

Approaches towards Elucidating the Metabolic Program of Hematopoietic Stem/Progenitor Cells


Abstract
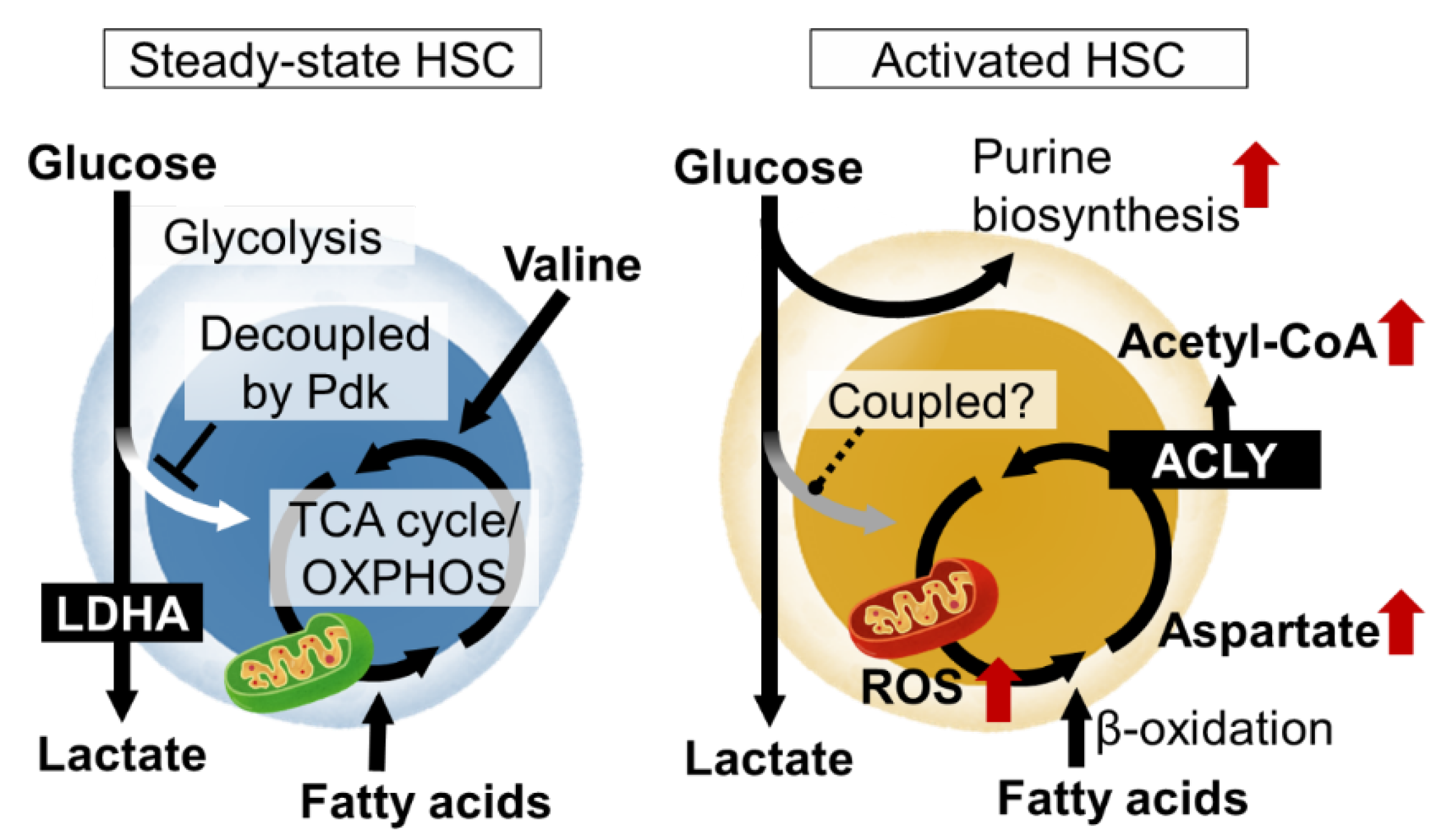
:1. Introduction: Metabolic Programs Regulating Stem Cell Identity and Function
2. Approaches to Metabolic Analysis of Stem Cells (Summarized in Table 1)
2.1. Deletion, Knockdown or Overexpression of Specific Genes

{kind=link}
| Name | Method | Advantage(s) | Disadvantages(s) |
|---|---|---|---|
| Deletion, knockdown or overexpression of specific genes | Deletion of gene function by gene targeting or editing; knockdown by vectors or oligonucleotides. | Clearly identifies the metabolic effects of each gene on stem cells. | Without spatiotemporally controlled genetic manipulation, only secondary metabolic effects on stem cells may be observed. |
| Classical metabolite assays using enzymatic reactions | Metabolite detection utilizing enzymatic reactions. | The principle is simple and easy to use. Many kits are commercially available. | Detection sensitivity may be low for use with small numbers of stem cells. There may be no simple enzymatic assay for unusual metabolites. |
| Mass spectrometry (MS) | Metabolites in intracellular extracts or extracellular fluid are separated by chromatography and measured with a mass spectrometer. | More sensitive than NMR and can identify many metabolites. Levels of unknown metabolites can be identified. Both fate tracking and spatial analysis of metabolites are possible. | Because it is a destructive test, time variation of metabolite levels in the same cell/tissue is unknown. Single cell analysis is under development. |
| Nuclear magnetic resonance (NMR) spectroscopy | Metabolites in a sample are identified by applying radio waves to the nuclei of molecules placed in a strong magnetic field, causing them to resonate. | Allows for nondestructive metabolite analysis. The fate of atoms derived from metabolite tracers can be accurately determined from which position they were incorporated. | Not sensitive enough to analyze a small number of stem cells. Untargeted analysis of metabolites is difficult. |
| Chemical probes | Cells take up chemical probes that interact with a metabolite and detect the changes in fluorescence intensity or fluorescence ratio. | A small number of cells can be analyzed. Combination of surface markers enables analysis of stem cells from many cells. Can analyze changes over time in a single cell as well as differences between cells. | Stem cells possess the ability to pump chemical probes out. Even if qualitative analysis is possible, absolute quantification can be difficult. |
| Genetic biosensors | Genetically encoded fluorescent biosensors to analyze metabolite levels and metabolic characteristics. | A small number of cells can be analyzed. Combination of surface markers enables analysis of stem cells from many cells. Can analyze changes over time in a single cell as well as differences between cells. | Need to select biosensors suitable for the number of metabolites in stem cells. Biosensors must be well expressed in stem cells. Ratiometric sensors are more quantitative. |
| Flux analyzer | Electrically measure oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) of cells in small wells. | The metabolic state of tens of cells at steady state or under stress load can be measured by adding inhibitors or metabolites over time. | At least tens of thousands of cells are required to obtain an average value that ignores the individuality of each cell. Note that this is a measurement in an in vitro medium and to what extent it reflects OCR and ECAR in the body. |
2.2. Classical Metabolite Assays Using Enzymatic Reactions
2.3. Metabolite Analysis by Mass Spectrometry (MS) and Nuclear Magnetic Resonance (NMR) Spectroscopy
2.4. Measurement Using Chemical Probes and Genetic Biosensors
2.5. Flux Analyzer
3. HSCs and Metabolic Program
3.1. HSCs and Their Metabolic Programs Which Maintain Individual Hematopoiesis
3.2. Metabolic Control of HSPCs in Steady State
3.2.1. Hypoxic Microenvironment and HSCs
3.2.2. Regulation of Glycolysis and Decoupling from TCA Cycle in HSCs
3.2.3. Mitochondrial Dynamics and Steady-State HSCs
3.2.4. Environmentally-Derived Metabolites That Maintain HSC Homeostasis
3.3. Metabolic Regulation of HSCs under Stress
3.3.1. Acute Stress and HSC Metabolism
3.3.2. Mitochondrial Activity and Acutely Stressed HSCs
3.3.3. Aging Stress and HSC Metabolism
3.4. Application of Metabolic Findings of HSC
3.4.1. Metabolic Optimization of HSC Culture
3.4.2. Non-Conditional Transplantation with the Removal of Metabolites from the Diet
4. Conclusions and Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandel, N.S. Carbohydrate Metabolism. Cold Spring Harb. Perspect. Biol. 2021, 13, a040568. [Google Scholar] [CrossRef]
- Chandel, N.S. Glycolysis. Cold Spring Harb. Perspect. Biol. 2021, 13, a040535. [Google Scholar] [CrossRef] [PubMed]
- Chandel, N.S. Mitochondria. Cold Spring Harb. Perspect. Biol. 2021, 13, a040543. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Thompson, C.B. Metabolic Regulation of Epigenetics. Cell Metab. 2012, 16, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem Cells, Cancer, and Cancer Stem Cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, K.A.; Lemischka, I.R. Stem Cells and Their Niches. Science 2006, 311, 1880–1885. [Google Scholar] [CrossRef] [Green Version]
- Suda, T.; Arai, F.; Hirao, A. Hematopoietic Stem Cells and Their Niche. Trends Immunol. 2005, 26, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Suda, T.; Takubo, K.; Semenza, G.L. Metabolic Regulation of Hematopoietic Stem Cells in the Hypoxic Niche. Cell Stem Cell 2011, 9, 298–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandel, N.S.; Jasper, H.; Ho, T.T.; Passegué, E. Metabolic Regulation of Stem Cell Function in Tissue Homeostasis and Organismal Ageing. Nat. Cell Biol. 2016, 18, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Nakamura-Ishizu, A.; Ito, K.; Suda, T. Hematopoietic Stem Cell Metabolism during Development and Aging. Dev. Cell 2020, 54, 239–255. [Google Scholar] [CrossRef]
- Nakamura-Ishizu, A.; Takizawa, H.; Suda, T. The Analysis, Roles and Regulation of Quiescence in Hematopoietic Stem Cells. Development 2014, 141, 4656–4666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, E.; Colla, S.; Liesa, M.; Moslehi, J.; Müller, F.L.; Guo, M.; Cooper, M.; Kotton, D.; Fabian, A.J.; Walkey, C.; et al. Telomere Dysfunction Induces Metabolic and Mitochondrial Compromise. Nature 2011, 470, 359–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palm, W.; Thompson, C.B. Nutrient Acquisition Strategies of Mammalian Cells. Nature 2017, 546, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Leach, F.R.; Webster, J.J. Commercially Available Firefly Luciferase Reagents. Methods Enzymol. 1986, 133, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Soga, T.; Ueno, Y.; Naraoka, H.; Ohashi, Y.; Tomita, M.; Nishioka, T. Simultaneous Determination of Anionic Intermediates for Bacillus Subtilis Metabolic Pathways by Capillary Electrophoresis Electrospray Ionization Mass Spectrometry. Anal. Chem. 2002, 74, 2233–2239. [Google Scholar] [CrossRef]
- Soga, T.; Ueno, Y.; Naraoka, H.; Matsuda, K.; Tomita, M.; Nishioka, T. Pressure-Assisted Capillary Electrophoresis Electrospray Ionization Mass Spectrometry for Analysis of Multivalent Anions. Anal. Chem. 2002, 74, 6224–6229. [Google Scholar] [CrossRef]
- Zhou, B.; Xiao, J.F.; Tuli, L.; Ressom, H.W. LC-MS-Based Metabolomics. Mol. BioSyst. 2012, 8, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Castiglione, F.; Ferro, M.; Mavroudakis, E.; Pellitteri, R.; Bossolasco, P.; Zaccheo, D.; Morbidelli, M.; Silani, V.; Mele, A.; Moscatelli, D.; et al. NMR Metabolomics for Stem Cell Type Discrimination. Sci. Rep. 2017, 7, 15808. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, S.; Kishimoto, S.; Saito, K.; Takakusagi, Y.; Munasinghe, J.P.; Devasahayam, N.; Hart, C.P.; Gillies, R.J.; Mitchell, J.B.; Krishna, M.C. Metabolic and Physiologic Imaging Biomarkers of the Tumor Microenvironment Predict Treatment Outcome with Radiation or a Hypoxia-Activated Prodrug in Mice. Cancer Res. 2018, 78, 3783–3792. [Google Scholar] [CrossRef] [Green Version]
- DeVilbiss, A.W.; Zhao, Z.; Martin-Sandoval, M.S.; Ubellacker, J.M.; Tasdogan, A.; Agathocleous, M.; Mathews, T.P.; Morrison, S.J. Metabolomic Profiling of Rare Cell Populations Isolated by Flow Cytometry from Tissues. Elife 2021, 10, e61980. [Google Scholar] [CrossRef]
- Emara, S.; Amer, S.; Ali, A.; Abouleila, Y.; Oga, A.; Masujima, T. Single-Cell Metabolomics. Adv. Exp. Med. Biol. 2017, 965, 323–343. [Google Scholar] [CrossRef]
- Yoshioka, K.; Takahashi, H.; Homma, T.; Saito, M.; Oh, K.B.; Nemoto, Y.; Matsuoka, H. A Novel Fluorescent Derivative of Glucose Applicable to the Assessment of Glucose Uptake Activity of Escherichia Coli. Biochim. Biophys. Acta 1996, 1289, 5–9. [Google Scholar] [CrossRef]
- Takubo, K.; Nagamatsu, G.; Kobayashi, C.I.; Nakamura-Ishizu, A.; Kobayashi, H.; Ikeda, E.; Goda, N.; Rahimi, Y.; Johnson, R.S.; Soga, T.; et al. Regulation of Glycolysis by Pdk Functions as a Metabolic Checkpoint for Cell Cycle Quiescence in Hematopoietic Stem Cells. Cell Stem Cell 2013, 12, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, L.; Stojkovic, M.; Dimmick, I.; Ahmad, S.; Stojkovic, P.; Hole, N.; Lako, M. Phenotypic Characterization of Murine Primitive Hematopoietic Progenitor Cells Isolated on Basis of Aldehyde Dehydrogenase Activity. Stem Cells 2004, 22, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Juopperi, T.A.; Schuler, W.; Yuan, X.; Collector, M.I.; Dang, C.V.; Sharkis, S.J. Isolation of Bone Marrow-Derived Stem Cells Using Density-Gradient Separation. Exp. Hematol. 2007, 35, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Cheng, A.; Luo, Y.; Lu, C.; Mattson, M.P.; Rao, M.S.; Furukawa, K. Membrane Properties of Rat Embryonic Multipotent Neural Stem Cells. J. Neurochem. 2004, 88, 212–226. [Google Scholar] [CrossRef]
- Umezawa, K.; Yoshida, M.; Kamiya, M.; Yamasoba, T.; Urano, Y. Rational Design of Reversible Fluorescent Probes for Live-Cell Imaging and Quantification of Fast Glutathione Dynamics. Nat. Chem. 2017, 9, 279–286. [Google Scholar] [CrossRef]
- Mächler, P.; Wyss, M.T.; Elsayed, M.; Stobart, J.; Gutierrez, R.; von Faber-Castell, A.; Kaelin, V.; Zuend, M.; San Martín, A.; Romero-Gómez, I.; et al. In Vivo Evidence for a Lactate Gradient from Astrocytes to Neurons. Cell Metab. 2016, 23, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Oki, T.; Mercier, F.; Kato, H.; Jung, Y.; McDonald, T.O.; Spencer, J.A.; Mazzola, M.C.; van Gastel, N.; Lin, C.P.; Michor, F.; et al. Imaging Dynamic MTORC1 Pathway Activity in Vivo Reveals Marked Shifts That Support Time-Specific Inhibitor Therapy in AML. Nat. Commun. 2021, 12, 245. [Google Scholar] [CrossRef]
- Watanuki, S.; Kobayashi, H.; Sorimachi, Y.; Yamamoto, M.; Okamoto, S.; Takubo, K. ATP Turnover and Glucose Dependency in Hematopoietic Stem/Progenitor Cells Are Increased by Proliferation and Differentiation. Biochem. Biophys. Res. Commun. 2019, 514, 287–294. [Google Scholar] [CrossRef]
- Ho, T.T.; Warr, M.R.; Adelman, E.R.; Lansinger, O.M.; Flach, J.; Verovskaya, E.V.; Figueroa, M.E.; Passegué, E. Autophagy Maintains the Metabolism and Function of Young and Old Stem Cells. Nature 2017, 543, 205–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orkin, S.H.; Zon, L.I. Hematopoiesis: An Evolving Paradigm for Stem Cell Biology. Cell 2008, 132, 631–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Agosto, J.A.; Mikkola, H.K.A.; Hartenstein, V.; Banerjee, U. The Hematopoietic Stem Cell and Its Niche: A Comparative View. Genes Dev. 2007, 21, 3044–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trumpp, A.; Essers, M.; Wilson, A. Awakening Dormant Haematopoietic Stem Cells. Nat. Rev. Immunol. 2010, 10, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Karigane, D.; Takubo, K. Metabolic Regulation of Hematopoietic and Leukemic Stem/Progenitor Cells under Homeostatic and Stress Conditions. Int. J. Hematol. 2017, 106, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Takubo, K.; Goda, N.; Yamada, W.; Iriuchishima, H.; Ikeda, E.; Kubota, Y.; Shima, H.; Johnson, R.S.; Hirao, A.; Suematsu, M.; et al. Regulation of the HIF-1alpha Level Is Essential for Hematopoietic Stem Cells. Cell Stem Cell 2010, 7, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Simsek, T.; Kocabas, F.; Zheng, J.; Deberardinis, R.J.; Mahmoud, A.I.; Olson, E.N.; Schneider, J.W.; Zhang, C.C.; Sadek, H.A. The Distinct Metabolic Profile of Hematopoietic Stem Cells Reflects Their Location in a Hypoxic Niche. Cell Stem Cell 2010, 7, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Nombela-Arrieta, C.; Pivarnik, G.; Winkel, B.; Canty, K.J.; Harley, B.; Mahoney, J.E.; Park, S.-Y.; Lu, J.; Protopopov, A.; Silberstein, L.E. Quantitative Imaging of Haematopoietic Stem and Progenitor Cell Localization and Hypoxic Status in the Bone Marrow Microenvironment. Nat. Cell Biol. 2013, 15, 533–543. [Google Scholar] [CrossRef]
- Spencer, J.A.; Ferraro, F.; Roussakis, E.; Klein, A.; Wu, J.; Runnels, J.M.; Zaher, W.; Mortensen, L.J.; Alt, C.; Turcotte, R.; et al. Direct Measurement of Local Oxygen Concentration in the Bone Marrow of Live Animals. Nature 2014, 508, 269–273. [Google Scholar] [CrossRef] [Green Version]
- Christodoulou, C.; Spencer, J.A.; Yeh, S.-C.A.; Turcotte, R.; Kokkaliaris, K.D.; Panero, R.; Ramos, A.; Guo, G.; Seyedhassantehrani, N.; Esipova, T.v.; et al. Live-Animal Imaging of Native Haematopoietic Stem and Progenitor Cells. Nature 2020, 578, 278–283. [Google Scholar] [CrossRef]
- Miharada, K.; Karlsson, G.; Rehn, M.; Rörby, E.; Siva, K.; Cammenga, J.; Karlsson, S. Cripto Regulates Hematopoietic Stem Cells as a Hypoxic-Niche-Related Factor through Cell Surface Receptor GRP78. Cell Stem Cell 2011, 9, 330–344. [Google Scholar] [CrossRef] [Green Version]
- Jakubison, B.L.; Sarkar, T.; Gudmundsson, K.O.; Singh, S.; Sun, L.; Morris, H.M.; Klarmann, K.D.; Keller, J.R. ID2 and HIF-1α Collaborate to Protect Quiescent Hematopoietic Stem Cells from Activation, Differentiation, and Exhaustion. J. Clin. Investig. 2022, 132, e152599. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Israelsen, W.J.; Lee, D.; Yu, V.W.C.; Jeanson, N.T.; Clish, C.B.; Cantley, L.C.; vander Heiden, M.G.; Scadden, D.T. Cell-State-Specific Metabolic Dependency in Hematopoiesis and Leukemogenesis. Cell 2014, 158, 1309–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, M.J.; Luchsinger, L.L.; Corrigan, D.J.; Williams, L.J.; Snoeck, H.-W. Dye-Independent Methods Reveal Elevated Mitochondrial Mass in Hematopoietic Stem Cells. Cell Stem Cell 2017, 21, 725–729.e4. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.-M.; Liu, X.; Shen, J.; Jovanovic, O.; Pohl, E.E.; Gerson, S.L.; Finkel, T.; Broxmeyer, H.E.; Qu, C.-K. Metabolic Regulation by the Mitochondrial Phosphatase PTPMT1 Is Required for Hematopoietic Stem Cell Differentiation. Cell Stem Cell 2013, 12, 62–74. [Google Scholar] [CrossRef] [Green Version]
- Guitart, A.V.; Panagopoulou, T.I.; Villacreces, A.; Vukovic, M.; Sepulveda, C.; Allen, L.; Carter, R.N.; van de Lagemaat, L.N.; Morgan, M.; Giles, P.; et al. Fumarate Hydratase Is a Critical Metabolic Regulator of Hematopoietic Stem Cell Functions. J. Exp. Med. 2017, 214, 719–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchsinger, L.L.; de Almeida, M.J.; Corrigan, D.J.; Mumau, M.; Snoeck, H.-W. Mitofusin 2 Maintains Haematopoietic Stem Cells with Extensive Lymphoid Potential. Nature 2016, 529, 528–531. [Google Scholar] [CrossRef] [Green Version]
- Ansó, E.; Weinberg, S.E.; Diebold, L.P.; Thompson, B.J.; Malinge, S.; Schumacker, P.T.; Liu, X.; Zhang, Y.; Shao, Z.; Steadman, M.; et al. The Mitochondrial Respiratory Chain Is Essential for Haematopoietic Stem Cell Function. Nat. Cell Biol. 2017, 19, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Wallscheid, N.; Buettner, F.; Sommerkamp, P.; Klimmeck, D.; Ladel, L.; Thalheimer, F.B.; Pastor-Flores, D.; Roma, L.P.; Renders, S.; Zeisberger, P.; et al. Vitamin A-Retinoic Acid Signaling Regulates Hematopoietic Stem Cell Dormancy. Cell 2017, 169, 807–823.e19. [Google Scholar] [CrossRef] [Green Version]
- Schönberger, K.; Obier, N.; Romero-Mulero, M.C.; Cauchy, P.; Mess, J.; Pavlovich, P.V.; Zhang, Y.W.; Mitterer, M.; Rettkowski, J.; Lalioti, M.-E.; et al. Multilayer Omics Analysis Reveals a Non-Classical Retinoic Acid Signaling Axis That Regulates Hematopoietic Stem Cell Identity. Cell Stem Cell 2022, 29, 131–148.e10. [Google Scholar] [CrossRef]
- Agathocleous, M.; Meacham, C.E.; Burgess, R.J.; Piskounova, E.; Zhao, Z.; Crane, G.M.; Cowin, B.L.; Bruner, E.; Murphy, M.M.; Chen, W.; et al. Ascorbate Regulates Haematopoietic Stem Cell Function and Leukaemogenesis. Nature 2017, 549, 476–481. [Google Scholar] [CrossRef] [Green Version]
- Karigane, D.; Kobayashi, H.; Morikawa, T.; Ootomo, Y.; Sakai, M.; Nagamatsu, G.; Kubota, Y.; Goda, N.; Matsumoto, M.; Nishimura, E.K.; et al. P38α Activates Purine Metabolism to Initiate Hematopoietic Stem/Progenitor Cell Cycling in Response to Stress. Cell Stem Cell 2016, 19, 192–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, M.; Moehrle, B.; Brown, A.; Eiwen, K.; Sakk, V.; Geiger, H. HPRT and Purine Salvaging Are Critical for Hematopoietic Stem Cell Function. Stem Cells 2019, 37, 1606–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umemoto, T.; Hashimoto, M.; Matsumura, T.; Nakamura-Ishizu, A.; Suda, T. Ca2+-Mitochondria Axis Drives Cell Division in Hematopoietic Stem Cells. J. Exp. Med. 2018, 215, 2097–2113. [Google Scholar] [CrossRef]
- Qi, L.; Martin-Sandoval, M.S.; Merchant, S.; Gu, W.; Eckhardt, M.; Mathews, T.P.; Zhao, Z.; Agathocleous, M.; Morrison, S.J. Aspartate Availability Limits Hematopoietic Stem Cell Function during Hematopoietic Regeneration. Cell Stem Cell 2021, 28, 1982–1999.e8. [Google Scholar] [CrossRef]
- Umemoto, T.; Johansson, A.; Ahmad, S.A.I.; Hashimoto, M.; Kubota, S.; Kikuchi, K.; Odaka, H.; Era, T.; Kurotaki, D.; Sashida, G.; et al. ATP Citrate Lyase Controls Hematopoietic Stem Cell Fate and Supports Bone Marrow Regeneration. EMBO J. 2022, 41, e109463. [Google Scholar] [CrossRef]
- Dong, S.; Wang, Q.; Kao, Y.-R.; Diaz, A.; Tasset, I.; Kaushik, S.; Thiruthuvanathan, V.; Zintiridou, A.; Nieves, E.; Dzieciatkowska, M.; et al. Chaperone-Mediated Autophagy Sustains Haematopoietic Stem-Cell Function. Nature 2021, 591, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Mansell, E.; Sigurdsson, V.; Deltcheva, E.; Brown, J.; James, C.; Miharada, K.; Soneji, S.; Larsson, J.; Enver, T. Mitochondrial Potentiation Ameliorates Age-Related Heterogeneity in Hematopoietic Stem Cell Function. Cell Stem Cell 2021, 28, 241–256.e6. [Google Scholar] [CrossRef]
- Sorimachi, Y.; Karigane, D.; Ootomo, Y.; Kobayashi, H.; Morikawa, T.; Otsu, K.; Kubota, Y.; Okamoto, S.; Goda, N.; Takubo, K. P38α Plays Differential Roles in Hematopoietic Stem Cell Activity Dependent on Aging Contexts. J. Biol. Chem. 2021, 296, 100563. [Google Scholar] [CrossRef]
- Kobayashi, H.; Morikawa, T.; Okinaga, A.; Hamano, F.; Hashidate-Yoshida, T.; Watanuki, S.; Hishikawa, D.; Shindou, H.; Arai, F.; Kabe, Y.; et al. Environmental Optimization Enables Maintenance of Quiescent Hematopoietic Stem Cells Ex Vivo. Cell Rep. 2019, 28, 145–158.e9. [Google Scholar] [CrossRef] [PubMed]
- Taya, Y.; Ota, Y.; Wilkinson, A.C.; Kanazawa, A.; Watarai, H.; Kasai, M.; Nakauchi, H.; Yamazaki, S. Depleting Dietary Valine Permits Nonmyeloablative Mouse Hematopoietic Stem Cell Transplantation. Science 2016, 354, 1152–1155. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.; Mahesula, S.; Mathews, T.P.; Martin-Sandoval, M.S.; Zhao, Z.; Piskounova, E.; Agathocleous, M. The Requirement for Pyruvate Dehydrogenase in Leukemogenesis Depends on Cell Lineage. Cell Metab. 2021, 33, 1777–1792.e8. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobayashi, H.; Watanuki, S.; Takubo, K. Approaches towards Elucidating the Metabolic Program of Hematopoietic Stem/Progenitor Cells. Cells 2022, 11, 3189. https://doi.org/10.3390/cells11203189
Kobayashi H, Watanuki S, Takubo K. Approaches towards Elucidating the Metabolic Program of Hematopoietic Stem/Progenitor Cells. Cells. 2022; 11(20):3189. https://doi.org/10.3390/cells11203189
Chicago/Turabian StyleKobayashi, Hiroshi, Shintaro Watanuki, and Keiyo Takubo. 2022. "Approaches towards Elucidating the Metabolic Program of Hematopoietic Stem/Progenitor Cells" Cells 11, no. 20: 3189. https://doi.org/10.3390/cells11203189
APA StyleKobayashi, H., Watanuki, S., & Takubo, K. (2022). Approaches towards Elucidating the Metabolic Program of Hematopoietic Stem/Progenitor Cells. Cells, 11(20), 3189. https://doi.org/10.3390/cells11203189