High Hepatic Uptake of Conjugated Bile Acids Promotes Colorectal Cancer—Associated Liver Metastasis" Cells 11, no. 23: 3810. https://doi.org/10.3390/cells11233810
APA StyleZheng, Z., Wei, J., Hou, X., Jia, F., Zhang, Z., Guo, H., Yuan, F., He, F., Ke, Z., Wang, Y., & Zhao, L. (2022). A High Hepatic Uptake of Conjugated Bile Acids Promotes Colorectal Cancer—Associated Liver Metastasis. Cells, 11(23), 3810. https://doi.org/10.3390/cells11233810