
Sex Steroids Effects on Asthma: A Network Perspective of Immune and Airway Cells


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
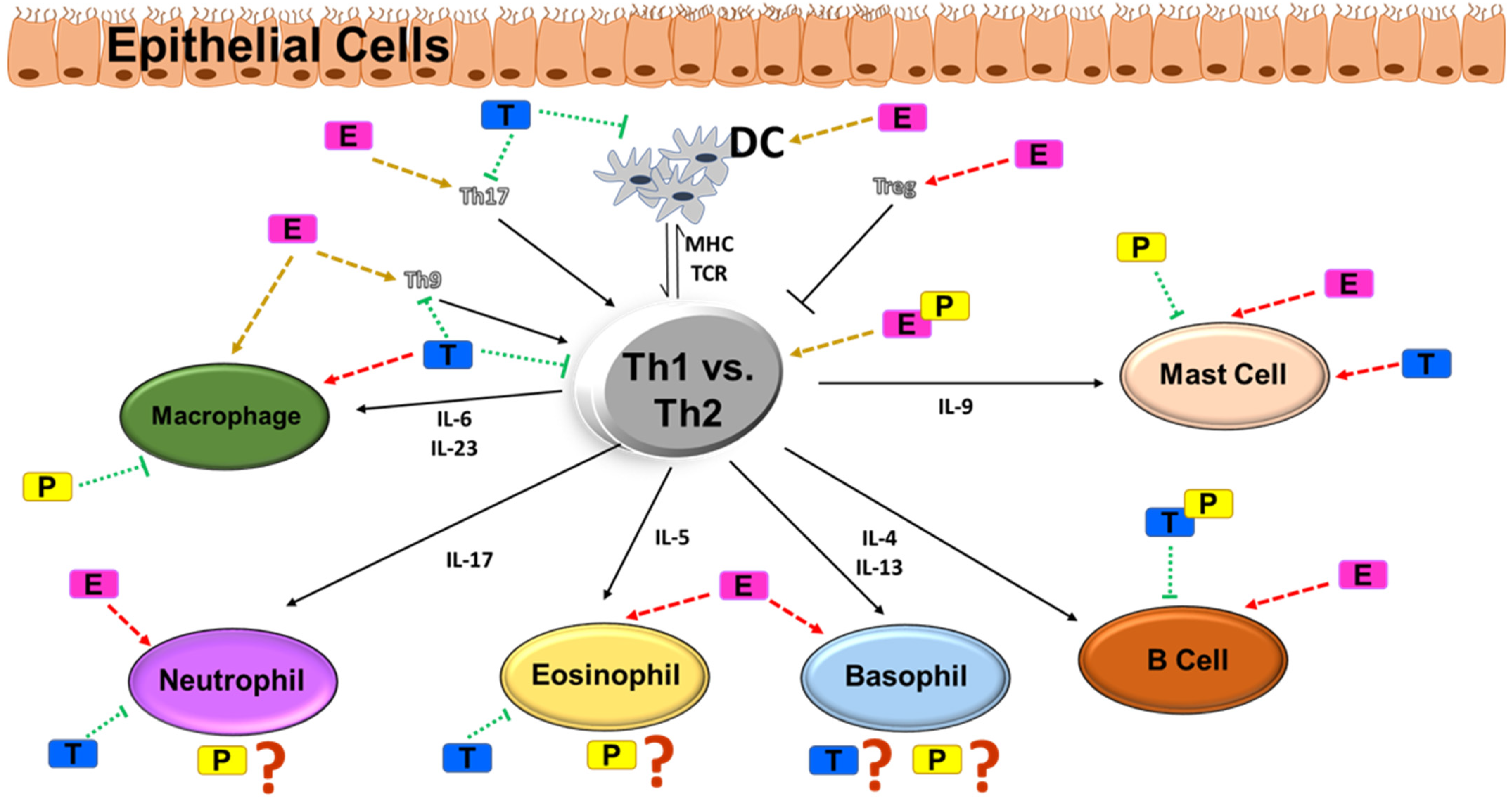
2. Airway Inflammation in Asthma
3. Sex Steroids and Immune Function
4. Biological Effects of Sex Steroids: Immune Cell Milieu in Asthma
4.1. Sex Steroids and Basophils
4.2. Sex Steroids and Eosinophils
4.3. Sex Steroids and Neutrophils
4.4. Sex Steroids and B Cells
4.5. Sex Steroids and T Cells
4.6. Sex Steroids and Monocytes/Macrophages/Dendritic Cells (Mononuclear Phagocyte System)
4.7. Sex Steroids and Natural Killer Cells
4.8. Sex Steroids and Mast Cells
4.9. Treg: A Window of Opportunity?
5. Sex Steroids and Structural Cells of the Airway
6. Coronavirus Disease: What’s Sex/Sex Steroids Got to Do with It?
7. Conclusions and Future Scope
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prevention of Asthma, Centers for Disease Control and Prevention. Most Recent National Asthma Data. Available online: https://www.cdc.gov/asthma/most_recent_national_asthma_data.htm (accessed on 15 May 2022).
- Borkar, N.A.; Sathish, V. Sex steroids and their influence in Lung Diseases across the lifespan. In Sex-Based Differences in Lung Physiology; Silveyra, P., Tigno, X.T., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Ward, C.; Pais, M.; Bish, R.; Reid, D.; Feltis, B.; Johns, D.; Walters, E.H. Airway inflammation, basement membrane thickening and bronchial hyperresponsiveness in asthma. Thorax 2002, 57, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Benayoun, L.; Druilhe, A.; Dombret, M.C.; Aubier, M.; Pretolani, M. Airway structural alterations selectively associated with severe asthma. Am. J. Respir Crit. Care Med. 2003, 167, 1360–1368. [Google Scholar] [CrossRef] [PubMed]
- Prakash, Y.S. Airway smooth muscle in airway reactivity and remodeling: What have we learned? Am. J. Physiol. Lung Cell Mol. Physiol. 2013, 305, L912–L933. [Google Scholar] [CrossRef] [PubMed]
- Prakash, Y.S. Emerging concepts in smooth muscle contributions to airway structure and function: Implications for health and disease. Am. J. Physiol. Lung Cell Mol. Physiol. 2016, 311, L1113–L1140. [Google Scholar] [CrossRef] [PubMed]
- An, S.S.; Bai, T.R.; Bates, J.H.; Black, J.L.; Brown, R.H.; Brusasco, V.; Chitano, P.; Deng, L.; Dowell, M.; Eidelman, D.H.; et al. Airway smooth muscle dynamics: A common pathway of airway obstruction in asthma. Eur. Respir. J. 2007, 29, 834–860. [Google Scholar] [CrossRef]
- Larche, M.; Robinson, D.S.; Kay, A.B. The role of T lymphocytes in the pathogenesis of asthma. J. Allergy Clin. Immunol. 2003, 111, 450–463. [Google Scholar] [CrossRef]
- Robinson, D.S. The role of the T cell in asthma. J. Allergy Clin. Immunol. 2010, 126, 1081–1091. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Dao Nguyen, X.; Robinson, D.S. Fluticasone propionate increases CD4CD25 T regulatory cell suppression of allergen-stimulated CD4CD25 T cells by an IL-10-dependent mechanism. J. Allergy Clin. Immunol. 2004, 114, 296–301. [Google Scholar] [CrossRef]
- Alexander, A.G.; Barnes, N.C.; Kay, A.B. Trial of cyclosporin in corticosteroid-dependent chronic severe asthma. Lancet 1992, 339, 324–328. [Google Scholar] [CrossRef]
- Reyes-Garcia, J.; Montano, L.M.; Carbajal-Garcia, A.; Wang, Y.X. Sex Hormones and Lung Inflammation. Adv. Exp. Med. Biol 2021, 1304, 259–321. [Google Scholar] [CrossRef] [PubMed]
- Yung, J.A.; Fuseini, H.; Newcomb, D.C. Hormones, sex, and asthma. Ann. Allergy Asthma Immunol. 2018, 120, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Laffont, S.; Blanquart, E.; Guery, J.C. Sex Differences in Asthma: A Key Role of Androgen-Signaling in Group 2 Innate Lymphoid Cells. Front. Immunol. 2017, 8, 1069. [Google Scholar] [CrossRef] [PubMed]
- Foo, Y.Z.; Nakagawa, S.; Rhodes, G.; Simmons, L.W. The effects of sex hormones on immune function: A meta-analysis. Biol. Rev. Camb. Philos. Soc. 2017, 92, 551–571. [Google Scholar] [CrossRef] [PubMed]
- Hellings, P.W.; Vandekerckhove, P.; Claeys, R.; Billen, J.; Kasran, A.; Ceuppens, J.L. Progesterone increases airway eosinophilia and hyper-responsiveness in a murine model of allergic asthma. Clin. Exp. Allergy 2003, 33, 1457–1463. [Google Scholar] [CrossRef] [PubMed]
- Kadel, S.; Ainsua-Enrich, E.; Hatipoglu, I.; Turner, S.; Singh, S.; Khan, S.; Kovats, S. A Major Population of Functional KLRG1(-) ILC2s in Female Lungs Contributes to a Sex Bias in ILC2 Numbers. Immunohorizons 2018, 2, 74–86. [Google Scholar] [CrossRef]
- Vermillion, M.S.; Ursin, R.L.; Attreed, S.E.; Klein, S.L. Estriol Reduces Pulmonary Immune Cell Recruitment and Inflammation to Protect Female Mice From Severe Influenza. Endocrinology 2018, 159, 3306–3320. [Google Scholar] [CrossRef]
- Kalidhindi, R.S.R.; Ambhore, N.S.; Balraj, P.; Schmidt, T.; Khan, M.N.; Sathish, V. Androgen receptor activation alleviates airway hyperresponsiveness, inflammation, and remodeling in a murine model of asthma. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 320, L803–L818. [Google Scholar] [CrossRef]
- Borkar, N.A.; Douri, D.; Pabelick, C.M.; Prakash, Y.; Sathish, V. Estrogen Influences Kisspeptins Expression in Human Airway Smooth Muscle Cells. Am. J. Respir. Crit. Care Med. 2021, 203, A4480. [Google Scholar] [CrossRef]
- Balraj, P.; Ambhore, N.S.; Borkar, N.A.; Pabelick, C.M.; Prakash, Y.; Sathish, V. Kisspeptin Attenuates Airway Smooth Muscle Cell Migration by Regulating Rho GTPase Signaling Pathway. Am. J. Respir. Crit. Care Med. 2022, 205, A1229. [Google Scholar] [CrossRef]
- Borkar, N.A.; Kalidhindi, R.S.R.; Ambhore, N.S.; Sathish, V. Role of Kisspeptins in Airway Hyperresponsiveness and Remodeling in a Mouse Model of Allergic Asthma. Am. J. Respir. Crit. Care Med. 2020, 201, A5617. [Google Scholar] [CrossRef]
- Borkar, N.A.; Ambhore, N.S.; Sathish, V. Kisspeptin Reduces Airway Hyperreactivity and Remodeling in Asthmatic Ovariectomized Mice. FASEB J. 2022, 36. [Google Scholar] [CrossRef]
- WHO Organization. Asthma. Available online: https://www.who.int/news-room/fact-sheets/detail/asthma (accessed on 15 May 2022).
- Trejo Bittar, H.E.; Yousem, S.A.; Wenzel, S.E. Pathobiology of severe asthma. Annu. Rev. Pathol. 2015, 10, 511–545. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef]
- Han, M.K.; Arteaga-Solis, E.; Blenis, J.; Bourjeily, G.; Clegg, D.J.; DeMeo, D.; Duffy, J.; Gaston, B.; Heller, N.M.; Hemnes, A.; et al. Female Sex and Gender in Lung/Sleep Health and Disease. Increased Understanding of Basic Biological, Pathophysiological, and Behavioral Mechanisms Leading to Better Health for Female Patients with Lung Disease. Am. J. Respir. Crit. Care Med. 2018, 198, 850–858. [Google Scholar] [CrossRef]
- Payne, D.N.; Rogers, A.V.; Adelroth, E.; Bandi, V.; Guntupalli, K.K.; Bush, A.; Jeffery, P.K. Early thickening of the reticular basement membrane in children with difficult asthma. Am. J. Respir. Crit. Care Med. 2003, 167, 78–82. [Google Scholar] [CrossRef]
- Holgate, S.T. Innate and adaptive immune responses in asthma. Nat. Med. 2012, 18, 673–683. [Google Scholar] [CrossRef]
- Boonpiyathad, T.; Sozener, Z.C.; Satitsuksanoa, P.; Akdis, C.A. Immunologic mechanisms in asthma. Semin. Immunol. 2019, 46, 101333. [Google Scholar] [CrossRef]
- Yao, Y.E.; Qin, C.C.; Yang, C.M.; Huang, T.X. γδT17/γδTreg cell subsets: A new paradigm for asthma treatment. J. Asthma 2021, 1–11. [Google Scholar] [CrossRef]
- Munitz, A.; Foster, P.S. T(H)9 cells: In front and beyond T(H)2. J. Allergy Clin. Immunol. 2012, 129, 1011–1013. [Google Scholar] [CrossRef]
- Pelaia, G.; Vatrella, A.; Busceti, M.T.; Gallelli, L.; Calabrese, C.; Terracciano, R.; Maselli, R. Cellular mechanisms underlying eosinophilic and neutrophilic airway inflammation in asthma. Mediat. Inflamm. 2015, 2015, 879783. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Kolls, J.K. Neutrophilic Inflammation in Asthma and Association with Disease Severity. Trends Immunol. 2017, 38, 942–954. [Google Scholar] [CrossRef]
- Brusselle, G.G.; Maes, T.; Bracke, K.R. Eosinophils in the spotlight: Eosinophilic airway inflammation in nonallergic asthma. Nat. Med. 2013, 19, 977–979. [Google Scholar] [CrossRef] [PubMed]
- Bouman, A.; Heineman, M.J.; Faas, M.M. Sex hormones and the immune response in humans. Hum. Reprod Update 2005, 11, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Zein, J.G.; Denson, J.L.; Wechsler, M.E. Asthma over the Adult Life Course: Gender and Hormonal Influences. Clin. Chest Med. 2019, 40, 149–161. [Google Scholar] [CrossRef]
- Borkar, N.A.; Roos, B.; Prakash, Y.S.; Sathish, V.; Pabelick, C.M. Nicotinic alpha7 acetylcholine receptor (alpha7nAChR) in human airway smooth muscle. Arch. Biochem. Biophys. 2021, 706, 108897. [Google Scholar] [CrossRef] [PubMed]
- Pawankar, R.; Hayashi, M.; Yamanishi, S.; Igarashi, T. The paradigm of cytokine networks in allergic airway inflammation. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 41–48. [Google Scholar] [CrossRef]
- Kuruvilla, M.E.; Lee, F.E.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef]
- Peters, M.C.; Wenzel, S.E. Intersection of biology and therapeutics: Type 2 targeted therapeutics for adult asthma. Lancet 2020, 395, 371–383. [Google Scholar] [CrossRef]
- Pavord, I.; Bahmer, T.; Braido, F.; Cosio, B.G.; Humbert, M.; Idzko, M.; Adamek, L. Severe T2-high asthma in the biologics era: European experts’ opinion. Eur Respir Rev. 2019, 28, 190054. [Google Scholar] [CrossRef]
- Kyriakopoulos, C.; Gogali, A.; Bartziokas, K.; Kostikas, K. Identification and treatment of T2-low asthma in the era of biologics. ERJ Open Res. 2021, 7, 00309-2020. [Google Scholar] [CrossRef]
- Svenningsen, S.; Nair, P. Asthma Endotypes and an Overview of Targeted Therapy for Asthma. Front. Med. (Lausanne) 2017, 4, 158. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.G.; Simpson, J.L.; Saltos, N. Heterogeneity of airway inflammation in persistent asthma: Evidence of neutrophilic inflammation and increased sputum interleukin-8. Chest 2001, 119, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Sze, E.; Bhalla, A.; Nair, P. Mechanisms and therapeutic strategies for non-T2 asthma. Allergy 2020, 75, 311–325. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, B.G.; Maarsingh, H.; Meurs, H.; Gosens, R. Airway structural components drive airway smooth muscle remodeling in asthma. Proc. Am. Thorac Soc. 2009, 6, 683–692. [Google Scholar] [CrossRef]
- Salter, B.; Pray, C.; Radford, K.; Martin, J.G.; Nair, P. Regulation of human airway smooth muscle cell migration and relevance to asthma. Respir Res. 2017, 18, 156. [Google Scholar] [CrossRef]
- Johnson, P.R.; Burgess, J.K. Airway smooth muscle and fibroblasts in the pathogenesis of asthma. Curr. Allergy Asthma Rep. 2004, 4, 102–108. [Google Scholar] [CrossRef]
- Bourdin, A.; Neveu, D.; Vachier, I.; Paganin, F.; Godard, P.; Chanez, P. Specificity of basement membrane thickening in severe asthma. J. Allergy Clin. Immunol. 2007, 119, 1367–1374. [Google Scholar] [CrossRef]
- Hamid, Q.; Tulic, M. Immunobiology of asthma. Annu. Rev. Physiol. 2009, 71, 489–507. [Google Scholar] [CrossRef]
- Gruenberg, D.; Busse, W. Biologic therapies for asthma. Curr. Opin. Pulm. Med. 2010, 16, 19–24. [Google Scholar] [CrossRef]
- Cabello, N.; Mishra, V.; Sinha, U.; DiAngelo, S.L.; Chroneos, Z.C.; Ekpa, N.A.; Cooper, T.K.; Caruso, C.R.; Silveyra, P. Sex differences in the expression of lung inflammatory mediators in response to ozone. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 309, L1150–L1163. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Mempel, M.; Schober, W.; Behrendt, H.; Ring, J. Gender difference, sex hormones, and immediate type hypersensitivity reactions. Allergy 2008, 63, 1418–1427. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, S.; Berthenet, K.; Garlanda, C. Sexual Dimorphism in Innate Immunity. Clin. Rev. Allergy Immunol. 2019, 56, 308–321. [Google Scholar] [CrossRef]
- Cooper, M.D.; Alder, M.N. The evolution of adaptive immune systems. Cell 2006, 124, 815–822. [Google Scholar] [CrossRef]
- Tesmer, L.A.; Lundy, S.K.; Sarkar, S.; Fox, D.A. Th17 cells in human disease. Immunol. Rev. 2008, 223, 87–113. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Lee, H.K. The Role of CD4(+) T Cells and Microbiota in the Pathogenesis of Asthma. Int J. Mol. Sci. 2021, 22, 11822. [Google Scholar] [CrossRef]
- Khan, M.A. Regulatory T cells mediated immunomodulation during asthma: A therapeutic standpoint. J. Transl. Med. 2020, 18, 456. [Google Scholar] [CrossRef]
- Chiarella, S.E.; Cardet, J.C.; Prakash, Y.S. Sex, Cells, and Asthma. Mayo Clin. Proc. 2021, 96, 1955–1969. [Google Scholar] [CrossRef]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef]
- Becerra-Diaz, M.; Song, M.; Heller, N. Androgen and Androgen Receptors as Regulators of Monocyte and Macrophage Biology in the Healthy and Diseased Lung. Front. Immunol. 2020, 11, 1698. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Sathish, V.; Martin, Y.N.; Prakash, Y.S. Sex steroid signaling: Implications for lung diseases. Pharmacol. Ther. 2015, 150, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Ngo, S.T.; Steyn, F.J.; McCombe, P.A. Gender differences in autoimmune disease. Front. Neuroendocr. 2014, 35, 347–369. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, A.; Sekhon, H.K.; Kaur, G. Sex hormones and immune dimorphism. Sci. World J. 2014, 2014, 159150. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Klein, R.S. Sex Drives Dimorphic Immune Responses to Viral Infections. J. Immunol. 2017, 198, 1782–1790. [Google Scholar] [CrossRef]
- Keselman, A.; Fang, X.; White, P.B.; Heller, N.M. Estrogen Signaling Contributes to Sex Differences in Macrophage Polarization during Asthma. J. Immunol. 2017, 199, 1573–1583. [Google Scholar] [CrossRef]
- Kovats, S. Estrogen receptors regulate innate immune cells and signaling pathways. Cell. Immunol. 2015, 294, 63–69. [Google Scholar] [CrossRef]
- Taneja, V. Sex Hormones Determine Immune Response. Front. Immunol. 2018, 9, 1931. [Google Scholar] [CrossRef]
- Namazi, M.R. The Th1-promoting effects of dehydroepiandrosterone can provide an explanation for the stronger Th1-immune response of women. Iran. J. Allergy Asthma Immunol. 2009, 8, 65–69. [Google Scholar]
- AbdulHussain, G.; Azizieh, F.; Makhseed, M.; Raghupathy, R. Effects of Progesterone, Dydrogesterone and Estrogen on the Production of Th1/Th2/Th17 Cytokines by Lymphocytes from Women with Recurrent Spontaneous Miscarriage. J. Reprod. Immunol. 2020, 140, 103132. [Google Scholar] [CrossRef]
- Piccinni, M.P.; Raghupathy, R.; Saito, S.; Szekeres-Bartho, J. Cytokines, Hormones and Cellular Regulatory Mechanisms Favoring Successful Reproduction. Front. Immunol. 2021, 12, 717808. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef] [PubMed]
- Heldring, N.; Pike, A.; Andersson, S.; Matthews, J.; Cheng, G.; Hartman, J.; Tujague, M.; Strom, A.; Treuter, E.; Warner, M.; et al. Estrogen receptors: How do they signal and what are their targets. Physiol. Rev. 2007, 87, 905–931. [Google Scholar] [CrossRef]
- Bonds, R.S.; Midoro-Horiuti, T. Estrogen effects in allergy and asthma. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 92–99. [Google Scholar] [CrossRef]
- Dosiou, C.; Hamilton, A.E.; Pang, Y.; Overgaard, M.T.; Tulac, S.; Dong, J.; Thomas, P.; Giudice, L.C. Expression of membrane progesterone receptors on human T lymphocytes and Jurkat cells and activation of G-proteins by progesterone. J. Endocrinol. 2008, 196, 67–77. [Google Scholar] [CrossRef]
- Zhao, X.J.; McKerr, G.; Dong, Z.; Higgins, C.A.; Carson, J.; Yang, Z.Q.; Hannigan, B.M. Expression of oestrogen and progesterone receptors by mast cells alone, but not lymphocytes, macrophages or other immune cells in human upper airways. Thorax 2001, 56, 205–211. [Google Scholar] [CrossRef]
- Khan, K.N.; Masuzaki, H.; Fujishita, A.; Kitajima, M.; Sekine, I.; Matsuyama, T.; Ishimaru, T. Estrogen and progesterone receptor expression in macrophages and regulation of hepatocyte growth factor by ovarian steroids in women with endometriosis. Hum. Reprod. 2005, 20, 2004–2013. [Google Scholar] [CrossRef] [PubMed]
- Teilmann, S.C.; Clement, C.A.; Thorup, J.; Byskov, A.G.; Christensen, S.T. Expression and localization of the progesterone receptor in mouse and human reproductive organs. J. Endocrinol. 2006, 191, 525–535. [Google Scholar] [CrossRef]
- Migliaccio, A.; Piccolo, D.; Castoria, G.; Di Domenico, M.; Bilancio, A.; Lombardi, M.; Gong, W.; Beato, M.; Auricchio, F. Activation of the Src/p21ras/Erk pathway by progesterone receptor via cross-talk with estrogen receptor. EMBO J. 1998, 17, 2008–2018. [Google Scholar] [CrossRef]
- Becerra-Diaz, M.; Strickland, A.B.; Keselman, A.; Heller, N.M. Androgen and Androgen Receptor as Enhancers of M2 Macrophage Polarization in Allergic Lung Inflammation. J. Immunol. 2018, 201, 2923–2933. [Google Scholar] [CrossRef]
- Siracusa, M.C.; Kim, B.S.; Spergel, J.M.; Artis, D. Basophils and allergic inflammation. J. Allergy Clin. Immunol. 2013, 132, 789–801, quiz 788. [Google Scholar] [CrossRef] [PubMed]
- Cocchiara, R.; Albeggiani, G.; Di Trapani, G.; Azzolina, A.; Lampiasi, N.; Rizzo, F.; Geraci, D. Modulation of rat peritoneal mast cell and human basophil histamine release by estrogens. Int. Arch. Allergy Appl. Immunol. 1990, 93, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Hall, O.J.; Limjunyawong, N.; Vermillion, M.S.; Robinson, D.P.; Wohlgemuth, N.; Pekosz, A.; Mitzner, W.; Klein, S.L. Progesterone-Based Therapy Protects Against Influenza by Promoting Lung Repair and Recovery in Females. PLoS Pathog. 2016, 12, e1005840. [Google Scholar] [CrossRef]
- Gagliano-Juca, T.; Pencina, K.M.; Guo, W.; Li, Z.; Huang, G.; Basaria, S.; Bhasin, S. Differential effects of testosterone on circulating neutrophils, monocytes, and platelets in men: Findings from two trials. Andrology 2020, 8, 1324–1331. [Google Scholar] [CrossRef] [PubMed]
- Humbles, A.A.; Lloyd, C.M.; McMillan, S.J.; Friend, D.S.; Xanthou, G.; McKenna, E.E.; Ghiran, S.; Gerard, N.P.; Yu, C.; Orkin, S.H.; et al. A critical role for eosinophils in allergic airways remodeling. Science 2004, 305, 1776–1779. [Google Scholar] [CrossRef]
- Nakagome, K.; Nagata, M. Involvement and Possible Role of Eosinophils in Asthma Exacerbation. Front. Immunol. 2018, 9, 2220. [Google Scholar] [CrossRef]
- Possa, S.S.; Leick, E.A.; Prado, C.M.; Martins, M.A.; Tiberio, I.F. Eosinophilic inflammation in allergic asthma. Front. Pharm. 2013, 4, 46. [Google Scholar] [CrossRef]
- Barnes, P.J. The cytokine network in asthma and chronic obstructive pulmonary disease. J. Clin. Investig. 2008, 118, 3546–3556. [Google Scholar] [CrossRef]
- Kaur, D.; Gomez, E.; Doe, C.; Berair, R.; Woodman, L.; Saunders, R.; Hollins, F.; Rose, F.R.; Amrani, Y.; May, R.; et al. IL-33 drives airway hyper-responsiveness through IL-13-mediated mast cell: Airway smooth muscle crosstalk. Allergy 2015, 70, 556–567. [Google Scholar] [CrossRef]
- Green, R.H.; Brightling, C.E.; McKenna, S.; Hargadon, B.; Parker, D.; Bradding, P.; Wardlaw, A.J.; Pavord, I.D. Asthma exacerbations and sputum eosinophil counts: A randomised controlled trial. Lancet 2002, 360, 1715–1721. [Google Scholar] [CrossRef]
- Monteseirin, J. Neutrophils and asthma. J. Investig. Allergol. Clin. Immunol. 2009, 19, 340–354. [Google Scholar] [PubMed]
- Riffo-Vasquez, Y.; Ligeiro de Oliveira, A.P.; Page, C.P.; Spina, D.; Tavares-de-Lima, W. Role of sex hormones in allergic inflammation in mice. Clin. Exp. Allergy 2007, 37, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Hamano, N.; Terada, N.; Maesako, K.; Numata, T.; Konno, A. Effect of sex hormones on eosinophilic inflammation in nasal mucosa. Allergy Asthma Proc. 1998, 19, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Busse, W.W. A role for neutrophils in asthma exacerbations. Nat. Med. 2017, 23, 658–659. [Google Scholar] [CrossRef] [PubMed]
- Sur, S.; Crotty, T.B.; Kephart, G.M.; Hyma, B.A.; Colby, T.V.; Reed, C.E.; Hunt, L.W.; Gleich, G.J. Sudden-onset fatal asthma. A distinct entity with few eosinophils and relatively more neutrophils in the airway submucosa? Am. Rev. Respir. Dis. 1993, 148, 713–719. [Google Scholar] [CrossRef]
- Cox, G. Glucocorticoid treatment inhibits apoptosis in human neutrophils. Separation of survival and activation outcomes. J. Immunol. 1995, 154, 4719–4725. [Google Scholar]
- Saffar, A.S.; Ashdown, H.; Gounni, A.S. The molecular mechanisms of glucocorticoids-mediated neutrophil survival. Curr. Drug Targets 2011, 12, 556–562. [Google Scholar] [CrossRef]
- Chen, F.; Yu, M.; Zhong, Y.; Wang, L.; Huang, H. Characteristics and Role of Neutrophil Extracellular Traps in Asthma. Inflammation 2022, 45, 6–13. [Google Scholar] [CrossRef]
- Liu, T.; Wang, F.P.; Wang, G.; Mao, H. Role of Neutrophil Extracellular Traps in Asthma and Chronic Obstructive Pulmonary Disease. Chin. Med. J. 2017, 130, 730–736. [Google Scholar] [CrossRef]
- Duvall, M.G.; Krishnamoorthy, N.; Levy, B.D. Non-type 2 inflammation in severe asthma is propelled by neutrophil cytoplasts and maintained by defective resolution. Allergol. Int. 2019, 68, 143–149. [Google Scholar] [CrossRef]
- Alexander, M.K. Sex difference in normal neutrophil count. Br. Med. J. 1975, 1, 683. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Blazkova, J.; Gupta, S.; Liu, Y.; Gaudilliere, B.; Ganio, E.A.; Bolen, C.R.; Saar-Dover, R.; Fragiadakis, G.K.; Angst, M.S.; Hasni, S.; et al. Multicenter Systems Analysis of Human Blood Reveals Immature Neutrophils in Males and During Pregnancy. J. Immunol. 2017, 198, 2479–2488. [Google Scholar] [CrossRef] [PubMed]
- Bain, B.J.; England, J.M. Normal haematological values: Sex difference in neutrophil count. Br. Med. J. 1975, 1, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Giaglis, S.; Stoikou, M.; Sur Chowdhury, C.; Schaefer, G.; Grimolizzi, F.; Rossi, S.W.; Hoesli, I.M.; Lapaire, O.; Hasler, P.; Hahn, S. Multimodal Regulation of NET Formation in Pregnancy: Progesterone Antagonizes the Pro-NETotic Effect of Estrogen and G-CSF. Front. Immunol. 2016, 7, 565. [Google Scholar] [CrossRef] [PubMed]
- Molloy, E.J.; O’Neill, A.J.; Grantham, J.J.; Sheridan-Pereira, M.; Fitzpatrick, J.M.; Webb, D.W.; Watson, R.W. Sex-specific alterations in neutrophil apoptosis: The role of estradiol and progesterone. Blood 2003, 102, 2653–2659. [Google Scholar] [CrossRef]
- Sakaguchi, H.; Fujimoto, J.; Aoki, I.; Tamaya, T. Expression of estrogen receptor alpha and beta in myometrium of premenopausal and postmenopausal women. Steroids 2003, 68, 11–19. [Google Scholar] [CrossRef]
- Robinson, D.P.; Hall, O.J.; Nilles, T.L.; Bream, J.H.; Klein, S.L. 17beta-estradiol protects females against influenza by recruiting neutrophils and increasing virus-specific CD8 T cell responses in the lungs. J. Virol. 2014, 88, 4711–4720. [Google Scholar] [CrossRef]
- Lee, G.T.; Kim, J.H.; Kwon, S.J.; Stein, M.N.; Hong, J.H.; Nagaya, N.; Billakanti, S.; Kim, M.M.; Kim, W.J.; Kim, I.Y. Dihydrotestosterone Increases Cytotoxic Activity of Macrophages on Prostate Cancer Cells via TRAIL. Endocrinology 2019, 160, 2049–2060. [Google Scholar] [CrossRef]
- Liu, C.Q.; Wu, S.Z.; Wang, Z.D.; Lai, W.Y.; Sun, F. Effect of testosterone on expression of androgen receptor in human monocytic cell line THP-1. Di Yi Jun Yi Da Xue Xue Bao 2004, 24, 389–391. [Google Scholar]
- Friedl, R.; Brunner, M.; Moeslinger, T.; Spieckermann, P.G. Testosterone inhibits expression of inducible nitric oxide synthase in murine macrophages. Life Sci. 2000, 68, 417–429. [Google Scholar] [CrossRef]
- Lai, J.J.; Lai, K.P.; Zeng, W.; Chuang, K.H.; Altuwaijri, S.; Chang, C. Androgen receptor influences on body defense system via modulation of innate and adaptive immune systems: Lessons from conditional AR knockout mice. Am. J. Pathol. 2012, 181, 1504–1512. [Google Scholar] [CrossRef] [PubMed]
- Buyon, J.P.; Korchak, H.M.; Rutherford, L.E.; Ganguly, M.; Weissmann, G. Female hormones reduce neutrophil responsiveness in vitro. Arthritis Rheum 1984, 27, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Bodel, P.; Dillard, G.M., Jr.; Kaplan, S.S.; Malawista, S.E. Anti-inflammatory effects of estradiol on human blood leukocytes. J. Lab. Clin. Med. 1972, 80, 373–384. [Google Scholar] [PubMed]
- Fahy, J.V. Eosinophilic and neutrophilic inflammation in asthma: Insights from clinical studies. Proc. Am. Thorac. Soc. 2009, 6, 256–259. [Google Scholar] [CrossRef]
- Wenzel, S.E. Asthma: Defining of the persistent adult phenotypes. Lancet 2006, 368, 804–813. [Google Scholar] [CrossRef]
- Chakir, J.; Shannon, J.; Molet, S.; Fukakusa, M.; Elias, J.; Laviolette, M.; Boulet, L.P.; Hamid, Q. Airway remodeling-associated mediators in moderate to severe asthma: Effect of steroids on TGF-beta, IL-11, IL-17, and type I and type III collagen expression. J. Allergy Clin. Immunol. 2003, 111, 1293–1298. [Google Scholar] [CrossRef]
- Molet, S.; Hamid, Q.; Davoine, F.; Nutku, E.; Taha, R.; Page, N.; Olivenstein, R.; Elias, J.; Chakir, J. IL-17 is increased in asthmatic airways and induces human bronchial fibroblasts to produce cytokines. J. Allergy Clin. Immunol. 2001, 108, 430–438. [Google Scholar] [CrossRef]
- Newcomb, D.C.; Cephus, J.Y.; Boswell, M.G.; Fahrenholz, J.M.; Langley, E.W.; Feldman, A.S.; Zhou, W.; Dulek, D.E.; Goleniewska, K.; Woodward, K.B.; et al. Estrogen and progesterone decrease let-7f microRNA expression and increase IL-23/IL-23 receptor signaling and IL-17A production in patients with severe asthma. J. Allergy Clin. Immunol. 2015, 136, 1025–1034.e1011. [Google Scholar] [CrossRef]
- Marin, D.P.; Bolin, A.P.; dos Santos Rde, C.; Curi, R.; Otton, R. Testosterone suppresses oxidative stress in human neutrophils. Cell Biochem. Funct. 2010, 28, 394–402. [Google Scholar] [CrossRef]
- Grimaldi, C.M.; Cleary, J.; Dagtas, A.S.; Moussai, D.; Diamond, B. Estrogen alters thresholds for B cell apoptosis and activation. J. Clin. Investig. 2002, 109, 1625–1633. [Google Scholar] [CrossRef]
- Buskiewicz, I.A.; Huber, S.A.; Fairweather, D. Chapter 4—Sex Hormone Receptor Expression in the Immune System. In Sex Differences in Physiology; Neigh, G.N., Mitzelfelt, M.M., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 45–60. [Google Scholar]
- Pawankar, R.; Yamagishi, S.; Yagi, T. Revisiting the roles of mast cells in allergic rhinitis and its relation to local IgE synthesis. Am. J. Rhinol. 2000, 14, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Stewart, P.; Johansen, H.; McRae, L.; Taylor, G. Sex difference in hospitalization due to asthma in relation to age. J. Clin. Epidemiol. 2003, 56, 180–187. [Google Scholar] [CrossRef]
- De Marco, R.; Locatelli, F.; Sunyer, J.; Burney, P. Differences in incidence of reported asthma related to age in men and women. A retrospective analysis of the data of the European Respiratory Health Survey. Am. J. Respir. Crit. Care Med. 2000, 162, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Masuzawa, T.; Miyaura, C.; Onoe, Y.; Kusano, K.; Ohta, H.; Nozawa, S.; Suda, T. Estrogen deficiency stimulates B lymphopoiesis in mouse bone marrow. J. Clin. Investig. 1994, 94, 1090–1097. [Google Scholar] [CrossRef]
- Narita, S.; Goldblum, R.M.; Watson, C.S.; Brooks, E.G.; Estes, D.M.; Curran, E.M.; Midoro-Horiuti, T. Environmental estrogens induce mast cell degranulation and enhance IgE-mediated release of allergic mediators. Environ. Health Perspect. 2007, 115, 48–52. [Google Scholar] [CrossRef]
- Zaitsu, M.; Narita, S.; Lambert, K.C.; Grady, J.J.; Estes, D.M.; Curran, E.M.; Brooks, E.G.; Watson, C.S.; Goldblum, R.M.; Midoro-Horiuti, T. Estradiol activates mast cells via a non-genomic estrogen receptor-alpha and calcium influx. Mol. Immunol. 2007, 44, 1977–1985. [Google Scholar] [CrossRef]
- Medina, K.L.; Kincade, P.W. Pregnancy-related steroids are potential negative regulators of B lymphopoiesis. Proc. Natl. Acad. Sci. USA 1994, 91, 5382–5386. [Google Scholar] [CrossRef]
- Keith, R.C.; Sokolove, J.; Edelman, B.L.; Lahey, L.; Redente, E.F.; Holers, V.M.; Sakaguchi, S.; Robinson, W.H.; Riches, D.W. Testosterone is protective in the sexually dimorphic development of arthritis and lung disease in SKG mice. Arthritis Rheum 2013, 65, 1487–1493. [Google Scholar] [CrossRef]
- Mizota, K.; Yoshida, A.; Uchida, H.; Fujita, R.; Ueda, H. Novel type of Gq/11 protein-coupled neurosteroid receptor sensitive to endocrine disrupting chemicals in mast cell line (RBL-2H3). Br. J. Pharm. 2005, 145, 545–550. [Google Scholar] [CrossRef]
- Lu, S.F.; Mo, Q.; Hu, S.; Garippa, C.; Simon, N.G. Dehydroepiandrosterone upregulates neural androgen receptor level and transcriptional activity. J. Neurobiol. 2003, 57, 163–171. [Google Scholar] [CrossRef]
- Mo, Q.; Lu, S.F.; Hu, S.; Simon, N.G. DHEA and DHEA sulfate differentially regulate neural androgen receptor and its transcriptional activity. Brain Res. Mol. Brain Res. 2004, 126, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. Human TH1 and TH2 subsets: Regulation of differentiation and role in protection and immunopathology. Int. Arch. Allergy Immunol. 1992, 98, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Kapsenberg, M.L.; Jansen, H.M.; Bos, J.D.; Wierenga, E.A. Role of type 1 and type 2 T helper cells in allergic diseases. Curr. Opin. Immunol. 1992, 4, 788–793. [Google Scholar] [CrossRef]
- Bergantini, L.; d’Alessandro, M.; Cameli, P.; Bono, C.; Perruzza, M.; Biagini, M.; Pini, L.; Bigliazzi, C.; Sestini, P.; Dotta, F.; et al. Regulatory T cell monitoring in severe eosinophilic asthma patients treated with mepolizumab. Scand. J. Immunol. 2021, 94, e13031. [Google Scholar] [CrossRef]
- Dembele, M.; Tao, S.; Massoud, A.H.; Miah, S.M.S.; Lelias, S.; De Groot, A.S.; Mazer, B.D. Tregitopes Improve Asthma by Promoting Highly Suppressive and Antigen-Specific Tregs. Front. Immunol. 2021, 12, 634509. [Google Scholar] [CrossRef]
- Ray, A.; Khare, A.; Krishnamoorthy, N.; Qi, Z.; Ray, P. Regulatory T cells in many flavors control asthma. Mucosal Immunol. 2010, 3, 216–229. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Sad, S. The expanding universe of T-cell subsets: Th1, Th2 and more. Immunol. Today 1996, 17, 138–146. [Google Scholar] [CrossRef]
- Kaiko, G.E.; Horvat, J.C.; Beagley, K.W.; Hansbro, P.M. Immunological decision-making: How does the immune system decide to mount a helper T-cell response? Immunology 2008, 123, 326–338. [Google Scholar] [CrossRef]
- Ying, S.; Robinson, D.S.; Varney, V.; Meng, Q.; Tsicopoulos, A.; Moqbel, R.; Durham, S.R.; Kay, A.B.; Hamid, Q. TNF alpha mRNA expression in allergic inflammation. Clin. Exp. Allergy 1991, 21, 745–750. [Google Scholar] [CrossRef]
- Berry, M.A.; Hargadon, B.; Shelley, M.; Parker, D.; Shaw, D.E.; Green, R.H.; Bradding, P.; Brightling, C.E.; Wardlaw, A.J.; Pavord, I.D. Evidence of a role of tumor necrosis factor alpha in refractory asthma. N. Engl. J. Med. 2006, 354, 697–708. [Google Scholar] [CrossRef]
- Truyen, E.; Coteur, L.; Dilissen, E.; Overbergh, L.; Dupont, L.J.; Ceuppens, J.L.; Bullens, D.M. Evaluation of airway inflammation by quantitative Th1/Th2 cytokine mRNA measurement in sputum of asthma patients. Thorax 2006, 61, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Naseer, T.; Minshall, E.M.; Leung, D.Y.; Laberge, S.; Ernst, P.; Martin, R.J.; Hamid, Q. Expression of IL-12 and IL-13 mRNA in asthma and their modulation in response to steroid therapy. Am. J. Respir. Crit. Care Med. 1997, 155, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Kroegel, C.; Julius, P.; Matthys, H.; Virchow, J.C., Jr.; Luttmann, W. Endobronchial secretion of interleukin-13 following local allergen challenge in atopic asthma: Relationship to interleukin-4 and eosinophil counts. Eur. Respir. J. 1996, 9, 899–904. [Google Scholar] [CrossRef]
- Keselman, A.; Heller, N. Estrogen Signaling Modulates Allergic Inflammation and Contributes to Sex Differences in Asthma. Front. Immunol. 2015, 6, 568. [Google Scholar] [CrossRef] [PubMed]
- Roved, J.; Westerdahl, H.; Hasselquist, D. Sex differences in immune responses: Hormonal effects, antagonistic selection, and evolutionary consequences. Horm. Behav. 2017, 88, 95–105. [Google Scholar] [CrossRef]
- Tam, A.; Morrish, D.; Wadsworth, S.; Dorscheid, D.; Man, S.F.; Sin, D.D. The role of female hormones on lung function in chronic lung diseases. BMC Womens Health 2011, 11, 24. [Google Scholar] [CrossRef]
- Fuseini, H.; Yung, J.A.; Cephus, J.Y.; Zhang, J.; Goleniewska, K.; Polosukhin, V.V.; Peebles, R.S., Jr.; Newcomb, D.C. Testosterone Decreases House Dust Mite-Induced Type 2 and IL-17A-Mediated Airway Inflammation. J. Immunol. 2018, 201, 1843–1854. [Google Scholar] [CrossRef]
- Giron-Gonzalez, J.A.; Moral, F.J.; Elvira, J.; Garcia-Gil, D.; Guerrero, F.; Gavilan, I.; Escobar, L. Consistent production of a higher TH1:TH2 cytokine ratio by stimulated T cells in men compared with women. Eur. J. Endocrinol. 2000, 143, 31–36. [Google Scholar] [CrossRef]
- Pignataro, F.S.; Bonini, M.; Forgione, A.; Melandri, S.; Usmani, O.S. Asthma and gender: The female lung. Pharm. Res. 2017, 119, 384–390. [Google Scholar] [CrossRef]
- Lelu, K.; Laffont, S.; Delpy, L.; Paulet, P.E.; Perinat, T.; Tschanz, S.A.; Pelletier, L.; Engelhardt, B.; Guery, J.C. Estrogen receptor alpha signaling in T lymphocytes is required for estradiol-mediated inhibition of Th1 and Th17 cell differentiation and protection against experimental autoimmune encephalomyelitis. J. Immunol. 2011, 187, 2386–2393. [Google Scholar] [CrossRef]
- Maselli, A.; Conti, F.; Alessandri, C.; Colasanti, T.; Barbati, C.; Vomero, M.; Ciarlo, L.; Patrizio, M.; Spinelli, F.R.; Ortona, E.; et al. Low expression of estrogen receptor beta in T lymphocytes and high serum levels of anti-estrogen receptor alpha antibodies impact disease activity in female patients with systemic lupus erythematosus. Biol Sex. Differ. 2016, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Tanabe, M.; Ito, W.; Ueki, S.; Konnno, Y.; Chihara, M.; Itoga, M.; Kobayashi, Y.; Moritoki, Y.; Kayaba, H.; et al. Gender difference in allergic airway remodelling and immunoglobulin production in mouse model of asthma. Respirology 2013, 18, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Adachi, Y.; Hasegawa, K.; Morimoto, M. Less sensitivity for late airway inflammation in males than females in BALB/c mice. Scand. J. Immunol. 2003, 57, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef] [PubMed]
- Miossec, P.; Kolls, J.K. Targeting IL-17 and TH17 cells in chronic inflammation. Nat. Rev. Drug Discov. 2012, 11, 763–776. [Google Scholar] [CrossRef]
- Tabarkiewicz, J.; Pogoda, K.; Karczmarczyk, A.; Pozarowski, P.; Giannopoulos, K. The Role of IL-17 and Th17 Lymphocytes in Autoimmune Diseases. Arch. Immunol. Exp. 2015, 63, 435–449. [Google Scholar] [CrossRef]
- Geissmann, F.; Manz, M.G.; Jung, S.; Sieweke, M.H.; Merad, M.; Ley, K. Development of monocytes, macrophages, and dendritic cells. Science 2010, 327, 656–661. [Google Scholar] [CrossRef]
- Hoffmann, F.; Ender, F.; Schmudde, I.; Lewkowich, I.P.; Kohl, J.; Konig, P.; Laumonnier, Y. Origin, Localization, and Immunoregulatory Properties of Pulmonary Phagocytes in Allergic Asthma. Front. Immunol. 2016, 7, 107. [Google Scholar] [CrossRef]
- D’Agostino, P.; Milano, S.; Barbera, C.; Di Bella, G.; La Rosa, M.; Ferlazzo, V.; Farruggio, R.; Miceli, D.M.; Miele, M.; Castagnetta, L.; et al. Sex hormones modulate inflammatory mediators produced by macrophages. Ann. N. Y. Acad. Sci. 1999, 876, 426–429. [Google Scholar] [CrossRef]
- Culley, F.J. Natural killer cells in infection and inflammation of the lung. Immunology 2009, 128, 151–163. [Google Scholar] [CrossRef]
- Abdullah, M.; Chai, P.S.; Chong, M.Y.; Tohit, E.R.; Ramasamy, R.; Pei, C.P.; Vidyadaran, S. Gender effect on in vitro lymphocyte subset levels of healthy individuals. Cell. Immunol. 2012, 272, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Lara, V.; Hernandez-Martinez, J.M.; Arrieta, O. Influence of estrogen in non-small cell lung cancer and its clinical implications. J. Thorac. Dis. 2018, 10, 482–497. [Google Scholar] [CrossRef] [PubMed]
- Kadel, S.; Kovats, S. Sex Hormones Regulate Innate Immune Cells and Promote Sex Differences in Respiratory Virus Infection. Front. Immunol. 2018, 9, 1653. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Alysandratos, K.D.; Angelidou, A.; Delivanis, D.A.; Sismanopoulos, N.; Zhang, B.; Asadi, S.; Vasiadi, M.; Weng, Z.; Miniati, A.; et al. Mast cells and inflammation. Biochim. Biophys. Acta 2012, 1822, 21–33. [Google Scholar] [CrossRef]
- Zierau, O.; Zenclussen, A.C.; Jensen, F. Role of female sex hormones, estradiol and progesterone, in mast cell behavior. Front. Immunol. 2012, 3, 169. [Google Scholar] [CrossRef]
- Reber, L.L.; Frossard, N. Targeting mast cells in inflammatory diseases. Pharmacol. Ther. 2014, 142, 416–435. [Google Scholar] [CrossRef]
- Jing, H.; Wang, Z.; Chen, Y. Effect of oestradiol on mast cell number and histamine level in the mammary glands of rat. Anat. Histol. Embryol. 2012, 41, 170–176. [Google Scholar] [CrossRef]
- Chen, W.; Beck, I.; Schober, W.; Brockow, K.; Effner, R.; Buters, J.T.; Behrendt, H.; Ring, J. Human mast cells express androgen receptors but treatment with testosterone exerts no influence on IgE-independent mast cell degranulation elicited by neuromuscular blocking agents. Exp. Derm. 2010, 19, 302–304. [Google Scholar] [CrossRef]
- Vasiadi, M.; Kempuraj, D.; Boucher, W.; Kalogeromitros, D.; Theoharides, T.C. Progesterone inhibits mast cell secretion. Int. J. Immunopathol. Pharm. 2006, 19, 787–794. [Google Scholar] [CrossRef]
- Ligeiro de Oliveira, A.P.; Oliveira-Filho, R.M.; da Silva, Z.L.; Borelli, P.; Tavares de Lima, W. Regulation of allergic lung inflammation in rats: Interaction between estradiol and corticosterone. Neuroimmunomodulation 2004, 11, 20–27. [Google Scholar] [CrossRef]
- Janicki, J.S.; Spinale, F.G.; Levick, S.P. Gender differences in non-ischemic myocardial remodeling: Are they due to estrogen modulation of cardiac mast cells and/or membrane type 1 matrix metalloproteinase. Pflug. Arch. 2013, 465, 687–697. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, Q.; Nian, S.; Ye, Y.; Liu, D.; Yu, H.; Xiong, H.; Pan, B.; Xiao, L.; Fan, C.; Yuan, Q. The Emerging Roles of T Helper Cell Subsets and Cytokines in Severe Neutrophilic Asthma. Inflammation 2021, 45, 1007–1022. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Yamaguchi, T.; Nomura, T.; Ono, M. Regulatory T cells and immune tolerance. Cell 2008, 133, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T. Regulatory T cells: How do they suppress immune responses? Int. Immunol. 2009, 21, 1105–1111. [Google Scholar] [CrossRef]
- Togashi, Y.; Shitara, K.; Nishikawa, H. Regulatory T cells in cancer immunosuppression—Implications for anticancer therapy. Nat. Rev. Clin. Oncol. 2019, 16, 356–371. [Google Scholar] [CrossRef]
- Leffler, J.; Stumbles, P.A.; Strickland, D.H. Immunological Processes Driving IgE Sensitisation and Disease Development in Males and Females. Int. J. Mol. Sci. 2018, 19, 1554. [Google Scholar] [CrossRef]
- Zhao, S.T.; Wang, C.Z. Regulatory T cells and asthma. J. Zhejiang Univ. Sci. B 2018, 19, 663–673. [Google Scholar] [CrossRef]
- Stampfli, M.R.; Cwiartka, M.; Gajewska, B.U.; Alvarez, D.; Ritz, S.A.; Inman, M.D.; Xing, Z.; Jordana, M. Interleukin-10 gene transfer to the airway regulates allergic mucosal sensitization in mice. Am. J. Respir. Cell Mol. Biol. 1999, 21, 586–596. [Google Scholar] [CrossRef]
- Hansen, G.; McIntire, J.J.; Yeung, V.P.; Berry, G.; Thorbecke, G.J.; Chen, L.; DeKruyff, R.H.; Umetsu, D.T. CD4(+) T helper cells engineered to produce latent TGF-beta1 reverse allergen-induced airway hyperreactivity and inflammation. J. Clin. Investig. 2000, 105, 61–70. [Google Scholar] [CrossRef]
- Nakao, A.; Miike, S.; Hatano, M.; Okumura, K.; Tokuhisa, T.; Ra, C.; Iwamoto, I. Blockade of transforming growth factor beta/Smad signaling in T cells by overexpression of Smad7 enhances antigen-induced airway inflammation and airway reactivity. J. Exp. Med. 2000, 192, 151–158. [Google Scholar] [CrossRef]
- Oh, J.W.; Seroogy, C.M.; Meyer, E.H.; Akbari, O.; Berry, G.; Fathman, C.G.; Dekruyff, R.H.; Umetsu, D.T. CD4 T-helper cells engineered to produce IL-10 prevent allergen-induced airway hyperreactivity and inflammation. J. Allergy Clin. Immunol. 2002, 110, 460–468. [Google Scholar] [CrossRef] [PubMed]
- John, M.; Lim, S.; Seybold, J.; Jose, P.; Robichaud, A.; O’Connor, B.; Barnes, P.J.; Chung, K.F. Inhaled corticosteroids increase interleukin-10 but reduce macrophage inflammatory protein-1alpha, granulocyte-macrophage colony-stimulating factor, and interferon-gamma release from alveolar macrophages in asthma. Am. J. Respir. Crit. Care Med. 1998, 157, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Zhong, Q.; Palmer, T.; Benner, A.; Wang, L.; Suresh, K.; Damico, R.; D’Alessio, F.R. Estradiol resolves pneumonia via ERbeta in regulatory T cells. JCI Insight 2021, 6, e133251. [Google Scholar] [CrossRef] [PubMed]
- Kalidhindi, R.S.R.; Ambhore, N.S.; Bhallamudi, S.; Loganathan, J.; Sathish, V. Role of Estrogen Receptors alpha and beta in a Murine Model of Asthma: Exacerbated Airway Hyperresponsiveness and Remodeling in ERbeta Knockout Mice. Front. Pharm. 2019, 10, 1499. [Google Scholar] [CrossRef]
- Velez-Ortega, A.C.; Temprano, J.; Reneer, M.C.; Ellis, G.I.; McCool, A.; Gardner, T.; Khosravi, M.; Marti, F. Enhanced generation of suppressor T cells in patients with asthma taking oral contraceptives. J. Asthma 2013, 50, 223–230. [Google Scholar] [CrossRef]
- Gandhi, V.D.; Cephus, J.Y.; Norlander, A.E.; Chowdhury, N.U.; Zhang, J.; Ceneviva, Z.J.; Tannous, E.; Polosukhin, V.V.; Putz, N.D.; Wickersham, N.; et al. Androgen receptor signaling promotes Treg suppressive function during allergic airway inflammation. J. Clin. Investig. 2022, 132, e153397. [Google Scholar] [CrossRef] [PubMed]
- Arruvito, L.; Sanz, M.; Banham, A.H.; Fainboim, L. Expansion of CD4+CD25+and FOXP3+ regulatory T cells during the follicular phase of the menstrual cycle: Implications for human reproduction. J. Immunol. 2007, 178, 2572–2578. [Google Scholar] [CrossRef] [PubMed]
- Weyand, C.M.; Goronzy, J.J. Aging of the Immune System. Mechanisms and Therapeutic Targets. Ann. Am. Thorac. Soc. 2016, 13 (Suppl. S5), S422–S428. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, T.; Shimizu, J.; Iida, R.; Yamazaki, S.; Sakaguchi, S. CD4+CD25+Foxp3+ T cells and CD4+CD25-Foxp3+ T cells in aged mice. J. Immunol. 2006, 176, 6586–6593. [Google Scholar] [CrossRef]
- Townsend, E.A.; Sathish, V.; Thompson, M.A.; Pabelick, C.M.; Prakash, Y.S. Estrogen effects on human airway smooth muscle involve cAMP and protein kinase A. Am. J. Physiol. Lung Cell Mol. Physiol. 2012, 303, L923–L928. [Google Scholar] [CrossRef]
- Zarazua, A.; Gonzalez-Arenas, A.; Ramirez-Velez, G.; Bazan-Perkins, B.; Guerra-Araiza, C.; Campos-Lara, M.G. Sexual Dimorphism in the Regulation of Estrogen, Progesterone, and Androgen Receptors by Sex Steroids in the Rat Airway Smooth Muscle Cells. Int. J. Endocrinol. 2016, 2016, 8423192. [Google Scholar] [CrossRef] [PubMed]
- Haggerty, C.L.; Ness, R.B.; Kelsey, S.; Waterer, G.W. The impact of estrogen and progesterone on asthma. Ann. Allergy Asthma Immunol. 2003, 90, 284–291. [Google Scholar] [CrossRef]
- Paech, K.; Webb, P.; Kuiper, G.G.; Nilsson, S.; Gustafsson, J.; Kushner, P.J.; Scanlan, T.S. Differential ligand activation of estrogen receptors ERalpha and ERbeta at AP1 sites. Science 1997, 277, 1508–1510. [Google Scholar] [CrossRef] [PubMed]
- Bhallamudi, S.; Connell, J.; Pabelick, C.M.; Prakash, Y.S.; Sathish, V. Estrogen receptors differentially regulate intracellular calcium handling in human nonasthmatic and asthmatic airway smooth muscle cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 318, L112–L124. [Google Scholar] [CrossRef] [PubMed]
- Ambhore, N.S.; Katragadda, R.; Raju Kalidhindi, R.S.; Thompson, M.A.; Pabelick, C.M.; Prakash, Y.S.; Sathish, V. Estrogen receptor beta signaling inhibits PDGF induced human airway smooth muscle proliferation. Mol. Cell Endocrinol. 2018, 476, 37–47. [Google Scholar] [CrossRef]
- Ambhore, N.S.; Kalidhindi, R.S.R.; Pabelick, C.M.; Hawse, J.R.; Prakash, Y.S.; Sathish, V. Differential estrogen-receptor activation regulates extracellular matrix deposition in human airway smooth muscle remodeling via NF-kappaB pathway. FASEB J. 2019, 33, 13935–13950. [Google Scholar] [CrossRef]
- Borkar, N.A.; Ambhore, N.S.; Kalidhindi, R.S.R.; Pabelick, C.M.; Prakash, Y.S.; Sathish, V. Kisspeptins inhibit human airway smooth muscle proliferation. JCI Insight 2022, 7, e152762. [Google Scholar] [CrossRef]
- Perusqia, M. Correlation between nongenomic action of C19-steroids and COVID-19 severity. Physiol. Res. 2021, 70, S135–S144. [Google Scholar] [CrossRef]
- Smith, L.C.; Moreno, S.; Robertson, L.; Robinson, S.; Gant, K.; Bryant, A.J.; Sabo-Attwood, T. Transforming growth factor beta1 targets estrogen receptor signaling in bronchial epithelial cells. Respir. Res. 2018, 19, 160. [Google Scholar] [CrossRef]
- Bylander, A.; Nutu, M.; Wellander, R.; Goksor, M.; Billig, H.; Larsson, D.G. Rapid effects of progesterone on ciliary beat frequency in the mouse fallopian tube. Reprod. Biol. Endocrinol. 2010, 8, 48. [Google Scholar] [CrossRef]
- Tofovic, S.P.; Zhang, X.; Jackson, E.K.; Zhu, H.; Petrusevska, G. 2-methoxyestradiol attenuates bleomycin-induced pulmonary hypertension and fibrosis in estrogen-deficient rats. Vasc. Pharm. 2009, 51, 190–197. [Google Scholar] [CrossRef]
- Wen, J.; Wang, J.; Tang, X.; Deng, S.; Dai, J.; Li, X.; Cai, W. DHT deteriorates the progression of monocrotaline-induced pulmonary arterial hypertension: Effects of endogenous and exogenous androgen. Am. J. Transl. Res. 2019, 11, 5752–5763. [Google Scholar] [PubMed]
- Chetty, A.; Nielsen, H.C. Targeting Airway Smooth Muscle Hypertrophy in Asthma: An Approach Whose Time Has Come. J. Asthma Allergy 2021, 14, 539–556. [Google Scholar] [CrossRef] [PubMed]
- Kalidhindi, R.S.R.; Borkar, N.A.; Ambhore, N.S.; Pabelick, C.M.; Prakash, Y.S.; Sathish, V. Sex steroids skew ACE2 expression in human airway: A contributing factor to sex differences in COVID-19? Am. J. Physiol. Lung Cell Mol. Physiol. 2020, 319, L843–L847. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. COVID-19. Available online: https://www.cdc.gov/coronavirus/2019-ncov/your-health/about-covid-19.html (accessed on 15 May 2022).
- Lombardi, C.; Gani, F.; Berti, A.; Comberiati, P.; Peroni, D.; Cottini, M. Asthma and COVID-19: A dangerous liaison? Asthma Res. Pract. 2021, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Morais-Almeida, M.; Pite, H.; Aguiar, R.; Ansotegui, I.; Bousquet, J. Asthma and the Coronavirus Disease 2019 Pandemic: A Literature Review. Int. Arch. Allergy Immunol. 2020, 181, 680–688. [Google Scholar] [CrossRef]
- Sunjaya, A.P.; Allida, S.M.; Di Tanna, G.L.; Jenkins, C. Asthma and risk of infection, hospitalization, ICU admission and mortality from COVID-19: Systematic review and meta-analysis. J. Asthma 2022, 59, 866–879. [Google Scholar] [CrossRef]
- Markle, J.G.; Fish, E.N. SeXX matters in immunity. Trends Immunol. 2014, 35, 97–104. [Google Scholar] [CrossRef]
- Klein, S.L.; Dhakal, S.; Ursin, R.L.; Deshpande, S.; Sandberg, K.; Mauvais-Jarvis, F. Biological sex impacts COVID-19 outcomes. PLoS Pathog. 2020, 16, e1008570. [Google Scholar] [CrossRef]
- Lu, W.; Eapen, M.S.; Singhera, G.K.; Markos, J.; Haug, G.; Chia, C.; Larby, J.; Brake, S.J.; Westall, G.P.; Jaffar, J.; et al. Angiotensin-Converting Enzyme 2 (ACE2), Transmembrane Peptidase Serine 2 (TMPRSS2), and Furin Expression Increases in the Lungs of Patients with Idiopathic Pulmonary Fibrosis (IPF) and Lymphangioleiomyomatosis (LAM): Implications for SARS-CoV-2 (COVID-19) Infections. J. Clin. Med. 2022, 11, 777. [Google Scholar] [CrossRef]
- Takahashi, T.; Ellingson, M.K.; Wong, P.; Israelow, B.; Lucas, C.; Klein, J.; Silva, J.; Mao, T.; Oh, J.E.; Tokuyama, M.; et al. Sex differences in immune responses that underlie COVID-19 disease outcomes. Nature 2020, 588, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Barranco, M.; Rivas-Garcia, L.; Quiles, J.L.; Redondo-Sanchez, D.; Aranda-Ramirez, P.; Llopis-Gonzalez, J.; Sanchez Perez, M.J.; Sanchez-Gonzalez, C. The spread of SARS-CoV-2 in Spain: Hygiene habits, sociodemographic profile, mobility patterns and comorbidities. Environ. Res. 2021, 192, 110223. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borkar, N.A.; Combs, C.K.; Sathish, V. Sex Steroids Effects on Asthma: A Network Perspective of Immune and Airway Cells. Cells 2022, 11, 2238. https://doi.org/10.3390/cells11142238
Borkar NA, Combs CK, Sathish V. Sex Steroids Effects on Asthma: A Network Perspective of Immune and Airway Cells. Cells. 2022; 11(14):2238. https://doi.org/10.3390/cells11142238
Chicago/Turabian StyleBorkar, Niyati A., Colin Kelly Combs, and Venkatachalem Sathish. 2022. "Sex Steroids Effects on Asthma: A Network Perspective of Immune and Airway Cells" Cells 11, no. 14: 2238. https://doi.org/10.3390/cells11142238
APA StyleBorkar, N. A., Combs, C. K., & Sathish, V. (2022). Sex Steroids Effects on Asthma: A Network Perspective of Immune and Airway Cells. Cells, 11(14), 2238. https://doi.org/10.3390/cells11142238