
Rhizobia Contribute to Salinity Tolerance in Common Beans (Phaseolus vulgaris L.)

 , ,
, ,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Rhizobia Exposure to Salinity
2.2. Salinity Exposure to Common Beans and Co-Cultivation with Rhizobia
2.3. RNA Extraction, Sequencing, and Bioinformatics Analysis
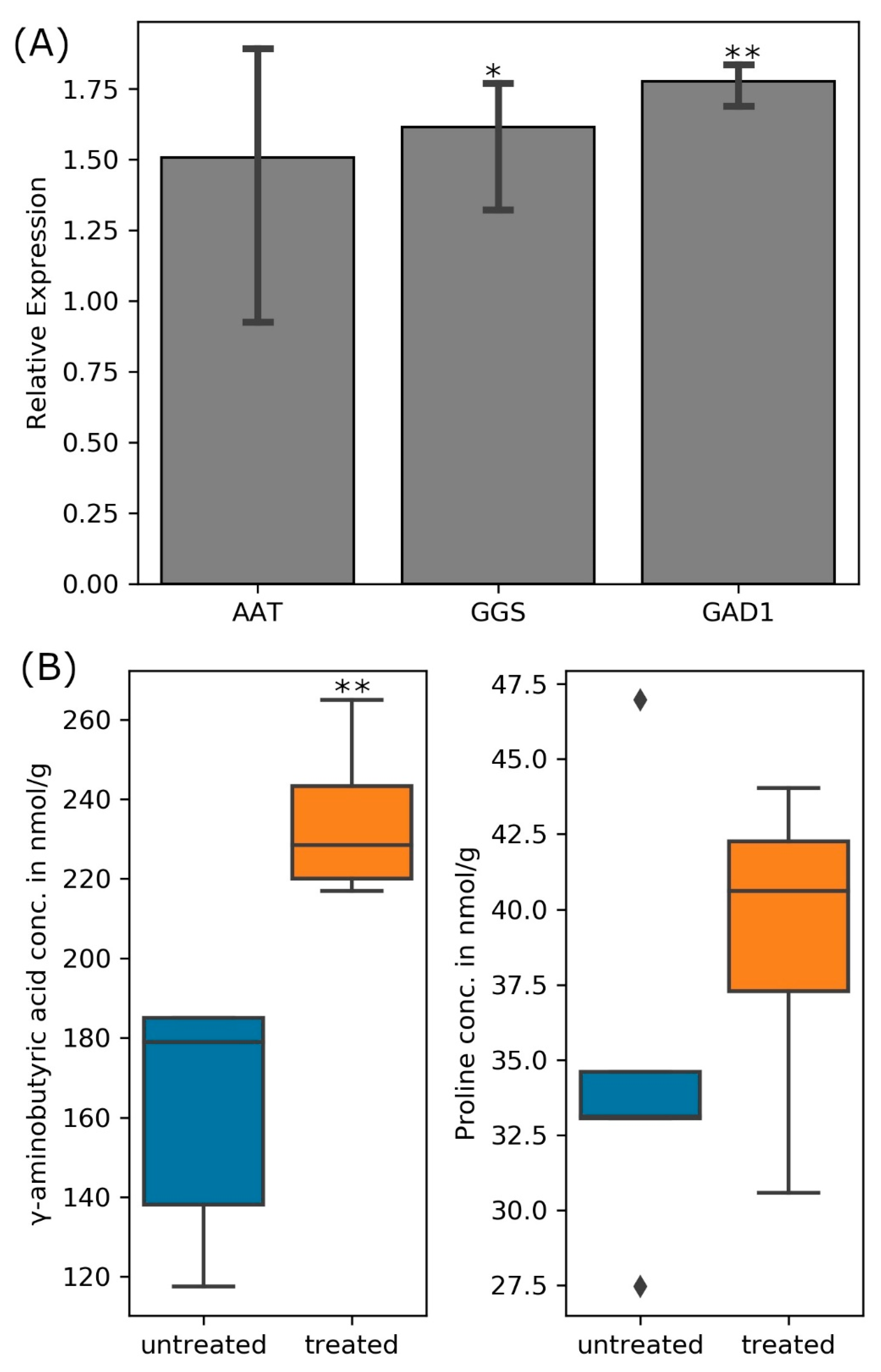
2.4. Quantitative Real-Time PCR (qPCR)
2.5. Quantification of γ-Aminobutyric Acid (GABA) and Amino Acids
3. Results and Discussion
3.1. Isolate S3 Is a Salt-Tolerant Rhizobia Strain
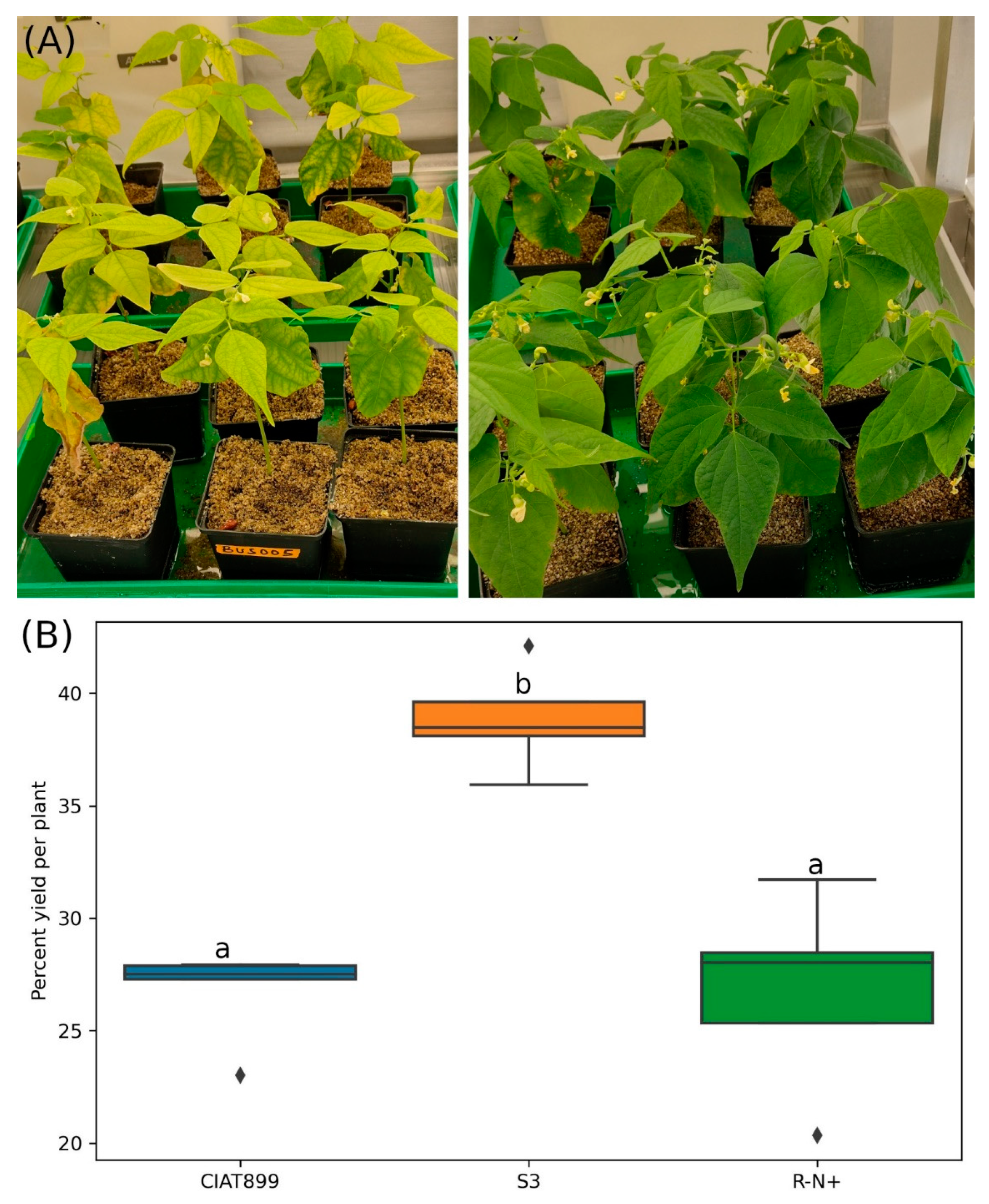
3.2. Isolate S3 Reduced the Effect of Salinity and Weight Loss in Common Beans
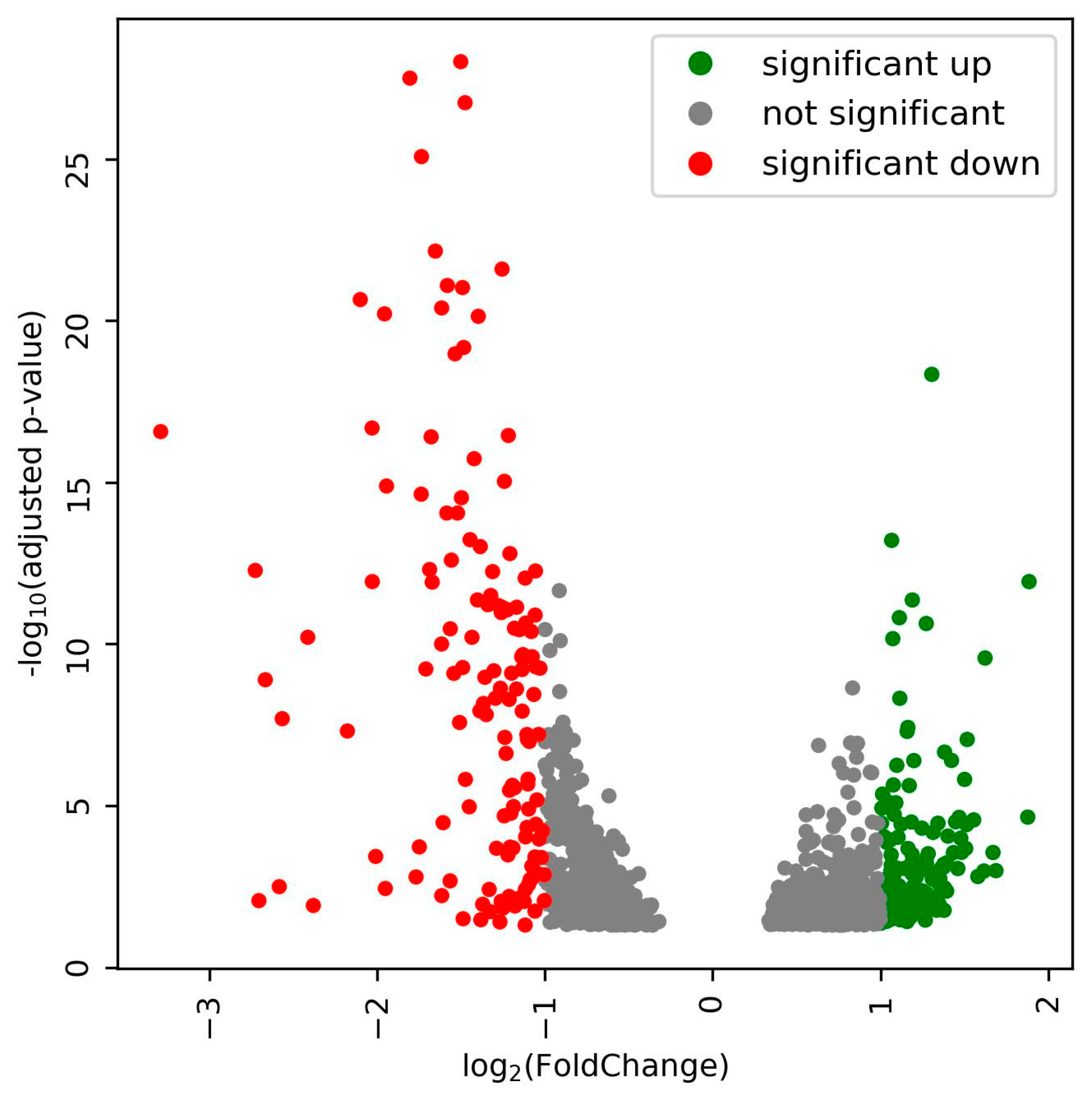
3.3. Rhizobia Gene Expression in the Nodules during Salinity Stress
3.4. Annotation of the Up-Regulated DEGs
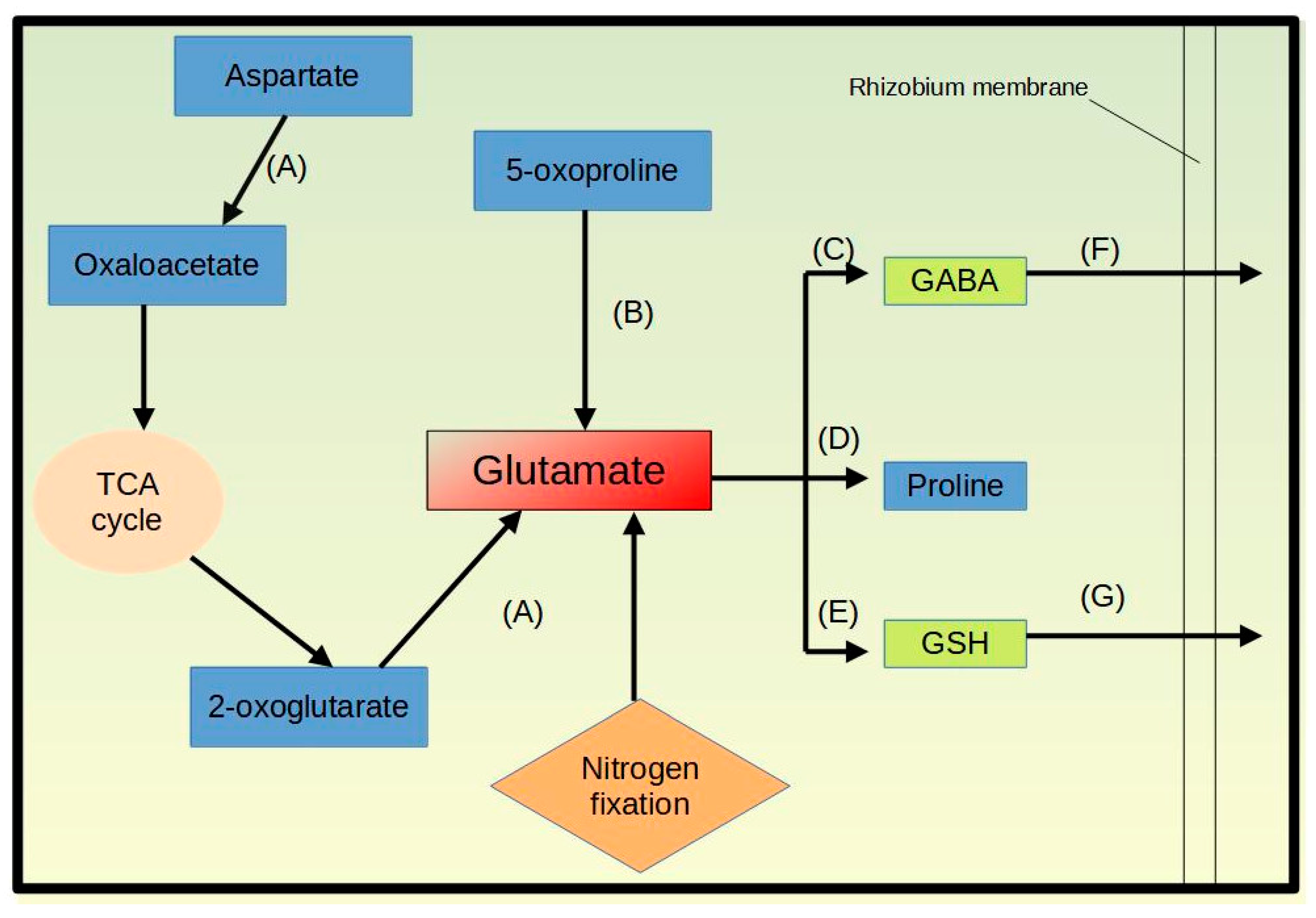
3.4.1. Amino Acid Transport and Metabolism
3.4.2. Inorganic Ion Transport and Metabolism
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant crop plants: Challenges and opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M. Gene expression profiling of plants under salt stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Kingsbury, R.W.; Epstein, E. Salt sensitivity in wheat: A case for specific ion toxicity. Plant Physiol. 1986, 80, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Volkov, V.; Beilby, M.J. Salinity tolerance in plants: Mechanisms and regulation of ion transport. Front. Plant Sci. 2017, 8, 1795. [Google Scholar] [CrossRef]
- Mugai, E.N. Salinity characterization of the Kenyan saline soils. Soil Sci. Plant Nutr. 2004, 50, 181–188. [Google Scholar] [CrossRef]
- Alhendawi, R.A.; Römheld, V.; Kirkby, E.A.; Marschner, H. Influence of increasing bicarbonate concentrations on plant growth, organic acid accumulation in roots and iron uptake by barley, sorghum, and maize. J. Plant Nutr. 1997, 20, 1731–1753. [Google Scholar] [CrossRef]
- Yang, X.; Römheld, V.; Marschner, H. Effect of bicarbonate on root growth and accumulation of organic acids in Zn-inefficient and Zn-efficient rice cultivars (Oryza sativa L.). Plant Soil 1994, 164, 1–7. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- de Oliveira, A.B.; Alencar, N.L.M.; Gomes-Filho, E. Comparison between the water and salt stress effects on plant growth and development. Responses Org. Water Stress 2013, 4, 67–94. [Google Scholar]
- Qadir, M.; Schubert, S. Degradation processes and nutrient constraints in sodic soils. Land Degrad. Dev. 2002, 13, 275–294. [Google Scholar] [CrossRef]
- Nawaz, K.; Hussain, K.; Majeed, A.; Khan, F.; Afghan, S.; Ali, K. Fatality of salt stress to plants: Morphological, physiological and biochemical aspects. Afr. J. Biotechnol. 2010, 9, 5475–5480. [Google Scholar]
- Jouyban, Z. The effects of salt stress on plant growth. Tech. J. Eng. Appl. Sci. 2012, 2, 7–10. [Google Scholar]
- Kochian, L.V.; Xin-Zhi, J.; Lucas, W.J. Potassium transport in corn roots: IV. Characterization of the linear component. Plant Physiol. 1985, 79, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.K.; Mahiwal, S. Role of Potassium in Plants; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Hassan, M.U.; Aamer, M.; Chattha, M.U.; Ullah, M.A.; Sulaman, S.; Nawaz, M.; Zhiqiang, W.; Yanqin, M.; Guoqin, H. The role of potassium in plants under drought stress: Mini review. J. Basic Appl. Sci. 2017, 13, 268–271. [Google Scholar] [CrossRef]
- Murillo-Amador, B.; Yamada, S.; Yamaguchi, T.; Rueda-Puente, E.; Ávila-Serrano, N.; García-Hernández, J.; López-Aguilar, R.; Troyo-Diéguez, E.; Nieto-Garibay, A. Influence of calcium silicate on growth, physiological parameters and mineral nutrition in two legume species under salt stress. J. Agron. Crop Sci. 2007, 193, 413–421. [Google Scholar] [CrossRef]
- Manishankar, P.; Wang, N.; Köster, P.; Alatar, A.A.; Kudla, J. Calcium signaling during salt stress and in the regulation of ion homeostasis. J. Exp. Bot. 2018, 69, 4215–4226. [Google Scholar] [CrossRef]
- Läuchli, A.; Grattan, S. Plant growth and development under salinity stress. In Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–32. [Google Scholar]
- Stoilova, T.; Pereira, G.; de Sousa, M. Morphological characterization of a small common bean (Phaseolus vulgaris L.) collection under different environments. J. Cent. Eur. Agric. 2013, 14, 1–11. [Google Scholar] [CrossRef]
- Ghassemi-Golezani, K.; Nikpour-Rashidabad, N.; Zehtab-Salmasi, S. Effect of salinity on yield and yield components of pinto bean cultivars. Int. J. Plant Anim. Environ. Sci. 2012, 2, 47–51. [Google Scholar]
- Lauchli, A. Salt Exclusion: An Adaptation of Legumes for Crops and Pastures under Saline Conditions; Wiley: Hoboken, NJ, USA, 1984. [Google Scholar]
- Cokkizgin, A. Salinity stress in common bean (Phaseolus vulgaris L.) seed germination. Not. Bot. Horti Agrobot. Cluj-Napoca 2012, 40, 177–182. [Google Scholar] [CrossRef]
- Faghire, M.; Bargaz, A.; Farissi, M.; Palma, F.; Mandri, B.; Lluch, C.; García, N.; Herrera-Cervera, J.; Oufdou, K.; Ghoulam, C. Effect of salinity on nodulation, nitrogen fixation and growth of common bean (Phaseolus vulgaris) inoculated with rhizobial strains isolated from the Haouz region of Morocco. Symbiosis 2011, 55, 69–75. [Google Scholar] [CrossRef]
- Delgado, M.; Ligero, F.; Lluch, C. Effects of salt stress on growth and nitrogen fixation by pea, faba-bean, common bean and soybean plants. Soil Biol. Biochem. 1994, 26, 371–376. [Google Scholar] [CrossRef]
- Serraj, R.; Vasquez-Diaz, H.; Drevon, J.-J. Effects of salt stress on nitrogen fixation, oxygen diffusion, and ion distribution in soybean, common bean, and alfalfa. J. Plant Nutr. 1998, 21, 475–488. [Google Scholar] [CrossRef]
- Gama, P.; Inanaga, S.; Tanaka, K.; Nakazawa, R. Physiological response of common bean (Phaseolus vulgaris L.) seedlings to salinity stress. Afr. J. Biotechnol. 2007, 6, 79–88. [Google Scholar]
- Fahmi, A.; Nagaty, H.; Eissa, R.; Hassan, M. Effects of salt stress on some nitrogen fixation parameters in faba bean. Pak. J. Biol. Sci. PJBS 2011, 14, 385–391. [Google Scholar] [CrossRef]
- Redondo, F.J.; Coba de la Peña, T.; Lucas, M.M.; Pueyo, J.J. Alfalfa nodules elicited by a flavodoxin-overexpressing Ensifer meliloti strain display nitrogen-fixing activity with enhanced tolerance to salinity stress. Planta 2012, 236, 1687–1700. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.; Keyster, M.; Ludidi, N. Response of soybean nodules to exogenously applied caffeic acid during NaCl-induced salinity. S. Afr. J. Bot. 2015, 96, 13–18. [Google Scholar] [CrossRef]
- Swaraj, K.; Bishnoi, N. Effect of salt stress on nodulation and nitrogen fixation in legumes. Indian J. Exp. Biol. 1999, 37, 843–848. [Google Scholar]
- Sombroek, W.G.; Braun, H.; Van der Pouw, B. Exploratory Soil Map and Agro-Climatic Zone Map of Kenya, 1980. Scale 1: 1,000,000; Kenya Soil Survey: Nairobi, Kenya, 1982. [Google Scholar]
- Ojwang, P.P.O.; Melis, R.; Songa, J.M.; Githiri, M.; Bett, C. Participatory plant breeding approach for host plant resistance to bean fly in common bean under semi-arid Kenya conditions. Euphytica 2009, 170, 383–393. [Google Scholar] [CrossRef]
- Katungi, E.; Farrow, A.; Mutuoki, T.; Gebeyehu, S.; Karanja, D.; Alamayehu, F.; Sperling, L.; Beebe, S.; Rubyogo, J.; Buruchara, R. Improving common bean productivity: An Analysis of socioeconomic factors in Ethiopia and Eastern Kenya. In Baseline Report Tropical Legumes II; Centro Internacional de Agricultura Tropical-CIAT: CIAT: Cali, Colombia, 2010; Volume 126. [Google Scholar]
- Zahir, Z.; Munir, A.; Asghar, H.; Shaharoona, B.; Arshad, M. Effectiveness of rhizobacteria containing ACC deaminase for growth promotion of peas (Pisum sativum) under drought conditions. J. Microbiol. Biotechnol. 2008, 18, 958–963. [Google Scholar]
- Lugtenberg, B.J.; Bloemberg, G.V. Microbe–plant interactions: Principles and mechanisms. Antonie Van Leeuwenhoek 2002, 81, 373–383. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Sridevi, M.; Mallaiah, K. Phosphate solubilization by Rhizobium strains. Indian J. Microbiol. 2009, 49, 98–102. [Google Scholar] [CrossRef]
- Guerinot, M. Iron uptake and metabolism in the rhizobia/legume symbioses. In Iron Nutrition and Interactions in Plants; Springer: Berlin/Heidelberg, Germany, 1991; pp. 239–249. [Google Scholar]
- Wekesa, C.S.; Furch, A.C.; Oelmüller, R. Isolation and characterization of high-efficiency rhizobia from Western Kenya nodulating with Common bean. Front. Microbiol. 2021, 12, 697567. [Google Scholar] [CrossRef]
- Wekesa, C.S. Isolation, Identification, and Characterization of Novel, Aluminium Tolerant Rhizobia Strains from Kenyan Soil. Ph.D. Thesis, Friedrich Schiller University Jena, Jena, Germany, 2022. [Google Scholar]
- Wekesa, C.; Muoma, J.O.; Reichelt, M.; Asudi, G.O.; Furch, A.C.; Oelmüller, R. The Cell Membrane of a Novel Rhizobium phaseoli Strain Is the Crucial Target for Aluminium Toxicity and Tolerance. Cells 2022, 11, 873. [Google Scholar] [CrossRef]
- Beal, J.; Farny, N.G.; Haddock-Angelli, T.; Selvarajah, V.; Baldwin, G.S.; Buckley-Taylor, R.; Gershater, M.; Kiga, D.; Marken, J.; Sanchania, V. Robust estimation of bacterial cell count from optical density. Commun. Biol. 2020, 3, 512. [Google Scholar] [CrossRef]
- Beck, D.P.; Materon, L.A.; Afandi, F. Practical Rhizobium-Legume Technology Manual; International Center for Agricultural Research in the Dry Areas (ICARDA): Aleppo, Syria, 1993. [Google Scholar]
- Andrews, S. FastQC: A Quality Control Tool for High throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 25 July 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Bedre, R. Bioinformatics Data Analysis and Visualization Toolkit. 2020, 6. Available online: https://github.com/reneshbedre/bioinfokit/tree/v0 (accessed on 21 August 2022).
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef]
- Leimbach, A. Bac-genomics-scripts: Bovine E. coli mastitis comparative genomics edition. Genome Announc. 2016, 4, 1–2. [Google Scholar]
- Kanehisa, M. The KEGG database. In Novartis Foundation Symposium; Wiley: Chichester, NY, USA, 2002; pp. 91–100. [Google Scholar]
- Moriya, Y.; Itoh, M.; Okuda, S.; Kanehisa, M. KAAS: KEGG automatic annotation server. Genome Inform. 2005, 5, 2005. [Google Scholar]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef]
- Zhang, J.D.; Ruschhaupt, M.; Biczok, R. ddCt method for qRT–PCR data analysis. Citeseer 2013, 48, 346–356. [Google Scholar]
- Scholz, S.S.; Reichelt, M.; Mekonnen, D.W.; Ludewig, F.; Mithöfer, A. Insect herbivory-elicited GABA accumulation in plants is a wound-induced, direct, systemic, and jasmonate-independent defense response. Front. Plant Sci. 2015, 6, 1128. [Google Scholar] [CrossRef]
- Miller, K.J.; Wood, J.M. Osmoadaptation by rhizosphere bacteria. Annu. Rev. Microbiol. 1996, 50, 101–137. [Google Scholar] [CrossRef]
- Soussi, M.; Santamaria, M.; Ocana, A.; Lluch, C. Effects of salinity on protein and lipopolysaccharide pattern in a salt-tolerant strain of Mesorhizobium ciceri. J. Appl. Microbiol. 2001, 90, 476–481. [Google Scholar] [CrossRef]
- Rady, M.M.; El-Shewy, A.A.; Seif El-Yazal, M.; Abdelaal, K.E. Response of salt-stressed common bean plant performances to foliar application of phosphorus (MAP). Int. Lett. Nat. Sci. 2018, 72. [Google Scholar] [CrossRef]
- Jebara, M.; Drevon, J.-J.; Aouani, M. Effects of hydroponic culture system and NaCl on interactions between common bean lines and native rhizobia from Tunisian soils. Agronomie 2001, 21, 601–605. [Google Scholar] [CrossRef]
- Estévez, J.; Dardanelli, M.; Megías, M.; Rodríguez-Navarro, D. Symbiotic performance of common bean and soybean co-inoculated with rhizobia and Chryseobacterium balustinum Aur9 under moderate saline conditions. Symbiosis 2009, 49, 29–36. [Google Scholar] [CrossRef]
- Cordovilla, M.; Ocana, A.; Ligero, F.; Lluch, C. Salinity effects on growth analysis and nutrient composition in four grain legumes-rhizobium symbiosis. J. Plant Nutr. 1995, 18, 1595–1609. [Google Scholar] [CrossRef]
- Chaiyen, P.; Tamanoi, F. Flavin-Dependent Enzymes: Mechanisms, Structures and Applications; Academic Press: Cambridge, MA, USA, 2020; pp. 1–566. [Google Scholar]
- Vinocur, B.; Altman, A. Recent advances in engineering plant tolerance to abiotic stress: Achievements and limitations. Curr. Opin. Biotechnol. 2005, 16, 123–132. [Google Scholar] [CrossRef]
- Pires, M.V.; Pereira Júnior, A.A.; Medeiros, D.B.; Daloso, D.M.; Pham, P.A.; Barros, K.A.; Engqvist, M.K.; Florian, A.; Krahnert, I.; Maurino, V.G. The influence of alternative pathways of respiration that utilize branched-chain amino acids following water shortage in Arabidopsis. Plant Cell Environ. 2016, 39, 1304–1319. [Google Scholar] [CrossRef]
- Shu, S.; Sun, J.; Guo, S.; Li, J.; Liu, C.; Wang, C. Effects of exogenous putrescine on PS II photochemistry and ion distribution of cucumber seedlings under salt stress. Acta Hortic. Sin. 2010, 37, 1065–1072. [Google Scholar]
- Sadak, M.S.; El-Hameid, A.; Asmaa, R.; Zaki, F.S.; Dawood, M.G.; El-Awadi, M.E. Physiological and biochemical responses of soybean (Glycine max L.) to cysteine application under sea salt stress. Bull. Natl. Res. Cent. 2020, 44, 1. [Google Scholar] [CrossRef]
- Hosie, A.H.; Allaway, D.; Galloway, C.; Dunsby, H.; Poole, P.S. Rhizobium leguminosarum has a second general amino acid permease with unusually broad substrate specificity and high similarity to branched-chain amino acid transporters (Bra/LIV) of the ABC family. J. Bacteriol. 2002, 184, 4071–4080. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, W.; Meng, Y.; Xie, T.; Li, L.; Li, J.; Wei, S. γ-Aminobutyric acid imparts partial protection from salt stress injury to maize seedlings by improving photosynthesis and upregulating osmoprotectants and antioxidants. Sci. Rep. 2017, 7, 43609. [Google Scholar] [CrossRef]
- Wu, X.; Jia, Q.; Ji, S.; Gong, B.; Li, J.; Lü, G.; Gao, H. Gamma-aminobutyric acid (GABA) alleviates salt damage in tomato by modulating Na+ uptake, the GAD gene, amino acid synthesis and reactive oxygen species metabolism. BMC Plant Biol. 2020, 20, 465. [Google Scholar] [CrossRef]
- Roxas, V.P.; Smith, R.K.; Allen, E.R.; Allen, R.D. Overexpression of glutathione S-transferase/glutathioneperoxidase enhances the growth of transgenic tobacco seedlings during stress. Nat. Biotechnol. 1997, 15, 988–991. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in plants: Biosynthesis and physiological role in environmental stress tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef]
- Martins, A.M.T.; Cordeiro, C.A.A.; Freire, A.M.J.P. In situ analysis of methylglyoxal metabolism in Saccharomyces cerevisiae. FEBS Lett. 2001, 499, 41–44. [Google Scholar] [CrossRef]
- Singla-Pareek, S.; Reddy, M.; Sopory, S. Genetic engineering of the glyoxalase pathway in tobacco leads to enhanced salinity tolerance. Proc. Natl. Acad. Sci. USA 2003, 100, 14672–14677. [Google Scholar] [CrossRef] [PubMed]
- Dey, G.; Banerjee, P.; Sharma, R.K.; Maity, J.P.; Etesami, H.; Shaw, A.K.; Huang, Y.-H.; Huang, H.-B.; Chen, C.-Y. Management of phosphorus in salinity-stressed agriculture for sustainable crop production by salt-tolerant phosphate-solubilizing bacteria—A review. Agronomy 2021, 11, 1552. [Google Scholar] [CrossRef]
- Grattan, S.; Grieve, C. Salinity–mineral nutrient relations in horticultural crops. Sci. Hortic. 1998, 78, 127–157. [Google Scholar] [CrossRef]
- Kageyama, H.; Tripathi, K.; Rai, A.K.; Cha-Um, S.; Waditee-Sirisattha, R.; Takabe, T. An alkaline phosphatase/phosphodiesterase, PhoD, induced by salt stress and secreted out of the cells of Aphanothece halophytica, a halotolerant cyanobacterium. Appl. Environ. Microbiol. 2011, 77, 5178–5183. [Google Scholar] [CrossRef] [PubMed]
- Umar, S.; Diva, I.; Anjum, N.A.; Iqbal, M.; Ahmad, I.; Pereira, E. Potassium-induced alleviation of salinity stress in Brassica campestris L. Cent. Eur. J. Biol. 2011, 6, 1054–1063. [Google Scholar] [CrossRef]
- Sikder, R.K.; Wang, X.; Zhang, H.; Gui, H.; Dong, Q.; Jin, D.; Song, M. Nitrogen enhances salt tolerance by modulating the antioxidant defense system and osmoregulation substance content in Gossypium hirsutum. Plants 2020, 9, 450. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, W.J.; Le, Q.T.; Hong, S.-W.; Lee, H. Growth performance can be increased under high nitrate and high salt stress through enhanced nitrate reductase activity in arabidopsis anthocyanin over-producing mutant plants. Front. Plant Sci. 2021, 12, 1115. [Google Scholar] [CrossRef]
- Kanwar, P.; Baby, D.; Bauer, P. Interconnection of iron and osmotic stress signalling in plants: Is FIT a regulatory hub to cross-connect abscisic acid responses? Plant Biol. 2021, 23, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Ma, J.F.; Yamaji, N.; Ueno, D.; Nomoto, K.; Iwashita, T. A specific transporter for iron (III)–phytosiderophore in barley roots. Plant J. 2006, 46, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Nozoye, T.; Nagasaka, S.; Kobayashi, T.; Takahashi, M.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 2011, 286, 5446–5454. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Geat, N.; Rajawat, M.V.S.; Prasanna, R.; Kar, A.; Singh, A.M.; Saxena, A.K. Prospecting endophytes from different Fe or Zn accumulating wheat genotypes for their influence as inoculants on plant growth, yield, and micronutrient content. Ann. Microbiol. 2018, 68, 815–833. [Google Scholar] [CrossRef]
- Ahmad, A.; Ismun, A.; Taib, M.; Ilias, M.A.; Ismail, A.; Othman, R.; Zainudin, S. Effects of salinity stress on carbohydrate metabolism in Cryptocoryne elliptica cultures. J. Trop. Plant Physiol. 2017, 9, 1–13. [Google Scholar]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H., Jr. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar] [CrossRef]
- Wu, D.; Cai, S.; Chen, M.; Ye, L.; Chen, Z.; Zhang, H.; Dai, F.; Wu, F.; Zhang, G. Tissue metabolic responses to salt stress in wild and cultivated barley. PLoS ONE 2013, 8, e55431. [Google Scholar] [CrossRef]
- Surówka, E.; Latowski, D.; Dziurka, M.; Rys, M.; Maksymowicz, A.; Żur, I.; Olchawa-Pajor, M.; Desel, C.; Krzewska, M.; Miszalski, Z. ROS-scavengers, osmoprotectants and violaxanthin de-epoxidation in salt-stressed Arabidopsis thaliana with different tocopherol composition. Int. J. Mol. Sci. 2021, 22, 11370. [Google Scholar] [CrossRef]
- Kaplan, F.; Sung, D.Y.; Guy, C.L. Roles of β-amylase and starch breakdown during temperatures stress. Physiol. Plant. 2006, 126, 120–128. [Google Scholar] [CrossRef]
- Purdy, S.J.; Bussell, J.D.; Nunn, C.P.; Smith, S.M. Leaves of the Arabidopsis maltose exporter1 mutant exhibit a metabolic profile with features of cold acclimation in the warm. PLoS ONE 2013, 8, e79412. [Google Scholar] [CrossRef]
- Rautengarten, C.; Birdseye, D.; Pattathil, S.; McFarlane, H.E.; Saez-Aguayo, S.; Orellana, A.; Persson, S.; Hahn, M.G.; Scheller, H.V.; Heazlewood, J.L. The elaborate route for UDP-arabinose delivery into the Golgi of plants. Proc. Natl. Acad. Sci. USA 2017, 114, 4261–4266. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zayed, O.; Zeng, F.; Liu, C.; Zhang, L.; Zhu, P.; Hsu, C.C.; Tuncil, Y.E.; Tao, W.A.; Carpita, N.C. Arabinose biosynthesis is critical for salt stress tolerance in Arabidopsis. New Phytol. 2019, 224, 274–290. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Q1 | Q3 | RT (min) | Internal Standard | IS Q1 | IS Q3 | DP | CE |
|---|---|---|---|---|---|---|---|---|
| GABA | 104.1 | 87.1 | 0.5 | 13C,15N-Ala | ||||
| Proline | 116.1 | 70.0 | 0.5 | 13C,15N-Pro | 122.1 | 75.0 | 20 | 19 |
| Valine | 118.1 | 72.2 | 0.5 | 13C,15N-Val | 124.1 | 77.2 | 20 | 13 |
| Leucine | 132.2 | 86.1 | 1.3 | 13C,15N-Leu | 139.2 | 92.1 | 20 | 13 |
| Isoleucine | 132.2 | 86.1 | 1.1 | 13C,15N-Ile | 139.2 | 92.1 | 20 | 13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wekesa, C.; Asudi, G.O.; Okoth, P.; Reichelt, M.; Muoma, J.O.; Furch, A.C.U.; Oelmüller, R. Rhizobia Contribute to Salinity Tolerance in Common Beans (Phaseolus vulgaris L.). Cells 2022, 11, 3628. https://doi.org/10.3390/cells11223628
Wekesa C, Asudi GO, Okoth P, Reichelt M, Muoma JO, Furch ACU, Oelmüller R. Rhizobia Contribute to Salinity Tolerance in Common Beans (Phaseolus vulgaris L.). Cells. 2022; 11(22):3628. https://doi.org/10.3390/cells11223628
Chicago/Turabian StyleWekesa, Clabe, George O. Asudi, Patrick Okoth, Michael Reichelt, John O. Muoma, Alexandra C. U. Furch, and Ralf Oelmüller. 2022. "Rhizobia Contribute to Salinity Tolerance in Common Beans (Phaseolus vulgaris L.)" Cells 11, no. 22: 3628. https://doi.org/10.3390/cells11223628
APA StyleWekesa, C., Asudi, G. O., Okoth, P., Reichelt, M., Muoma, J. O., Furch, A. C. U., & Oelmüller, R. (2022). Rhizobia Contribute to Salinity Tolerance in Common Beans (Phaseolus vulgaris L.). Cells, 11(22), 3628. https://doi.org/10.3390/cells11223628