

Mechanisms of Regulation in Intraflagellar Transport


Abstract
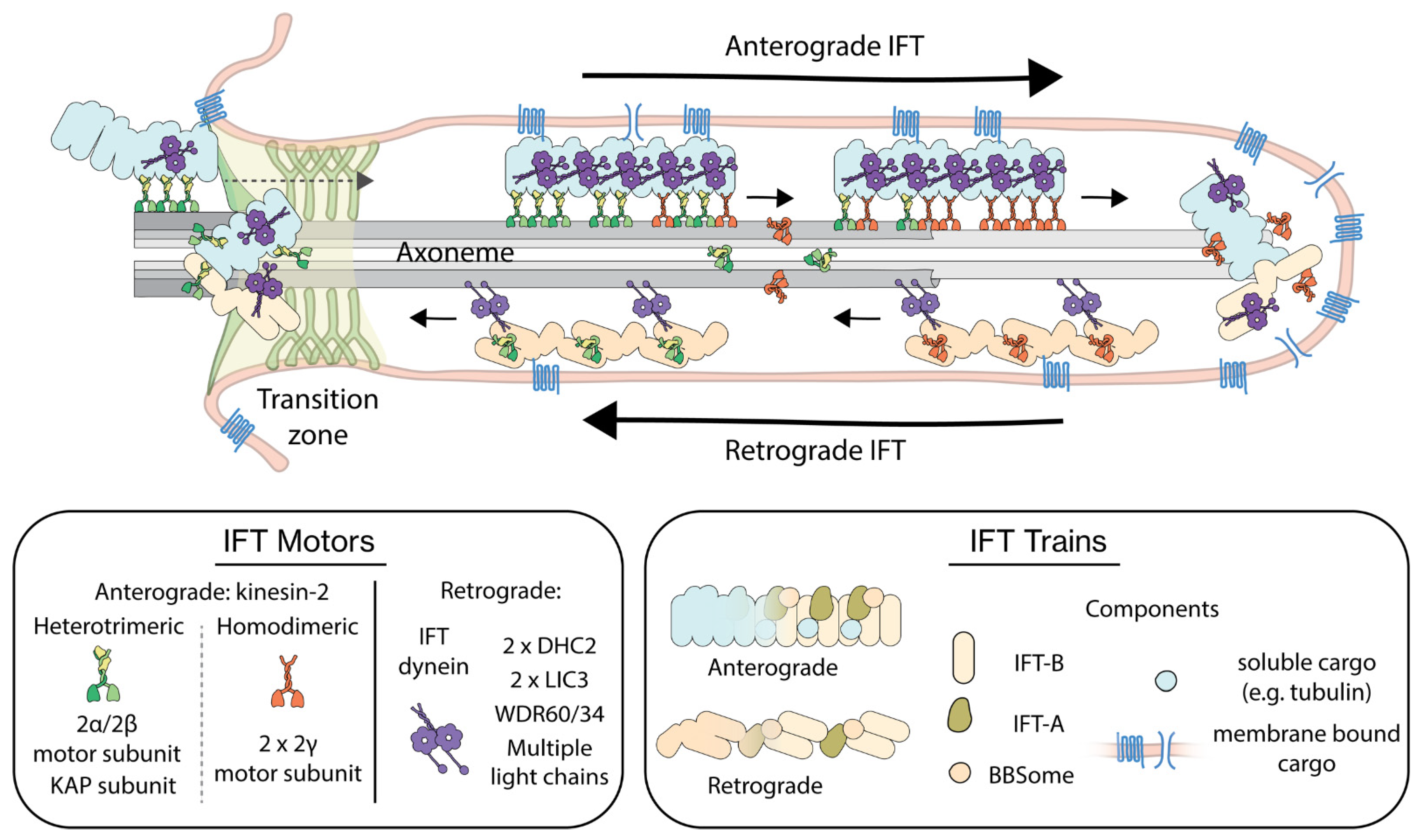
1. Introduction
2. IFT Regulation from the Perspective of the Track
2.1. The Tubulin Code
2.2. Polarity of the Axoneme and Directionality of IFT Trains
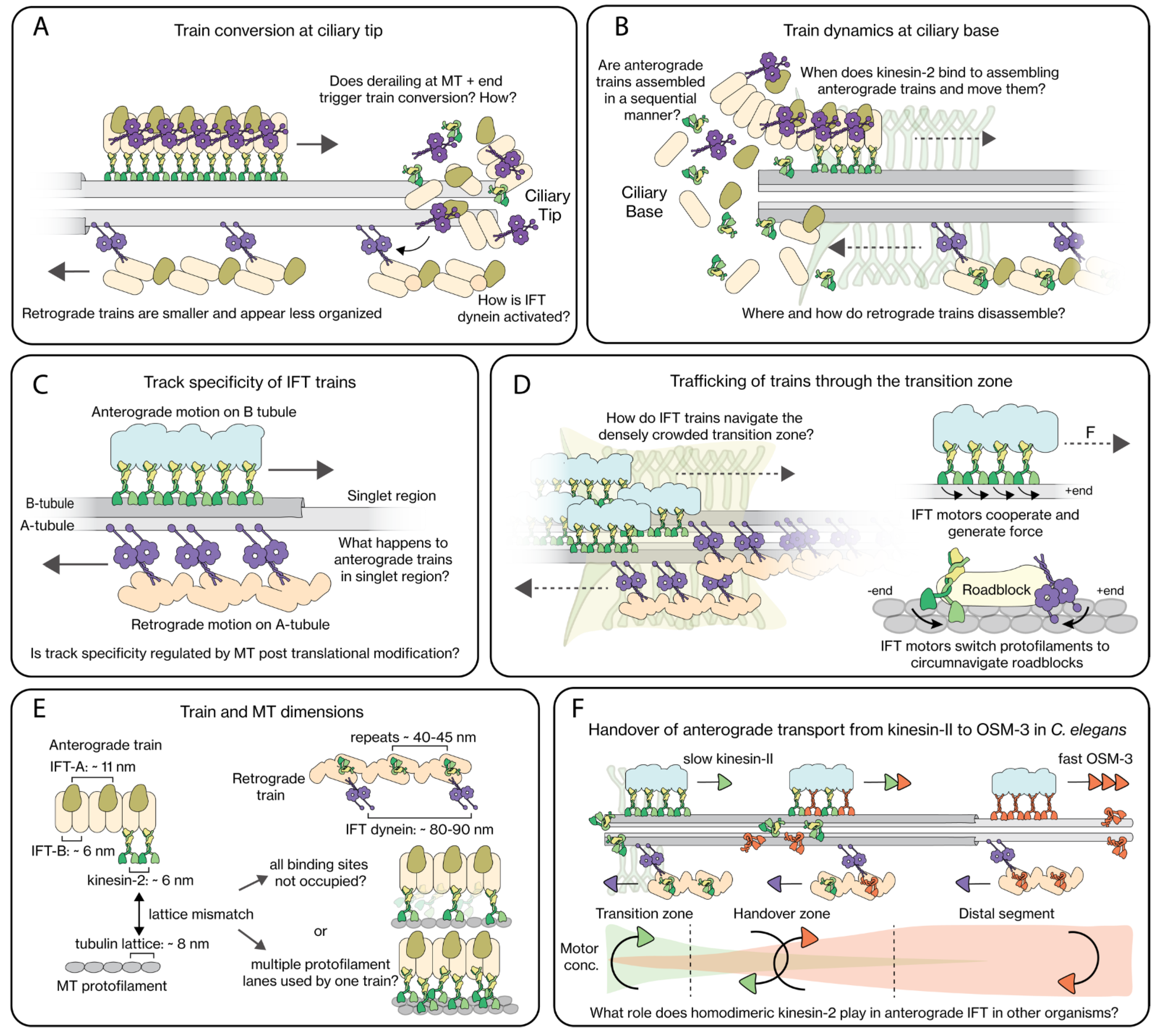
2.3. IFT-Train Turnaround at the Ciliary Tip
2.4. IFT-Train Assembly at the Ciliary Base
2.5. Track Specificity of IFT Trains
3. IFT Regulation from the Perspective of the Motors
3.1. The Properties of IFT Motors
3.2. Interactions of IFT Motors with IFT Trains
3.3. Trafficking across the Transition Zone
3.4. Collective Motor Behavior
3.5. Dynamic Interactions of IFT Motors with Trains
3.6. IFT and Variations in Axoneme Length
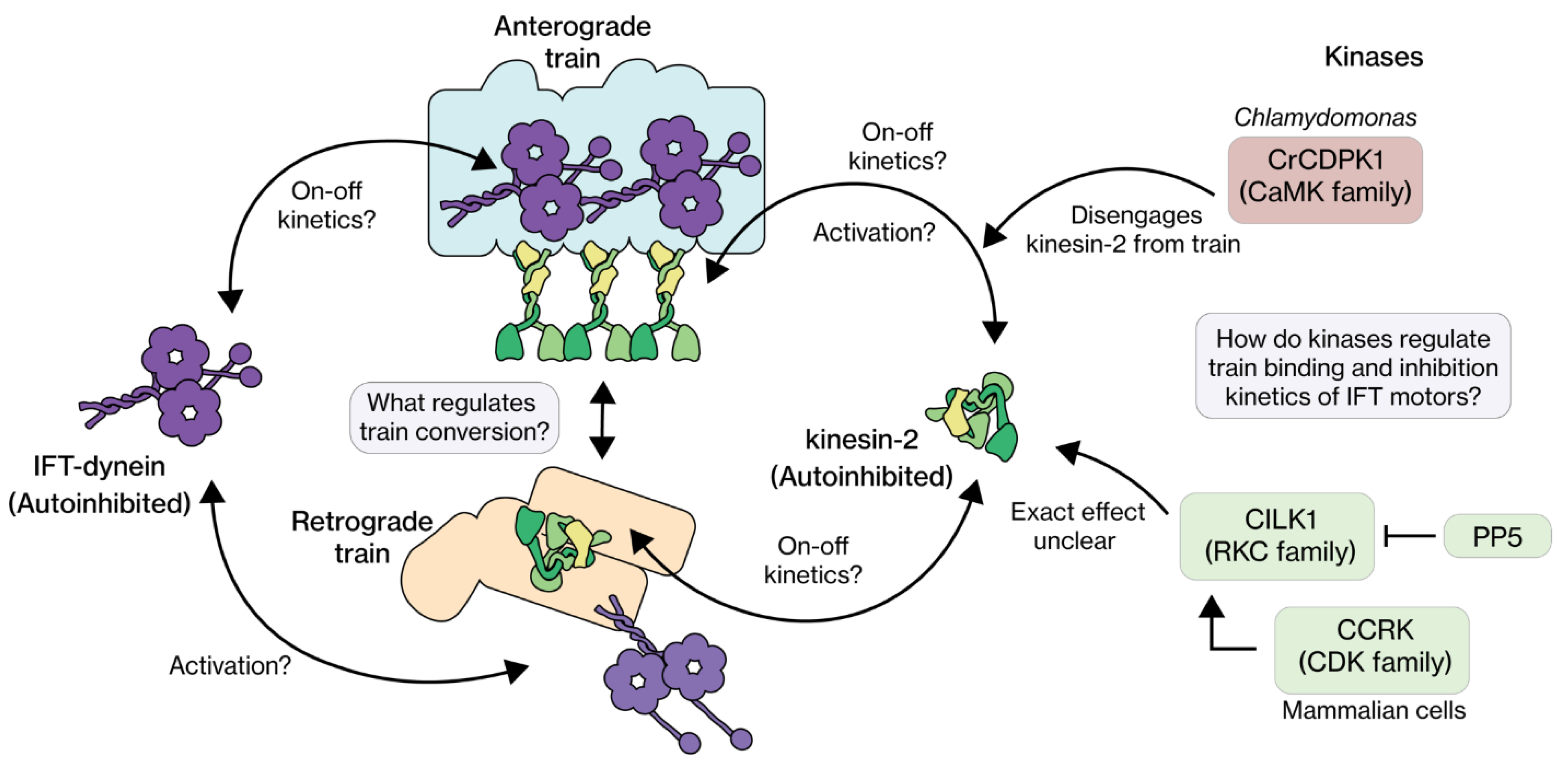
4. IFT Regulation from the Perspective of Regulatory Proteins
4.1. RCK
4.2. CDK
4.3. CaMK
4.4. CDKL
4.5. Regulatory Proteins: Concluding Remarks
5. Open Questions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobell, C. Antony van Leeuwenhoek and his “Little Animals”: Being Some Account of the Father of Protozoology and Bacteriology and his Multifarious Discoveries in these Disciplines; Collected, Translated, and Edited, from his Printed Works, Unpublished Manuscripts, and Contemporary Records. Published on the 300th Anniversary of His Birth; Harcourt, Brace and Company: New York, NY, USA, 1932; p. 119. [Google Scholar]
- Conduit, S.E.; Vanhaesebroeck, B. Phosphoinositide lipids in primary cilia biology. Biochem. J. 2020, 477, 3541–3565. [Google Scholar] [CrossRef] [PubMed]
- Cavalier-Smith, T. The phagotrophic origin of eukaryotes and phylogenetic classification of Protozoa. Int. J. Syst. Evol. Microbiol. 2002, 52, 297–354. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.R. Evolution of cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028290. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Mitchell, D.R.; Jékely, G. How did the cilium evolve? Curr. Top. Dev. Biol. 2008, 85, 63–82. [Google Scholar] [PubMed]
- Carvalho-Santos, Z.; Azimzadeh, J.; Pereira-Leal, J.B.; Bettencourt-Dias, M. Tracing the origins of centrioles, cilia, and flagella. J. Cell Biol. 2011, 195, 341. [Google Scholar] [CrossRef][Green Version]
- Lee, J.H.; Gleeson, J.G. The role of primary cilia in neuronal function. Neurobiol. Dis. 2010, 38, 167–172. [Google Scholar] [CrossRef]
- Gitten, J.; Dede, D.; Fennell, E.; Quisling, R.; Maria, B.L. Neurobehavioral development in Joubert syndrome. J. Child Neurol. 1998, 13, 391–397. [Google Scholar] [CrossRef]
- Kozminski, K.G.; Johnson, K.A.; Forscher, P.; Rosenbaum, J.L. A motility in the eukaryotic flagellum unrelated to flagellar beating. Proc. Natl. Acad. Sci. USA 1993, 90, 5519–5523. [Google Scholar] [CrossRef]
- Jordan, M.A.; Pigino, G. The structural basis of intraflagellar transport at a glance. J. Cell Sci. 2021, 134, jcs247163. [Google Scholar] [CrossRef]
- Nakamura, K.; Noguchi, T.; Takahara, M.; Omori, Y.; Furukawa, T.; Katoh, Y.; Nakayama, K. Anterograde trafficking of ciliary MAP kinase–like ICK/CILK1 by the intraflagellar transport machinery is required for intraciliary retrograde protein trafficking. J. Biol. Chem. 2020, 295, 13363–13376. [Google Scholar] [CrossRef]
- Scholey, J.M. Kinesin-2: A family of heterotrimeric and homodimeric motors with diverse intracellular transport functions. Annu. Rev. Cell Dev. Biol. 2013, 29, 443–469. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Witman, G.B. Dynein and intraflagellar transport. Exp. Cell Res. 2015, 334, 26. [Google Scholar] [CrossRef] [PubMed]
- Jordan, M.A.; Diener, D.R.; Stepanek, L.; Pigino, G. The cryo-EM structure of intraflagellar transport trains reveals how dynein is inactivated to ensure unidirectional anterograde movement in cilia. Nat. Cell Biol. 2018, 20, 1250–1255. [Google Scholar] [CrossRef]
- Bhogaraju, S.; Cajanek, L.; Fort, C.; Blisnick, T.; Weber, K.; Taschner, M.; Mizuno, N.; Lamla, S.; Bastin, P.; Nigg, E.A. Molecular basis of tubulin transport within the cilium by IFT74 and IFT81. Science 2013, 341, 1009–1012. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Ishida, Y.; Hirano, T.; Katoh, Y.; Nakayama, K. Cooperation of the IFT-A complex with the IFT-B complex is required for ciliary retrograde protein trafficking and GPCR import. Mol. Biol. Cell 2021, 32, 45–56. [Google Scholar] [CrossRef]
- Lechtreck, K.F.; Van De Weghe, J.C.; Harris, J.A.; Liu, P. Protein transport in growing and steady-state cilia. Traffic 2017, 18, 277–286. [Google Scholar] [CrossRef]
- Vincensini, L.; Blisnick, T.; Bastin, P. 1001 model organisms to study cilia and flagella. Biol. Cell 2011, 103, 109–130. [Google Scholar] [CrossRef]
- Zehr, E.A.; Roll-Mecak, A. A look under the hood of the machine that makes cilia beat. Nat. Struct. Mol. Biol. 2022, 29, 416–418. [Google Scholar] [CrossRef]
- Nechipurenko, I.V.; Sengupta, P. The rise and fall of basal bodies in the nematode Caenorhabditis elegans. Cilia 2017, 6, 9. [Google Scholar] [CrossRef]
- Soares, H.; Carmona, B.; Nolasco, S.; Viseu Melo, L.; Goncalves, J. Cilia Distal Domain: Diversity in Evolutionarily Conserved Structures. Cells 2019, 8, 160. [Google Scholar] [CrossRef]
- Chien, A.; Shih, S.M.; Bower, R.; Tritschler, D.; Porter, M.E.; Yildiz, A. Dynamics of the IFT machinery at the ciliary tip. Elife 2017, 6, e28606. [Google Scholar] [CrossRef] [PubMed]
- Prevo, B.; Scholey, J.M.; Peterman, E.J.G. Intraflagellar transport: Mechanisms of motor action, cooperation, and cargo delivery. FEBS J. 2017, 284, 2905–2931. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Katoh, Y. Architecture of the IFT ciliary trafficking machinery and interplay between its components. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 179–196. [Google Scholar] [CrossRef] [PubMed]
- Webb, S.; Mukhopadhyay, A.G.; Roberts, A.J. Intraflagellar transport trains and motors: Insights from structure. Semin. Cell Dev. Biol. 2020, 107, 82–90. [Google Scholar] [CrossRef]
- Scholey, J.M. Intraflagellar transport motors in cilia: Moving along the cell’s antenna. J. Cell Biol. 2008, 180, 23–29. [Google Scholar] [CrossRef]
- Vuolo, L.; Stevenson, N.L.; Mukhopadhyay, A.G.; Roberts, A.J.; Stephens, D.J. Cytoplasmic dynein-2 at a glance. J. Cell Sci. 2020, 133, jcs240614. [Google Scholar] [CrossRef]
- Chaya, T.; Furukawa, T. Post-translational modification enzymes as key regulators of ciliary protein trafficking. J. Biochem. 2021, 169, 633–642. [Google Scholar] [CrossRef]
- Avasthi, P.; Marshall, W.F. Stages of ciliogenesis and regulation of ciliary length. Differentiation 2012, 83, S30–S42. [Google Scholar] [CrossRef]
- Ishikawa, H.; Marshall, W.F. Ciliogenesis: Building the cell’s antenna. Nat. Rev. Mol. Cell Biol. 2011, 12, 222–234. [Google Scholar] [CrossRef]
- Santos, N.; Reiter, J.F. Building it up and taking it down: The regulation of vertebrate ciliogenesis. Dev. Dyn. 2008, 237, 1972–1981. [Google Scholar] [CrossRef]
- Bruggeman, C.W.; Haasnoot, G.H.; Danne, N.; van Krugten, J.; Peterman, E.J.G. Differentiated dynamic response in C. elegans chemosensory cilia. bioRxiv 2022. [Google Scholar] [CrossRef]
- Nicastro, D.; Fu, X.; Heuser, T.; Tso, A.; Porter, M.E.; Linck, R.W. Cryo-electron tomography reveals conserved features of doublet microtubules in flagella. Proc. Natl. Acad. Sci. USA 2011, 108, E845–E853. [Google Scholar] [CrossRef] [PubMed]
- Sui, H.; Downing, K.H. Molecular architecture of axonemal microtubule doublets revealed by cryo-electron tomography. Nature 2006, 442, 475–478. [Google Scholar] [CrossRef]
- Williams, C.L.; McIntyre, J.C.; Norris, S.R.; Jenkins, P.M.; Zhang, L.; Pei, Q.; Verhey, K.; Martens, J.R. Direct evidence for BBSome-associated intraflagellar transport reveals distinct properties of native mammalian cilia. Nat. Commun. 2014, 5, 5813. [Google Scholar] [CrossRef] [PubMed]
- Perkins, L.A.; Hedgecock, E.M.; Thomson, J.N.; Culotti, J.G. Mutant sensory cilia in the nematode Caenorhabditis elegans. Dev. Biol. 1986, 117, 456–487. [Google Scholar] [CrossRef]
- Nievergelt, A.P.; Zykov, I.; Diener, D.; Chhatre, A.; Buchholz, T.-O.; Delling, M.; Diez, S.; Jug, F.; Štěpánek, L.; Pigino, G. Conversion of anterograde into retrograde trains is an intrinsic property of intraflagellar transport. Curr. Biol. 2022; in press. [Google Scholar] [CrossRef]
- Vaughan, S.; Gull, K. Basal body structure and cell cycle-dependent biogenesis in Trypanosoma brucei. Cilia 2015, 5, 5. [Google Scholar] [CrossRef]
- Gui, M.; Ma, M.; Sze-Tu, E.; Wang, X.; Koh, F.; Zhong, E.D.; Berger, B.; Davis, J.H.; Dutcher, S.K.; Zhang, R.; et al. Structures of radial spokes and associated complexes important for ciliary motility. Nat. Struct. Mol. Biol. 2021, 28, 29–37. [Google Scholar] [CrossRef]
- Gui, M.; Wang, X.; Dutcher, S.K.; Brown, A.; Zhang, R. Ciliary central apparatus structure reveals mechanisms of microtubule patterning. Nat. Struct. Mol. Biol. 2022, 29, 483–492. [Google Scholar] [CrossRef]
- Han, L.; Rao, Q.; Yang, R.; Wang, Y.; Chai, P.; Xiong, Y.; Zhang, K. Cryo-EM structure of an active central apparatus. Nat. Struct. Mol. Biol. 2022, 29, 472–482. [Google Scholar] [CrossRef]
- Ma, M.; Stoyanova, M.; Rademacher, G.; Dutcher, S.K.; Brown, A.; Zhang, R. Structure of the Decorated Ciliary Doublet Microtubule. Cell 2019, 179, 909–922 e912. [Google Scholar] [CrossRef] [PubMed]
- Janke, C.; Magiera, M.M. The tubulin code and its role in controlling microtubule properties and functions. Nat. Rev. Mol. Cell Biol. 2020, 21, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Roll-Mecak, A. The Tubulin Code in Microtubule Dynamics and Information Encoding. Dev. Cell 2020, 54, 7–20. [Google Scholar] [CrossRef]
- Thazhath, R.; Liu, C.; Gaertig, J. Polyglycylation domain of β-tubulin maintains axonemal architecture and affects cytokinesis in Tetrahymena. Nat. Cell Biol. 2002, 4, 256–259. [Google Scholar] [CrossRef]
- Sirajuddin, M.; Rice, L.M.; Vale, R.D. Regulation of microtubule motors by tubulin isotypes and post-translational modifications. Nat. Cell Biol. 2014, 16, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Akella, J.S.; Barr, M.M. The tubulin code specializes neuronal cilia for extracellular vesicle release. Dev. Neurobiol. 2021, 81, 231–252. [Google Scholar] [CrossRef]
- Hao, L.; Thein, M.; Brust-Mascher, I.; Civelekoglu-Scholey, G.; Lu, Y.; Acar, S.; Prevo, B.; Shaham, S.; Scholey, J.M. Intraflagellar transport delivers tubulin isotypes to sensory cilium middle and distal segments. Nat. Cell Biol. 2011, 13, 790–798. [Google Scholar] [CrossRef]
- Hurd, D.D.; Miller, R.M.; Nunez, L.; Portman, D.S. Specific alpha- and beta-tubulin isotypes optimize the functions of sensory Cilia in Caenorhabditis elegans. Genetics 2010, 185, 883–896. [Google Scholar] [CrossRef]
- Kimura, Y.; Tsutsumi, K.; Konno, A.; Ikegami, K.; Hameed, S.; Kaneko, T.; Kaplan, O.I.; Teramoto, T.; Fujiwara, M.; Ishihara, T.; et al. Environmental responsiveness of tubulin glutamylation in sensory cilia is regulated by the p38 MAPK pathway. Sci. Rep. 2018, 8, 8392. [Google Scholar] [CrossRef]
- Power, K.M.; Akella, J.S.; Gu, A.; Walsh, J.D.; Bellotti, S.; Morash, M.; Zhang, W.; Ramadan, Y.H.; Ross, N.; Golden, A.; et al. Mutation of NEKL-4/NEK10 and TTLL genes suppress neuronal ciliary degeneration caused by loss of CCPP-1 deglutamylase function. PLoS Genet. 2020, 16, e1009052. [Google Scholar] [CrossRef]
- Lechtreck, K.F.; Geimer, S. Distribution of polyglutamylated tubulin in the flagellar apparatus of green flagellates. Cell Motil. Cytoskelet. 2000, 47, 219–235. [Google Scholar] [CrossRef]
- Hancock, W.O. Bidirectional cargo transport: Moving beyond tug of war. Nat. Rev. Mol. Cell Biol. 2014, 15, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, J.L.; Mekonnen, B.; Mengoni, I.; Liu, P.; Jordan, M.; Diener, D.; Pigino, G.; Lechtreck, K. In vivo imaging shows continued association of several IFT-A, IFT-B and dynein complexes while IFT trains U-turn at the tip. J. Cell Sci. 2021, 134, jcs259010. [Google Scholar] [CrossRef] [PubMed]
- Mijalkovic, J.; van Krugten, J.; Oswald, F.; Acar, S.; Peterman, E.J.G. Single-Molecule Turnarounds of Intraflagellar Transport at the C. elegans Ciliary Tip. Cell Rep. 2018, 25, 1701–1707.e2. [Google Scholar] [CrossRef]
- Dentler, W.L. Structures linking the tips of ciliary and flagellar microtubules to the membrane. J. Cell Sci. 1980, 42, 207–220. [Google Scholar] [CrossRef]
- Miller, J.M.; Wang, W.; Balczon, R.; Dentler, W.L. Ciliary microtubule capping structures contain a mammalian kinetochore antigen. J. Cell Biol. 1990, 110, 703–714. [Google Scholar] [CrossRef]
- Chaya, T.; Omori, Y.; Kuwahara, R.; Furukawa, T. ICK is essential for cell type-specific ciliogenesis and the regulation of ciliary transport. EMBO J. 2014, 33, 1227–1242. [Google Scholar] [CrossRef]
- Buisson, J.; Chenouard, N.; Lagache, T.; Blisnick, T.; Olivo-Marin, J.C.; Bastin, P. Intraflagellar transport proteins cycle between the flagellum and its base. J. Cell Sci. 2013, 126, 327–338. [Google Scholar] [CrossRef]
- Hibbard, J.V.K.; Vazquez, N.; Satija, R.; Wallingford, J.B. Protein turnover dynamics suggest a diffusion-to-capture mechanism for peri-basal body recruitment and retention of intraflagellar transport proteins. Mol. Biol. Cell 2021, 32, 1171–1180. [Google Scholar] [CrossRef]
- Prevo, B.; Mangeol, P.; Oswald, F.; Scholey, J.M.; Peterman, E.J.G. Functional differentiation of cooperating kinesin-2 motors orchestrates cargo import and transport in C. elegans cilia. Nat. Cell Biol. 2015, 17, 1536–1545. [Google Scholar] [CrossRef]
- Wingfield, J.L.; Mengoni, I.; Bomberger, H.; Jiang, Y.Y.; Walsh, J.D.; Brown, J.M.; Picariello, T.; Cochran, D.A.; Zhu, B.; Pan, J.; et al. IFT trains in different stages of assembly queue at the ciliary base for consecutive release into the cilium. Elife 2017, 6, e26609. [Google Scholar] [CrossRef] [PubMed]
- Van den Hoek, H.; Klena, N.; Jordan, M.A.; Alvarez Viar, G.; Righetto, R.D.; Schaffer, M.; Erdmann, P.S.; Wan, W.; Geimer, S.; Plitzko, J.M. In situ architecture of the ciliary base reveals the stepwise assembly of intraflagellar transport trains. Science 2022, 377, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Blacque, O.E.; Sanders, A.A. Compartments within a compartment: What C. elegans can tell us about ciliary subdomain composition, biogenesis, function, and disease. Organogenesis 2014, 10, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Stepanek, L.; Pigino, G. Microtubule doublets are double-track railways for intraflagellar transport trains. Science 2016, 352, 721–724. [Google Scholar] [CrossRef]
- Maheshwari, A.; Obbineni, J.M.; Bui, K.H.; Shibata, K.; Toyoshima, Y.Y.; Ishikawa, T. alpha- and beta-Tubulin Lattice of the Axonemal Microtubule Doublet and Binding Proteins Revealed by Single Particle Cryo-Electron Microscopy and Tomography. Structure 2015, 23, 1584–1595. [Google Scholar] [CrossRef]
- Johnson, K.A. The axonemal microtubules of the Chlamydomonas flagellum differ in tubulin isoform content. J. Cell Sci. 1998, 111 Pt 3, 313–320. [Google Scholar] [CrossRef]
- Stepp, W.L.; Merck, G.; Mueller-Planitz, F.; Okten, Z. Kinesin-2 motors adapt their stepping behavior for processive transport on axonemes and microtubules. EMBO Rep. 2017, 18, 1947–1956. [Google Scholar] [CrossRef]
- Xie, C.; Li, L.; Li, M.; Shao, W.; Zuo, Q.; Huang, X.; Chen, R.; Li, W.; Brunnbauer, M.; Okten, Z.; et al. Optimal sidestepping of intraflagellar transport kinesins regulates structure and function of sensory cilia. EMBO J. 2020, 39, e103955. [Google Scholar] [CrossRef]
- Kiesel, P.; Alvarez Viar, G.; Tsoy, N.; Maraspini, R.; Gorilak, P.; Varga, V.; Honigmann, A.; Pigino, G. The molecular structure of mammalian primary cilia revealed by cryo-electron tomography. Nat. Struct. Mol. Biol. 2020, 27, 1115–1124. [Google Scholar] [CrossRef]
- Bertiaux, E.; Mallet, A.; Fort, C.; Blisnick, T.; Bonnefoy, S.; Jung, J.; Lemos, M.; Marco, S.; Vaughan, S.; Trepout, S.; et al. Bidirectional intraflagellar transport is restricted to two sets of microtubule doublets in the trypanosome flagellum. J. Cell Biol. 2018, 217, 4284–4297. [Google Scholar] [CrossRef]
- Ou, G.; Scholey, J.M. Motor Cooperation during Mitosis and Ciliogenesis. Annu. Rev. Cell Dev. Biol. 2022, 38. [Google Scholar] [CrossRef] [PubMed]
- Snow, J.J.; Ou, G.; Gunnarson, A.L.; Walker, M.R.S.; Zhou, H.M.; Brust-Mascher, I.; Scholey, J.M. Two anterograde intraflagellar transport motors cooperate to build sensory cilia on C. elegans neurons. Nat. Cell Biol. 2004, 6, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Pang, Y.; Wu, Q.; Hu, Z.; Han, X.; Xu, Y.; Deng, H.; Pan, J. FLA8/KIF3B phosphorylation regulates kinesin-II interaction with IFT-B to control IFT entry and turnaround. Dev. Cell 2014, 30, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Tanaka, Y.; Okada, Y.; Takeda, S.; Harada, A.; Kanai, Y.; Kido, M.; Hirokawa, N. Randomization of left–right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein. Cell 1998, 95, 829–837. [Google Scholar] [CrossRef]
- Miller, M.S.; Esparza, J.M.; Lippa, A.M.; Lux III, F.G.; Cole, D.G.; Dutcher, S.K. Mutant kinesin-2 motor subunits increase chromosome loss. Mol. Biol. Cell 2005, 16, 3810–3820. [Google Scholar] [CrossRef]
- Chana, M.; Tripet, B.P.; Mant, C.T.; Hodges, R.S. The role of unstructured highly charged regions on the stability and specificity of dimerization of two-stranded alpha-helical coiled-coils: Analysis of the neck-hinge region of the kinesin-like motor protein Kif3A. J. Struct. Biol. 2002, 137, 206–219. [Google Scholar] [CrossRef]
- Vukajlovic, M.; Dietz, H.; Schliwa, M.; Okten, Z. How kinesin-2 forms a stalk. Mol. Biol. Cell 2011, 22, 4279–4287. [Google Scholar] [CrossRef]
- Oh, Y.S.; Wang, E.J.; Gailey, C.D.; Brautigan, D.L.; Allen, B.L.; Fu, Z. Ciliopathy-associated protein kinase ICK requires its non-catalytic carboxyl-terminal domain for regulation of ciliogenesis. Cells 2019, 8, 677. [Google Scholar] [CrossRef]
- Doodhi, H.; Ghosal, D.; Krishnamurthy, M.; Jana, S.C.; Shamala, D.; Bhaduri, A.; Sowdhamini, R.; Ray, K. KAP, the accessory subunit of kinesin-2, binds the predicted coiled-coil stalk of the motor subunits. Biochemistry 2009, 48, 2248–2260. [Google Scholar] [CrossRef]
- De Marco, V.; De Marco, A.; Goldie, K.N.; Correia, J.J.; Hoenger, A. Dimerization properties of a Xenopus laevis kinesin-II carboxy-terminal stalk fragment. EMBO Rep. 2003, 4, 717–722. [Google Scholar] [CrossRef]
- Funabashi, T.; Katoh, Y.; Okazaki, M.; Sugawa, M.; Nakayama, K. Interaction of heterotrimeric kinesin-II with IFT-B-connecting tetramer is crucial for ciliogenesis. J. Cell Biol. 2018, 217, 2867–2876. [Google Scholar] [CrossRef] [PubMed]
- Mueller, J.; Perrone, C.A.; Bower, R.; Cole, D.G.; Porter, M.E. The FLA3 KAP subunit is required for localization of kinesin-2 to the site of flagellar assembly and processive anterograde intraflagellar transport. Mol. Biol. Cell 2005, 16, 1341–1354. [Google Scholar] [CrossRef] [PubMed]
- Brunnbauer, M.; Mueller-Planitz, F.; Kosem, S.; Ho, T.H.; Dombi, R.; Gebhardt, J.C.; Rief, M.; Okten, Z. Regulation of a heterodimeric kinesin-2 through an unprocessive motor domain that is turned processive by its partner. Proc. Natl. Acad. Sci. USA 2010, 107, 10460–10465. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Acar, S.; Scholey, J.M. Torque generation by one of the motor subunits of heterotrimeric kinesin-2. Biochem. Biophys. Res. Commun. 2010, 401, 53–57. [Google Scholar] [CrossRef]
- Andreasson, J.O.; Shastry, S.; Hancock, W.O.; Block, S.M. The Mechanochemical Cycle of Mammalian Kinesin-2 KIF3A/B under Load. Curr. Biol. 2015, 25, 1166–1175. [Google Scholar] [CrossRef]
- Dentler, W. Intraflagellar transport (IFT) during assembly and disassembly of Chlamydomonas flagella. J. Cell Biol. 2005, 170, 649–659. [Google Scholar] [CrossRef]
- Broekhuis, J.R.; Verhey, K.J.; Jansen, G. Regulation of cilium length and intraflagellar transport by the RCK-kinases ICK and MOK in renal epithelial cells. PLoS ONE 2014, 9, e108470. [Google Scholar] [CrossRef]
- Engelke, M.F.; Waas, B.; Kearns, S.E.; Suber, A.; Boss, A.; Allen, B.L.; Verhey, K.J. Acute inhibition of heterotrimeric kinesin-2 function reveals mechanisms of intraflagellar transport in mammalian cilia. Curr. Biol. 2019, 29, 1137–1148. e1134. [Google Scholar] [CrossRef]
- Sonar, P.; Youyen, W.; Cleetus, A.; Wisanpitayakorn, P.; Mousavi, S.I.; Stepp, W.L.; Hancock, W.O.; Tuzel, E.; Okten, Z. Kinesin-2 from C. reinhardtii Is an Atypically Fast and Auto-inhibited Motor that Is Activated by Heterotrimerization for Intraflagellar Transport. Curr. Biol. 2020, 30, 1160–1166.e5. [Google Scholar] [CrossRef]
- Li, S.; Wan, K.Y.; Chen, W.; Tao, H.; Liang, X.; Pan, J. Functional exploration of heterotrimeric kinesin-II in IFT and ciliary length control in Chlamydomonas. Elife 2020, 9, e58868. [Google Scholar] [CrossRef]
- Friedman, D.S.; Vale, R.D. Single-molecule analysis of kinesin motility reveals regulation by the cargo-binding tail domain. Nat. Cell Biol. 1999, 1, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Ou, G.; Civelekoglu-Scholey, G.; Blacque, O.E.; Endres, N.F.; Tao, L.; Mogilner, A.; Leroux, M.R.; Vale, R.D.; Scholey, J.M. Mechanism of transport of IFT particles in C. elegans cilia by the concerted action of kinesin-II and OSM-3 motors. J. Cell Biol. 2006, 174, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Oswald, F.; Prevo, B.; Acar, S.; Peterman, E.J.G. Interplay between Ciliary Ultrastructure and IFT-Train Dynamics Revealed by Single-Molecule Super-resolution Imaging. Cell Rep. 2018, 25, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Craft, J.M.; Harris, J.A.; Hyman, S.; Kner, P.; Lechtreck, K.F. Tubulin transport by IFT is upregulated during ciliary growth by a cilium-autonomous mechanism. J. Cell Biol. 2015, 208, 223–237. [Google Scholar] [CrossRef]
- Craft Van De Weghe, J.; Harris, J.A.; Kubo, T.; Witman, G.B.; Lechtreck, K.F. Diffusion rather than intraflagellar transport likely provides most of the tubulin required for axonemal assembly in Chlamydomonas. J. Cell Sci. 2020, 133, jcs249805. [Google Scholar] [CrossRef]
- Kubo, T.; Brown, J.M.; Bellve, K.; Craige, B.; Craft, J.M.; Fogarty, K.; Lechtreck, K.F.; Witman, G.B. Together, the IFT81 and IFT74 N-termini form the main module for intraflagellar transport of tubulin. J. Cell Sci. 2016, 129, 2106–2119. [Google Scholar]
- Jiang, L.; Tam, B.M.; Ying, G.; Wu, S.; Hauswirth, W.W.; Frederick, J.M.; Moritz, O.L.; Baehr, W. Kinesin family 17 (osmotic avoidance abnormal-3) is dispensable for photoreceptor morphology and function. FASEB J. 2015, 29, 4866–4880. [Google Scholar] [CrossRef]
- Funabashi, T.; Katoh, Y.; Michisaka, S.; Terada, M.; Sugawa, M.; Nakayama, K. Ciliary entry of KIF17 is dependent on its binding to the IFT-B complex via IFT46-IFT56 as well as on its nuclear localization signal. Mol. Biol. Cell 2017, 28, 624–633. [Google Scholar] [CrossRef]
- Hammond, J.W.; Blasius, T.L.; Soppina, V.; Cai, D.; Verhey, K.J. Autoinhibition of the kinesin-2 motor KIF17 via dual intramolecular mechanisms. J. Cell Biol. 2010, 189, 1013–1025. [Google Scholar] [CrossRef]
- Imanishi, M.; Endres, N.F.; Gennerich, A.; Vale, R.D. Autoinhibition regulates the motility of the C. elegans intraflagellar transport motor OSM-3. J. Cell Biol. 2006, 174, 931–937. [Google Scholar] [CrossRef]
- Mohamed, M.A.A.; Stepp, W.L.; Okten, Z. Reconstitution reveals motor activation for intraflagellar transport. Nature 2018, 557, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Leaf, A.; Von Zastrow, M. Dopamine receptors reveal an essential role of IFT-B, KIF17, and Rab23 in delivering specific receptors to primary cilia. Elife 2015, 4, e06996. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, P.M.; Hurd, T.W.; Zhang, L.; McEwen, D.P.; Brown, R.L.; Margolis, B.; Verhey, K.J.; Martens, J.R. Ciliary targeting of olfactory CNG channels requires the CNGB1b subunit and the kinesin-2 motor protein, KIF17. Curr. Biol. 2006, 16, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Verhey, K.J.; Hammond, J.W. Traffic control: Regulation of kinesin motors. Nat. Rev. Mol. Cell Biol. 2009, 10, 765–777. [Google Scholar] [CrossRef]
- Hirokawa, N.; Takemura, R. Kinesin superfamily proteins and their various functions and dynamics. Exp. Cell Res. 2004, 301, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Verhey, K.J.; Dishinger, J.; Kee, H.L. Kinesin motors and primary cilia. Biochem. Soc. Trans. 2011, 39, 1120–1125. [Google Scholar] [CrossRef]
- Roberts, A.J. Emerging mechanisms of dynein transport in the cytoplasm versus the cilium. Biochem. Soc. Trans. 2018, 46, 967–982. [Google Scholar] [CrossRef]
- Reck-Peterson, S.L.; Redwine, W.B.; Vale, R.D.; Carter, A.P. The cytoplasmic dynein transport machinery and its many cargoes. Nat. Rev. Mol. Cell Biol. 2018, 19, 382–398. [Google Scholar] [CrossRef]
- Signor, D.; Wedaman, K.P.; Orozco, J.T.; Dwyer, N.D.; Bargmann, C.I.; Rose, L.S.; Scholey, J.M. Role of a class DHC1b dynein in retrograde transport of IFT motors and IFT raft particles along cilia, but not dendrites, in chemosensory neurons of living Caenorhabditis elegans. J. Cell Biol. 1999, 147, 519–530. [Google Scholar] [CrossRef]
- Palmer, K.J.; Hughes, H.; Stephens, D.J. Specificity of cytoplasmic dynein subunits in discrete membrane-trafficking steps. Mol. Biol. Cell 2009, 20, 2885–2899. [Google Scholar] [CrossRef]
- Pazour, G.J.; Dickert, B.L.; Witman, G.B. The DHC1b (DHC2) isoform of cytoplasmic dynein is required for flagellar assembly. J. Cell Biol. 1999, 144, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Asante, D.; Stevenson, N.L.; Stephens, D.J. Subunit composition of the human cytoplasmic dynein-2 complex. J. Cell Sci. 2014, 127, 4774–4787. [Google Scholar] [CrossRef] [PubMed]
- Toropova, K.; Zalyte, R.; Mukhopadhyay, A.G.; Mladenov, M.; Carter, A.P.; Roberts, A.J. Structure of the dynein-2 complex and its assembly with intraflagellar transport trains. Nat. Struct. Mol. Biol. 2019, 26, 823–829. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, J.; Li, S.; Lechtreck, K.; Pan, J. IFT54 directly interacts with kinesin-II and IFT dynein to regulate anterograde intraflagellar transport. EMBO J. 2021, 40, e105781. [Google Scholar] [CrossRef] [PubMed]
- Ou, G.; Blacque, O.E.; Snow, J.J.; Leroux, M.R.; Scholey, J.M. Functional coordination of intraflagellar transport motors. Nature 2005, 436, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Cleetus, A.; Merck, G.; Mueller-Planitz, F.; Okten, Z. The physiological cargo adaptor of kinesin-2 functions as an evolutionary conserved lockpick. Proc. Natl. Acad. Sci. USA 2022, 119, e2109378119. [Google Scholar] [CrossRef] [PubMed]
- Williamson, S.M.; Silva, D.A.; Richey, E.; Qin, H. Probing the role of IFT particle complex A and B in flagellar entry and exit of IFT-dynein in Chlamydomonas. Protoplasma 2012, 249, 851–856. [Google Scholar] [CrossRef]
- Nachury, M.V.; Seeley, E.S.; Jin, H. Trafficking to the ciliary membrane: How to get across the periciliary diffusion barrier? Annu. Rev. Cell Dev. Biol. 2010, 26, 59–87. [Google Scholar] [CrossRef]
- Garcia-Gonzalo, F.R.; Reiter, J.F. Open Sesame: How Transition Fibers and the Transition Zone Control Ciliary Composition. Cold Spring Harb. Perspect. Biol. 2017, 9, a028134. [Google Scholar] [CrossRef]
- Wood, C.R.; Huang, K.; Diener, D.R.; Rosenbaum, J.L. The cilium secretes bioactive ectosomes. Curr. Biol. 2013, 23, 906–911. [Google Scholar] [CrossRef]
- Wang, J.; Barr, M.M. Ciliary Extracellular Vesicles: Txt Msg Organelles. Cell Mol. Neurobiol. 2016, 36, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Deane, J.A.; Cole, D.G.; Seeley, E.S.; Diener, D.R.; Rosenbaum, J.L. Localization of intraflagellar transport protein IFT52 identifies basal body transitional fibers as the docking site for IFT particles. Curr. Biol. 2001, 11, 1586–1590. [Google Scholar] [CrossRef]
- Brunnbauer, M.; Dombi, R.; Ho, T.H.; Schliwa, M.; Rief, M.; Okten, Z. Torque generation of kinesin motors is governed by the stability of the neck domain. Mol. Cell 2012, 46, 147–158. [Google Scholar] [CrossRef]
- Mitra, A.; Ruhnow, F.; Girardo, S.; Diez, S. Directionally biased sidestepping of Kip3/kinesin-8 is regulated by ATP waiting time and motor-microtubule interaction strength. Proc. Natl. Acad. Sci. USA 2018, 115, E7950–E7959. [Google Scholar] [CrossRef] [PubMed]
- Hoeprich, G.J.; Thompson, A.R.; McVicker, D.P.; Hancock, W.O.; Berger, C.L. Kinesin’s neck-linker determines its ability to navigate obstacles on the microtubule surface. Biophys. J. 2014, 106, 1691–1700. [Google Scholar] [CrossRef]
- Kushwaha, V.S.; Acar, S.; Miedema, D.M.; Denisov, D.V.; Schall, P.; Peterman, E.J.G. The crowding dynamics of the motor protein kinesin-II. PLoS ONE 2020, 15, e0228930. [Google Scholar] [CrossRef]
- Can, S.; Dewitt, M.A.; Yildiz, A. Bidirectional helical motility of cytoplasmic dynein around microtubules. Elife 2014, 3, e03205. [Google Scholar] [CrossRef]
- Mitra, A.; Ruhnow, F.; Nitzsche, B.; Diez, S. Impact-Free Measurement of Microtubule Rotations on Kinesin and Cytoplasmic-Dynein Coated Surfaces. PLoS ONE 2015, 10, e0136920. [Google Scholar] [CrossRef]
- Ferro, L.S.; Can, S.; Turner, M.A.; ElShenawy, M.M.; Yildiz, A. Kinesin and dynein use distinct mechanisms to bypass obstacles. Elife 2019, 8, e48629. [Google Scholar] [CrossRef]
- Tanenbaum, M.E.; Vale, R.D.; McKenney, R.J. Cytoplasmic dynein crosslinks and slides anti-parallel microtubules using its two motor domains. Elife 2013, 2, e00943. [Google Scholar] [CrossRef]
- Qiu, W.; Derr, N.D.; Goodman, B.S.; Villa, E.; Wu, D.; Shih, W.; Reck-Peterson, S.L. Dynein achieves processive motion using both stochastic and coordinated stepping. Nat. Struct. Mol. Biol. 2012, 19, 193–200. [Google Scholar] [CrossRef]
- Milic, B.; Andreasson, J.O.L.; Hogan, D.W.; Block, S.M. Intraflagellar transport velocity is governed by the number of active KIF17 and KIF3AB motors and their motility properties under load. Proc. Natl. Acad. Sci. USA 2017, 114, E6830–E6838. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, H.W., 3rd; Hendricks, A.G.; Ikeda, K.; Shuman, H.; Rodionov, V.; Ikebe, M.; Goldman, Y.E.; Holzbaur, E.L. Force-dependent detachment of kinesin-2 biases track switching at cytoskeletal filament intersections. Biophys. J. 2012, 103, 48–58. [Google Scholar] [CrossRef]
- Laib, J.A.; Marin, J.A.; Bloodgood, R.A.; Guilford, W.H. The reciprocal coordination and mechanics of molecular motors in living cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3190–3195. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, R.T.; Diehl, M.R.; Kolomeisky, A.B. Collective dynamics of processive cytoskeletal motors. Soft Matter 2016, 12, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Grover, R.; Fischer, J.; Schwarz, F.W.; Walter, W.J.; Schwille, P.; Diez, S. Transport efficiency of membrane-anchored kinesin-1 motors depends on motor density and diffusivity. Proc. Natl. Acad. Sci. USA 2016, 113, E7185–E7193. [Google Scholar] [CrossRef]
- Leduc, C.; Ruhnow, F.; Howard, J.; Diez, S. Detection of fractional steps in cargo movement by the collective operation of kinesin-1 motors. Proc. Natl. Acad. Sci. USA 2007, 104, 10847–10852. [Google Scholar] [CrossRef]
- De-Castro, A.R.G.; Rodrigues, D.R.M.; De-Castro, M.J.G.; Vieira, N.; Vieira, C.; Carvalho, A.X.; Gassmann, R.; Abreu, C.M.C.; Dantas, T.J. WDR60-mediated dynein-2 loading into cilia powers retrograde IFT and transition zone crossing. J. Cell Biol. 2022, 221, jcb202010178. [Google Scholar] [CrossRef]
- Toropova, K.; Mladenov, M.; Roberts, A.J. Intraflagellar transport dynein is autoinhibited by trapping of its mechanical and track-binding elements. Nat. Struct. Mol. Biol. 2017, 24, 461–468. [Google Scholar] [CrossRef]
- Goodman, B.S.; Reck-Peterson, S.L. Engineering defined motor ensembles with DNA origami. Methods Enzymol. 2014, 540, 169–188. [Google Scholar] [CrossRef]
- Zhang, Z.; Danne, N.; Meddens, B.; Heller, I.; Peterman, E.J.G. Direct imaging of intraflagellar-transport turnarounds reveals that motors detach, diffuse, and reattach to opposite-direction trains. Proc. Natl. Acad. Sci. USA 2021, 118, e2115089118. [Google Scholar] [CrossRef] [PubMed]
- Mijalkovic, J.; Prevo, B.; Oswald, F.; Mangeol, P.; Peterman, E.J.G. Ensemble and single-molecule dynamics of IFT dynein in Caenorhabditis elegans cilia. Nat. Commun. 2017, 8, 14591. [Google Scholar] [CrossRef] [PubMed]
- Conkar, D.; Firat-Karalar, E.N. Microtubule-associated proteins and emerging links to primary cilium structure, assembly, maintenance, and disassembly. FEBS J. 2021, 288, 786–798. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Engelke, M.F.; Blasius, T.L.; Verhey, K.J. Hedgehog-induced ciliary trafficking of kinesin-4 motor KIF7 requires intraflagellar transport but not KIF7’s microtubule binding. Mol. Biol. Cell 2022, 33, br1. [Google Scholar] [CrossRef]
- He, M.; Subramanian, R.; Bangs, F.; Omelchenko, T.; Liem Jr, K.F.; Kapoor, T.M.; Anderson, K.V. The kinesin-4 protein Kif7 regulates mammalian Hedgehog signalling by organizing the cilium tip compartment. Nat. Cell Biol. 2014, 16, 663–672. [Google Scholar] [CrossRef]
- Ma, R.; Hendel, N.L.; Marshall, W.F.; Qin, H. Speed and Diffusion of Kinesin-2 Are Competing Limiting Factors in Flagellar Length-Control Model. Biophys. J. 2020, 118, 2790–2800. [Google Scholar] [CrossRef]
- Firestone, A.J.; Weinger, J.S.; Maldonado, M.; Barlan, K.; Langston, L.D.; O’Donnell, M.; Gelfand, V.I.; Kapoor, T.M.; Chen, J.K. Small-molecule inhibitors of the AAA+ ATPase motor cytoplasmic dynein. Nature 2012, 484, 125–129. [Google Scholar] [CrossRef]
- Mijalkovic, J.; Girard, J.; van Krugten, J.; van Loo, J.; Zhang, Z.; Loseva, E.; Oswald, F.; Peterman, E.J.G. Cutting off ciliary protein import: Intraflagellar transport after dendritic femtosecond-laser ablation. Mol. Biol. Cell 2020, 31, 324–334. [Google Scholar] [CrossRef]
- Nachury, M.V.; Mick, D.U. Establishing and regulating the composition of cilia for signal transduction. Nat. Rev. Mol. Cell Biol. 2019, 20, 389–405. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Wen, X.; Chih, B.; Nelson, C.D.; Lane, W.S.; Scales, S.J.; Jackson, P.K. TULP3 bridges the IFT-A complex and membrane phosphoinositides to promote trafficking of G protein-coupled receptors into primary cilia. Genes Dev. 2010, 24, 2180–2193. [Google Scholar] [CrossRef]
- Abe, S.; Yagi, T.; Ishiyama, S.; Hiroe, M.; Marumo, F.; Ikawa, Y. Molecular cloning of a novel serine/threonine kinase, MRK, possibly involved in cardiac development. Oncogene 1995, 11, 2187–2195. [Google Scholar] [PubMed]
- Togawa, K.; Yan, Y.X.; Inomoto, T.; Slaugenhaupt, S.; Rustgi, A.K. Intestinal cell kinase (ICK) localizes to the crypt region and requires a dual phosphorylation site found in map kinases. J. Cell. Physiol. 2000, 183, 129–139. [Google Scholar] [CrossRef]
- Bladt, F.; Birchmeier, C. Characterization and expression analysis of the murine rck gene: A protein kinase with a potential function in sensory cells. Differentiation 1993, 53, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Matsushime, H.; Jinno, A.; Takagi, N.; Shibuya, M. A novel mammalian protein kinase gene (mak) is highly expressed in testicular germ cells at and after meiosis. Mol. Cell. Biol. 1990, 10, 2261–2268. [Google Scholar] [PubMed]
- Burghoorn, J.; Dekkers, M.P.; Rademakers, S.; de Jong, T.; Willemsen, R.; Jansen, G. Mutation of the MAP kinase DYF-5 affects docking and undocking of kinesin-2 motors and reduces their speed in the cilia of Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2007, 104, 7157–7162. [Google Scholar] [CrossRef]
- Miyata, Y.; Akashi, M.; Nishida, E. Molecular cloning and characterization of a novel member of the MAP kinase superfamily. Genes Cells 1999, 4, 299–309. [Google Scholar] [CrossRef]
- Berman, S.A.; Wilson, N.F.; Haas, N.A.; Lefebvre, P.A. A novel MAP kinase regulates flagellar length in Chlamydomonas. Curr. Biol. 2003, 13, 1145–1149. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, C.; Galaktionov, K. p42, a novel cyclin-dependent kinase-activating kinase in mammalian cells. J. Biol. Chem. 2004, 279, 4507–4514. [Google Scholar] [CrossRef]
- Phirke, P.; Efimenko, E.; Mohan, S.; Burghoorn, J.; Crona, F.; Bakhoum, M.W.; Trieb, M.; Schuske, K.; Jorgensen, E.M.; Piasecki, B.P. Transcriptional profiling of C. elegans DAF-19 uncovers a ciliary base-associated protein and a CDK/CCRK/LF2p-related kinase required for intraflagellar transport. Dev. Biol. 2011, 357, 235–247. [Google Scholar] [CrossRef]
- Tam, L.-W.; Wilson, N.F.; Lefebvre, P.A. A CDK-related kinase regulates the length and assembly of flagella in Chlamydomonas. J. Cell Biol. 2007, 176, 819–829. [Google Scholar] [CrossRef]
- Schulman, H.; Greengard, P. Stimulation of brain membrane protein phosphorylation by calcium and an endogenous heat-stable protein. Nature 1978, 271, 478–479. [Google Scholar] [CrossRef] [PubMed]
- Hudmon, A.; Schulman, H. Neuronal Ca2+/calmodulin-dependent protein kinase II: The role of structure and autoregulation in cellular function. Annu. Rev. Biochem. 2002, 71, 473. [Google Scholar] [CrossRef] [PubMed]
- Robatzek, M.; Thomas, J.H. Calcium/calmodulin-dependent protein kinase II regulates Caenorhabditis elegans locomotion in concert with a Go/Gq signaling network. Genetics 2000, 156, 1069–1082. [Google Scholar] [CrossRef]
- Reiner, D.J.; Newton, E.M.; Tian, H.; Thomas, J.H. Diverse behavioural defects caused by mutations in Caenorhabditis elegans unc-43 CaM kinase II. Nature 1999, 402, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Pazour, G.J.; Agrin, N.; Leszyk, J.; Witman, G.B. Proteomic analysis of a eukaryotic cilium. J. Cell Biol. 2005, 170, 103–113. [Google Scholar] [CrossRef]
- Montini, E.; Andolfi, G.; Caruso, A.; Buchner, G.; Walpole, S.M.; Mariani, M.; Consalez, G.; Trump, D.; Ballabio, A.; Franco, B. Identification and characterization of a novel serine–threonine kinase gene from the Xp22 region. Genomics 1998, 51, 427–433. [Google Scholar] [CrossRef]
- Canning, P.; Park, K.; Gonçalves, J.; Li, C.; Howard, C.J.; Sharpe, T.D.; Holt, L.J.; Pelletier, L.; Bullock, A.N.; Leroux, M.R. CDKL family kinases have evolved distinct structural features and ciliary function. Cell Rep. 2018, 22, 885–894. [Google Scholar] [CrossRef]
- Park, K.; Li, C.; Tsiropoulou, S.; Gonçalves, J.; Kondratev, C.; Pelletier, L.; Blacque, O.E.; Leroux, M.R. CDKL kinase regulates the length of the ciliary proximal segment. Curr. Biol. 2021, 31, 2359–2373. e2357. [Google Scholar]
- Tam, L.-W.; Ranum, P.T.; Lefebvre, P.A. CDKL5 regulates flagellar length and localizes to the base of the flagella in Chlamydomonas. Mol. Biol. Cell 2013, 24, 588–600. [Google Scholar] [CrossRef]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef]
- Fu, Z.; Gailey, C.D.; Wang, E.J.; Brautigan, D.L. Ciliogenesis associated kinase 1: Targets and functions in various organ systems. FEBS Lett. 2019, 593, 2990–3002. [Google Scholar] [CrossRef]
- Jiang, X.; Shao, W.; Chai, Y.; Huang, J.; Mohamed, M.A.; Ökten, Z.; Li, W.; Zhu, Z.; Ou, G. DYF-5/MAK–dependent phosphorylation promotes ciliary tubulin unloading. Proc. Natl. Acad. Sci. USA 2022, 119, e2207134119. [Google Scholar] [CrossRef] [PubMed]
- Asleson, C.M.; Lefebvre, P.A. Genetic analysis of flagellar length control in Chlamydomonas reinhardtii: A new long-flagella locus and extragenic suppressor mutations. Genetics 1998, 148, 693–702. [Google Scholar] [PubMed]
- Yi, P.; Xie, C.; Ou, G. The kinases male germ cell-associated kinase and cell cycle-related kinase regulate kinesin-2 motility in Caenorhabditis elegans neuronal cilia. Traffic 2018, 19, 522–535. [Google Scholar]
- Okamoto, S.; Chaya, T.; Omori, Y.; Kuwahara, R.; Kubo, S.; Sakaguchi, H.; Furukawa, T. Ick ciliary kinase is essential for planar cell polarity formation in inner ear hair cells and hearing function. J. Neurosci. 2017, 37, 2073–2085. [Google Scholar] [PubMed]
- Wang, Y.; Ren, Y.; Pan, J. Regulation of flagellar assembly and length in Chlamydomonas by LF4, a MAPK-related kinase. FASEB J. 2019, 33, 6431–6441. [Google Scholar]
- Gailey, C.D.; Wang, E.J.; Jin, L.; Ahmadi, S.; Brautigan, D.L.; Li, X.; Xu, W.; Scott, M.M.; Fu, Z. Phosphosite T674A mutation in kinesin family member 3A fails to reproduce tissue and ciliary defects characteristic of CILK1 loss of function. Dev. Dyn. 2021, 250, 263–273. [Google Scholar] [CrossRef]
- Hirano, T.; Katoh, Y.; Nakayama, K. Intraflagellar transport-A complex mediates ciliary entry and retrograde trafficking of ciliary G protein–coupled receptors. Mol. Biol. Cell 2017, 28, 429–439. [Google Scholar]
- Hamada, Y.; Tsurumi, Y.; Nozaki, S.; Katoh, Y.; Nakayama, K. Interaction of WDR60 intermediate chain with TCTEX1D2 light chain of the dynein-2 complex is crucial for ciliary protein trafficking. Mol. Biol. Cell 2018, 29, 1628–1639. [Google Scholar] [CrossRef]
- Takahara, M.; Katoh, Y.; Nakamura, K.; Hirano, T.; Sugawa, M.; Tsurumi, Y.; Nakayama, K. Ciliopathy-associated mutations of IFT122 impair ciliary protein trafficking but not ciliogenesis. Hum. Mol. Genet. 2018, 27, 516–528. [Google Scholar] [CrossRef]
- Wang, W.; Allard, B.A.; Pottorf, T.S.; Wang, H.H.; Vivian, J.L.; Tran, P.V. Genetic interaction of mammalian IFT-A paralogs regulates cilia disassembly, ciliary entry of membrane protein, Hedgehog signaling, and embryogenesis. FASEB J. 2020, 34, 6369–6381. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Wan, H.; Tan, G. Cell cycle-related kinase in carcinogenesis. Oncol. Lett. 2012, 4, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Zhou, J.; Xiong, Z.; Sun, H.; Yang, W.; Mok, M.T.; Wang, J.; Li, J.; Liu, M.; Tang, W. Cell cycle-related kinase reprograms the liver immune microenvironment to promote cancer metastasis. Cell. Mol. Immunol. 2021, 18, 1005–1015. [Google Scholar] [CrossRef]
- Snouffer, A.; Brown, D.; Lee, H.; Walsh, J.; Lupu, F.; Norman, R.; Lechtreck, K.; Ko, H.W.; Eggenschwiler, J. Cell Cycle-Related Kinase (CCRK) regulates ciliogenesis and Hedgehog signaling in mice. PLoS Genet. 2017, 13, e1006912. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Nakamura, K.; Satoda, Y.; Katoh, Y.; Nakayama, K. CCRK/CDK20 regulates ciliary retrograde protein trafficking via interacting with BROMI/TBC1D32. PLoS ONE 2021, 16, e0258497. [Google Scholar] [CrossRef]
- Ko, H.W.; Norman, R.X.; Tran, J.; Fuller, K.P.; Fukuda, M.; Eggenschwiler, J.T. Broad-minded links cell cycle-related kinase to cilia assembly and hedgehog signal transduction. Dev. Cell 2010, 18, 237–247. [Google Scholar] [CrossRef]
- Fu, Z.; Larson, K.A.; Chitta, R.K.; Parker, S.A.; Turk, B.E.; Lawrence, M.W.; Kaldis, P.; Galaktionov, K.; Cohn, S.M.; Shabanowitz, J. Identification of yin-yang regulators and a phosphorylation consensus for male germ cell-associated kinase (MAK)-related kinase. Mol. Cell. Biol. 2006, 26, 8639–8654. [Google Scholar] [CrossRef]
- Chen, M.X.; McPartlin, A.E.; Brown, L.; Chen, Y.H.; Barker, H.M.; Cohen, P. A novel human protein serine/threonine phosphatase, which possesses four tetratricopeptide repeat motifs and localizes to the nucleus. EMBO J. 1994, 13, 4278–4290. [Google Scholar] [CrossRef]
- Fu, Z.; Schroeder, M.J.; Shabanowitz, J.; Kaldis, P.; Togawa, K.; Rustgi, A.K.; Hunt, D.F.; Sturgill, T.W. Activation of a nuclear Cdc2-related kinase within a mitogen-activated protein kinase-like TDY motif by autophosphorylation and cyclin-dependent protein kinase-activating kinase. Mol. Cell. Biol. 2005, 25, 6047–6064. [Google Scholar] [CrossRef]
- Ichinose, S.; Ogawa, T.; Hirokawa, N. Mechanism of activity-dependent cargo loading via the phosphorylation of KIF3A by PKA and CaMKIIa. Neuron 2015, 87, 1022–1035. [Google Scholar] [CrossRef]
- Lisman, J.; Schulman, H.; Cline, H. The molecular basis of CaMKII function in synaptic and behavioural memory. Nat. Rev. Neurosci. 2002, 3, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Guillaud, L.; Wong, R.; Hirokawa, N. Disruption of KIF17–Mint1 interaction by CaMKII-dependent phosphorylation: A molecular model of kinesin–cargo release. Nat. Cell Biol. 2008, 10, 19–29. [Google Scholar] [CrossRef] [PubMed]
- McVicker, D.P.; Millette, M.M.; Dent, E.W. Signaling to the microtubule cytoskeleton: An unconventional role for C a MKII. Dev. Neurobiol. 2015, 75, 423–434. [Google Scholar] [CrossRef]
- Guillaud, L.; Setou, M.; Hirokawa, N. KIF17 dynamics and regulation of NR2B trafficking in hippocampal neurons. J. Neurosci. 2003, 23, 131–140. [Google Scholar] [CrossRef]
- Wong, R.W.-C.; Setou, M.; Teng, J.; Takei, Y.; Hirokawa, N. Overexpression of motor protein KIF17 enhances spatial and working memory in transgenic mice. Proc. Natl. Acad. Sci. USA 2002, 99, 14500–14505. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhu, X.; Wu, Q.; Pan, J. Ciliary length sensing regulates IFT entry via changes in FLA8/KIF3B phosphorylation to control ciliary assembly. Curr. Biol. 2018, 28, 2429–2435. e2423. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jensen, V.L.; Park, K.; Kennedy, J.; Garcia-Gonzalo, F.R.; Romani, M.; De Mori, R.; Bruel, A.-L.; Gaillard, D.; Doray, B. MKS5 and CEP290 dependent assembly pathway of the ciliary transition zone. PLoS Biol. 2016, 14, e1002416. [Google Scholar] [CrossRef] [PubMed]
- Omori, Y.; Chaya, T.; Katoh, K.; Kajimura, N.; Sato, S.; Muraoka, K.; Ueno, S.; Koyasu, T.; Kondo, M.; Furukawa, T. Negative regulation of ciliary length by ciliary male germ cell-associated kinase (Mak) is required for retinal photoreceptor survival. Proc. Natl. Acad. Sci. USA 2010, 107, 22671–22676. [Google Scholar] [CrossRef]
- Maurya, A.K.; Rogers, T.; Sengupta, P. A CCRK and a MAK kinase modulate cilia branching and length via regulation of axonemal microtubule dynamics in Caenorhabditis elegans. Curr. Biol. 2019, 29, 1286–1300. e1284. [Google Scholar] [CrossRef]
- Roux, K.J.; Kim, D.I.; Raida, M.; Burke, B. A promiscuous biotin ligase fusion protein identifies proximal and interacting proteins in mammalian cells. J. Cell Biol. 2012, 196, 801–810. [Google Scholar] [CrossRef]
- Varjosalo, M.; Sacco, R.; Stukalov, A.; Van Drogen, A.; Planyavsky, M.; Hauri, S.; Aebersold, R.; Bennett, K.L.; Colinge, J.; Gstaiger, M. Interlaboratory reproducibility of large-scale human protein-complex analysis by standardized AP-MS. Nat. Methods 2013, 10, 307–314. [Google Scholar] [CrossRef]
- Bauer, A.; Kuster, B. Affinity purification-mass spectrometry: Powerful tools for the characterization of protein complexes. Eur. J. Biochem. 2003, 270, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Lacey, S.E.; Foster, H.E.; Pigino, G. The Molecular Structure of Anterograde Intraflagellar Transport Trains. bioRxiv 2022. [Google Scholar] [CrossRef]
- Bansal, A.; Shikha, S.; Zhang, Y. Towards translational optogenetics. Nat. Biomed. Eng. 2022, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.; He, L.; Huang, Y.; Zhou, Y. Optophysiology: Illuminating cell physiology with optogenetics. Physiol. Rev. 2022, 102, 1263–1325. [Google Scholar] [CrossRef]
- Yesbolatova, A.; Saito, Y.; Kitamoto, N.; Makino-Itou, H.; Ajima, R.; Nakano, R.; Nakaoka, H.; Fukui, K.; Gamo, K.; Tominari, Y. The auxin-inducible degron 2 technology provides sharp degradation control in yeast, mammalian cells, and mice. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Martinez, M.A.; Matus, D.Q. Auxin-mediated protein degradation in Caenorhabditis elegans. Bio-Protocol 2020, 10, e3589. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mammals | C. elegans | Chlamydomonas | |
|---|---|---|---|
| RCK | CILK1 and MRK [152,153] | - | - |
| MAK [154,155] | Dyf-5 [156] | - | |
| MOK [157] | - | LF4 [158] | |
| CDK | CCRK/CDK20/p42 [159] | Dyf-18 [160] | LF2 [161] |
| CaMK | CaMKII [162,163] | UNC-43 [164,165] | CrCDPK1 [166] |
| CDKL | CDKL5 [167,168] | CDKL-1 [169] | LF5 [170] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mul, W.; Mitra, A.; Peterman, E.J.G. Mechanisms of Regulation in Intraflagellar Transport. Cells 2022, 11, 2737. https://doi.org/10.3390/cells11172737
Mul W, Mitra A, Peterman EJG. Mechanisms of Regulation in Intraflagellar Transport. Cells. 2022; 11(17):2737. https://doi.org/10.3390/cells11172737
Chicago/Turabian StyleMul, Wouter, Aniruddha Mitra, and Erwin J. G. Peterman. 2022. "Mechanisms of Regulation in Intraflagellar Transport" Cells 11, no. 17: 2737. https://doi.org/10.3390/cells11172737
APA StyleMul, W., Mitra, A., & Peterman, E. J. G. (2022). Mechanisms of Regulation in Intraflagellar Transport. Cells, 11(17), 2737. https://doi.org/10.3390/cells11172737