

Evaluation of Malondialdehyde Levels, Oxidative Stress and Host–Bacteria Interactions: Escherichia coli and Salmonella Derby


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Bacterial Suspension/Extract Preparation
2.3. MDA Determination
2.4. Statistical Analysis
3. Results
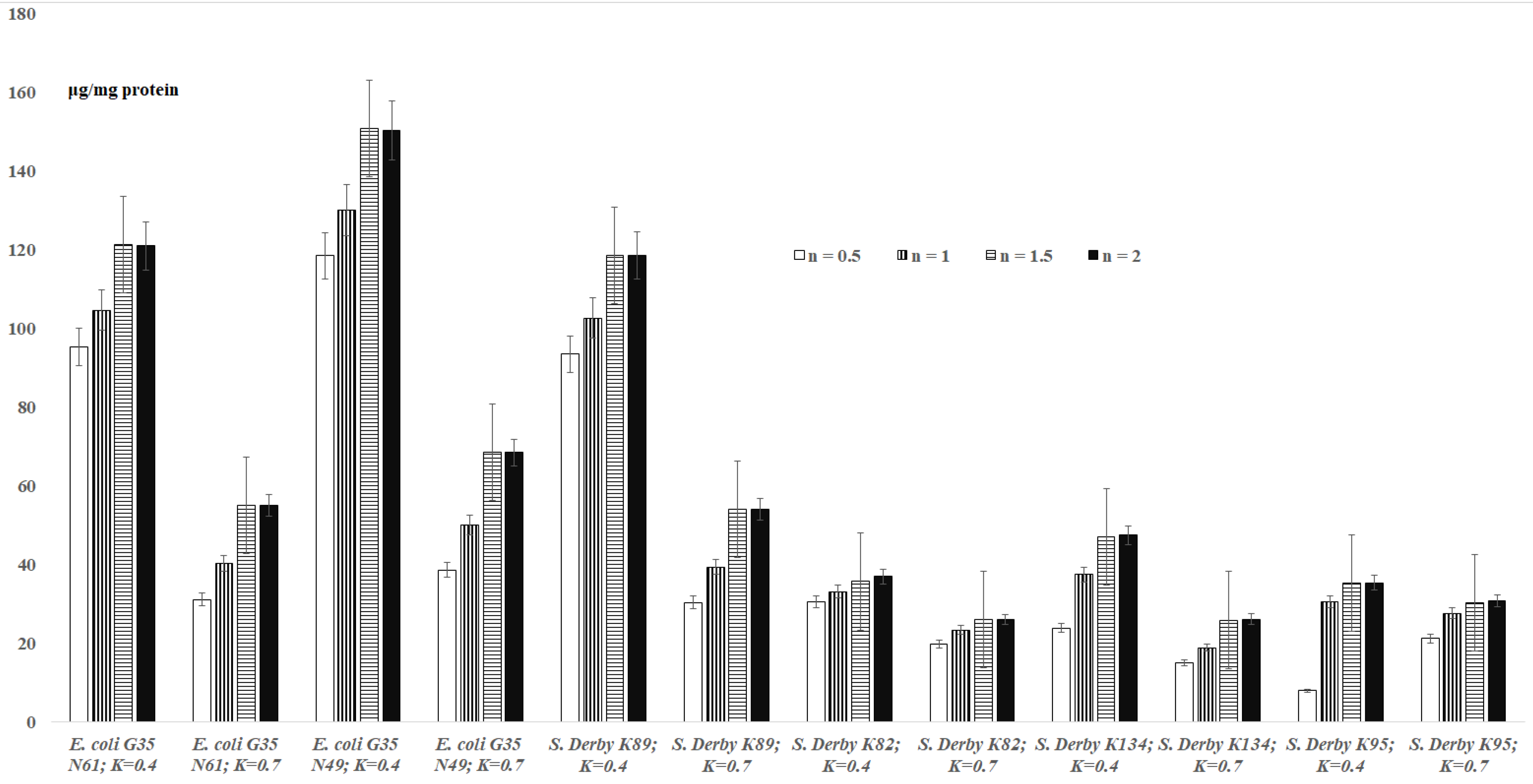
3.1. TBARs Levels in the Bacterial Suspensions
3.2. TBARs Levels in the Bacterial Extracts
3.3. TBARs Levels in the Logarithmic and Stationary Phase E. coli Cells
3.4. TBARs Levels for the E. coli G35 Cells
4. Discussion
4.1. TBARs Levels in the Bacterial Suspensions and Extracts
4.1.1. Comparative Evaluation of TBARs Levels in the Bacterial Cells
4.1.2. Comparative Evaluation of TBARs Levels in the Cells of E. coli G35
4.1.3. Comparative Evaluation of TBARs Levels in the UV-Sensitive Cells of S. Derby K134
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.M.; Rock, C. Membrane Lipid Homeostasis in Bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Araújo, J.R.; Tazi, A.; Burlen-Defranoux, O.; Vichier-Guerre, S.; Nigro, G.; Licandro, H.; Demignot, S.; Sansonetti, P.J. Fermentation Products of Commensal Bacteria Alter Enterocyte Lipid Metabolism. Cell Host Microbe 2020, 27, 358–375. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Mih, N.; Monk, J.M.; Fang, X.; Catoiu, E.; Heckmann, D.; Yang, L.; Palsson, B. Adaptations of Escherichia coli Strains to Oxidative Stress are Reflected in Properties of their Structural Proteomes. BMC Bioinform. 2020, 21, 162. [Google Scholar] [CrossRef]
- Imlay, J.A. Where in the World do Bacteria Experience Oxidative Stress? Environ. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef]
- Hsu, K.C.; Hsu, P.F.; Chen, Y.C.; Lin, H.C.; Hung, C.C.; Chen, P.C.; Huang, Y.L. Oxidative Stress During Bacterial Growth Characterized through Microdialysis Sampling Coupled with HPLC/fluorescence Detection of Malondialdehyde. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2016, 1019, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Toto, A.; Wild, P.; Graille, M.; Turcu, V.; Crézé, C.; Hemmendinger, M.; Sauvain, J.J.; Bergamaschi, E.; GusevaCanu, I.; Hopf, N.B. Urinary Malondialdehyde (MDA) Concentrations in the General Population-A Systematic Literature Review and Meta-Analysis. Toxics 2022, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Pepoyan, A.Z. Role of Membranes in UV Resistance of Enterobacteriaceae Cells. Ph.D. Thesis, Institute of Molecular Biology, NAS RA, Yerevan, Armenia, 2002; p. 222. [Google Scholar]
- Zhuravlev, A.V.; Vetrovoy, O.V.; Savvateeva-Popova, E.V. Enzymatic and Non-enzymatic Pathways of Kynurenines’ Dimerization: The Molecular Factors for Oxidative Stress Development. PLoS Comput. Biol. 2018, 14, e1006672. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.H.A.; Hozzein, W.N.; Mousa, A.S.M.; Rabie, W.; Alkhalifah, D.H.M.; Selim, S.; AbdElgawad, H. Heat Stress as an Innovative Approach to Enhance the Antioxidant Production in Pseudooceanicola and Bacillus Isolates. Sci. Rep. 2020, 10, 15076. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.D.; Miklos, A.E.; Chiao, A.C.; Sun, Z.Z.; Lux, M.W. Methodologies for Preparation of Prokaryotic Extracts for Cell-free Expression Systems. Synth. Syst. Biotechnol. 2020, 5, 252–267. [Google Scholar] [CrossRef] [PubMed]
- Balayan, M.H.; Manvelyan, A.M.; Marutyan, S.; Isajanyan, M.; Tsaturyan, V.V.; Pepoyan, A.Z.; Marotta, F.; Torok, T. Impact of Lactobacillus acidophilus INMIA 9602 Er-2 and Escherichia coli M-17 on Some Clinical Blood Characteristics of Familial Mediterranean Fever Disease Patients from the Armenian Cohort. Int. J. Probiotics Prebiotics 2015, 10, 91–95. [Google Scholar]
- Khan, R.; Petersen, F.C.; Shekhar, S. Commensal Bacteria: An Emerging Player in Defense Against Respiratory Pathogens. Front. Immunol. 2019, 10, 1203. [Google Scholar] [CrossRef] [PubMed]
- Ragonnaud, E.; Biragyn, A. Gut Microbiota as the Key Controllers of "Healthy" Aging of Elderly People. Immun. Ageing. 2021, 18, 2. [Google Scholar] [CrossRef]
- Pepoyan, A.Z.; Tsaturyan, V.V.; Badalyan, M.; Weeks, R.; Kamiya, S.; Chikindas, M.L. Blood Protein Polymorphisms and the Gut Bacteria: Impact of Probiotic Lactobacillus acidophilus Narine on Salmonella Carriage in Sheep. Benef. Microbes 2020, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pepoyan, A.Z.; Balayan, M.H.; Manvelyan, A.M.; Galstyan, L.; Pepoyan, S.; Petrosyan, S.D.; Tsaturyan, V.V.; Kamiya, S.; Torok, T.; Chikindas, M. Probiotic Lactobacillus acidophilus strain INMIA 9602 Er317/402 Administration Reduces the Numbers of Candida albicans and Abundance of enterobacteria in the Gut Microbiota of Familial Mediterranean Fever Patients. Front. Immunol. 2018, 9, 1426. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.L.; Larkinson, M.L.Y.; Clarke, T.B. Immunological Design of Commensal Communities to Treat Intestinal Infection and Inflammation. PLoS Pathog. 2021, 17, e1009191. [Google Scholar] [CrossRef] [PubMed]
- Bok, E.; Mazurek, J.; Myc, A.; Stosik, M.; Wojciech, M.; Baldy-Chudzik, K. Comparison of Commensal Escherichia coli Isolates from Adults and Young Children in Lubuskie Province, Poland: Virulence Potential, Phylogeny and Antimicrobial Resistance. Int. J. Environ. Res. Public Health 2018, 15, 617. [Google Scholar] [CrossRef]
- Mazurek-Popczyk, J.; Pisarska, J.; Bok, E.; Baldy-Chudzik, K. Antibacterial Activity of Bacteriocinogenic Commensal Escherichia coli Against Zoonotic Strains Resistant and Sensitive to Antibiotics. Antibiotics 2020, 9, 411. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Vazquez, U.; Verstraeten, S.; Martin, R.; Chain, F.; Langella, P.; Thomas, M.; Cherbuy, C. The Commensal Escherichia coli CEC15 Reinforces Intestinal Defences in Gnotobiotic Mice and is Protective in a Chronic Colitis Mouse Model. Sci. Rep. 2019, 9, 11431. [Google Scholar] [CrossRef] [PubMed]
- Mirzoyan, N.S.; Pepoyan, A.Z.; Trchounian, A.H. Modification of the Biophysical Characteristics of Membranes Incommensal Escherichia coli Strains from Breast Cancer Patients. FEMS Microbiol. Lett. 2006, 254, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Stepanyan, K.; Balayan, M.H.; Vassilian, A.; Pepoyan, A.Z.; Trchounian, A.H. Growth Peculiarities and Some Characteristics of Membrane for Probiotic Strain of Escherichia coli. Memb. Cell Biol. 2007, 1, 333–335. [Google Scholar] [CrossRef]
- Balayan, M.H.; Mirzabekyan, S.S.; Pepoyan, A.Z.; Trchounian, A.H. Some Peculiarities of Growth and Functional Activity of Escherichia coli Strain from Probiotic Formula “ASAP”. World Acad. Sci. Eng. Technol. 2010, 44, 858–862. [Google Scholar]
- Gasparyan, G.; Balayan, M.H.; Grigoryan, A.; Hakopyan, A.; Manvelyan, A.M.; Mirzabekyan, S.S.; Pepoyan, A.Z. Growth Peculiarities of Commensal Escherichia coli Isolates from the Gut Microflora of Crohn’s Disease Patients. Biofizika 2013, 58, 690–696. (In Russian) [Google Scholar] [CrossRef]
- Pepoyan, A.Z.; Balayan, M.H.; Manvelyan, A.M.; Tsaturyan, V.V. Growth and Motility of Gut Commensal Escherichia coli in Health and Disease. Biophys. J. 2014, 106, 726. [Google Scholar] [CrossRef]
- Pepoyan, A.Z.; Balayan, M.H.; Manvelyan, A.M.; Mamikonyan, V.; Isajanyan, M.; Tsaturyan, V.V.; Kamiya, S.; Netrebov, V.; Chikindas, M. Lactobacillus acidophilus INMIA 9602 Er-2 strain 317/402 Probiotic Regulates Growth of Commensal Escherichia coli in Gut Microbiota of Familial Mediterranean Fever Disease Subjects. Lett. Appl. Microbiol. 2017, 64, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Pepoyan, A.Z.; Mirzoyan, N.S.; Saakyan, M.O.; Kirakosyan, L.A.; KapareзяH, K.G. Some Peculiarities of Antioxidative System of E. coli G35 Bacterial Strains. Rep. Acad. Sci. 2002, 102, 70–75. [Google Scholar]
- Pepoyan, A.Z.; Ktsoyan, Z.A.; Shaginyan, A.A.; Osipyan, L.M.; Karagezyan, K.G. The Role of Membrane Interactions in Lipid-peroxidation Processes in Plasmid and Unplasmid Cells of Salmonella derby. Biophysika. 1991, 36, 475–479. (In Russian) [Google Scholar]
- Shahinyan, A.; Garibyan, J.; Pepoyan, A.; Karapetyan, O. Cancerolitic Action of E. coli. J. Nat. Sci. 2003, 1, 53–58. [Google Scholar]
- Lowry, O.H.; Rosebrouh, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Vladimirov, Y.A.; Archakov, A.I. Lipid Peroxidation in Biological Membranes; Nauka: Moscow, Russia, 1972; p. 252. (In Russian) [Google Scholar]
- Yakovleva, V.I.; Maloreeva, I.V.; Zueva, M.N.; Andreeva, V.P.; Gubnetskiy, L.S.; Shcherbakova, V.N.; Berezin, I.V. Synthesis of L-aspartic Acid from Ammonium Fumarate by Free and Immobilized Cells of Escherichia Coli. Prikl. Biokhim. Mikrobiol. 1979, 15, 328–336. [Google Scholar]
- Leyane, T.S.; Jere, S.W.; Houreld, N.N. Oxidative Stress in Ageing and Chronic Degenerative Pathologies: Molecular Mechanisms Involved in Counteracting Oxidative Stress and Chronic Inflammation. Int. J. Mol. Sci. 2022, 23, 7273. [Google Scholar] [CrossRef]
- Imlay, J.A. The Molecular Mechanisms and Physiological Consequences of Oxidative Stress: Lessons from a Model Bacterium. Nat. Publ. Gr. 2013, 11, 443–454. [Google Scholar] [CrossRef]
- Fasnacht, M.; Polacek, N. Oxidative Stress in Bacteria and the Central Dogma of Molecular Biology. Front. Mol. Biosci. 2021, 8, 671037. [Google Scholar] [CrossRef]
- Karapetyan, A.O. Biological Antagonism of Some Representatives of the Intestinal Microflora and Tumor Cells. Ph.D. Thesis, Moscow State University, Moscow, Russia, 1987; p. 101. (In Russian). [Google Scholar]
- Shachlamov, W.A.; Karapetyan, A.O.; Karapetyan, O.G. Materials of Actual Quest. of Modern Histology; Nauka: Moscow, Russia, 1988; p. 101. (In Russian) [Google Scholar]
- Nersesyan, L.E.; Garibyan, D.V.; Garibjanyan, B.T.; Shahinyan, A.A. The Change in the Structure of Tumor DNA under the Combined Action of Bacterial Culture, E. coli and Doxorubicin A In Vivo. Electron. J. Nat. Sci. 2004, 3, 36–39. [Google Scholar]
- Manvelyan, A.; Balayan, M.; Pepoyan, E.; Aloyan, S.; Pepoyan, A. Laboratory Assessment of Gut Dysbiosis. Biol. J. RA 2007, 12, 35–39. (In Russian) [Google Scholar]
- Tang, L.; Zhou, Y.J.; Zhu, S.; Liang, G.D.; Zhuang, H.; Zhao, M.; Chang, X.Y.; Li, H.N.; Liu, Z.; Guo, Z.R.; et al. E. coli Diversity: Low in Colorectal Cancer. BMC Med. Genom. 2020, 13, 59. [Google Scholar] [CrossRef]
- Shandilya, S.; Kumar, S.; Kumar Jha, N.; Kumar Kesari, K.; Ruokolainen, J. Interplay of Gut Microbiota and Oxidative Stress: Perspective on Neurodegeneration and Neuroprotection. J. Adv. Res. 2021, 38, 223–244. [Google Scholar] [CrossRef]
- Ni, Q.; Zhang, P.; Li, Q.; Han, Z. Oxidative Stress and Gut Microbiome in Inflammatory Skin Diseases. Front. Cell Dev. Biol. 2022, 10, 849985. [Google Scholar] [CrossRef]
- Kulchitskaya, M. Development of a Probiotic Preparation Ocarin for Medicine and Veterinary Medicine. Ph.D. Thesis, Nizhny Novgorod State Agricultural Academy, Nizhny Novgorod, Russia, 2002; p. 193. [Google Scholar]
- Alizadeh, S.; Esmaeili, A.; Omidi, Y. Anti-cancer Properties of Escherichia coli Nissle 1917 against HT-29 Colon Cancer Cells through Regulation of Bax/Bcl-xL and AKT/PTEN Signaling Pathways. Iran. J. Basic Med. Sci. 2020, 23, 886–893. [Google Scholar] [CrossRef]
- Pepoyan, A.Z.; Manvelyan, A.M.; Balayan, M.H.; McCabe, G.; Tsaturyan, V.V.; Melnikov, V.G.; Chikindas, M.L.; Weeks, R.; Karlyshev, A.V. The Effectiveness of Potential Probiotics Lactobacillus rhamnosus Vahe and Lactobacillus delbrueckii IAHAHI in Irradiated Rats Depends on the Nutritional Stage of the Host. Probiotics Antimicro. Prot. 2020, 12, 1439–1450. [Google Scholar] [CrossRef]
- Harutyunyan, N.; Kushugulova, A.; Hovhannisyan, N.; Pepoyan, A. One Health Probiotics as Biocontrol Agents: One Health Tomato Probiotics. Plants 2022, 11, 1334. [Google Scholar] [CrossRef]
- Stones, D.H.; Krachler, A.M. Against the Tide: The Role of Bacterial Adhesion in Host Colonization. Biochem. Soc. Trans. 2016, 44, 1571–1580. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L.A. Host-pathogen Interactions: Basic Concepts of Microbial Commensalism, Colonization, Infection, and Disease. Infect. Immun. 2000, 68, 6511–6518. [Google Scholar] [CrossRef]
- Chautrand, T.; Souak, D.; Chevalier, S.; Duclairoir-Poc, C. Gram-Negative Bacterial Envelope Homeostasis under Oxidative and Nitrosative Stress. Microorganisms 2022, 10, 924. [Google Scholar] [CrossRef]
- Knaus, U.G.; Hertzberger, R.; Pircalabioru, G.G.; Yousefi, S.P.; Branco Dos Santos, F. Pathogen Control at the Intestinal Mucosa-H2O2 to the Rescue. Gut Microbes 2017, 8, 67–74. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strains | Logarithmic Phase of Growth | Stationary Phase of Growth | |||
|---|---|---|---|---|---|
| OD600 = 0.1 | OD600 = 0.2 | OD600 = 0.2 | OD600 = 0.4 | OD600 = 0.7 | |
| E. coli G35 N61 | 143.36 | 55.58 ± 2.78 | 37.06 ± 1.85 p * < 0.05 | 121.3 ± 2.4 | 55.02 |
| E. coli G35 N49 | 178.15 | 69.07 ± 3.45 | 46.05 ± 2.3 p * < 0.05 | 150.74 ± 1.75 | 68.37 |
| TBARs Levels | Strains | |
|---|---|---|
| E. coli G35 N61 | E. coli G35 N49 | |
| Suspension | 121.3 ± 2.4 (75.55) * | 150.74 ± 1.75 (93.4) * p < 0.05 |
| Extract | 40.72 ± 1.05 | 34.2 ± 1.23 p < 0.05 |
| Cell-free supernatant | 0.625 ± 0.08 | 0.171 ± 0.06 p < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsaturyan, V.; Poghosyan, A.; Toczyłowski, M.; Pepoyan, A. Evaluation of Malondialdehyde Levels, Oxidative Stress and Host–Bacteria Interactions: Escherichia coli and Salmonella Derby. Cells 2022, 11, 2989. https://doi.org/10.3390/cells11192989
Tsaturyan V, Poghosyan A, Toczyłowski M, Pepoyan A. Evaluation of Malondialdehyde Levels, Oxidative Stress and Host–Bacteria Interactions: Escherichia coli and Salmonella Derby. Cells. 2022; 11(19):2989. https://doi.org/10.3390/cells11192989
Chicago/Turabian StyleTsaturyan, Vardan, Armen Poghosyan, Michał Toczyłowski, and Astghik Pepoyan. 2022. "Evaluation of Malondialdehyde Levels, Oxidative Stress and Host–Bacteria Interactions: Escherichia coli and Salmonella Derby" Cells 11, no. 19: 2989. https://doi.org/10.3390/cells11192989
APA StyleTsaturyan, V., Poghosyan, A., Toczyłowski, M., & Pepoyan, A. (2022). Evaluation of Malondialdehyde Levels, Oxidative Stress and Host–Bacteria Interactions: Escherichia coli and Salmonella Derby. Cells, 11(19), 2989. https://doi.org/10.3390/cells11192989