
The Mechanical Microenvironment Regulates Axon Diameters Visualized by Cryo-Electron Tomography



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Thy1-YFP Transgenic Mice and Brain Tissue Fixation and Sectioning
2.2. Confocal Imaging, 3D Reconstitution and Quantification of Axon Diameter
2.3. Hippocampal Neuron Culture on Cover Glasses, Immunostaining and Imaging
2.4. Hippocampal and Cortical Neuron Culture on EM Grids and Vitrification
2.5. Cryo-Electron Tomography (Cryo-ET)
2.6. 3D Reconstruction and Segmentation
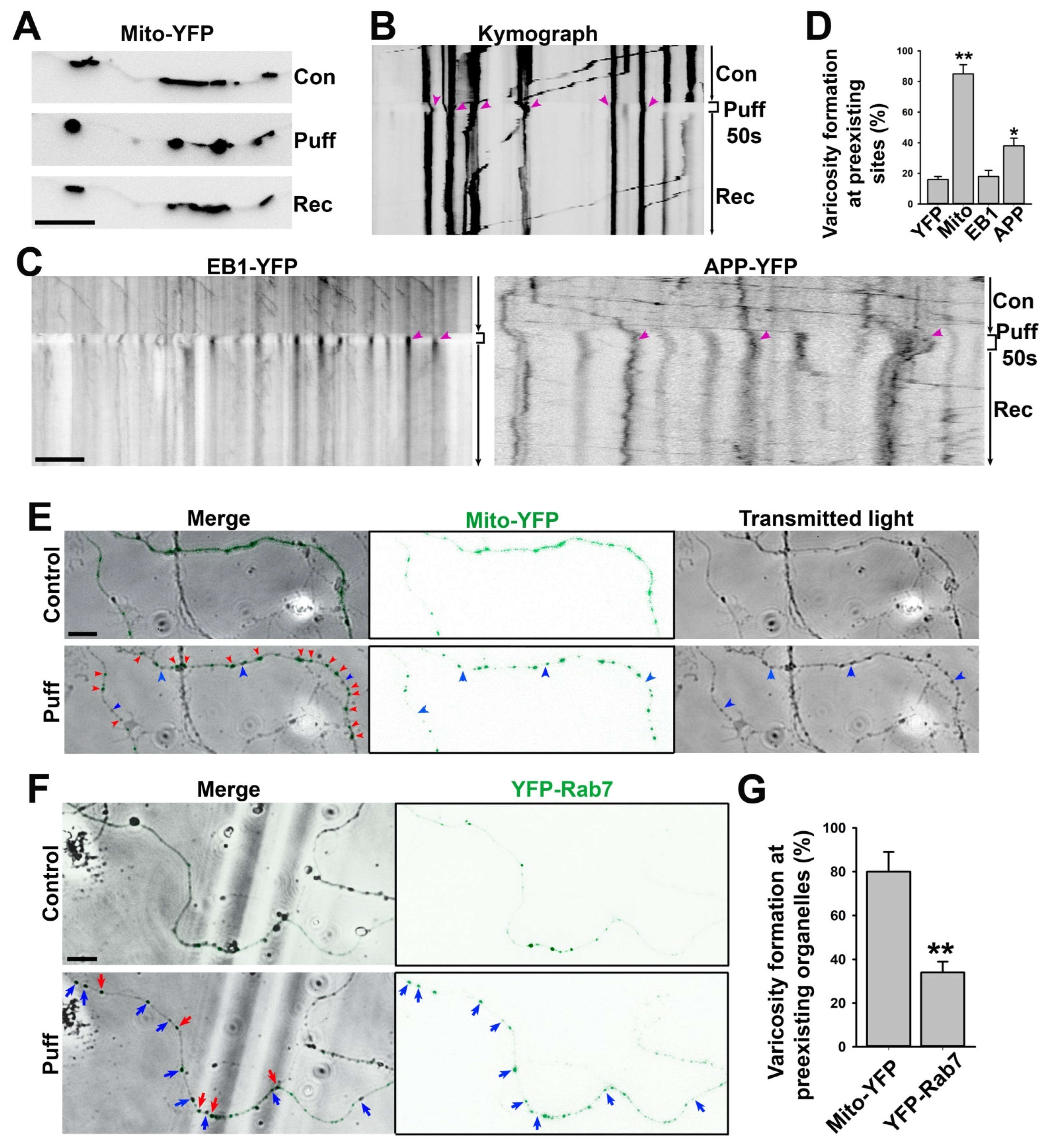
2.7. Puffing-Induced Axonal Varicosity Formation and Live-Cell Imaging
3. Results
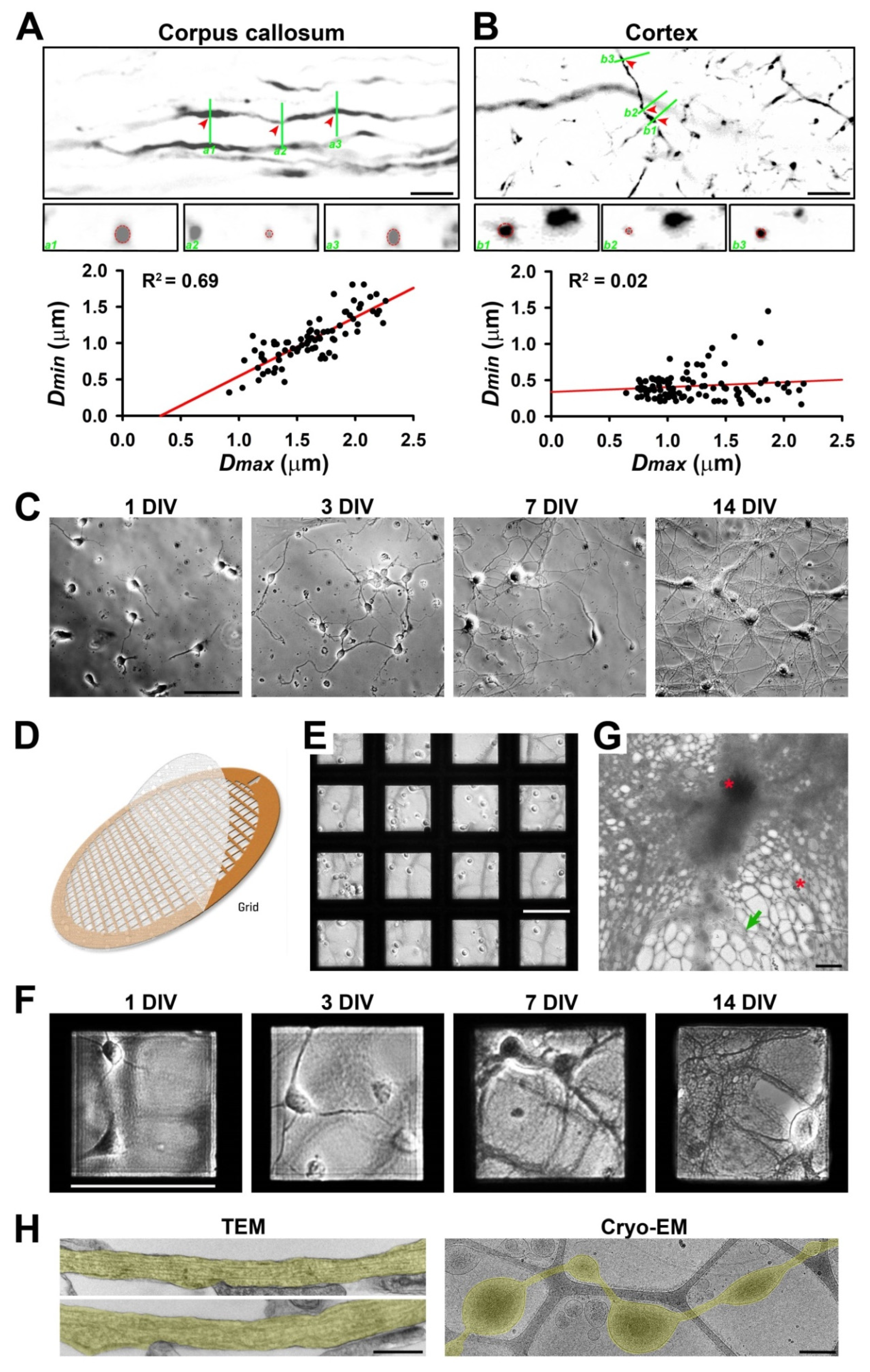
3.1. Diameter Variation of Individual Axons Correlates with the Complexity of Microenvironment
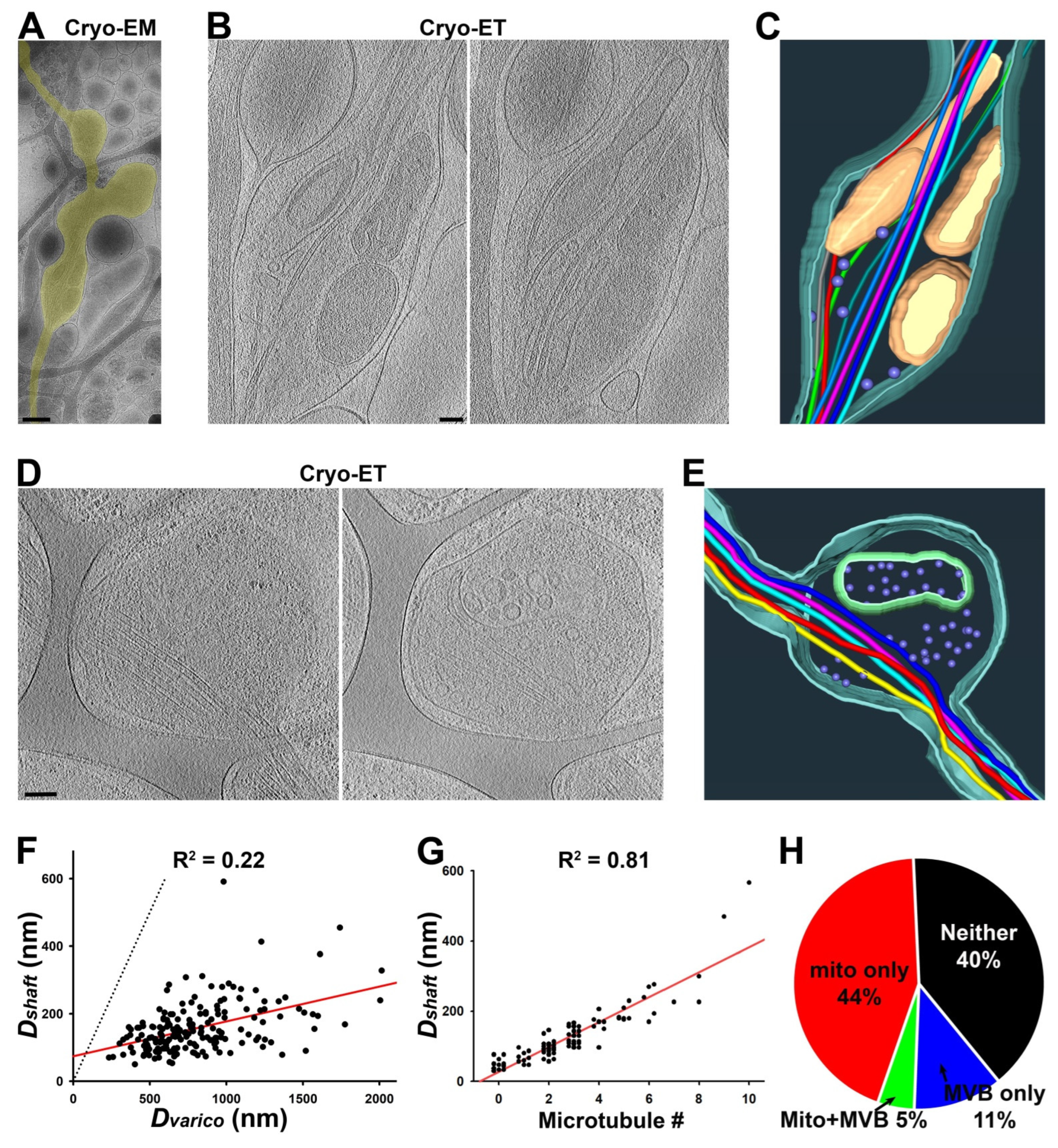
3.2. Ultrastructures of Axonal Varicosities Induced by Non-Uniform Extracellular Support
3.3. A Majority of Puffing-Induced Axonal Varicosities Formed around Mitochondrial Clusters
3.4. Axon Branch Points Often Possess Enlarged Diameters and Are Highly Heterogeneous
3.5. Bundled Axons Tend to Have Relatively More Uniform Diameters with Fewer Axonal Varicosities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Debanne, D. Information processing in the axon. Nat. Rev. Neurosci. 2004, 5, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Gu, C. Rapid and Reversible Development of Axonal Varicosities: A New Form of Neural Plasticity. Front. Mol. Neurosci. 2021, 14, 610857. [Google Scholar] [CrossRef] [PubMed]
- Browne, K.D.; Chen, X.H.; Meaney, D.F.; Smith, D.H. Mild traumatic brain injury and diffuse axonal injury in swine. J. Neurotrauma 2011, 28, 1747–1755. [Google Scholar] [CrossRef]
- Reeves, T.M.; Smith, T.L.; Williamson, J.C.; Phillips, L.L. Unmyelinated axons show selective rostrocaudal pathology in the corpus callosum after traumatic brain injury. J. Neuropathol. Exp. Neurol. 2012, 71, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Johnson, V.E.; Stewart, W.; Smith, D.H. Axonal pathology in traumatic brain injury. Exp. Neurol. 2013, 246, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Weimer, R.M.; Kallop, D.; Olsen, O.; Wu, Z.; Renier, N.; Uryu, K.; Tessier-Lavigne, M. Regulation of axon degeneration after injury and in development by the endogenous calpain inhibitor calpastatin. Neuron 2013, 80, 1175–1189. [Google Scholar] [CrossRef]
- Gu, Y.; Jukkola, P.; Wang, Q.; Esparza, T.; Zhao, Y.; Brody, D.; Gu, C. Polarity of varicosity initiation in central neuron mechanosensation. J. Cell Biol. 2017, 216, 2179–2199. [Google Scholar] [CrossRef]
- Sun, C.; Lin, Q.; Cheng, Y.; Zhao, Y.; Gu, C. Immediate induction of varicosities by transverse compression but not uniaxial stretch in axon mechanosensation. Acta Neuropathol. Commun. 2022, 10, 7. [Google Scholar] [CrossRef]
- Greenberg, M.M.; Leitao, C.; Trogadis, J.; Stevens, J.K. Irregular geometries in normal unmyelinated axons: A 3D serial EM analysis. J. Neurocytol. 1990, 19, 978–988. [Google Scholar] [CrossRef]
- Shepherd, G.M.; Harris, K.M. Three-dimensional structure and composition of CA3→CA1 axons in rat hippocampal slices: Implications for presynaptic connectivity and compartmentalization. J. Neurosci. 1998, 18, 8300–8310. [Google Scholar] [CrossRef]
- Giacci, M.K.; Bartlett, C.A.; Huynh, M.; Kilburn, M.R.; Dunlop, S.A.; Fitzgerald, M. Three dimensional electron microscopy reveals changing axonal and myelin morphology along normal and partially injured optic nerves. Sci. Rep. 2018, 8, 3979. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Zhong, G.; Zhuang, X. Actin, spectrin, and associated proteins form a periodic cytoskeletal structure in axons. Science 2013, 339, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Leterrier, C. Putting the axonal periodic scaffold in order. Curr. Opin. Neurobiol. 2021, 69, 33–40. [Google Scholar] [CrossRef] [PubMed]
- D’Este, E.; Kamin, D.; Velte, C.; Gottfert, F.; Simons, M.; Hell, S.W. Subcortical cytoskeleton periodicity throughout the nervous system. Sci. Rep. 2016, 6, 22741. [Google Scholar] [CrossRef] [PubMed]
- D’Este, E.; Kamin, D.; Balzarotti, F.; Hell, S.W. Ultrastructural anatomy of nodes of Ranvier in the peripheral nervous system as revealed by STED microscopy. Proc. Natl. Acad. Sci. USA 2017, 114, E191–E199. [Google Scholar] [CrossRef]
- He, J.; Zhou, R.; Wu, Z.; Carrasco, M.A.; Kurshan, P.T.; Farley, J.E.; Simon, D.J.; Wang, G.; Han, B.; Hao, J.; et al. Prevalent presence of periodic actin-spectrin-based membrane skeleton in a broad range of neuronal cell types and animal species. Proc. Natl. Acad. Sci. USA 2016, 113, 6029–6034. [Google Scholar] [CrossRef]
- Costa, A.R.; Sousa, S.C.; Pinto-Costa, R.; Mateus, J.C.; Lopes, C.D.; Costa, A.C.; Rosa, D.; Machado, D.; Pajuelo, L.; Wang, X.; et al. The membrane periodic skeleton is an actomyosin network that regulates axonal diameter and conduction. eLife 2020, 9, e55471. [Google Scholar] [CrossRef]
- Vassilopoulos, S.; Gibaud, S.; Jimenez, A.; Caillol, G.; Leterrier, C. Ultrastructure of the axonal periodic scaffold reveals a braid-like organization of actin rings. Nat. Commun. 2019, 10, 5803. [Google Scholar] [CrossRef]
- Paul, D.M.; Mantell, J.; Borucu, U.; Coombs, J.; Surridge, K.J.; Squire, J.M.; Verkade, P.; Dodding, M.P. In situ cryo-electron tomography reveals filamentous actin within the microtubule lumen. J. Cell Biol. 2020, 219, 9. [Google Scholar] [CrossRef]
- Foster, H.E.; Ventura Santos, C.; Carter, A.P. A cryo-ET survey of microtubules and intracellular compartments in mammalian axons. J. Cell Biol. 2022, 221, e202103154. [Google Scholar] [CrossRef]
- Fischer, T.D.; Dash, P.K.; Liu, J.; Waxham, M.N. Morphology of mitochondria in spatially restricted axons revealed by cryo-electron tomography. PLoS Biol. 2018, 16, e2006169. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, P.C.; Giandomenico, S.L.; Ganeva, I.; Wozny, M.R.; Sutcliffe, M.; Lancaster, M.A.; Kukulski, W. Electron cryo-tomography reveals the subcellular architecture of growing axons in human brain organoids. eLife 2021, 10, e70269. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.T.; Tao, C.L.; Zhang, X.; Xia, W.; Shi, D.Q.; Qi, L.; Xu, C.; Sun, R.; Li, X.W.; Lau, P.M.; et al. Mesophasic organization of GABAA receptors in hippocampal inhibitory synapses. Nat. Neurosci. 2020, 23, 1589–1596. [Google Scholar] [CrossRef]
- Trinkaus, V.A.; Riera-Tur, I.; Martinez-Sanchez, A.; Bauerlein, F.J.B.; Guo, Q.; Arzberger, T.; Baumeister, W.; Dudanova, I.; Hipp, M.S.; Hartl, F.U.; et al. In situ architecture of neuronal alpha-Synuclein inclusions. Nat. Commun. 2021, 12, 2110. [Google Scholar] [CrossRef]
- Ibiricu, I.; Huiskonen, J.T.; Dohner, K.; Bradke, F.; Sodeik, B.; Grunewald, K. Cryo electron tomography of herpes simplex virus during axonal transport and secondary envelopment in primary neurons. PLoS Pathog. 2011, 7, e1002406. [Google Scholar] [CrossRef] [PubMed]
- Jukkola, P.I.; Lovett-Racke, A.E.; Zamvil, S.S.; Gu, C. K+ channel alterations in the progression of experimental autoimmune encephalomyelitis. Neurobiol. Dis. 2012, 47, 280–293. [Google Scholar] [CrossRef][Green Version]
- Jukkola, P.; Gu, Y.; Lovett-Racke, A.E.; Gu, C. Suppression of Inflammatory Demyelinaton and Axon Degeneration through Inhibiting Kv3 Channels. Front. Mol. Neurosci. 2017, 10, 344. [Google Scholar] [CrossRef]
- Barry, J.; Gu, Y.; Jukkola, P.; O’Neill, B.; Gu, H.; Mohler, P.J.; Rajamani, K.T.; Gu, C. Ankyrin-G directly binds to Kinesin-1 to transport voltage-gated Na+ channels into axons. Dev. Cell 2014, 28, 117–131. [Google Scholar] [CrossRef]
- Jukkola, P.; Gray, V.; Guerrero, T.; Gu, C. Astrocytes differentially respond to inflammatory autoimmune insults and imbalances of neural activity. Acta Neuropathol. Commun. 2013, 1, 70. [Google Scholar] [CrossRef]
- Nickel, M.; Eid, F.; Jukkola, P.; Gu, C. Copper chelation and autoimmunity differentially impact myelin in the hippocampal-prefrontal circuit. J. Neuroimmunol. 2019, 334, 576998. [Google Scholar] [CrossRef]
- Barry, J.; Xu, M.; Gu, Y.; Dangel, A.W.; Jukkola, P.; Shrestha, C.; Gu, C. Activation of conventional kinesin motors in clusters by Shaw voltage-gated K+ channels. J. Cell Sci. 2013, 126 Pt 9, 2027–2041. [Google Scholar] [CrossRef] [PubMed]
- Rice, J.; Weiner, J.L.; Coutellier, L.; Gu, C. Region-Specific Interneuron Demyelination and Heightened Anxiety Induced by Adolescent Binge Alcohol Drinking. Acta Neuropathol. Commun. 2019, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Cao, R.; Xiao, R.; Zhu, M.X.; Gu, C. The axon-dendrite targeting of Kv3 (Shaw) channels is determined by a targeting motif that associates with the T1 domain and ankyrin G. J. Neurosci. 2007, 27, 14158–14170. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Gu, Y.; Barry, J.; Gu, C. Kinesin I transports tetramerized Kv3 channels through the axon initial segment via direct binding. J. Neurosci. 2010, 30, 15987–16001. [Google Scholar] [CrossRef]
- Hagen, W.J.H.; Wan, W.; Briggs, J.A.G. Implementation of a cryo-electron tomography tilt-scheme optimized for high resolution subtomogram averaging. J. Struct. Biol. 2017, 197, 191–198. [Google Scholar] [CrossRef]
- Kremer, J.R.; Mastronarde, D.N.; McIntosh, J.R. Computer visualization of three-dimensional image data using IMOD. J. Struct. Biol. 1996, 116, 71–76. [Google Scholar] [CrossRef]
- Gu, Y.; Gu, C. Dynamics of Kv1 channel transport in axons. PLoS ONE 2010, 5, e11931. [Google Scholar] [CrossRef]
- Servello, D.; Gu, Y.; Gu, C. A Microbiomechanical System for Studying Varicosity Formation and Recovery in Central Neuron Axons. J. Vis. Exp. 2018, 134, e57202. [Google Scholar] [CrossRef]
- Dotti, C.G.; Sullivan, C.A.; Banker, G.A. The establishment of polarity by hippocampal neurons in culture. J. Neurosci. 1988, 8, 1454–1468. [Google Scholar] [CrossRef]
- Horton, A.C.; Ehlers, M.D. Neuronal polarity and trafficking. Neuron 2003, 40, 277–295. [Google Scholar] [CrossRef]
- Shi, S.H.; Jan, L.Y.; Jan, Y.N. Hippocampal neuronal polarity specified by spatially localized mPar3/mPar6 and PI 3-kinase activity. Cell 2003, 112, 63–75. [Google Scholar] [CrossRef]
- Gardner, A.; Jukkola, P.; Gu, C. Myelination of rodent hippocampal neurons in culture. Nat. Protoc. 2013, 7, 1774–1782. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Tian, J.H.; Pan, P.Y.; Zald, P.; Li, C.; Deng, C.; Sheng, Z.H. Docking of axonal mitochondria by syntaphilin controls their mobility and affects short-term facilitation. Cell 2008, 132, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Schwarz, T.L. The mechanism of Ca2+-dependent regulation of kinesin-mediated mitochondrial motility. Cell 2009, 136, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Press, B.; Wandinger-Ness, A. Rab 7: An important regulator of late endocytic membrane traffic. J. Cell Biol. 1995, 131 Pt 1, 1435–1452. [Google Scholar] [CrossRef] [PubMed]
- Miaczynska, M.; Zerial, M. Mosaic organization of the endocytic pathway. Exp. Cell Res. 2002, 272, 8–14. [Google Scholar] [CrossRef][Green Version]
- Kalil, K.; Dent, E.W. Branch management: Mechanisms of axon branching in the developing vertebrate CNS. Nat. Rev. Neurosci. 2014, 15, 7–18. [Google Scholar] [CrossRef]
- Bodakuntla, S.; Nedozralova, H.; Basnet, N.; Mizuno, N. Cytoskeleton and Membrane Organization at Axon Branches. Front. Cell Dev. Biol. 2021, 9, 707486. [Google Scholar] [CrossRef]
- Prokop, A. Cytoskeletal organization of axons in vertebrates and invertebrates. J. Cell Biol. 2020, 219, 7. [Google Scholar] [CrossRef]
- Dubey, P.; Jorgenson, K.; Roy, S. Actin Assemblies in the Axon Shaft-some Open Questions. Curr. Opin. Neurobiol. 2018, 51, 163–167. [Google Scholar] [CrossRef]
- Wang, T.; Li, W.; Martin, S.; Papadopulos, A.; Joensuu, M.; Liu, C.; Jiang, A.; Shamsollahi, G.; Amor, R.; Lanoue, V.; et al. Radial contractility of actomyosin rings facilitates axonal trafficking and structural stability. J. Cell Biol. 2020, 219, e201902001. [Google Scholar] [CrossRef] [PubMed]
- Chereau, R.; Saraceno, G.E.; Angibaud, J.; Cattaert, D.; Nagerl, U.V. Superresolution imaging reveals activity-dependent plasticity of axon morphology linked to changes in action potential conduction velocity. Proc. Natl. Acad. Sci. USA 2017, 114, 1401–1406. [Google Scholar] [CrossRef] [PubMed]
- Kaur, C.; Ling, E.A. Transitory cystic cavities in the developing mammalian brain-normal or anomalous? J. Anat. 2017, 230, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Gerber, Y.N.; Saint-Martin, G.P.; Bringuier, C.M.; Bartolami, S.; Goze-Bac, C.; Noristani, H.N.; Perrin, F.E. CSF1R Inhibition Reduces Microglia Proliferation, Promotes Tissue Preservation and Improves Motor Recovery after Spinal Cord Injury. Front. Cell Neurosci. 2018, 12, 368. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.T.; Gilpin, K.; Adnan, A. Effects of Focal Axonal Swelling Level on the Action Potential Signal Transmission. J. Comput. Neurosci. 2020, 48, 253–263. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, D.; Deng, B.; Sun, C.; McComb, D.W.; Gu, C. The Mechanical Microenvironment Regulates Axon Diameters Visualized by Cryo-Electron Tomography. Cells 2022, 11, 2533. https://doi.org/10.3390/cells11162533
Ma D, Deng B, Sun C, McComb DW, Gu C. The Mechanical Microenvironment Regulates Axon Diameters Visualized by Cryo-Electron Tomography. Cells. 2022; 11(16):2533. https://doi.org/10.3390/cells11162533
Chicago/Turabian StyleMa, Di, Binbin Deng, Chao Sun, David W. McComb, and Chen Gu. 2022. "The Mechanical Microenvironment Regulates Axon Diameters Visualized by Cryo-Electron Tomography" Cells 11, no. 16: 2533. https://doi.org/10.3390/cells11162533
APA StyleMa, D., Deng, B., Sun, C., McComb, D. W., & Gu, C. (2022). The Mechanical Microenvironment Regulates Axon Diameters Visualized by Cryo-Electron Tomography. Cells, 11(16), 2533. https://doi.org/10.3390/cells11162533