
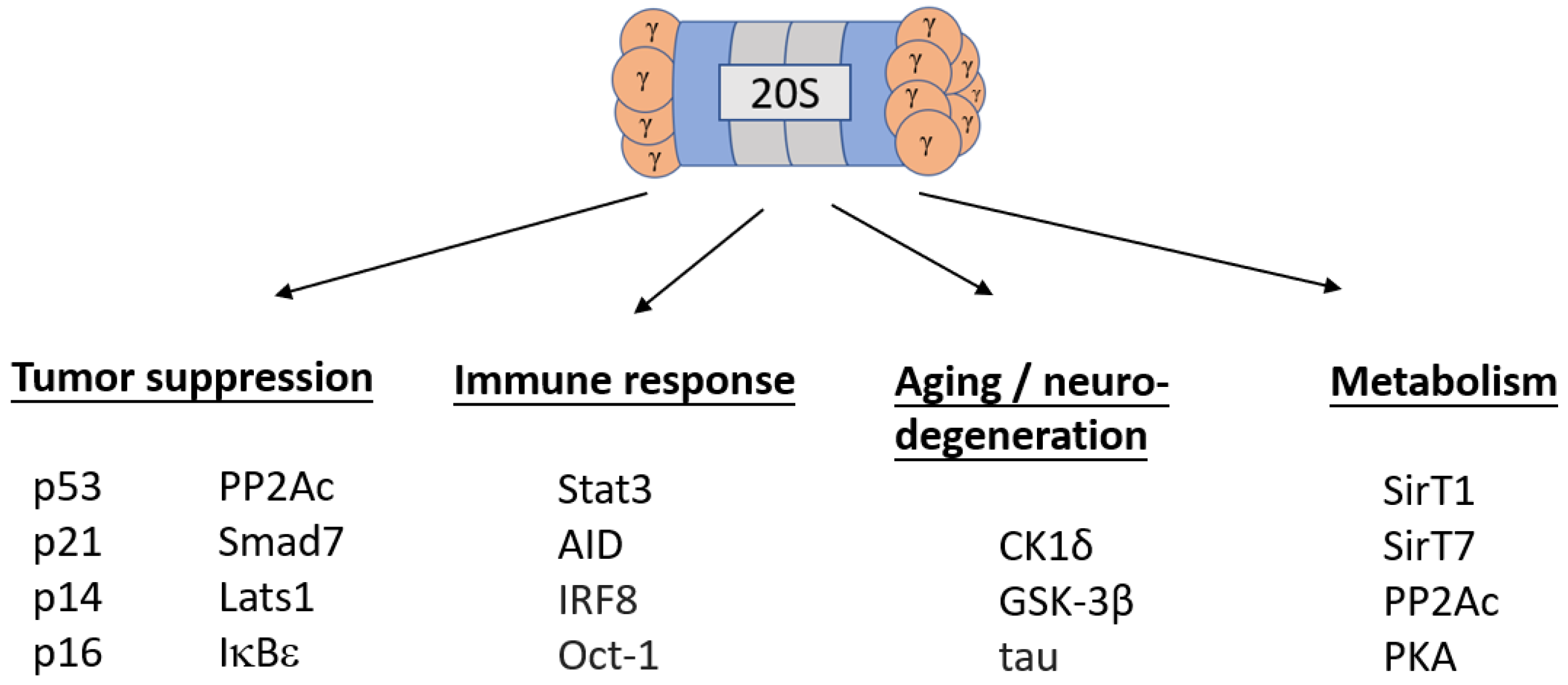
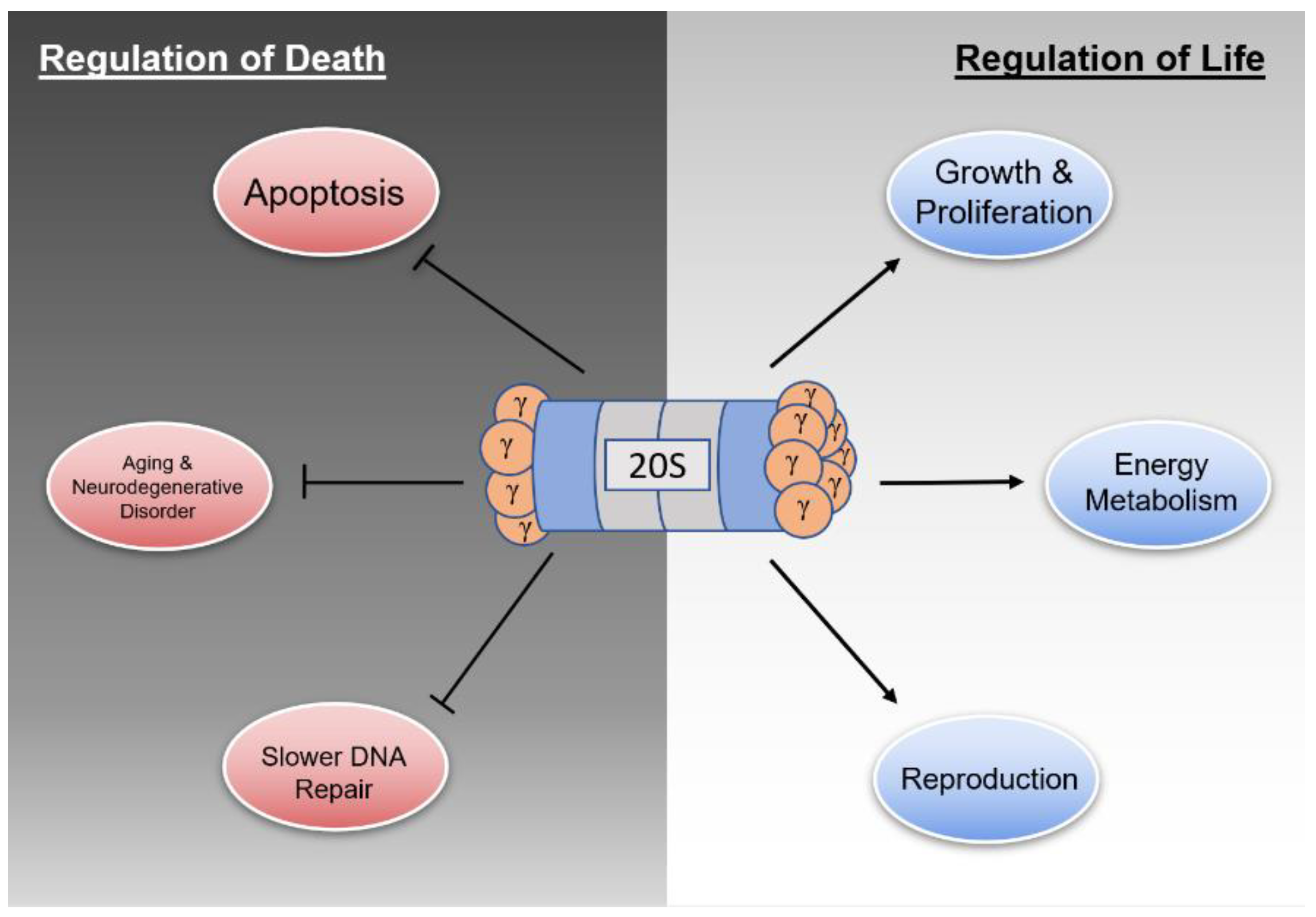
Regulation of Life & Death by REGγ


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Regulation of Life
2.1. Cell Growth and Proliferation
2.2. Energy Metabolism
2.3. Reproduction
3. Regulation of Death
3.1. Apoptosis
3.2. Aging & Neurodegenerative Disease
3.3. DNA Damage Repair & Chromosomal Stability
4. Regulation of the Regulator
4.1. Transcriptional Regulation
4.2. Post-transcriptional Regulation
4.3. Post-translational Regulation
5. Conclusions and Future Directions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hochstrasser, M. Ubiquitin, Proteasomes, and the Regulation of Intracellular Degradation. Curr. Opin. Cell Biol. 1995, 7, 215–223. [Google Scholar] [CrossRef]
- Tanaka, K. The Proteasome: Overview of Structure and Functions. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 12–36. [Google Scholar] [CrossRef]
- Cascio, P. PA28γ: New Insights on an Ancient Proteasome Activator. Biomolecules 2021, 11, 228. [Google Scholar] [CrossRef]
- Li, J.; Rechsteiner, M. Molecular Dissection of the 11S REG (PA28) Proteasome. Biochimie 2001, 83, 373–383. [Google Scholar] [CrossRef]
- Wójcik, C.; Tanaka, K.; Paweletz, N.; Naab, U.; Wilk, S. Proteasome Activator (PA28) Subunits, α, β and γ (Ki Antigen) in NT2 Neuronal Precursor Cells and HeLa S3 Cells. Eur. J. Cell Biol. 1998, 77, 151–160. [Google Scholar] [CrossRef]
- Zannini, L.; Lecis, D.; Buscemi, G.; Carlessi, L.; Gasparini, P.; Fontanella, E.; Lisanti, S.; Barton, L.; Delia, D. REGgamma Proteasome Activator Is Involved in the maintenance of Chromosomal Stability. Cell Cycle 2008, 7, 504–512. [Google Scholar] [CrossRef]
- Fesquet, D.; Llères, D.; Grimaud, C.; Viganò, C.; Méchali, F.; Boulon, S.; Coux, O.; Bonne-Andrea, C.; Baldin, V. The 20S Proteasome Activator PA28γ Controls the compaction of Chromatin. J. Cell Sci. 2021, 134, jcs257717. [Google Scholar] [CrossRef]
- Fabre, B.; Lambour, T.; Garrigues, L.; Ducoux-Petit, M.; Amalric, F.; Monsarrat, B.; Burlet-Schiltz, O.; Bousquet-Dubouch, M.-P. Label-Free Quantitative Proteomics Reveals the Dynamics of Proteasome Complexes Composition and Stoichiometry in a Wide Range of Human Cell Lines. J. Proteome Res. 2014, 13, 3027–3037. [Google Scholar] [CrossRef]
- Li, X.; Lonard, D.M.; Jung, S.Y.; Malovannaya, A.; Feng, Q.; Qin, J.; Tsai Sophia, Y.; Tsai, M.-J.; O’Malley, B.W. The SRC-3/AIB1 Coactivator Is Degraded in a Ubiquitin- and ATP-Independent Manner by the REGgamma Proteasome. Cell 2006, 124, 381–392. [Google Scholar] [CrossRef]
- Li, L.; Zhao, D.; Wei, H.; Yao, L.; Dang, Y.; Amjad, A.; Xu, J.; Liu, J.; Guo, L.; Li, D.; et al. REGγ Deficiency Promotes Premature Aging via the Casein 1 Pathway. Proc. Natl. Acad. Sci. USA 2013, 110, 11005–11010. [Google Scholar] [CrossRef]
- Li, X.; Amazit, L.; Long, W.; Lonard, D.M.; Monaco, J.J.; O’Malley, B.W. Ubiquitin- and ATP-Independent Proteolytic Turnover of P21 by the REGgamma-Proteasome Pathway. Mol. Cell 2007, 26, 831–842. [Google Scholar] [CrossRef]
- Chen, X.; Barton, L.F.; Chi, Y.; Clurman, B.E.; Roberts, J.M. Ubiquitin-Independent Degradation of Cell-Cycle Inhibitors by the REGgamma Proteasome. Mol. Cell 2007, 26, 843–852. [Google Scholar] [CrossRef]
- Wang, Q.; Gao, X.; Yu, T.; Yuan, L.; Dai, J.; Wang, W.; Chen, G.; Jiao, C.; Zhou, W.; Huang, Q.; et al. REGγ Controls Hippo Signaling and Reciprocal-ΚB–YAP Regulation to Promote Colon Cancer. Clin. Cancer Res. 2018, 24, 2015–2025. [Google Scholar] [CrossRef]
- Yao, L.; Zhou, L.; Xuan, Y.; Zhang, P.; Wang, X.; Wang, T.; Meng, T.; Xue, Y.; Ma, X.; Shah, A.S.; et al. The Proteasome Activator REGγ Counteracts Expression and Autoimmunity. J. Autoimmun. 2019, 103, 102282. [Google Scholar] [CrossRef]
- Liu, S.; Zheng, L.-L.; Zhu, Y.-M.; Shen, H.-J.; Zhong, Q.; Huang, J.; Li, C.; Liu, Z.; Yao, M.-D.; Ou, R.-M.; et al. Knockdown of REGγ Inhibits the Proliferation and migration and Promotes the Apoptosis of Multiple Myeloma Cells by downregulating NF-ΚB Signal Pathway. Hematology 2018, 23, 277–283. [Google Scholar] [CrossRef]
- Jiao, C.; Li, L.; Zhang, P.; Zhang, L.; Li, K.; Fang, R.; Yuan, L.; Shi, K.; Pan, L.; Guo, Q.; et al. REGγ Ablation Impedes Dedifferentiation of Anaplastic Carcinoma and Accentuates Radio-Therapeutic Response by regulating the Smad7-TGF-β Pathway. Cell Death Differ. 2020, 27, 497–508. [Google Scholar] [CrossRef]
- Tong, L.; Shen, S.; Huang, Q.; Fu, J.; Wang, T.; Pan, L.; Zhang, P.; Chen, G.; Huang, T.; Li, K.; et al. Proteasome-Dependent Degradation of Smad7 Is Critical for Lung Metastasis. Cell Death Differ. 2020, 27, 1795–1806. [Google Scholar] [CrossRef]
- Uchimura, Y.; Barton, L.F.; Rada, C.; Neuberger, M.S. REG-γ Associates with and Modulates the Abundance of nuclear Activation-Induced Deaminase. J. Exp. Med. 2011, 208, 2385–2391. [Google Scholar] [CrossRef]
- Zhou, L.; Yao, L.; Zhang, Q.; Xie, W.; Wang, X.; Zhang, H.; Xu, J.; Lin, Q.; Li, Q.; Xuan, Y.; et al. REGγ Controls Th17 Cell Differentiation and Autoimmune by Regulating Dendritic Cells. Cell. Mol. Immunol. 2020, 17, 1136–1147. [Google Scholar] [CrossRef]
- Fan, J.; Liu, L.; Liu, Q.; Cui, Y.; Yao, B.; Zhang, M.; Gao, Y.; Fu, Y.; Dai, H.; Pan, J.; et al. CKIP-1 Limits Foam Cell Formation and Inhibits atherosclerosis by Promoting Degradation of Oct-1 by REGγ. Nat. Commun. 2019, 10, 425. [Google Scholar] [CrossRef]
- Dong, S.; Jia, C.; Zhang, S.; Fan, G.; Li, Y.; Shan, P.; Sun, L.; Xiao, W.; Li, L.; Zheng, Y.; et al. The REGγ Proteasome Regulates Hepatic Lipid metabolism through Inhibition of Autophagy. Cell Metab. 2013, 18, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Fan, G.; Shan, P.; Qiu, X.; Dong, S.; Liao, L.; Yu, C.; Wang, T.; Gu, X.; Li, Q.; et al. Regulation of Energy Homeostasis by the Ubiquitin-Independent Proteasome. Nat. Commun. 2016, 7, 12497. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Lai, L.; Zuo, Q.; Dai, F.; Wu, L.; Wang, Y.; Zhou, Q.; Liu, J.; Liu, J.; Li, L.; et al. PKA Turnover by the REGγ-Proteasome Modulates FoxO1 Activity and VEGF-Induced Angiogenesis. J. Mol. Cell. Cardiol. 2014, 72, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Meng, B.; Dong, H.; Jing, T.; Wu, N.; Yang, Y.; Huang, L.; Moses, R.E.; O’Malley, B.W.; Mei, B.; et al. Upregulation of GSK3β Contributes to Brain Disorders in elderly REGγ-Knockout Mice. Neuropsychopharmacology 2016, 41, 1340–1349. [Google Scholar] [CrossRef]
- Murata, S.; Kawahara, H.; Tohma, S.; Yamamoto, K.; Kasahara, M.; Nabeshima, Y.; Tanaka, K.; Chiba, T. Growth Retardation in Mice Lacking the Proteasome Activator. J. Biol. Chem. 1999, 274, 38211–38215. [Google Scholar] [CrossRef]
- Barton, L.F.; Runnels, H.A.; Schell, T.D.; Cho, Y.; Gibbons, R.; Tevethia, S.S.; Deepe Jr, G.S.; Monaco, J.J. Immune Defects in 28-KDa Proteasome Activator Gamma-Deficient. J. Immunol. 2004, 172, 3948–3954. [Google Scholar] [CrossRef]
- Masson, P.; Lundgren, J.; Young, P. Drosophila Proteasome Regulator REGgamma: Transcriptional by DNA Replication-Related Factor DREF and evidence for a Role in Cell Cycle Progression. J. Mol. Biol. 2003, 327, 1001–1012. [Google Scholar] [CrossRef]
- He, J.; Cui, L.; Zeng, Y.; Wang, G.; Zhou, P.; Yang, Y.; Ji, L.; Zhao, Y.; Chen, J.; Wang, Z.; et al. REGγ Is Associated with Multiple Oncogenic Pathways in human Cancers. BMC Cancer 2012, 12, 75. [Google Scholar] [CrossRef]
- He, X.; Li, Y.; Chen, Q.; Zheng, L.; Lou, J.; Lin, C.; Gong, J.; Zhu, Y.; Wu, Y. O-GlcNAcylation and Stablization of SIRT7 Promote Pancreatic Progression by Blocking the SIRT7-REGγ. Cell Death Differ. 2022. [Google Scholar] [CrossRef]
- Chen, H.; Gao, X.; Sun, Z.; Wang, Q.; Zuo, D.; Pan, L.; Li, K.; Chen, J.; Chen, G.; Hu, K.; et al. REGγ Accelerates Melanoma Formation by Regulating/β-Catenin Signalling Pathway. Exp. Dermatol. 2017, 26, 1118–1124. [Google Scholar] [CrossRef]
- Li, L.; Dang, Y.; Zhang, J.; Yan, W.; Zhai, W.; Chen, H.; Li, K.; Tong, L.; Gao, X.; Amjad, A.; et al. REGγ Is Critical for Skin Carcinogenesis by modulating the Wnt/β-Catenin Pathway. Nat. Commun. 2015, 6, 6875. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yang, M.; Lin, Z.; Mael, S.K.; Li, Y.; Zhang, L.; Kong, Y.; Zhang, Y.; Ren, Y.; Li, J.; et al. REGγ Drives Lgr5+ Stem Cells to Potentiate Radiation Intestinal Regeneration. Sci. China Life Sci. 2021. [Google Scholar] [CrossRef] [PubMed]
- Rigoulet, M.; Bouchez, C.L.; Paumard, P.; Ransac, S.; Cuvellier, S.; Duvezin-Caubet, S.; Mazat, J.P.; Devin, A. Cell Energy Metabolism: An Update. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148276. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Xuan, Y.; Zhang, H.; Yang, B.; Ma, X.; Wang, T.; Meng, T.; Sun, W.; Wei, H.; Ma, X.; et al. Reciprocal REGγ-MTORC1 Regulation Promotes Glycolytic in Hepatocellular Carcinoma. Oncogene 2021, 40, 677–692. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Haratake, K.; Miyahara, H.; Chiba, T. Proteasome Activators, PA28γ and PA200, Play Roles in Male Fertility. Sci. Rep. 2016, 6, 23171. [Google Scholar] [CrossRef]
- Clotaire, D.Z.J.; Wei, Y.; Yu, X.; Ousman, T.; Hua, J. Functions of Promyelocytic Leukaemia Zinc Finger (Plzf) in Male Stem Cell Development and Differentiation. Reprod. Fertil. Dev. 2019, 31, 1315. [Google Scholar] [CrossRef]
- Gao, X.; Chen, H.; Liu, J.; Shen, S.; Wang, Q.; Clement, T.M.; Deskin, B.J.; Chen, C.; Zhao, D.; Wang, L.; et al. The REGγ-Proteasome Regulates Spermatogenesis partially by P53-PLZF Signaling. Stem Cell Rep. 2019, 13, 559–571. [Google Scholar] [CrossRef]
- Magni, M.; Ruscica, V.; Buscemi, G.; Kim, J.-E.; Nachimuthu, B.T.; Fontanella, E.; Delia, D.; Zannini, L. Chk2 and REGγ-Dependent DBC1 Regulation in DNA Induced Apoptosis. Nucleic Acids Res. 2014, 42, 13150–13160. [Google Scholar] [CrossRef]
- Shi, Y.; Luo, X.; Li, P.; Tan, J.; Wang, X.; Xiang, T.; Ren, G. MiR-7-5p Suppresses Cell Proliferation and Induces Apoptosis of breast Cancer Cells Mainly by Targeting REGγ. Cancer Lett. 2015, 358, 27–36. [Google Scholar] [CrossRef]
- Shu, Y.; Long, J.; Guo, W.; Ye, W. MicroRNA-195-5p Inhibitor Prevents the Development of osteoarthritis by Targeting REGγ. Mol. Med. Rep. 2019, 19, 4561–4568. [Google Scholar] [CrossRef]
- Liu, J.; Yu, G.; Zhao, Y.; Zhao, D.; Wang, Y.; Wang, L.; Liu, J.; Li, L.; Zeng, Y.; Dang, Y.; et al. REGgamma Modulates P53 Activity by Regulating Its Cellular. J. Cell Sci. 2010, 123, 4076–4084. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How Does P53 Induce Apoptosis and How Does This Relate To-Mediated Tumour Suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Moncsek, A.; Gruner, M.; Meyer, H.; Lehmann, A.; Kloetzel, P.-M.; Stohwasser, R. Evidence for Anti-Apoptotic Roles of Proteasome Activator via Inhibiting Caspase Activity. Apoptosis 2015, 20, 1211–1228. [Google Scholar] [CrossRef] [PubMed]
- Uriarte, M.; Sen Nkwe, N.; Tremblay, R.; Ahmed, O.; Messmer, C.; Mashtalir, N.; Barbour, H.; Masclef, L.; Voide, M.; Viallard, C.; et al. Starvation-Induced Proteasome Assemblies in the Nucleus Link Acid Supply to Apoptosis. Nat. Commun. 2021, 12, 6984. [Google Scholar] [CrossRef] [PubMed]
- Naumova, O.Y.; Palejev, D.; Vlasova, N.V.; Lee, M.; Rychkov, S.Y.; Babich, O.N.; Vaccarino, F.M.; Grigorenko, E.L. Age-Related Changes of Gene Expression in the Neocortex: Preliminary Data on RNA-Seq of the Transcriptome in Three Distinct Cortical Areas. Dev. Psychopathol. 2012, 24, 1427–1442. [Google Scholar] [CrossRef]
- Satoh, J.-I.; Yamamoto, Y.; Asahina, N.; Kitano, S.; Kino, Y. RNA-Seq Data Mining: Downregulation of NeuroD6 Serves as a possible Biomarker for Alzheimer’s Disease Brains. Dis. Markers 2014, 2014, 123165. [Google Scholar] [CrossRef]
- Seo, H.; Sonntag, K.-C.; Kim, W.; Cattaneo, E.; Isacson, O. Proteasome Activator Enhances Survival of Huntington’s Disease Model Cells. PLoS ONE 2007, 2, e238. [Google Scholar] [CrossRef]
- Jeon, J.; Kim, W.; Jang, J.; Isacson, O.; Seo, H. Gene Therapy by Proteasome Activator, PA28γ, Improves Coordination and Proteasome Function in Huntington’s YAC128 Mice. Neuroscience 2016, 324, 20–28. [Google Scholar] [CrossRef]
- Levy-Barda, A.; Lerenthal, Y.; Davis, A.J.; Chung, Y.M.; Essers, J.; Shao, Z.; van Vliet, N.; Chen, D.J.; Hu, M.C.-T.; Kanaar, R.; et al. Involvement of the Nuclear Proteasome Activator PA28γ in the Cellular Response to DNA Double-Strand Breaks. Cell Cycle 2011, 10, 4300–4310. [Google Scholar] [CrossRef]
- Liedtke, V.; Schröder, C.; Roggenbuck, D.; Weiss, R.; Stohwasser, R.; Schierack, P.; Rödiger, S.; Schenk, L. LEDGF/P75 Is Required for an Efficient DNA Damage Response. Int. J. Mol. Sci. 2021, 22, 5866. [Google Scholar] [CrossRef]
- Ali, A.; Wang, Z.; Fu, J.; Ji, L.; Liu, J.; Li, L.; Wang, H.; Chen, J.; Caulin, C.; Myers, J.N.; et al. Differential Regulation of the REGγ-Proteasome pathway by P53/TGF-β Signalling and Mutant P53 in Cancer Cells. Nat. Commun. 2013, 4, 2667. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bao, W.; Jiang, F.; Che, Q.; Chen, Z.; Wang, F.; Tong, H.; Dai, C.; He, X.; Liao, Y.; et al. Mutant P53 (P53-R248Q) Functions as an Oncogene in Promoting Cancer by up-Regulating REGγ. Cancer Lett. 2015, 360, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yang, X.; Wang, X.; Gu, J.; Zhou, D.; Wang, Y.; Yin, B.; Guo, J.; Zhou, M. Silencing CDR1as Enhances the Sensitivity of Breast Cancer to Drug Resistance by Acting as a MiR-7 Sponge to down-Regulate REGγ. J. Cell. Mol. Med. 2019, 23, 4921–4932. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, M.; Xue, Y. Downregulation of CircRNA CDR1as Specifically Triggered-Dose Diosbulbin-B Induced Gastric Cancer Cell Death by regulating MiR-7-5p/REGγ Axis. Biomed. Pharmacother. 2019, 120, 109462. [Google Scholar] [CrossRef]
- Gao, X.; Wang, Q.; Wang, Y.; Liu, J.; Liu, S.; Liu, J.; Zhou, X.; Zhou, L.; Chen, H.; Pan, L.; et al. The REGγ Inhibitor NIP30 Increases Sensitivity to chemotherapy in P53-Deficient Tumor Cells. Nat. Commun. 2020, 11, 3904. [Google Scholar] [CrossRef]
- Yu, Z.; Wei, X.; Liu, L.; Sun, H.; Fang, T.; Wang, L.; Li, Y.; Sui, W.; Wang, K.; He, Y.; et al. Indirubin-3′-Monoxime Acts as Proteasome Inhibitor: Therapeutic in Multiple Myeloma. EBioMedicine 2022, 78, 103950. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, L.; Zhou, P.; Wang, G.; Zeng, Y.; Wang, Y.; Liu, J.; Zhang, B.; Liu, S.; Luo, H.; et al. Regulation of REGγ Cellular Distribution and function by SUMO Modification. Cell Res. 2011, 21, 807–816. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, S.; Zuo, Q.; Wu, L.; Ji, L.; Zhai, W.; Xiao, J.; Chen, J.; Li, X. Oxidative Challenge Enhances REGγ-Proteasome-Dependent Degradation. Free Radic. Biol. Med. 2015, 82, 42–49. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Funderburk, K.E.; Kang, J.; Li, H.J. Regulation of Life & Death by REGγ. Cells 2022, 11, 2281. https://doi.org/10.3390/cells11152281
Funderburk KE, Kang J, Li HJ. Regulation of Life & Death by REGγ. Cells. 2022; 11(15):2281. https://doi.org/10.3390/cells11152281
Chicago/Turabian StyleFunderburk, Keaton E., Jungseog Kang, and Henry J. Li. 2022. "Regulation of Life & Death by REGγ" Cells 11, no. 15: 2281. https://doi.org/10.3390/cells11152281
APA StyleFunderburk, K. E., Kang, J., & Li, H. J. (2022). Regulation of Life & Death by REGγ. Cells, 11(15), 2281. https://doi.org/10.3390/cells11152281