
Cell Cycle Regulation by Integrin-Mediated Adhesion




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Integrins
2. Cell Cycle and Cell Adhesion Crosstalk
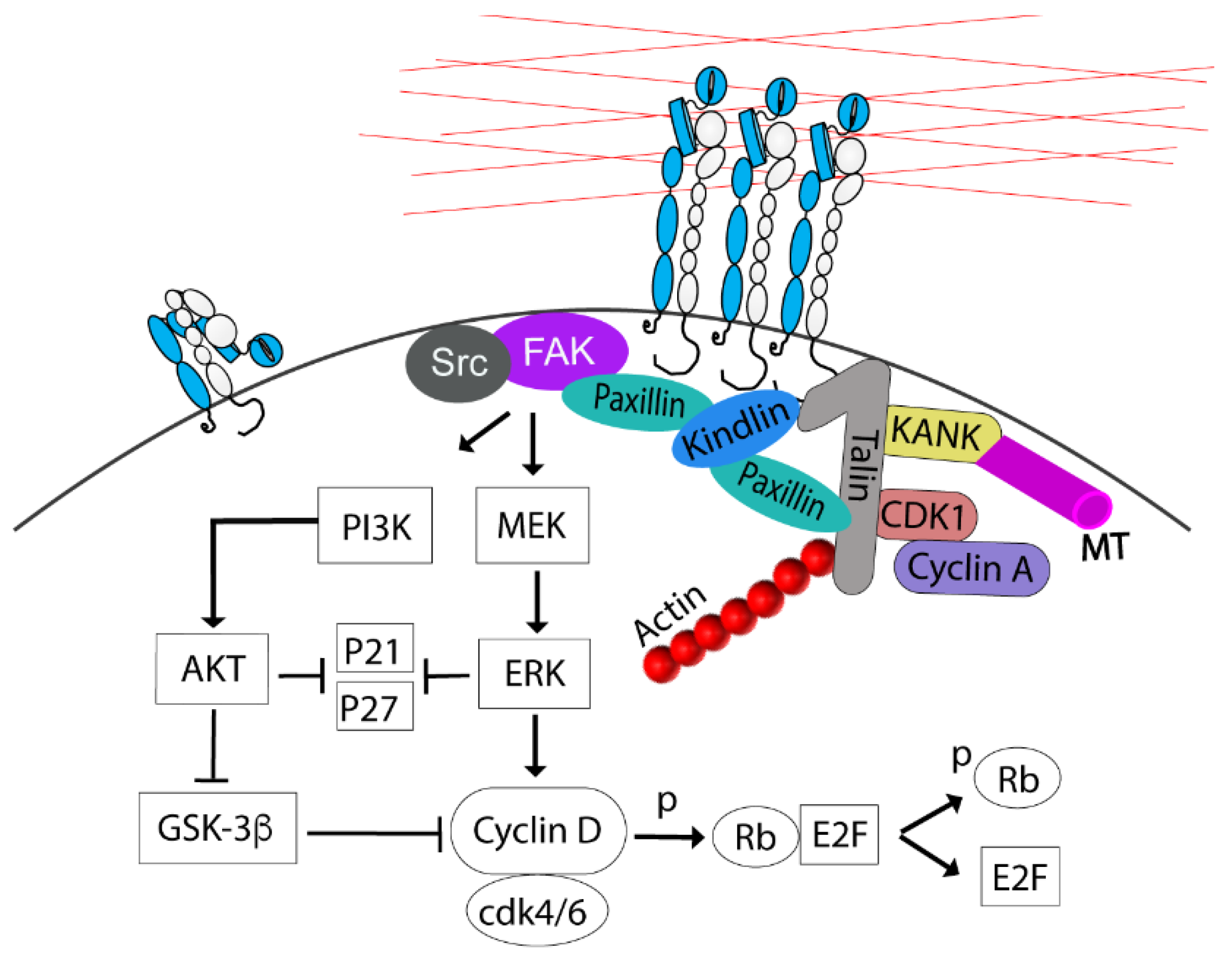
2.1. G1 to S Transition
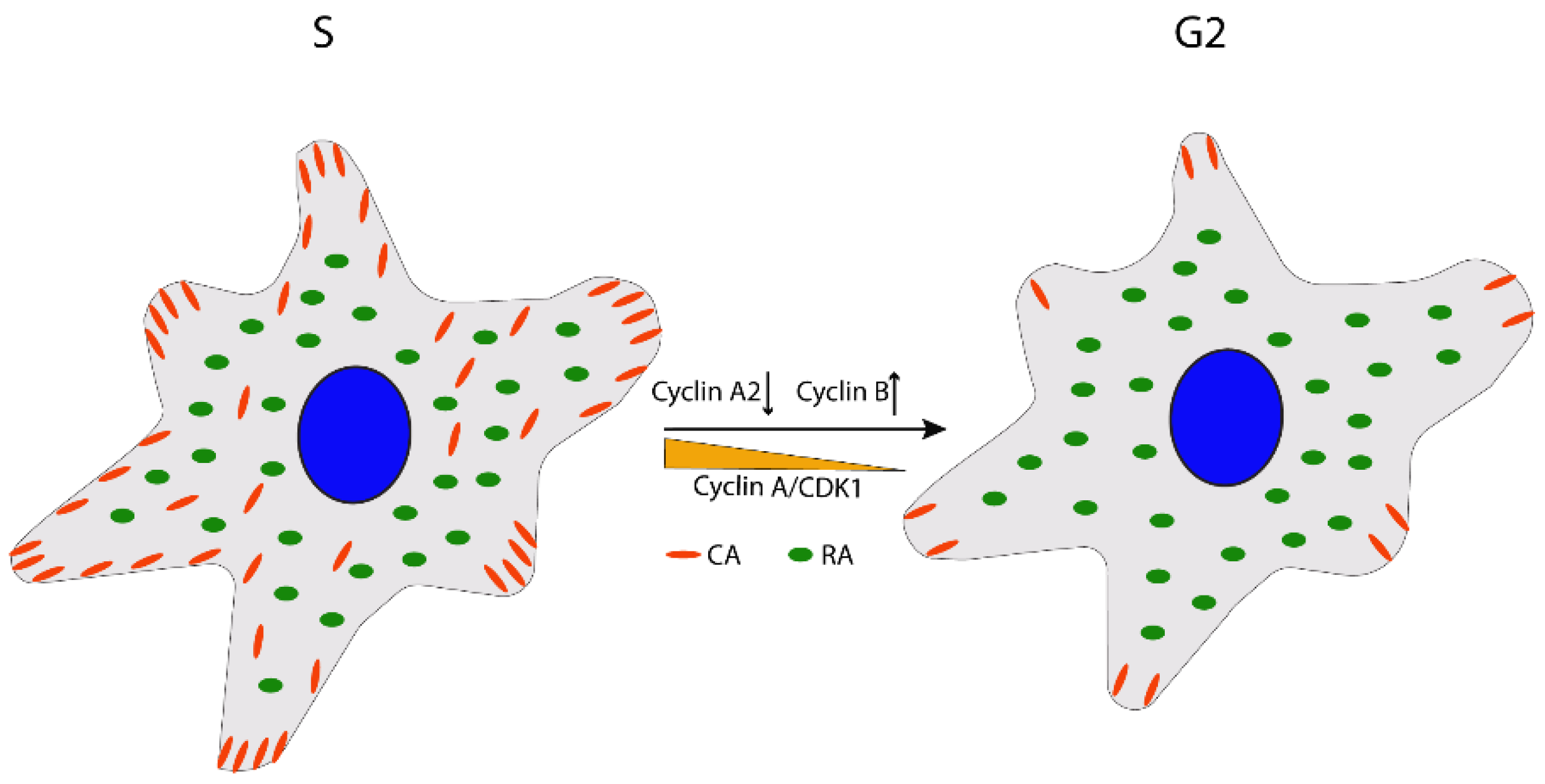
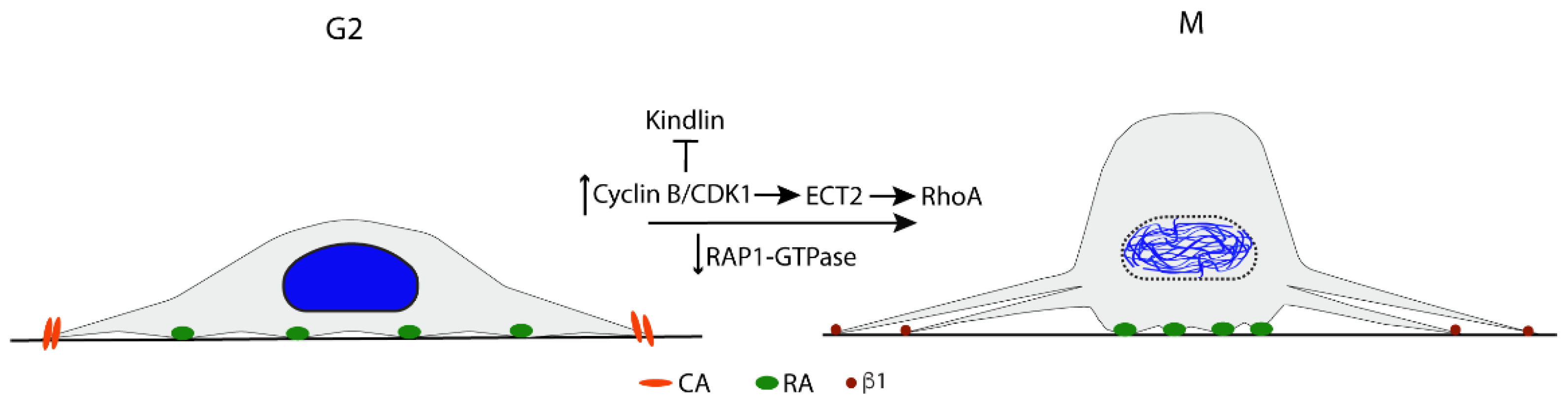
2.2. G2/M Transition
Cell Rounding
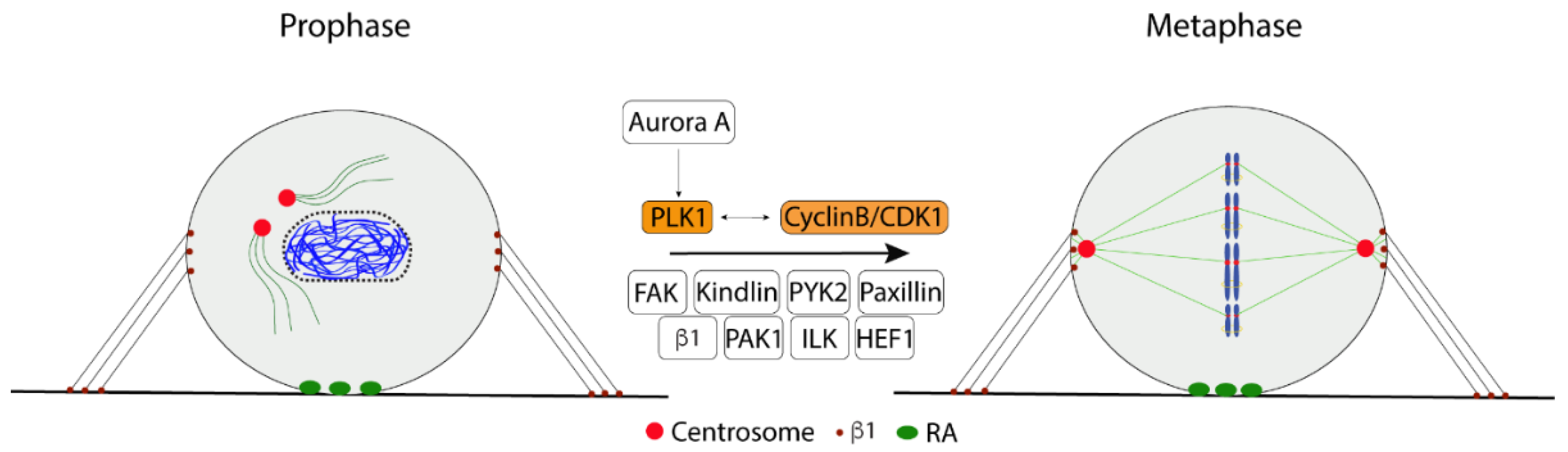
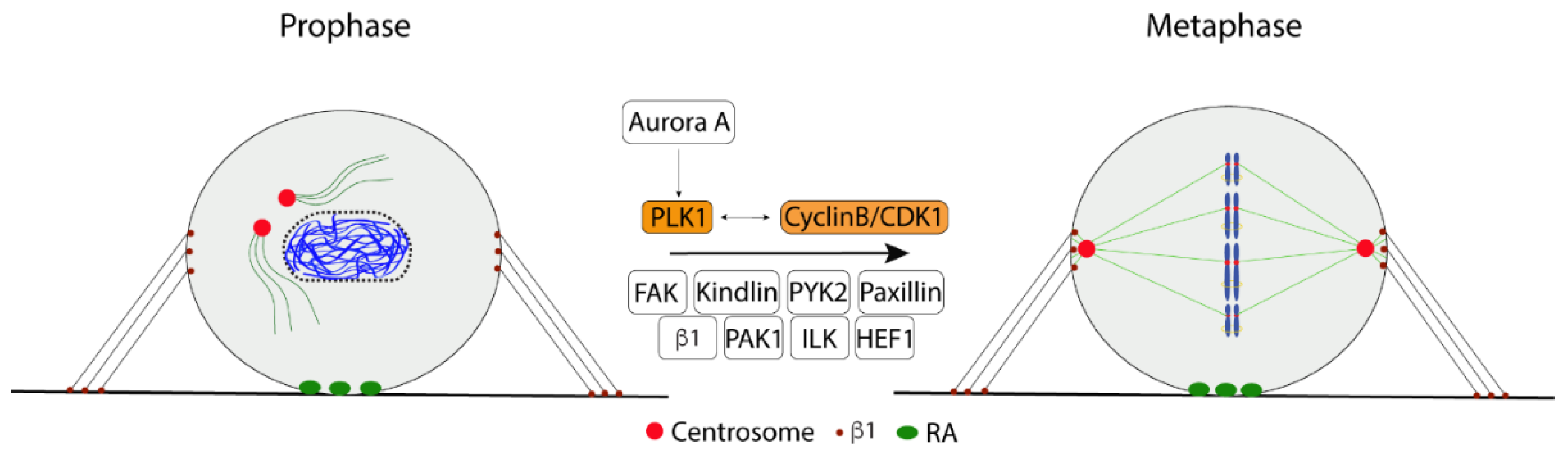
2.3. Bipolar Mitotic Spindle Formation
2.4. Spindle Orientation
2.5. Chromosome Condensation
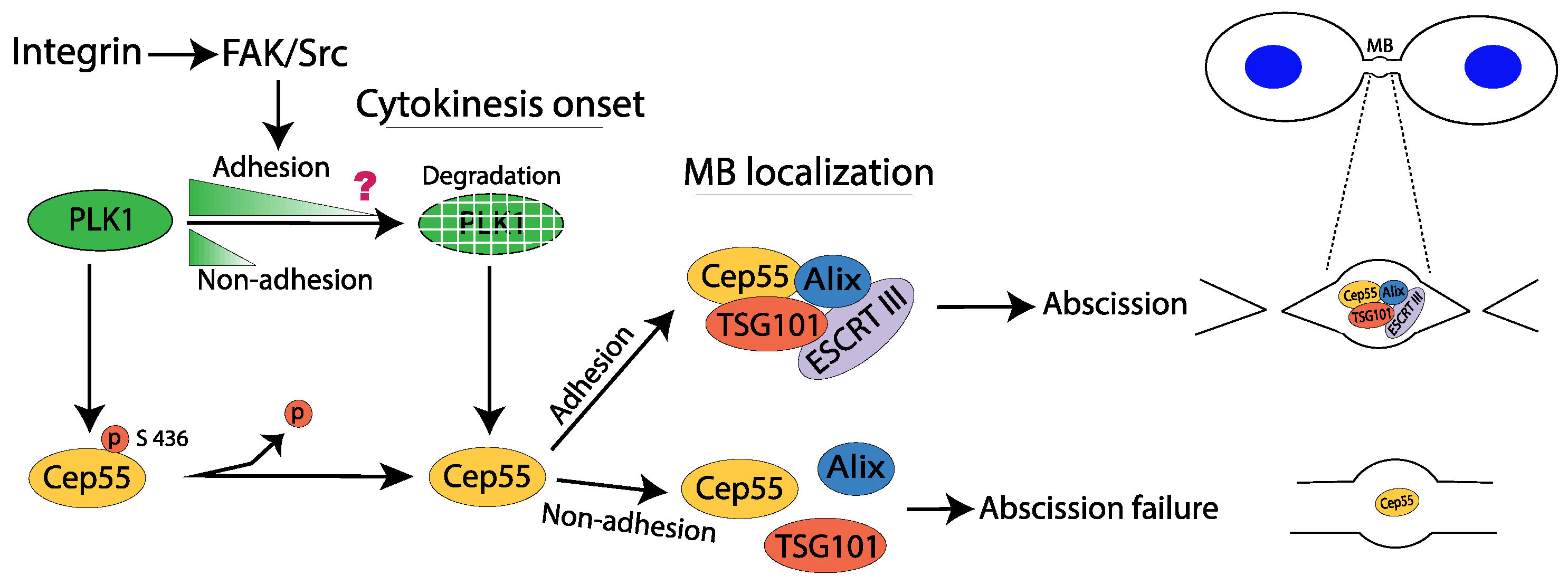
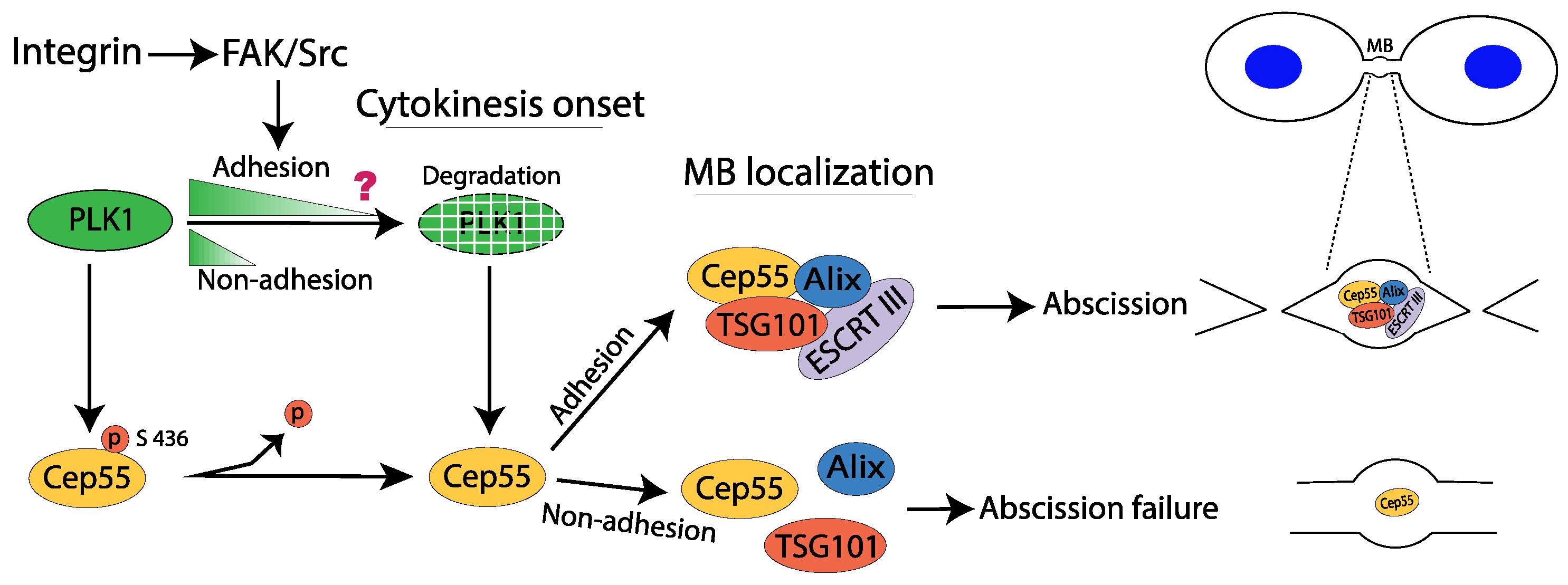
2.6. Cytokinesis
Consequences of Failed Cytokinesis
3. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miranti, C.K.; Brugge, J.S. Sensing the environment: A historical perspective on integrin signal transduction. Nat. Cell Biol. 2002, 4, E83–E90. [Google Scholar] [CrossRef] [PubMed]
- Geiger, B.; Bershadsky, A.; Pankov, R.; Yamada, K.M. Transmembrane crosstalk between the extracellular matrix—Cytoskeleton crosstalk. Nat. Rev. Mol. Cell Biol. 2001, 2, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Zhu, L.; Bromberger, T.; Yang, J.; Yang, Q.; Liu, J.; Plow, E.F.; Moser, M.; Qin, J. Mechanism of integrin activation by talin and its cooperation with kindlin. Nat. Commun. 2022, 13, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Lagarrigue, F.; Tan, B.; Du, Q.; Fan, Z.; Lopez-Ramirez, M.A.; Gingras, A.R.; Wang, H.; Qi, W.; Sun, H. Direct binding of Rap1 to Talin1 and to MRL proteins promotes integrin activation in CD4+ T cells. J. Immunol. 2022, 208, 1378–1388. [Google Scholar] [CrossRef]
- Lagarrigue, F.; Paul, D.S.; Gingras, A.R.; Valadez, A.J.; Sun, H.; Lin, J.; Cuevas, M.N.; Ablack, J.N.G.; Ramirez, M.A.L.; Bergmeier, W.; et al. Talin1 is the principal platelet Rap1 effector of Integrin activation. Blood 2020, 136, 1180–1190. [Google Scholar] [CrossRef]
- Wang, J.; Dong, X.; Zhao, B.; Li, J.; Lu, C.; Springer, T.A. Atypical interactions of integrin alphaVbeta8 with pro-TGF-beta1. Proc. Natl. Acad. Sci. USA. 2017, 114, E4168–E4174. [Google Scholar]
- Lock, J.G.; Jones, M.C.; Askari, J.A.; Gong, X.; Oddone, A.; Olofsson, H.; Göransson, S.; Lakadamyali, M.; Humphries, M.J.; Strömblad, S. Reticular adhesions are a distinct class of cell-matrix adhesions that mediate attachment during mitosis. Nat. Cell Biol. 2018, 20, 1290–1302. [Google Scholar] [CrossRef]
- Litjens, S.H.; de Pereda, J.M.; Sonnenberg, A. Current insights into the formation and breakdown of hemidesmosomes. Trends Cell Biol. 2006, 16, 376–383. [Google Scholar] [CrossRef]
- Winograd-Katz, S.E.; Fässler, R.; Geiger, B.; Legate, K.R. The integrin adhesome: From genes and proteins to human disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 273–288. [Google Scholar] [CrossRef]
- Acebrón, I.; Righetto, R.D.; Schoenherr, C.; De Buhr, S.; Redondo, P.; Culley, J.; Rodríguez, C.F.; Daday, C.; Biyani, N.; Llorca, O.; et al. Structural basis of Focal Adhesion Kinase activation on lipid membranes. EMBO J. 2020, 39, e104743. [Google Scholar] [CrossRef]
- Calalb, M.B.; Polte, T.R.; Hanks, S.K. Tyrosine phosphorylation of focal adhesion kinase at sites in the catalytic domain regulates kinase activity: A role for Src family kinases. Mol. Cell. Biol. 1995, 15, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Appeddu, P.A.; Isoda, H.; Guan, J.L. Phosphorylation of tyrosine 397 in focal adhesion kinase is required for binding phosphatidylinositol 3-kinase. J. Biol. Chem. 1996, 271, 26329–26334. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, B.P.; Gough, R.E.; Ammon, Y.C.; van de Willige, D.; Post, H.; Jacquemet, G.; Altelaar, A.M.; Heck, A.J.; Goult, B.T.; Akhmanova, A. Talin-KANK1 interaction controls the recruitment of cortical microtubule stabilizing complexes to focal adhesions. Elife 2016, 5, e18124. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ouyang, M.; Van den Dries, K.; McGhee, E.J.; Tanaka, K.; Anderson, M.D.; Groisman, A.; Goult, B.T.; Anderson, K.I.; Schwartz, M.A. Talin tension sensor reveals novel features of focal adhesion force transmission and mechanosensitivity. J. Cell Biol. 2016, 213, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Zuidema, A.; Wang, W.; Kreft, M.; Te Molder, L.; Hoekman, L.; Bleijerveld, O.B.; Nahidiazar, L.; Janssen, H.; Sonnenberg, A. Mechanisms of integrin alphaVbeta5 clustering in flat clathrin lattices. J. Cell Sci. 2018, 131, 221317. [Google Scholar] [CrossRef]
- Te Molder, L.; de Pereda, J.M.; Sonnenberg, A. Regulation of hemidesmosome dynamics and cell signaling by integrin alpha6beta4. J. Cell Sci. 2021, 134, 259004. [Google Scholar] [CrossRef]
- Dontu, G.; Wicha, M.S. Survival of mammary stem cells in suspension culture: Implications for stem cell biology and neoplasia. J. Mammary Gland Biol. Neoplasia. 2005, 10, 75–86. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Ingram, P.N.; Fouladdel, S.; McDermott, S.P.; Azizi, E.; Wicha, M.S.; Yoon, E. High-Throughput Single-Cell Derived Sphere Formation for Cancer Stem-Like Cell Identification and Analysis. Sci. Rep. 2016, 6, 27301. [Google Scholar] [CrossRef]
- Aplin, A.E.; Howe, A.K.; Juliano, R.L. Cell adhesion molecules, signal transduction and cell growth. Curr. Opin. Cell Biol. 1999, 11, 737–744. [Google Scholar] [CrossRef]
- Schwartz, M.A.; Assoian, R.K. Integrins and cell proliferation: Regulation of cyclin-dependent kinases via cytoplasmic signaling pathways. J. Cell Sci. 2001, 114, 2553–2560. [Google Scholar] [CrossRef]
- Kamranvar, S.A.; Gupta, D.K.; Huang, Y.; Gupta, R.K.; Johansson, S. Integrin signaling via FAK-Src controls cytokinetic abscission by decelerating PLK1 degradation and subsequent recruitment of CEP55 at the midbody. Oncotarget 2016, 7, 30820–30830. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.C.; Askari, J.A.; Humphries, J.D.; Humphries, M.J. Cell adhesion is regulated by CDK1 during the cell cycle. J. Cell Biol. 2018, 217, 3203–3218. [Google Scholar] [CrossRef] [PubMed]
- Assoian, R.K.; Schwartz, M.A. Coordinate signaling by integrins and receptor tyrosine kinases in the regulation of G1 phase cell-cycle progression. Curr. Opin. Genet. Dev. 2001, 11, 48–53. [Google Scholar] [CrossRef]
- Zhu, X.; Ohtsubo, M.; Böhmer, R.M.; Roberts, J.M.; Assoian, R.K. Adhesion-dependent cell cycle progression linked to the expression of cyclin D1, activation of cyclin E-cdk2, and phosphorylation of the retinoblastoma protein. J. Cell Biol. 1996, 133, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, K.; Block, K.; Nayak, B.K.; Livi, C.B.; Venkatachalam, M.A.; Sudarshan, S. PI3K regulation of the SKP-2/p27 axis through mTORC2. Oncogene 2013, 32, 2027–2036. [Google Scholar] [CrossRef]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef]
- Zhao, J.; Pestell, R.; Guan, J.L. Transcriptional activation of cyclin D1 promoter by FAK contributes to cell cycle progression. Mol. Biol. Cell 2001, 12, 4066–4077. [Google Scholar] [CrossRef]
- Zhao, J.; Bian, Z.; Yee, K.; Chen, B.P.; Chien, S.; Guan, J.L. Identification of transcription factor KLF8 as a downstream target of focal adhesion kinase in its regulation of cyclin D1 and cell cycle progression. Mol. Cell 2003, 11, 1503–1515. [Google Scholar] [CrossRef]
- Diehl, J.A.; Cheng, M.; Roussel, M.F.; Sherr, C.J. Glycogen synthase kinase-3beta regulates cyclin D1 proteolysis and subcellular localization. Genes Dev. 1998, 12, 3499–3511. [Google Scholar] [CrossRef]
- Streuli, C.H.; Akhtar, N. Signal co-operation between integrins and other receptor systems. Biochem. J. 2009, 418, 491–506. [Google Scholar] [CrossRef]
- Assoian, R.K. Anchorage-dependent cell cycle progression. J. Cell Biol. 1997, 136, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.; Jacquemet, G.; Byron, A.; Jones, M.C.; Warwood, S.; Selley, J.N.; Knight, D.; Humphries, J.; Humphries, M. Defining the phospho-adhesome through the phosphoproteomic analysis of integrin signalling. Nat. Commun. 2015, 6, 6265. [Google Scholar] [CrossRef] [PubMed]
- Gough, R.E.; Jones, M.C.; Zacharchenko, T.; Le, S.; Yu, M.; Jacquemet, G.; Muench, S.P.; Yan, J.; Humphries, J.D.; Jørgensen, C.; et al. Talin mechanosensitivity is modulated by a direct interaction with cyclin-dependent kinase-1. J. Biol. Chem. 2021, 297, 100837. [Google Scholar] [CrossRef] [PubMed]
- Elbæk, C.R.; Petrosius, V.; Sørensen, C.S. WEE1 kinase limits CDK activities to safeguard DNA replication and mitotic entry. Mutat. Res. Mol. Mech. Mutagen. 2020, 819–820, 111694. [Google Scholar] [CrossRef]
- Gheghiani, L.; Loew, D.; Lombard, B.; Mansfeld, J.; Gavet, O. PLK1 activation in late G2 sets up commitment to mitosis. Cell Rep. 2017, 19, 2060–2073. [Google Scholar] [CrossRef]
- Kamranvar, S.A.; Gupta, D.K.; Wasberg, A.; Liu, L.; Roig, J.; Johansson, S. Integrin-Mediated Adhesion Promotes Centrosome Separation in Early Mitosis. Cells 2022, 11, 1360. [Google Scholar] [CrossRef]
- Vianay, B.; Senger, F.; Alamos, S.; Anjur-Dietrich, M.; Bearce, E.; Cheeseman, B.; Lee, L.; Théry, M. Variation in traction forces during cell cycle progression. Biol. Cell 2018, 110, 91–96. [Google Scholar] [CrossRef]
- Dao, V.T.; Dupuy, A.G.; Gavet, O.; Caron, E.; de Gunzburg, J. Dynamic changes in Rap1 activity are required for cell retraction and spreading during mitosis. J. Cell Sci. 2009, 122, 2996–3004. [Google Scholar] [CrossRef]
- Marchesi, S.; Montani, F.; Deflorian, G.; D’Antuono, R.; Cuomo, A.; Bologna, S.; Mazzoccoli, C.; Bonaldi, T.; Di Fiore, P.P.; Nicassio, F. DEPDC1B coordinates de-adhesion events and cell-cycle progression at mitosis. Dev. Cell 2014, 31, 420–433. [Google Scholar] [CrossRef]
- Lancaster, O.M.; Le Berre, M.; Dimitracopoulos, A.; Bonazzi, D.; Zlotek-Zlotkiewicz, E.; Picone, R.; Duke, T.; Piel, M.; Baum, B. Mitotic rounding alters cell geometry to ensure efficient bipolar spindle formation. Dev. Cell 2013, 25, 270–283. [Google Scholar] [CrossRef]
- Jefford, C.E.; Irminger-Finger, I. Mechanisms of chromosome instability in cancers. Crit. Rev. Oncol. 2006, 59, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, A.V.; Baum, B.; Matthews, H.K. The mechanics of mitotic cell rounding. Front. Cell Dev. Biol. 2020, 8, 687. [Google Scholar] [CrossRef] [PubMed]
- Matthews, H.K.; Delabre, U.; Rohn, J.L.; Guck, J.; Kunda, P.; Baum, B. Changes in Ect2 localization couple actomyosin-dependent cell shape changes to mitotic progression. Dev. Cell 2012, 23, 371–383. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.-P.; Aretz, J.; Fässler, R. CDK1–cyclin-B1-induced kindlin degradation drives focal adhesion disassembly at mitotic entry. Nat. Cell Biol. 2022, 24, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Gavet, O.; Pines, J. Progressive activation of CyclinB1-Cdk1 coordinates entry to mitosis. Dev. Cell. 2010, 18, 533–543. [Google Scholar] [CrossRef]
- Ezratty, E.J.; Partridge, M.A.; Gundersen, G.G. Microtubule-induced focal adhesion disassembly is mediated by dynamin and focal adhesion kinase. Nat. Cell Biol. 2005, 7, 581–590. [Google Scholar] [CrossRef]
- Moreno-Layseca, P.; Icha, J.; Hamidi, H.; Ivaska, J. Integrin trafficking in cells and tissues. Nat. Cell Biol. 2019, 21, 122–132. [Google Scholar] [CrossRef]
- Yamakita, Y.; Totsukawa, G.; Yamashiro, S.; Fry, D.; Zhang, X.; Hanks, S.K.; Matsumura, F. Dissociation of FAK/p130(CAS)/c-Src complex during mitosis: Role of mitosis-specific serine phosphorylation of FAK. J. Cell Biol. 1999, 144, 315–324. [Google Scholar] [CrossRef]
- Reverte, C.G.; Benware, A.; Jones, C.W.; LaFlamme, S.E. Perturbing integrin function inhibits microtubule growth from centrosomes, spindle assembly, and cytokinesis. J. Cell Biol. 2006, 174, 491–497. [Google Scholar] [CrossRef]
- Bertran, M.T.; Sdelci, S.; Regue, L.; Avruch, J.; Caelles, C.; Roig, J. Nek9 is a Plk1-activated kinase that controls early centrosome separation through Nek6/7 and Eg5. EMBO J. 2011, 30, 2634–2647. [Google Scholar] [CrossRef]
- Pintard, L.; Archambault, V. A unified view of spatio-temporal control of mitotic entry: Polo kinase as the key. Open Biol. 2018, 8, 180114. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; Morisaki, T.; Nishiyama, Y.; Marumoto, T.; Tada, K.; Hara, T.; Masuko, N.; Inagaki, M.; Hatakeyama, K.; Saya, H. Zyxin, a regulator of actin filament assembly, targets the mitotic apparatus by interacting with H-Warts/Lats1 tumor suppressor. J. Cell Biol. 2000, 149, 1073–1086. [Google Scholar] [CrossRef] [PubMed]
- Herreros, L.; Rodríguez-Fernández, J.L.; Brown, M.C.; Alonso-Lebrero, J.L.; Cabañas, C.; Sánchez-Madrid, F.; Longo, N.; Turner, C.E.; Sánchez-Mateos, P. Paxillin localizes to the lymphocyte microtubule organizing center and associates with the microtubule cytoskeleton. J. Biol. Chem. 2000, 275, 26436–26440. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Fernández, J.L.; Gómez, M.; Luque, A.; Hogg, N.; Sánchez-Madrid, F.; Cabañas, C. The interaction of activated integrin lymphocyte function-associated antigen 1 with ligand intercellular adhesion molecule 1 induces activation and redistribution of focal adhesion kinase and proline-rich tyrosine kinase 2 in T lymphocytes. Mol. Biol. Cell 1999, 10, 1891–1907. [Google Scholar] [CrossRef] [PubMed]
- Pugacheva, E.N.; Golemis, E.A. The focal adhesion scaffolding protein HEF1 regulates activation of the Aurora-A and Nek2 kinases at the centrosome. Nat. Cell Biol. 2005, 7, 937–946. [Google Scholar] [CrossRef]
- Fielding, A.; Dobreva, I.; McDonald, P.C.; Foster, L.J.; Dedhar, S. Integrin-linked kinase localizes to the centrosome and regulates mitotic spindle organization. J. Cell Biol. 2008, 180, 681–689. [Google Scholar] [CrossRef]
- Maroto, B.; Ye, M.B.; von Lohneysen, K.; Schnelzer, A.; Knaus, U.G. P21-activated kinase is required for mitotic progression and regulates Plk1. Oncogene 2008, 27, 4900–4908. [Google Scholar] [CrossRef]
- Patel, H.; Stavrou, I.; Shrestha, R.L.; Draviam, V.; Frame, M.C.; Brunton, V.G. Kindlin1 regulates microtubule function to ensure normal mitosis. J. Mol. Cell Biol. 2016, 8, 338–348. [Google Scholar] [CrossRef]
- Patel, H.; Zich, J.; Serrels, B.; Rickman, C.; Hardwick, K.G.; Frame, M.C.; Brunton, V.G. Kindlin-1 regulates mitotic spindle formation by interacting with integrins and Plk-1. Nat. Commun. 2013, 4, 2056. [Google Scholar] [CrossRef]
- Tan, H.F.; Tan, S.M. The focal adhesion protein kindlin-2 controls mitotic spindle assembly by inhibiting histone deacetylase 6 and maintaining α-tubulin acetylation. J. Biol. Chem. 2020, 295, 5928–5943. [Google Scholar] [CrossRef]
- Rea, K.; Sensi, M.; Anichini, A.; Canevari, S.; Tomassetti, A. EGFR/MEK/ERK/CDK5-dependent integrin-independent FAK phosphorylated on serine 732 contributes to microtubule depolymerization and mitosis in tumor cells. Cell Death Dis. 2013, 4, e815. [Google Scholar] [CrossRef] [PubMed]
- Fischer, E.; Legue, E.; Doyen, A.; Nato, F.; Nicolas, J.-F.; Torres, V.; Yaniv, M.; Pontoglio, M. Defective planar cell polarity in polycystic kidney disease. Nat. Genet. 2006, 38, 21–23. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, C. Spindle orientation, asymmetric division and tumour suppression in Drosophila stem cells. Nat. Rev. Genet. 2007, 8, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, F.; Nishida, E. Spindle orientation in animal cell mitosis: Roles of integrin in the control of spindle axis. J. Cell. Physiol. 2007, 213, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima, F.; Matsumura, S.; Morimoto, H.; Mitsushima, M.; Nishida, E. PtdIns(3,4,5)P3 regulates spindle orientation in adherent cells. Dev. Cell. 2007, 13, 796–811. [Google Scholar] [CrossRef] [PubMed]
- Schuyler, S.C.; Pellman, D. Microtubule “plus-end-tracking proteins”: The end is just the beginning. Cell. 2001, 105, 421–424. [Google Scholar] [CrossRef]
- Woodard, G.E.; Huang, N.N.; Cho, H.; Miki, T.; Tall, G.G.; Kehrl, J.H. Ric-8A and Giα recruit LGN, NuMA, and dynein to the cell cortex to help orient the mitotic spindle. Mol. Cell. Biol. 2010, 30, 3519–3530. [Google Scholar] [CrossRef]
- Okumura, M.; Natsume, T.; Kanemaki, M.T.; Kiyomitsu, T. Dynein-Dynactin-NuMA clusters generate cortical spindle-pulling forces as a multi-arm ensemble. Elife. 2018, 7, e36559. [Google Scholar] [CrossRef]
- Maier, B.; Kirsch, M.; Anderhub, S.; Zentgraf, H.; Krämer, A. The novel actin/focal adhesion-associated protein MISP is involved in mitotic spindle positioning in human cells. Cell Cycle 2013, 12, 1457–1471. [Google Scholar] [CrossRef]
- Petridou, N.I.; Skourides, P.A. A ligand-independent integrin beta1 mechanosensory complex guides spindle orientation. Nat Commun 2016, 7, 10899. [Google Scholar] [CrossRef]
- Humphries, J.D.; Byron, A.; Bass, M.D.; Craig, S.E.; Pinney, J.W.; Knight, D.; Humphries, M.J. proteomic analysis of integrin-associated complexes identifies RCC2 as a dual regulator of Rac1 and Arf6. Sci. Signal. 2009, 2, ra51. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, S.J.; Gontarczyk, A.M.; Alghamdi, A.A.; Ellison, T.S.; Johnson, R.T.; Fowler, W.J.; Kirkup, B.M.; Silva, B.C.; Harry, B.E.; Schneider, J.G.; et al. The beta3-integrin endothelial adhesome regulates microtubule-dependent cell migration. EMBO Rep. 2018, 19, e44578. [Google Scholar] [CrossRef] [PubMed]
- Mollinari, C.; Reynaud, C.; Martineau-Thuillier, S.; Monier, S.; Kieffer, S.; Garin, J.; Andreassen, P.R.; Boulet, A.; Goud, B.; Kleman, J.-P.; et al. The mammalian passenger protein TD-60 is an RCC1 family member with an essential role in prometaphase to metaphase progression. Dev. Cell. 2003, 5, 295–307. [Google Scholar] [CrossRef]
- Papini, D.; Langemeyer, L.; Abad, M.A.; Kerr, A.; Samejima, I.; Eyers, P.A.; Jeyaprakash, A.A.; Higgins, J.M.G.; Barr, F.A.; Earnshaw, W.C. TD-60 links RalA GTPase function to the CPC in mitosis. Nat. Commun. 2015, 6, 7678. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.C.; Cowell, C.A.M.; Hammond, C.L.; Bergen, D.J.M.; Roper, J.A.; Feng, Y.; Rendall, T.; Race, P.R.; Bass, M.D. Coronin-1C and RCC2 guide mesenchymal migration by trafficking Rac1 and controlling GEF exposure. J. Cell Sci. 2014, 127, 4292–4307. [Google Scholar] [CrossRef]
- Hognas, G.; Tuomi, S.; Veltel, S.; Mattila, E.; Murumagi, A.; Edgren, H.; Kallioniemi, O.; Ivaska, J. Cytokinesis failure due to derailed integrin traffic induces aneuploidy and oncogenic transformation in vitro and in vivo. Oncogene 2011, 31, 3597–3606. [Google Scholar] [CrossRef]
- Thullberg, M.; Gad, A.; Le Guyader, S.; Strömblad, S. Oncogenic H-Ras V12 promotes anchorage-independent cytokinesis in human fibroblasts. Proc. Natl. Acad. Sci. USA 2007, 104, 20338–20343. [Google Scholar] [CrossRef]
- Mierzwa, B.; Gerlich, D.W. Cytokinetic abscission: Molecular mechanisms and temporal control. Dev. Cell. 2014, 31, 525–538. [Google Scholar] [CrossRef]
- Taneja, N.; Fenix, A.; Rathbun, L.; Millis, B.A.; Tyska, M.J.; Hehnly, H.; Burnette, D.T. Focal adhesions control cleavage furrow shape and spindle tilt during mitosis. Sci. Rep. 2016, 6, 29846. [Google Scholar] [CrossRef]
- Mathew, S.S.; Nieves, B.; Sequeira, S.; Sambandamoorthy, S.; Pumiglia, K.; Larsen, M.; LaFlamme, S.E. integrins promote cytokinesis through the RSK signaling Axis. J. Cell Sci. 2013, 127, 534–545. [Google Scholar] [CrossRef]
- Pellinen, T.; Tuomi, S.; Arjonen, A.; Wolf, M.; Edgren, H.; Meyer, H.; Grosse, R.; Kitzing, T.; Rantala, J.K.; Kallioniemi, O.; et al. Integrin trafficking regulated by Rab21 is necessary for cytokinesis. Dev. Cell 2008, 15, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Sambandamoorthy, S.; Mathew-Steiner, S.; Varney, S.; Zuidema, J.M.; Gilbert, R.J.; Van De Water, L.; LaFlamme, S.E. Matrix compliance and the regulation of cytokinesis. Biol. Open 2015, 4, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Bastos, R.N.; Barr, F.A. Plk1 negatively regulates Cep55 recruitment to the midbody to ensure orderly abscission. J. Cell Biol. 2010, 191, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Ganem, N.J.; Godinho, S.A.; Pellman, D. A mechanism linking extra centrosomes to chromosomal instability. Nature 2009, 460, 278–282. [Google Scholar] [CrossRef]
- Godinho, S.A.; Kwon, M.; Pellman, D. Centrosomes and cancer: How cancer cells divide with too many centrosomes. Cancer Metastasis Rev. 2009, 28, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Fava, L.L.; Schuler, F.; Sladky, V.; Haschka, M.D.; Soratroi, C.; Eiterer, L.; Demetz, E.; Weiss, G.; Geley, S.; Nigg, E.A.; et al. The PIDDosome activates p53 in response to supernumerary centrosomes. Genes Dev. 2017, 31, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Burigotto, M.; Fava, L.L. The PIDDosome: Centrosome guardian and backup on the DNA damage response. Mol. Cell Oncol. 2021, 8, 1893625. [Google Scholar] [CrossRef]
- Gupta, D.K.; Kamranvar, S.A.; Du, J.; Liu, L.; Johansson, S. Septin and Ras regulate cytokinetic abscission in detached cells. Cell Div. 2019, 14, 1–15. [Google Scholar] [CrossRef]
- Kumari, R.; Jat, P. Mechanisms of cellular senescence: Cell cycle arrest and senescence associated secretory phenotype. Front. Cell Dev. Biol. 2021, 9, 645593. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Saqcena, M.; Foster, D.A. Synthetic lethality in KRas-driven cancer cells created by glutamine deprivation. Oncoscience 2015, 2, 807–808. [Google Scholar] [CrossRef]
- Lee, I.H.; Finkel, T. Metabolic regulation of the cell cycle. Curr. Opin. Cell Biol. 2013, 25, 724–729. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamranvar, S.A.; Rani, B.; Johansson, S. Cell Cycle Regulation by Integrin-Mediated Adhesion. Cells 2022, 11, 2521. https://doi.org/10.3390/cells11162521
Kamranvar SA, Rani B, Johansson S. Cell Cycle Regulation by Integrin-Mediated Adhesion. Cells. 2022; 11(16):2521. https://doi.org/10.3390/cells11162521
Chicago/Turabian StyleKamranvar, Siamak A., Bhavna Rani, and Staffan Johansson. 2022. "Cell Cycle Regulation by Integrin-Mediated Adhesion" Cells 11, no. 16: 2521. https://doi.org/10.3390/cells11162521
APA StyleKamranvar, S. A., Rani, B., & Johansson, S. (2022). Cell Cycle Regulation by Integrin-Mediated Adhesion. Cells, 11(16), 2521. https://doi.org/10.3390/cells11162521