
Molecular Defense Response of Pine Trees (Pinus spp.) to the Parasitic Nematode Bursaphelenchus xylophilus



Abstract
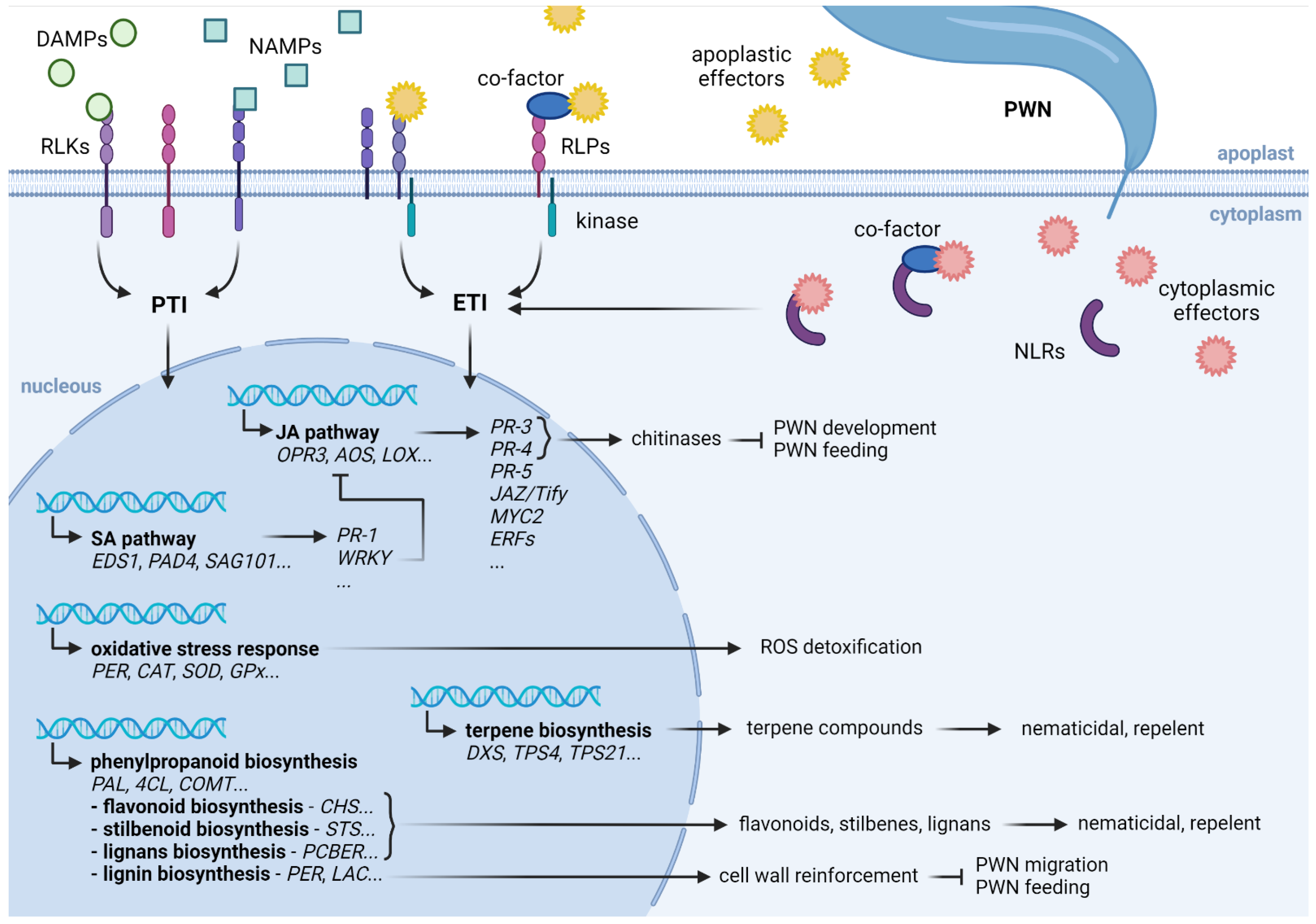
1. Introduction
2. Defense Mechanisms Highlighted by the Transcriptional Response to PWN
2.1. Phytohormone Signaling
2.2. Secondary Metabolism
2.2.1. Biosynthesis of Terpene Compounds

{kind=link}
| Secondary Metabolite | Type of Compound | Species of Origin | Effect on PWN | References |
|---|---|---|---|---|
| α-humulene | Sesquiterpene | Pma | repellent | [64] |
| Calarene | Sesquiterpene | Pma | repellent | [64] |
| β-bisabolene | Sesquiterpene | Pma | repellent | [64] |
| Dihydroabietane | Diterpene | Pma | repellent | [64] |
| α-pinene | Monoterpene | Pma | repellent | [64] |
| Pma | nematicide | [63] | ||
| β-pinene | Monoterpene | Pma | nematicide | [63] |
| β-myrcene | Monoterpene | Pma | nematicide | [63] |
| D-limonene | Monoterpene | Pma | nematicide | [63] |
| Longifolene | Sesquiterpene | Pma | nematicide | [63] |
| Pinosylvin monomethyl ether (PME) | Stilbene | Pma, Pst, Ppa | nematicide | [64] |
| Pst | nematicide | [66] | ||
| Dihydropinosylvin monomethyl ether (DPME) | Stilbene | - | nematicide | [64] |
| Pst | nematicide | [66] | ||
| Pst | nematicide | [67] | ||
| Pinosylvin | Stilbene | - | nematicide | [64] |
| Methyl ferulate | Phenolic compound | Pma | nematicide | [64] |
| Ferulic acid | Phenolic acid | - | nematicide | [64] |
| (-)-nortrachelogenin | Lignan | Pma | nematicide | [64] |
| (+)-pinoresinol | Lignan | Pma | nematicide | [64] |
2.2.2. Phenylpropanoids Biosynthesis
2.2.3. Lignin Biosynthesis and Cell Wall Reinforcement
2.3. Oxidative Stress Response
2.4. Plant Defense Response Genes
2.5. Resistance Genes
3. Post-Transcriptional Regulation Mediated by Small RNAs in Pine Response to PWN
3.1. MicroRNA-Mediated Response to PWN
3.2. Trans-Kingdom RNA Silencing in PWD
4. Defense Response Induced by the Application of Phytohormones and Secondary Metabolites
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jones, J.T.; Moens, M.; Mota, M.; Li, H.; Kikuchi, T. Bursaphelenchus xylophilus: Opportunities in Comparative Genomics and Molecular Host-Parasite Interactions. Mol. Plant Pathol. 2008, 9, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.N.; Kim, J.H.; Ahn, J.Y.; Kim, S.; Cho, B.K.; Kim, Y.H.; Min, J. A Short Review of the Pinewood Nematode, Bursaphelenchus xylophilus. Toxicol. Environ. Health Sci. 2020, 12, 297–304. [Google Scholar] [CrossRef]
- Webster, J.; Mota, M. PineWilt Disease: Global Issues, Trade and Economic Impact. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M., Vieira, P., Eds.; Springer: Berlin, Germany, 2008; pp. 1–4. [Google Scholar]
- Futai, K. Pine Wilt in Japan: From First Incidence to the Present. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 5–10. [Google Scholar]
- Mota, M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First Report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef]
- Zhao, B.G. Pine Wilt Disease in China. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 18–25. ISBN 978-4-431-75655-2. [Google Scholar]
- Sniezko, R.A.; Koch, J. Breeding Trees Resistant to Insects and Diseases: Putting Theory into Application. Biol. Invasions 2017, 19, 3377–3400. [Google Scholar] [CrossRef]
- Naidoo, S.; Slippers, B.; Plett, J.M.; Coles, D.; Oates, C.N. The Road to Resistance in Forest Trees. Front. Plant Sci. 2019, 10, 273. [Google Scholar] [CrossRef]
- Nose, M.; Shiraishi, S. Breeding for Resistance to Pine Wilt Disease. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 334–350. [Google Scholar]
- Carrasquinho, I.; Lisboa, A.; Inácio, M.L.; Gonçalves, E. Genetic Variation in Susceptibility to Pine Wilt Disease of Maritime Pine Half-Sib Families. Ann. For. Sci. 2018, 75, 85. [Google Scholar] [CrossRef]
- Menéndez-Gutiérrez, M.; Alonso, M.; Toval, G.; Díaz, R. Testing of Selected Pinus pinaster Half-Sib Families for Tolerance to Pinewood Nematode (Bursaphelenchus xylophilus). Forestry 2017, 91, 38–48. [Google Scholar] [CrossRef]
- Toda, T.; Kurinobu, S. Realized Genetic Gains Observed in Progeny Tolerance of Selected Red Pine (Pinus densiflora) and Black Pine (P. thunbergii) to Pine Wilt Disease. Silvae Genet. 2002, 51, 42–44. [Google Scholar]
- Xu, L.-Y.; Zhang, J.; Gao, J.-B.; Chen, X.-L.; Jiang, C.-W.; Hao, Y.-P. Study on the Disease Resistance of Candidate Clones in Pinus massoniana to Bursaphelenchus xylophilus. China For. Sci. Technol. 2012, 26, 27–30. [Google Scholar]
- Ribeiro, B.; Espada, M.; Vu, T.; Nóbrega, F.; Mota, M.; Carrasquinho, I. Pine Wilt Disease: Detection of the Pinewood Nematode (Bursaphelenchus xylophilus) as a Tool for a Pine Breeding Programme. For. Pathol. 2012, 42, 521–525. [Google Scholar] [CrossRef]
- Menéndez-Gutiérrez, M.; Villar, L.; Díaz, R. Virulence of Seven Pathogenic Bursaphelenchus xylophilus Isolates in Pinus pinaster and Pinus radiata Seedlings and Its Relation with Multiplication. For. Pathol. 2021, 51, e12677. [Google Scholar] [CrossRef]
- Aikawa, T.; Kikuchi, T. Estimation of Virulence of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) Based on Its Reproductive Ability. Nematology 2007, 9, 371–377. [Google Scholar] [CrossRef]
- Akiba, M.; Ishihara, M.; Sahashi, N.; Nakamura, K.; Ohira, M.; Toda, T. Virulence of Bursaphelenchus xylophilus Isolated from Naturally Infested Pine Forests to Five Resistant Families of Pinus thunbergii. Plant Dis. 2012, 96, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Ye, J.; Lin, S.; Wu, X.; Li, D.; Nian, B. Deciphering the Molecular Variations of Pine Wood Nematode Bursaphelenchus xylophilus with Different Virulence. PLoS ONE 2016, 11, e0156040. [Google Scholar] [CrossRef] [PubMed]
- Palomares-Rius, J.E.; Tsai, I.J.; Karim, N.; Akiba, M.; Kato, T.; Maruyama, H.; Takeuchi, Y.; Kikuchi, T. Genome-Wide Variation in the Pinewood Nematode Bursaphelenchus xylophilus and Its Relationship with Pathogenic Traits. BMC Genom. 2015, 16, 845. [Google Scholar] [CrossRef] [PubMed]
- Mota, M.M.; Takemoto, S.; Takeuchi, Y.; Hara, N.; Futai, K. Comparative Studies between Portuguese and Japanese Isolates of the Pinewood Nematode, Bursaphelenchus xylophilus. J. Nematol. 2006, 38, 429–433. [Google Scholar]
- Valadas, V.; Laranjo, M.; Mota, M.; Oliveira, S. Molecular Characterization of Portuguese Populations of the Pinewood Nematode Bursaphelenchus xylophilus Using Cytochrome b and Cellulase Genes. J. Helminthol. 2012, 87, 457–466. [Google Scholar] [CrossRef]
- Zipfel, C. Plant Pattern-Recognition Receptors. Trends Immunol. 2014, 35, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Gillet, F.X.; Bournaud, C.; De Souza, J.D.A.; Grossi-De-Sa, M.F. Plant-Parasitic Nematodes: Towards Understanding Molecular Players in Stress Responses. Ann. Bot. 2017, 119, 775–789. [Google Scholar] [CrossRef]
- Couto, D.; Zipfel, C. Regulation of Pattern Recognition Receptor Signaling in Plants. Nat. Rev. Immunol. 2016, 16, 537–552. [Google Scholar] [CrossRef]
- Holbein, J.; Grundler, F.M.W.; Siddique, S. Plant Basal Resistance to Nematodes: An Update. J. Exp. Bot. 2016, 67, erw005. [Google Scholar] [CrossRef]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling Mechanisms in Pattern-Triggered Immunity (PTI). Mol. Plant 2015, 8, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-Triggered Immunity: From Pathogen Perception to Robust Defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Kadota, Y.; Shirasu, K. Plant Immune Responses to Parasitic Nematodes. Front. Plant Sci. 2019, 10, 1165. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Putker, V.; Goverse, A. Molecular and Cellular Mechanisms Involved in Host-Specific Resistance to Cyst Nematodes in Crops. Front. Plant Sci. 2021, 12, 641582. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Bos, J.I.B. Effector Proteins That Modulate Plant-Insect Interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Erb, M.; Reymond, P. Molecular Interactions between Plants and Insect Herbivores. Annu. Rev. Plant Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef]
- Dodds, P.N.; Rathjen, J.P. Plant Immunity: Towards an Integrated View of Plant–Pathogen Interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef]
- Trudgill, D.L. Resistance to and Tolerance of Plant Parasitic Nematodes in Plants. Annu. Rev. Phytopathol. 1991, 29, 167–192. [Google Scholar] [CrossRef]
- Hirao, T.; Fukatsu, E.; Watanabe, A. Characterization of Resistance to Pine Wood Nematode Infection in Pinus thunbergii Using Suppression Subtractive Hybridization. BMC Plant Biol. 2012, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Nose, M.; Shiraishi, S. Comparison of the Gene Expression Profiles of Resistant and Non-Resistant Japanese Black Pine Inoculated with Pine Wood Nematode Using a Modified LongSAGE Technique. For. Pathol. 2011, 41, 143–155. [Google Scholar] [CrossRef]
- Liu, Q.; Wei, Y.; Xu, L.; Hao, Y.; Chen, X.; Zhou, Z. Transcriptomic Profiling Reveals Differentially Expressed Genes Associated with Pine Wood Nematode Resistance in Masson Pine (Pinus massoniana Lamb.). Sci. Rep. 2017, 7, 4693. [Google Scholar] [CrossRef] [PubMed]
- Modesto, I.; Sterck, L.; Arbona, V.; Gómez-Cadenas, A.; Carrasquinho, I.; Van de Peer, Y.; Miguel, C.M. Insights Into the Mechanisms Implicated in Pinus pinaster Resistance to Pinewood Nematode. Front. Plant Sci. 2021, 12, 690857. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, R.; Li, D.; Wang, F. Transcriptomic and Coexpression Network Analyses Revealed Pine Chalcone Synthase Genes Associated with Pine Wood Nematode Infection. Int. J. Mol. Sci. 2021, 22, 11195. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Liu, Z.Y.; Zhang, K.; Lu, Q.; Liang, J.; Zhang, X.Y. Characterization of the Pinus massoniana Transcriptional Response to Bursaphelenchus xylophilus Infection Using Suppression Subtractive Hybridization. Int. J. Mol. Sci. 2013, 14, 11356–11375. [Google Scholar] [CrossRef]
- Xie, W.; Liang, G.; Huang, A.; Zhang, F.; Guo, W. Comparative Study on the MRNA Expression of Pinus massoniana Infected by Bursaphelenchus xylophilus. J. For. Res. 2020, 31, 75–86. [Google Scholar] [CrossRef]
- Zheng, H.; Xu, M.; Xu, F.; Ye, J. A Comparative Proteomics Analysis of Pinus massoniana Inoculated with Bursaphelenchus xylophilus. Pak. J. Bot. 2015, 47, 1271–1280. [Google Scholar]
- Gao, J.; Pan, T.; Chen, X.; Wei, Q.; Xu, L. Proteomic Analysis of Masson Pine with High Resistance to Pine Wood Nematodes. PLoS ONE 2022, 17, e0273010. [Google Scholar] [CrossRef]
- Shin, H.; Lee, H.; Woo, K.S.; Noh, E.W.; Koo, Y.B.; Lee, K.J. Identification of Genes Upregulated by Pinewood Nematode Inoculation in Japanese Red Pine. Tree Physiol. 2009, 29, 411–421. [Google Scholar] [CrossRef]
- Lee, I.H.; Han, H.; Koh, Y.H.; Kim, I.S.; Lee, S.W.; Shim, D. Comparative Transcriptome Analysis of Pinus densiflora Following Inoculation with Pathogenic (Bursaphelenchus xylophilus) or Non-Pathogenic Nematodes (B. thailandae). Sci. Rep. 2019, 9, 12180. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, D.; Trindade, C.; Usié, A.; Meireles, B.; Barbosa, P.; Fortes, A.; Pesquita, C.; Costa, R.; Ramos, A. Expression Profiling in Pinus pinaster in Response to Infection with the Pine Wood Nematode Bursaphelenchus xylophilus. Forests 2017, 8, 279. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Carrasquinho, I.; António, C. Primary Metabolite Adjustments Associated With Pinewood Nematode Resistance in Pinus pinaster. Front. Plant Sci. 2021, 12, 777681. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.S.; Pinheiro, M.; Silva, A.I.; Egas, C.; Vasconcelos, M.W. Searching for Resistance Genes to Bursaphelenchus xylophilus Using High Throughput Screening. BMC Genom. 2012, 13, 599. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, D.; Trindade, C.; Usié, A.; Meireles, B.; Fortes, A.M.; Guimarães, J.B.; Simões, F.; Costa, R.L.; Ramos, A.M. Comparative Transcriptomic Response of Two Pinus Species to Infection with the Pine Wood Nematode Bursaphelenchus xylophilus. Forests 2020, 11, 204. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2011, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Caarls, L.; Pieterse, C.M.J.; Wees, S.C.M. Van How Salicylic Acid Takes Transcriptional Control over Jasmonic Acid Signaling. Front. Palnt Sci. 2015, 6, 170. [Google Scholar] [CrossRef]
- Campos, M.L.; Kang, J.H.; Howe, G.A. Jasmonate-Triggered Plant Immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef]
- De Vleesschauwer, D.; Xu, J.; Höfte, M. Making Sense of Hormone-Mediated Defense Networking: From Rice to Arabidopsis. Front. Plant Sci. 2014, 5, 611. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Langer, S.; Carrasquinho, I.; Bergström, E.; Larson, T.; Thomas-Oates, J.; António, C. Pinus pinaster Early Hormonal Defense Responses to Pinewood Nematode (Bursaphelenchus xylophilus) Infection. Metabolites 2021, 11, 227. [Google Scholar] [CrossRef]
- Vos, I.A.; Verhage, A.; Watt, L.G.; Vlaardingerbroek, I.; Schuurink, R.C.; Pieterse, C.M.J.; van Wees, S.C.M. Abscisic Acid Is Essential for Rewiring of Jasmonic Acid-Dependent Defenses during Herbivory. bioRxiv 2019, 1–41. [Google Scholar] [CrossRef]
- Ton, J.; Flors, V.; Mauch-Mani, B. The Multifaceted Role of ABA in Disease Resistance. Trends Plant Sci. 2009, 14, 310–317. [Google Scholar] [CrossRef]
- Nahar, K.; Kyndt, T.; Nzogela, Y.B.; Gheysen, G. Abscisic Acid Interacts Antagonistically with Classical Defense Pathways in Rice—Migratory Nematode Interaction. New Phytol. 2012, 196, 901–913. [Google Scholar] [CrossRef]
- Fukuda, K.; Hogetsu, T.; Suzuki, K. Ethylene Production during Symptom Development of Pine-wilt Disease. Eur. J. For. Pathol. 1994, 24, 193–202. [Google Scholar] [CrossRef]
- Fukuda, K. Physiological Process of the Symptom Development and Resistance Mechanism in Pine Wilt Disease. J. For. Res. 1997, 2, 171–181. [Google Scholar] [CrossRef]
- Keeling, C.I.; Bohlmann, J. Genes, Enzymes and Chemicals of Terpenoid Diversity in the Constitutive and Induced Defense of Conifers against Insects and Pathogens. New Phytol. 2006, 170, 657–675. [Google Scholar] [CrossRef] [PubMed]
- Eyles, A.; Bonello, P.; Ganley, R.; Mohammed, C. Induced Resistance to Pests and Pathogens in Trees. New Phytol. 2010, 185, 893–908. [Google Scholar] [CrossRef]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in Defense against Pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, Q.; Zhou, Z.; Yin, H.; Xie, Y.; Wei, Y. Two Terpene Synthases in Resistant Pinus massoniana Contribute to Defense against Bursaphelenchus xylophilus. Plant Cell Environ. 2020, 44, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Suga, T.; Ohta, S.; Munesada, K.; Ide, N.; Kurokawa, M.; Shimizu, M.; Ohta, E. Endogenous Pine Wood Nematicidal Substances in Pines, Pinus massoniana, P. strobus and P. palustris. Phytochemistry 1993, 33, 1395–1401. [Google Scholar] [CrossRef]
- Trindade, H.; Sena, I.; Figueiredo, A.C. Characterization of α-Pinene Synthase Gene in Pinus pinaster and P. pinea in Vitro Cultures and Differential Gene Expression Following Bursaphelenchus xylophilus Inoculation. Acta Physiol. Plant. 2016, 38, 143. [Google Scholar] [CrossRef]
- Hwang, H.S.; Han, J.Y.; Choi, Y.E. Enhanced Accumulation of Pinosylvin Stilbenes and Related Gene Expression in Pinus strobus after Infection of Pine Wood Nematode. Tree Physiol. 2021, 41, 1972–1987. [Google Scholar] [CrossRef] [PubMed]
- Hanawa, F.; Yamada, T.; Nakashima, T. Phytoalexins from Pinus strobus Bark Infected with Pinewood Nematode, Bursaphelenchus xylophilus. Phytochemistry 2001, 57, 223–228. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Mendes, M.D.; Lima, A.S.; Barbosa, P.M.; Ascens, L.; Pedro, L.G.; Mota, M.M.; Figueiredo, A.C. Pinus halepensis, Pinus pinaster, Pinus pinea and Pinus sylvestris Essential Oils Chemotypes and Monoterpene Hydrocarbon Enantiomers, before and after Inoculation with the Pinewood Nematode Bursaphelenchus xylophilus. Chem. Biodivers. 2017, 14, e1600153. [Google Scholar] [CrossRef] [PubMed]
- da Silva, M.N.; Solla, A.; Sampedro, L.; Zas, R.; Vasconcelos, M.W. Susceptibility to the Pinewood Nematode (PWN) of Four Pine Species Involved in Potential Range Expansion across Europe. Tree Physiol. 2015, 35, 987–999. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, E.; Figueiredo, A.C.; Barroso, J.G.; Henriques, J.; Sousa, E.; Bonifácio, L. Effect of Monochamus galloprovincialis Feeding on Pinus pinaster and Pinus pinea, Oleoresin and Insect Volatiles. Phytochemistry 2020, 169, 112159. [Google Scholar] [CrossRef] [PubMed]
- Vogt, T. Phenylpropanoid Biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Chin, S.; Behm, C.; Mathesius, U. Functions of Flavonoids in Plant–Nematode Interactions. Plants 2018, 7, 85. [Google Scholar] [CrossRef]
- Pimentel, C.S.; Gonçalves, E.V.; Firmino, P.N.; Calvão, T.; Fonseca, L.; Abrantes, I.; Correia, O.; Máguas, C. Differences in Pine Species Constitutive and Inducible Defenses Determining the Susceptibility to the Pinewood Nematode. Plant Pathol. 2016, 66, 131–139. [Google Scholar] [CrossRef]
- Trindade, S.; Canas, S.; In, M.L.; Pereira-lorenzo, S.; Sousa, E.; Naves, P. Phenolic Compounds Regulating the Susceptibility of Adult Pine Species to Bursaphelenchus xylophilus. Forests 2022, 13, 500. [Google Scholar] [CrossRef]
- Kuroda, H.; Goto, S.; Kazumi, E.; Kuroda, K. The Expressed Genes of Japanese Red Pine (Pinus densiflora) Involved in the Pine Wilt Disease Severity. BMC Proc. 2011, 5, P92. [Google Scholar] [CrossRef]
- Wuyts, N.; Swennen, R.; De Waele, D. Effects of Plant Phenylpropanoid Pathway Products and Selected Terpenoids and Alkaloids on the Behaviour of the Plant-Parasitic Nematodes Radopholus similis, Pratylenchus penetrans and Meloidogyne incognita. Nematology 2006, 8, 89–101. [Google Scholar] [CrossRef]
- Kusumoto, D.; Yonemichi, T.; Inoue, H.; Hirao, T.; Watanabe, A.; Yamada, T. Comparison of Histological Responses and Tissue Damage Expansion between Resistant and Susceptible Pinus thunbergii Infected with Pine Wood Nematode Bursaphelenchus xylophilus. J. For. Res. 2014, 19, 285–294. [Google Scholar] [CrossRef]
- Nairn, C.J.; Lennon, D.M.; Wood-Jones, A.; Nairn, A.V.; Dean, J.F.D. Carbohydrate-Related Genes and Cell Wall Biosynthesis in Vascular Tissues of Loblolly Pine (Pinus taeda). Tree Physiol. 2008, 28, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C. Occurrence and Properties of Plant Pathogenesis-Related Proteins. In Pathogenesis-Related Proteins in Plants; Datta, S.K., Muthukrishman, S., Eds.; CRC Press LLC: Boca Raton, FL, USA, 1999; ISBN 0849306973. [Google Scholar]
- Fukushige, H.; Futai, K. Characteristics of Egg Shells and the Morphology of Female Tail-Tips of Bursaphelenchus xylophilus, B. mucronatus and Some Strains of Related Species from France. Jpn. J. Nematol. 1985, 15, 49–54. [Google Scholar]
- Mercer, C.F.; Greenwood, D.R.; Grant, J.L. Effect of Plant and Microbial Chitinases on the Eggs and Juveniles of Meloidogyne hapla Chitwood (Nematoda: Tylenchida). Nematologica 1992, 38, 227–236. [Google Scholar] [CrossRef]
- Vandenborre, G.; Smagghe, G.; Van Damme, E.J.M. Plant Lectins as Defense Proteins against Phytophagous Insects. Phytochemistry 2011, 72, 1538–1550. [Google Scholar] [CrossRef]
- Ripoll, C.; Favery, B.; Lecomte, P.; Van Damme, E.; Peumans, W.; Abad, P.; Jouanin, L. Evaluation of the Ability of Lectin from Snowdrop (Galanthus nivalis) to Protect Plants against Root-Knot Nematodes. Plant Sci. 2003, 164, 517–523. [Google Scholar] [CrossRef]
- Kang, J.S.; Koh, Y.H.; Moon, Y.S.; Lee, S.H. Molecular Properties of a Venom Allergen-like Protein Suggest a Parasitic Function in the Pinewood Nematode Bursaphelenchus xylophilus. Int. J. Parasitol. 2012, 42, 63–70. [Google Scholar] [CrossRef]
- Hu, L.J.; Wu, X.Q.; Li, H.Y.; Zhao, Q.; Wang, Y.C.; Ye, J.R. An Effector, BxSapB1, Induces Cell Death and Contributes to Virulence in the Pine Wood Nematode Bursaphelenchus xylophilus. Mol. Plant-Microbe Interact. 2019, 32, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.J.; Wu, X.Q.; Li, H.Y.; Wang, Y.C.; Huang, X.; Wang, Y.; Li, Y. BxCDP1 from the Pine Wood Nematode Bursaphelenchus xylophilus Is Recognized as a Novel Molecular Pattern. Mol. Plant Pathol. 2020, 21, 923–935. [Google Scholar] [CrossRef]
- Milligan, S.B.; Bodeau, J.; Yaghoobi, J.; Kaloshian, I.; Zabel, P.; Williamson, V.M. The Root Knot Nematode Resistance Gene Mi from Tomato Is a Member of the Leucine Zipper, Nucleotide Binding, Leucine-Rich Repeat Family of Plant Genes. Plant Cell 1998, 10, 1307–1319. [Google Scholar] [CrossRef]
- Rossi, M.; Goggin, F.L.; Milligan, S.B.; Kaloshian, I.; Ullman, D.E.; Williamson, V.M. The Nematode Resistance Gene Mi of Tomato Confers Resistance against the Potato Aphid. Proc. Natl. Acad. Sci. USA. 1998, 95, 9750–9754. [Google Scholar] [CrossRef]
- Nombela, G.; Williamson, V.M.; Muñiz, M. The Root-Knot Nematode Resistance Gene Mi-1.2 of Tomato Is Responsible for Resistance against the Whitefly Bemisia Tabaci. Mol. Plant-Microbe Interact. 2003, 16, 645–649. [Google Scholar] [CrossRef]
- Casteel, C.L.; Walling, L.L.; Paine, T.D. Behavior and Biology of the Tomato Psyllid, Bactericerca Cockerelli, in Response to the Mi-1.2 Gene. Entomol. Exp. Appl. 2006, 121, 67–72. [Google Scholar] [CrossRef]
- Tang, Y.; Yan, X.; Gu, C.; Yuan, X. Biogenesis, Trafficking, and Function of Small RNAs in Plants. Front. Plant Sci. 2022, 13, 825477. [Google Scholar] [CrossRef]
- Brant, E.J.; Budak, H. Plant Small Non-Coding RNAs and Their Roles in Biotic Stresses. Front. Plant Sci. 2018, 9, 1038. [Google Scholar] [CrossRef]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J. Role of MiRNAs and SiRNAs in Biotic and Abiotic Stress Responses of Plants. Biochim. Biophys. Acta—Gene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef]
- Dong, Q.; Hu, B.; Zhang, C. MicroRNAs and Their Roles in Plant Development. Front. Plant Sci. 2022, 13, 315. [Google Scholar] [CrossRef]
- Huang, J.; Yang, M.; Zhang, X. The Function of Small RNAs in Plant Biotic Stress Response. J. Integr. Plant Biol. 2016, 58, 312–327. [Google Scholar] [CrossRef]
- Khanna, K.; Ohri, P.; Bhardwaj, R. Genetic Toolbox and Regulatory Circuits of Plant-Nematode Associations. Plant Physiol. Biochem. 2021, 165, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Kulshrestha, C.; Pathak, H.; Kumar, D.; Dave, S.; Sudan, J. Elucidating Micro RNAs Role in Different Plant–Pathogen Interactions. Mol. Biol. Rep. 2020, 47, 8219–8227. [Google Scholar] [CrossRef] [PubMed]
- Jaubert-Possamai, S.; Noureddine, Y.; Favery, B. MicroRNAs, New Players in the Plant–Nematode Interaction. Front. Plant Sci. 2019, 10, 1180. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.D.; Ganguly, K.; Lim, K.T. RNAi-Based Gene Silencing in Plant-Parasitic Nematodes: A Road toward Crop Improvements. In Nanobiotechnology for Plant Protection, CRISPR and RNAi Systems; Abd-Elsalam, K.A., Lim, K.-T., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 579–596. ISBN 9780128219102. [Google Scholar]
- Hewezi, T. Epigenetic Mechanisms in Nematode-Plant Interactions. Annu. Rev. Phytopathol. 2020, 58, 119–138. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Huang, A.; Li, H.; Feng, L.; Zhang, F.; Guo, W. Identification and Comparative Analysis of MicroRNAs in Pinus massoniana Infected by Bursaphelenchus xylophilus. Plant Growth Regul. 2017, 83, 223–232. [Google Scholar] [CrossRef]
- He, Z.; Webster, S.; He, S.Y. Growth–Defense Trade-Offs in Plants. Curr. Biol. 2022, 32, R634–R639. [Google Scholar] [CrossRef]
- Modesto, I.; Inácio, V.; Van de Peer, Y.; Miguel, C.M. MicroRNA-Mediated Post-Transcriptional Regulation of Pinus pinaster Response and Resistance to Pinewood Nematode. Sci. Rep. 2022, 12, 5160. [Google Scholar] [CrossRef]
- Cai, Q.; He, B.; Kogel, K.H.; Jin, H. Cross-Kingdom RNA Trafficking and Environmental RNAi—Nature’s Blueprint for Modern Crop Protection Strategies. Curr. Opin. Microbiol. 2018, 46, 58–64. [Google Scholar] [CrossRef]
- Gualtieri, C.; Leonetti, P.; Macovei, A. Plant MiRNA Cross-Kingdom Transfer Targeting Parasitic and Mutualistic Organisms as a Tool to Advance Modern Agriculture. Front. Plant Sci. 2020, 11, 930. [Google Scholar] [CrossRef]
- Zhao, J.H.; Guo, H.S. Trans-Kingdom RNA Interactions Drive the Evolutionary Arms Race between Hosts and Pathogens. Curr. Opin. Genet. Dev. 2019, 58–59, 62–69. [Google Scholar] [CrossRef]
- Karimi, H.Z.; Innes, R.W. Molecular Mechanisms Underlying Host-Induced Gene Silencing. Plant Cell 2022, 34, 3183–3199. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.T.; Sheng, D.P.; Ye, J.R.; Cheng, F.M.; Zhang, X.Y.; Qiu, X.W. Study on the Function of Cyp-33D3 Gene of Bursaphelenchus xylophilus with RNA Interference Method. For. Res. 2020, 33, 32–38. [Google Scholar] [CrossRef]
- Qiu, X.; Yang, L.; Ye, J.; Wang, W.; Zhao, T.; Hu, H.; Zhou, G. Silencing of Cyp-33C9 Gene Affects the Reproduction and Pathogenicity of the Pine Wood Nematode, Bursaphelenchus xylophilus. Int. J. Mol. Sci. 2019, 20, 4520. [Google Scholar] [CrossRef] [PubMed]
- Sheng, D.; Zhang, X.; Feng, M.; Ye, J.; Qiu, X. Construction and Its Function Analysis of RNA Interference Vector of Cytochrome Cyp-13A11 Gene in Bursaphelenchus xylophilus. Beijing Linye Daxue Xuebao/J. Beijing For. Univ. 2021, 43, 96–102. [Google Scholar] [CrossRef]
- Wang, F.; Chen, Q.; Zhang, R.; Li, D.; Ling, Y.; Song, R. The Anti-Phytoalexin Gene Bx-Cathepsin W Supports the Survival of Bursaphelenchus xylophilus under Pinus massoniana Phytoalexin Stress. BMC Genom. 2019, 20, 779. [Google Scholar] [CrossRef]
- Shinya, R.; Kirino, H.; Morisaka, H.; Takeuchi-Kaneko, Y.; Futai, K.; Ueda, M. Comparative Secretome and Functional Analyses Reveal Glycoside Hydrolase Family 30 and Cysteine Peptidase as Virulence Determinants in the Pinewood Nematode Bursaphelenchus xylophilus. Front. Plant Sci. 2021, 12, 640459. [Google Scholar] [CrossRef]
- Espada, M.; Silva, A.C.; Eves Van Den Akker, S.; Cock, P.J.A.; Mota, M.; Jones, J.T. Identification and Characterization of Parasitism Genes from the Pinewood Nematode Bursaphelenchus xylophilus Reveals a Multilayered Detoxification Strategy. Mol. Plant Pathol. 2016, 17, 286–295. [Google Scholar] [CrossRef]
- Shinya, R.; Morisaka, H.; Kikuchi, T.; Takeuchi, Y.; Ueda, M.; Futai, K. Secretome Analysis of the Pine Wood Nematode Bursaphelenchus xylophilus Reveals the Tangled Roots of Parasitism and Its Potential for Molecular Mimicry. PLoS ONE 2013, 8, 16–18. [Google Scholar] [CrossRef]
- Cardoso, J.M.S.; Anjo, S.I.; Manadas, B.; Silva, H.; Abrantes, I.; Nakamura, K.; Fonseca, L. Virulence Biomarkers of Bursaphelenchus xylophilus: A Proteomic Approach. Front. Plant Sci. 2022, 12, 822289. [Google Scholar] [CrossRef]
- Dinh, P.T.Y.; Zhang, L.; Brown, C.R.; Elling, A.A. Plant-Mediated RNA Interference of Effector Gene Mc16D10L Confers Resistance against Meloidogyne chitwoodi in Diverse Genetic Backgrounds of Potato and Reduces Pathogenicity of Nematode Offspring. Nematology 2014, 16, 669–682. [Google Scholar] [CrossRef]
- Ibrahim, H.M.M.; Alkharouf, N.W.; Meyer, S.L.F.; Aly, M.A.M.; Gamal El-Din, A.E.K.Y.; Hussein, E.H.A.; Matthews, B.F. Post-Transcriptional Gene Silencing of Root-Knot Nematode in Transformed Soybean Roots. Exp. Parasitol. 2011, 127, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Shivakumara, T.N.; Papolu, P.K.; Dutta, T.K.; Kamaraju, D.; Chaudhary, S.; Rao, U. RNAi-Induced Silencing of an Effector Confers Transcriptional Oscillation in Another Group of Effectors in the Root-Knot Nematode, Meloidogyne incognita. Nematology 2016, 18, 857–870. [Google Scholar] [CrossRef]
- Jeon, H.W.; Park, A.R.; Sung, M.; Kim, N.; Mannaa, M.; Han, G.; Kim, J.; Koo, Y.; Seo, Y.S.; Kim, J.C. Systemic Acquired Resistance-Mediated Control of Pine Wilt Disease by Foliar Application With Methyl Salicylate. Front. Plant Sci. 2022, 12, 812414. [Google Scholar] [CrossRef] [PubMed]
- Mannaa, M.; Han, G.; Jeon, H.W.; Kim, J.; Kim, N.; Park, A.R.; Kim, J.-C.; Seo, Y.-S. Influence of Resistance-Inducing Chemical Elicitors against Pine Wilt Disease on the Rhizosphere Microbiome. Microorganisms 2020, 8, 884. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Cardoso, A.R.; Ferreira, D.; Brito, M.; Pintado, M.E.; Vasconcelos, M.W. Chitosan as a Biocontrol Agent against the Pinewood Nematode (Bursaphelenchus xylophilus). For. Pathol. 2014, 44, 420–423. [Google Scholar] [CrossRef]
- Nunes da Silva, M.; Santos, C.S.; Cruz, A.; López-Villamor, A.; Vasconcelos, M.W. Chitosan Increases Pinus pinaster Tolerance to the Pinewood Nematode (Bursaphelenchus xylophilus) by Promoting Plant Antioxidative Metabolism. Sci. Rep. 2021, 11, 3781. [Google Scholar] [CrossRef]
- Bektas, Y.; Eulgem, T. Synthetic Plant Defense Elicitors. Front. Plant Sci. 2015, 5, 804. [Google Scholar] [CrossRef]
- Park, J.; Jeon, H.W.; Jung, H.; Lee, H.H.; Kim, J.; Park, A.R.; Kim, N.; Han, G.; Kim, J.C.; Seo, Y.S. Comparative Transcriptome Analysis of Pine Trees Treated with Resistance-Inducing Substances against the Nematode Bursaphelenchus xylophilus. Genes 2020, 11, 1000. [Google Scholar] [CrossRef]
- Chen, R.; Huang, K.; Pan, S.; Xu, T.; Tan, J.; Hao, D. Jasmonate Induced Terpene-Based Defense in Pinus massoniana Depresses Monochamus alternatus Adult Feeding. Pest Manag. Sci. 2021, 77, 731–740. [Google Scholar] [CrossRef]
- Sarmast, M.K. Genetic Transformation and Somaclonal Variation in Conifers. Plant Biotechnol. Rep. 2016, 10, 309–325. [Google Scholar] [CrossRef]
- Poovaiah, C.; Phillips, L.; Geddes, B.; Reeves, C.; Sorieul, M.; Thorlby, G. Genome Editing with CRISPR/Cas9 in Pinus radiata (D. Don). BMC Plant Biol. 2021, 21, 363. [Google Scholar] [CrossRef]
- Kumar, J.; Pratap, A.; Kumar, S. (Eds.) Plant Phenomics: An Overview. In Phenomics in Crop Plants: Trends, Options and Limitations; Springer: New Delhi, India, 2015; pp. 1–296. ISBN 9788132222262. [Google Scholar]
- Yu, R.; Luo, Y.; Zhou, Q.; Zhang, X.; Wu, D.; Ren, L. Early Detection of Pine Wilt Disease Using Deep Learning Algorithms and UAV-Based Multispectral Imagery. For. Ecol. Manag. 2021, 497, 119493. [Google Scholar] [CrossRef]

| Host Species | Type of Study | S/R | Type of Analysis | Time Points | Plant Material | Isolate and Origin | Inoc. PWNs | Expm. Conditions | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| P. thunbergii | Gene exp. (LongSAGE) | S species, R varieties | S vs. R | 72 hpi | 2–3 yo seedlings | Shimabara, P. thunbergii | 5000 | Field | [10] |
| P. thunbergii | Gene exp. (SSH) | S species, R varieties | S vs. R | 24 hpi, 72 hpi, 7 dpi, 14 dpi | 2 yo grafts | Ka-4, P. thunbergii | 10,000 | Field | [35] |
| P. thunbergii, P. massoniana | Gene exp. (RNA-seq.) | S species | S response | 24 hpi, 48 hpi, 72 hpi, 96 hpi, 5 dpi, 6 dpi | 3 yo seedlings | Unknown | 10,000 | Greenhouse | [39] |
| P. massoniana | Gene exp. (SSH) | S species | S response | 24 hpi, 72 hpi | 3 yo seedlings | BXY03, P. massoniana | 1500 | Growth chamber | [40] |
| P. massoniana | Gene exp. (RNA-seq.) | S species, R varieties | S vs. R | 24 hpi, 15 dpi, 30 dpi | 4 yo ramets | Guangzhu-3B, unknown | 10,000 | Nursery | [37] |
| P. massoniana | Gene exp. (RNA-seq.) | S species | S response | 24 hpi, 48 hpi, 72 hpi | 2 yo seedlings | Unknown, P. massoniana | 2000 | Greenhouse | [41] |
| P. massoniana | Proteomics | S species | S vs. R | 14 dpi | 2 yo seedlings | Unknown | 1000 | Unknown | [42] |
| P. massoniana | Proteomics | S species | S vs. R | Constitutive response | grafts | - | - | Field | [43] |
| P. densiflora | Gene exp. (ACP, SSH) | S species | S response | 21 hpi, 24 hpi, 7 dpi | 4 yo and 8 yo seedlings | Unknown, P. thunbergii | 6000 to 30,000 | Field and nursery | [44] |
| P. densiflora | Gene exp. (RNA-seq.) | S species | Pathogenic vs. non-pathogenic PWN | 28 dpi | adult trees (11–13 m) | Unknown, P. densiflora | 60,000 | Field | [45] |
| P. pinaster | Gene exp. (RNA-seq.) | S species | S response | 6 hpi, 24 hpi, 48 hpi, 7 dpi | 3 yo seedlings | Unknown | 2000 | Field | [46] |
| P. pinaster | Gene exp. (RNA-seq.) | S species, R varieties | S vs. R | 72 hpi | 4 yo seedlings | Bx013.003, P. pinaster | 500 | Greenhouse | [38] |
| P. pinaster | Metabolomics | S species, R varieties | S vs. R | 14 dpi, 21 dpi, 28 dpi, 35 dpi | 2 yo seedlings | Bx013.003, P. pinaster | 500 | Greenhouse | [47] |
| P. pinaster, P. pinea | Gene exp. (pyroseq.) | S and R species | S vs. R | 24 hpi | 2 yo seedlings | HF, P. pinaster | 1000 | Growth chamber | [48] |
| P. pinaster, P. yunnanensis | Gene exp. (RNA-seq.) | S and R species | S vs. R | 6 hpi, 24 hpi, 48 hpi, 7 dpi | 3 yo seedlings | Unknown | 2000 | Field | [49] |
| Pathway | Genes | P. densiflora | P. massoniana | P. thunbergii | P. pinaster | P. pinea | P. yunnanensis | P. strobus |
|---|---|---|---|---|---|---|---|---|
| Phenylpropanoid biosynthesis | phenylalanine ammonia-lyase (PAL) | up | up; S < R | up | ||||
| 4-coumarate-CoA ligase (4CL) | up | up; S < R | up | |||||
| caffeoyl-CoA O-methyltransferase (CCoAOMT) | up | up | up | |||||
| caffeic acid O-methyltransferase (COMT) | up | up | ||||||
| Flavonoid biosynthesis | chalcone synthase (CHS) | up | up | up | up; S < R | up | ||
| chalcone isomerase | up | up | ||||||
| flavonol synthase (FLS) | up | |||||||
| flavonoid hydroxilase | up | |||||||
| leucoanthocyanidin dioxygenase (LDOX) | up | up; S > R | up; S < R | |||||
| leucoanthocyanidin reductase (LAR) | up; S < R | |||||||
| anthocyanidin synthase (ANS) | up; S < R | |||||||
| Stilbenoid biosynthesis | pinosylvin synthase (STS) | up | up | up | up | |||
| pinosylvin O-methyltransferase (PMT) | up | |||||||
| Lignans biosynthesis | phenylcoumaran benzylic ether reductase | up | ||||||
| Lignin biosynthesis | cinnamoyl-CoA reductase (CCR) | up | up | up | ||||
| cinnamyl-alcohool dehydrogenase (CAD) | up; down in S | up | ||||||
| peroxidase (PER) | up | up; S < R | up; S < R | up; S < R | up | |||
| laccase (LAC) | up; S < R | |||||||
| Transcription factors | bHLH | up | up | up | ||||
| MYB | up | up | up | up | ||||
| WRKY | up | up | up | up | ||||
| References | [44,45] | [37,39,40,41] | [35,36,39] | [38,46] | [48] | [49] | [66] | |
| Genes | P. densiflora | P. massoniana | P. thunbergii | P. pinaster | P. yunnanensis |
|---|---|---|---|---|---|
| superoxide dismutase | up | down, S < R | up | up | |
| glutathione reductase | down, S < R | ||||
| glutathione peroxidase (GPx) | up | up | |||
| L-ascorbate peroxidase | up | down | |||
| catalase (CAT) | down, S < R | S < R | |||
| catalase isozyme | up | ||||
| peroxidase (PER) | up | S < R | S < R | S < R | up |
| glutathione S-transferase | up | up; S < R | up | ||
| peroxiredoxin | up | ||||
| thioredoxin | up | up | |||
| References | [44] | [37,40] | [35,36] | [38,46] | [49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Modesto, I.; Mendes, A.; Carrasquinho, I.; Miguel, C.M. Molecular Defense Response of Pine Trees (Pinus spp.) to the Parasitic Nematode Bursaphelenchus xylophilus. Cells 2022, 11, 3208. https://doi.org/10.3390/cells11203208
Modesto I, Mendes A, Carrasquinho I, Miguel CM. Molecular Defense Response of Pine Trees (Pinus spp.) to the Parasitic Nematode Bursaphelenchus xylophilus. Cells. 2022; 11(20):3208. https://doi.org/10.3390/cells11203208
Chicago/Turabian StyleModesto, Inês, André Mendes, Isabel Carrasquinho, and Célia M. Miguel. 2022. "Molecular Defense Response of Pine Trees (Pinus spp.) to the Parasitic Nematode Bursaphelenchus xylophilus" Cells 11, no. 20: 3208. https://doi.org/10.3390/cells11203208
APA StyleModesto, I., Mendes, A., Carrasquinho, I., & Miguel, C. M. (2022). Molecular Defense Response of Pine Trees (Pinus spp.) to the Parasitic Nematode Bursaphelenchus xylophilus. Cells, 11(20), 3208. https://doi.org/10.3390/cells11203208