
LPAL2 Suppresses Tumor Growth and Metastasis of Hepatocellular Carcinoma by Modulating MMP9 Expression

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagents
2.2. HCC Specimens
2.3. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
2.4. Immunoblot Analysis
2.5. Establishment of Cell Lines Stably Overexpressing LPAL2-Targeting shRNAs
2.6. In Vitro Migration and Invasion Assays
2.7. Cell Viability and Growth Assays
2.8. Cell Sorting and Flow Cytometry
2.9. Sphere-Formation Assay
2.10. Animal Models
2.11. Integration of the Protein-Protein Interaction (PPI) Network
2.12. Statistical Analysis
3. Results
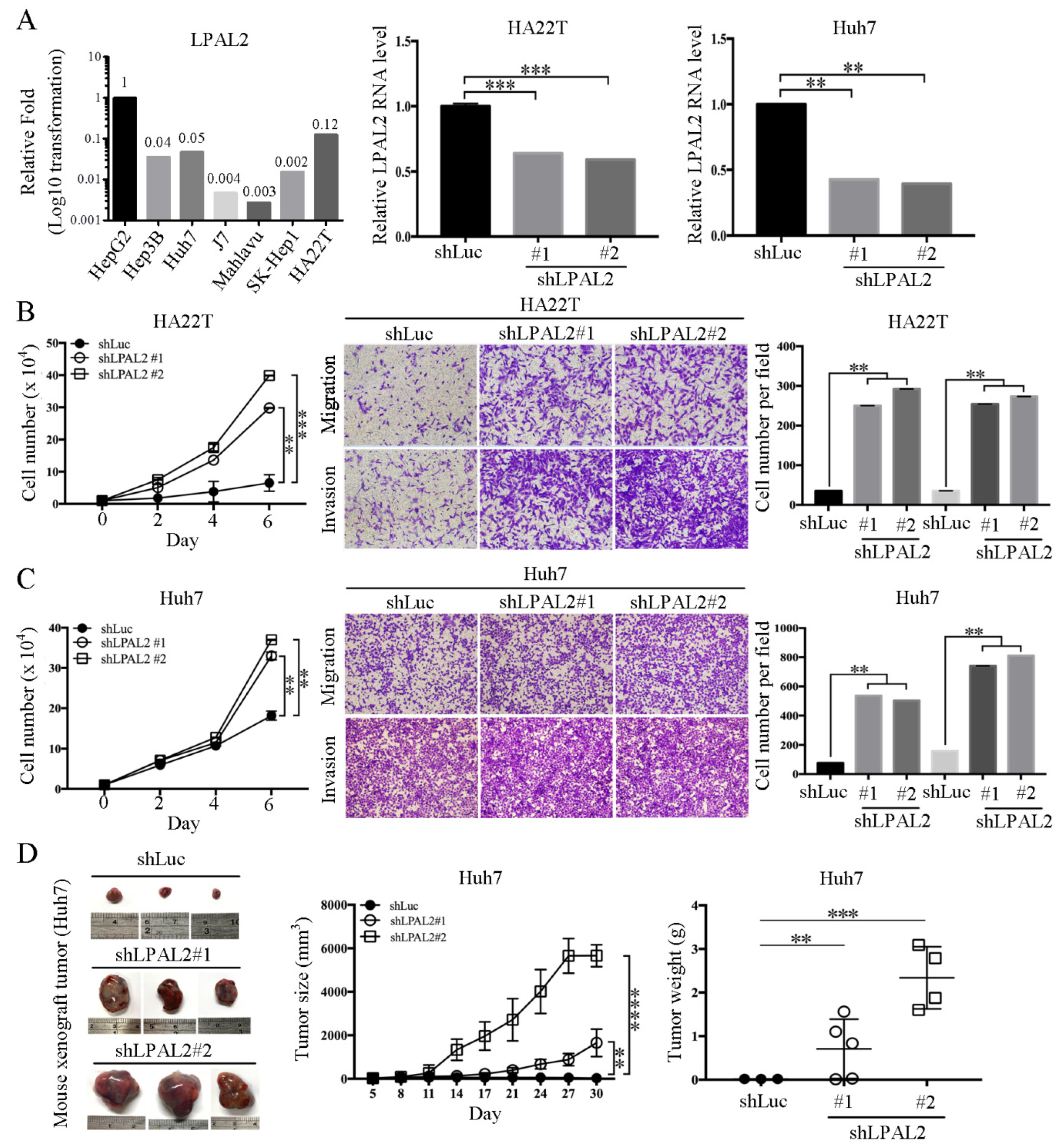
3.1. Elevated Expression of LPAL2 in HCC Is Positively Correlated with Clinical Outcome
3.2. Knockdown of LPAL2 Enhances Tumor Growth, Migration, and Invasion
3.3. MMP9 Is Regulated by LPAL2
3.4. Clinical Correlation of LPAL2 with MMP9
3.5. LPAL2 Alleviates Doxorubicin Resistance and Cancer Stem Cell Phenotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farzaneh, Z.; Vosough, M.; Agarwal, T.; Farzaneh, M. Critical signaling pathways governing hepatocellular carcinoma behavior; small molecule-based approaches. Cancer Cell Int. 2021, 21, 208. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, E.; Lindoso, R.S.; Sensi, F.; Pucciarelli, S.; Bussolati, B.; Agostini, M.; Collino, F. Intrinsic and Extrinsic Modulators of the Epithelial to Mesenchymal Transition: Driving the Fate of Tumor Microenvironment. Front. Oncol. 2020, 10, 1122. [Google Scholar] [CrossRef]
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.J.; Calin, G.A. Junk DNA and the long non-coding RNA twist in cancer genetics. Oncogene 2015, 34, 5003–5011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Hu, X.; Yang, L.; Mo, Y.Y. Role of Pseudogenes in Tumorigenesis. Cancers 2018, 10, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, Y.; Zhang, L.; Huang, Y.; He, T.; Zhang, L.; Zhao, X.; Zhou, X.; Zhou, D.; Yan, Y.; Zhou, J.; et al. Pseudogene PDIA3P1 promotes cell proliferation, migration and invasion, and suppresses apoptosis in hepatocellular carcinoma by regulating the p53 pathway. Cancer Lett. 2017, 407, 76–83. [Google Scholar] [CrossRef]
- Cao, S.W.; Huang, J.L.; Chen, J.; Hu, Y.W.; Hu, X.M.; Ren, T.Y.; Zheng, S.H.; Lin, J.D.; Tang, J.; Zheng, L.; et al. Long non-coding RNA UBE2CP3 promotes tumor metastasis by inducing epithelial-mesenchymal transition in hepatocellular carcinoma. Oncotarget 2017, 8, 65370–65385. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, J.; Zeng, Y.; Wei, J.; Jing, J.; Li, G.; Su, L.; Tang, X.; Wu, T.; Zhou, L. Functional Variant in the SLC22A3-LPAL2-LPA Gene Cluster Contributes to the Severity of Coronary Artery Disease. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1989–1996. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Hou, S.Y.; Qi, X.; Deng, M.; Cao, J.M.; Tong, B.D.; Xiong, W. LncRNA LPAL2/miR-1287-5p/EGFR Axis Modulates TED-Derived Orbital Fibroblast Activation Through Cell Adhesion Factors. J. Clin. Endocrinol. Metab. 2021, 106, e2866–e2886. [Google Scholar] [CrossRef]
- Lin, Y.H.; Wu, M.H.; Liu, Y.C.; Lyu, P.C.; Yeh, C.T.; Lin, K.H. LINC01348 suppresses hepatocellular carcinoma metastasis through inhibition of SF3B3-mediated EZH2 pre-mRNA splicing. Oncogene 2021, 40, 4675–4685. [Google Scholar] [CrossRef]
- Jiang, H.; Li, H. Prognostic values of tumoral MMP2 and MMP9 overexpression in breast cancer: A systematic review and meta-analysis. BMC Cancer 2021, 21, 149. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Chen, X. Drug resistance and combating drug resistance in cancer. Cancer Drug Resist. 2019, 2, 141–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.; Kim, Y.K. Cancer Stem Cells as a Potential Target to Overcome Multidrug Resistance. Front. Oncol. 2020, 10, 764. [Google Scholar] [CrossRef] [PubMed]
- Lou, W.; Ding, B.; Fu, P. Pseudogene-Derived lncRNAs and Their miRNA Sponging Mechanism in Human Cancer. Front. Cell Dev. Biol. 2020, 8, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.; Jiang, Z.; Zeng, S.; Wu, H.; Zhang, Z.; Guo, X.; Dong, K.; Wang, J.; Shang, L.; Li, L. A new candidate oncogenic lncRNA derived from pseudogene WFDC21P promotes tumor progression in gastric cancer. Cell Death Dis. 2021, 12, 903. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Ma, T.; Chen, M.; Zou, Z.; Zhang, Z. The pseudogene-derived long non-coding RNA SFTA1P suppresses cell proliferation, migration, and invasion in gastric cancer. Biosci. Rep. 2018, 38, BSR20171193. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.L.; Cao, S.W.; Ou, Q.S.; Yang, B.; Zheng, S.H.; Tang, J.; Chen, J.; Hu, Y.W.; Zheng, L.; Wang, Q. The long non-coding RNA PTTG3P promotes cell growth and metastasis via up-regulating PTTG1 and activating PI3K/AKT signaling in hepatocellular carcinoma. Mol. Cancer 2018, 17, 93. [Google Scholar] [CrossRef]
- Pan, W.; Li, W.; Zhao, J.; Huang, Z.; Zhao, J.; Chen, S.; Wang, C.; Xue, Y.; Huang, F.; Fang, Q.; et al. lncRNA-PDPK2P promotes hepatocellular carcinoma progression through the PDK1/AKT/Caspase 3 pathway. Mol. Oncol. 2019, 13, 2246–2258. [Google Scholar] [CrossRef] [Green Version]
- Huang, H. Matrix Metalloproteinase-9 (MMP-9) as a Cancer Biomarker and MMP-9 Biosensors: Recent Advances. Sensors 2018, 18, 3249. [Google Scholar] [CrossRef] [Green Version]
- Phi, L.T.H.; Sari, I.N.; Yang, Y.G.; Lee, S.H.; Jun, N.; Kim, K.S.; Lee, Y.K.; Kwon, H.Y. Cancer Stem Cells (CSCs) in Drug Resistance and their Therapeutic Implications in Cancer Treatment. Stem Cells Int. 2018, 2018, 5416923. [Google Scholar] [CrossRef] [Green Version]
- Spallarossa, P.; Altieri, P.; Garibaldi, S.; Ghigliotti, G.; Barisione, C.; Manca, V.; Fabbi, P.; Ballestrero, A.; Brunelli, C.; Barsotti, A. Matrix metalloproteinase-2 and -9 are induced differently by doxorubicin in H9c2 cells: The role of MAP kinases and NAD(P)H oxidase. Cardiovasc. Res. 2006, 69, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, R.; Ng, K.P.; Cicek, M.; Kelleher, C.; Niculaita, R.; Casey, G.; Sizemore, N. Role of IKK and oscillatory NFkappaB kinetics in MMP-9 gene expression and chemoresistance to 5-fluorouracil in RKO colorectal cancer cells. Mol. Carcinog. 2007, 46, 402–413. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | n = 157 | LPAL2 Mean a ± SE | pb |
|---|---|---|---|
| Age (years) | |||
| <65 | 99 | 0.7794 ± 0.1007 | 0.8944 |
| ≥65 | 58 | 0.6136 ± 0.0790 | |
| Gender | |||
| Male | 79 | 0.5916 ± 0.0754 | 0.0134 |
| Female | 78 | 0.8462 ± 0.1171 | |
| Cirrhosis | |||
| No | 94 | 0.7073 ± 0.1004 | 0.2685 |
| Yes | 63 | 0.7344 ± 0.0903 | |
| AFP | |||
| Low | 98 | 0.6554 ± 0.0701 | 0.5379 |
| High | 55 | 0.8294 ± 0.1552 | |
| Tumor type | |||
| Solitary | 125 | 0.7400 ± 0.0838 | 0.9809 |
| Multiple | 32 | 0.6329 ± 0.1036 | |
| Tumor size | |||
| <5 cm | 91 | 0.7961 ± 0.1032 | 0.2172 |
| ≥5 cm | 66 | 0.6107 ± 0.0856 | |
| Vascular invasion | |||
| No | 91 | 0.7787 ± 0.1017 | 0.1937 |
| Yes | 66 | 0.6346 ± 0.0896 | |
| Pathological stage | |||
| I | 75 | 0.7298 ± 0.0859 | 0.5887 |
| II | 51 | 0.8018 ± 0.1627 | |
| III | 31 | 0.5521 ± 0.1050 | |
| Grading | |||
| 1 | 4 | 0.7812 ± 0.1094 | 0.2256 |
| 2 | 110 | 0.6516 ± 0.0686 | |
| 3 | 43 | 0.8824 ± 0.1848 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-H.; Liu, Y.-C.; Chen, C.-Y.; Chi, H.-C.; Wu, M.-H.; Huang, P.-S.; Chang, C.-C.; Lin, T.-K.; Yeh, C.-T.; Lin, K.-H. LPAL2 Suppresses Tumor Growth and Metastasis of Hepatocellular Carcinoma by Modulating MMP9 Expression. Cells 2022, 11, 2610. https://doi.org/10.3390/cells11162610
Lin Y-H, Liu Y-C, Chen C-Y, Chi H-C, Wu M-H, Huang P-S, Chang C-C, Lin T-K, Yeh C-T, Lin K-H. LPAL2 Suppresses Tumor Growth and Metastasis of Hepatocellular Carcinoma by Modulating MMP9 Expression. Cells. 2022; 11(16):2610. https://doi.org/10.3390/cells11162610
Chicago/Turabian StyleLin, Yang-Hsiang, Yu-Chin Liu, Cheng-Yi Chen, Hsiang-Cheng Chi, Meng-Han Wu, Po-Shuan Huang, Cheng-Chih Chang, Tzu-Kang Lin, Chau-Ting Yeh, and Kwang-Huei Lin. 2022. "LPAL2 Suppresses Tumor Growth and Metastasis of Hepatocellular Carcinoma by Modulating MMP9 Expression" Cells 11, no. 16: 2610. https://doi.org/10.3390/cells11162610
APA StyleLin, Y.-H., Liu, Y.-C., Chen, C.-Y., Chi, H.-C., Wu, M.-H., Huang, P.-S., Chang, C.-C., Lin, T.-K., Yeh, C.-T., & Lin, K.-H. (2022). LPAL2 Suppresses Tumor Growth and Metastasis of Hepatocellular Carcinoma by Modulating MMP9 Expression. Cells, 11(16), 2610. https://doi.org/10.3390/cells11162610