
High Expression of a Cancer Stemness-Related Gene, Chromobox 8 (CBX8), in Normal Tissue Adjacent to the Tumor (NAT) Is Associated with Poor Prognosis of Colorectal Cancer Patients

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Specimens
2.2. Blood Immune Cell Profile of CRC Patients
2.3. RNA Extraction and Quantitative PCR (qPCR) Analysis
2.4. Expression Correlation of CBX8 with Other Pluripotency-Associated Genes Using an Online Database
2.5. Immunohistochemistry
2.6. Statistical Analysis
3. Results
3.1. CBX8 Gene and Protein Expressions in CRC and Normal Tissue Adjacent to Tumor (NAT) Specimens and Their Clinicopathological Significances
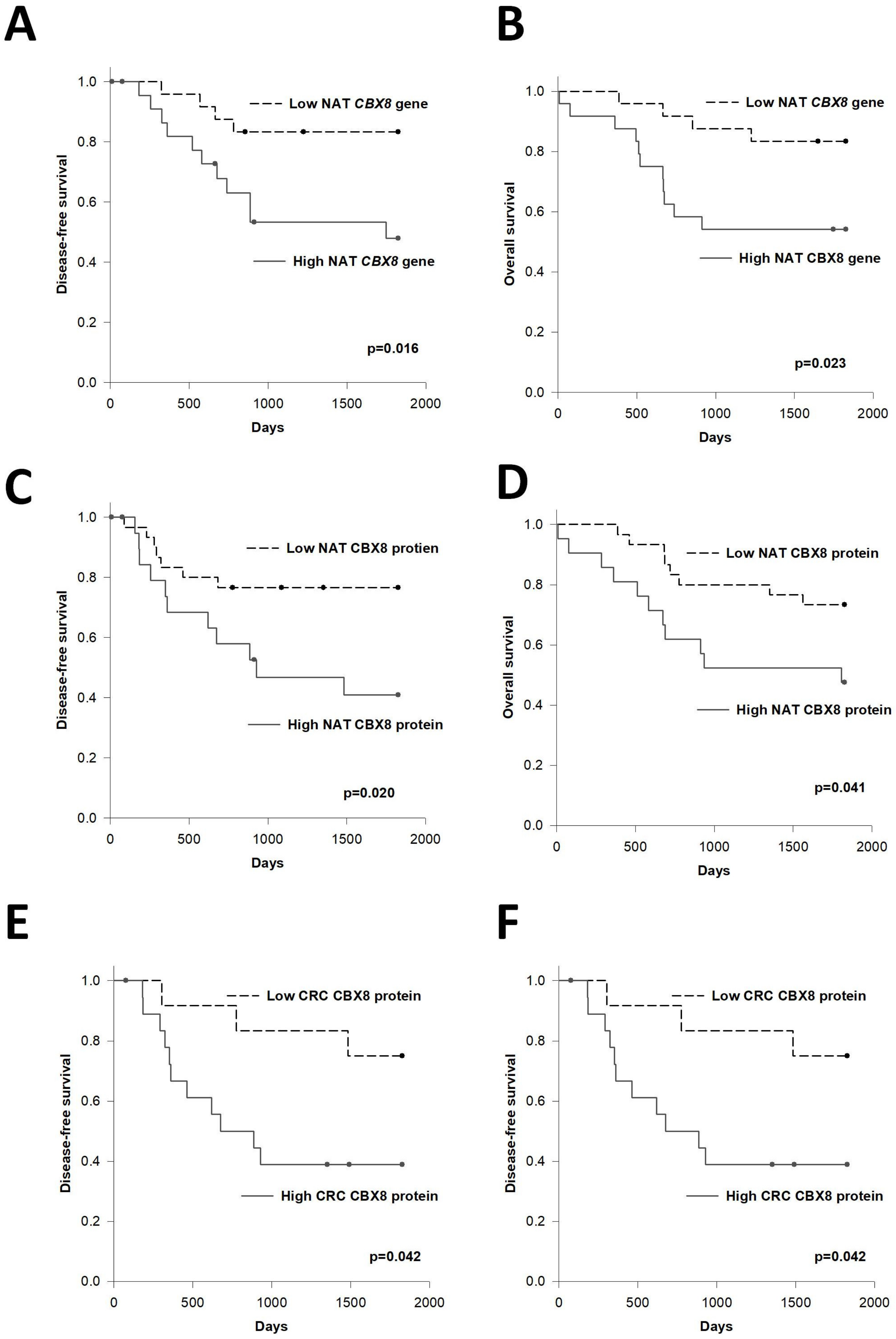
3.2. Prognostic Significance of CBX8
3.3. Association between CBX8 Expression and Immune Cell Profile of CRC Patients
3.4. Association of CBX8 with Pluripotency-Associated Genes in CRC and NAT
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allemani, C.; Weir, H.K.; Carreira, H.; Harewood, R.; Spika, D.; Wang, X.S.; Bannon, F.; Ahn, J.V.; Johnson, C.J.; Bonaventure, A.; et al. Global surveillance of cancer survival 1995–2009: Analysis of individual data for 25,676,887 patients from 279 population-based registries in 67 countries (CONCORD-2). Lancet 2015, 385, 977–1010. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.; Park, D.J.; Yang, D.; El-Khoueiry, A.; Sherrod, A.; Groshen, S.; Streeter, O.; Iqbal, S.; Danenberg, K.D.; Lenz, H.J. Gene expression in tumor-adjacent normal tissue is associated with recurrence in patients with rectal cancer treated with adjuvant chemoradiation. Pharm. Genom. 2006, 16, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Kuan, J.C.; Wu, C.C.; Sun, C.A.; Chu, C.M.; Lin, F.G.; Hsu, C.H.; Kan, P.C.; Lin, S.C.; Yang, T.; Chou, Y.C. DNA methylation combinations in adjacent normal colon tissue predict cancer recurrence: Evidence from a clinical cohort study. PLoS ONE 2015, 10, e0123396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassi, A.; Perilli, L.; Albertoni, L.; Tessarollo, S.; Mescoli, C.; Urso, E.D.L.; Fassan, M.; Rugge, M.; Zanovello, P. A coordinate deregulation of microRNAs expressed in mucosa adjacent to tumor predicts relapse after resection in localized colon cancer. Mol. Cancer 2018, 17, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.; Qin, F.; Yuan, H.; He, B.; Yang, N.; Zhang, Y.; Ren, H.; Zeng, Y. Normal tissue adjacent to tumor expression profile analysis developed and validated a prognostic model based on Hippo-related genes in hepatocellular carcinoma. Cancer Med. 2021, 10, 3139–3152. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Feng, Y.; Ye, J.; Han, Z.; Liang, Y.; Chen, Q.; Xu, X.; Huang, Y.; Jia, Z.; Zhong, W. Prediction of Biochemical Recurrence-Free Survival of Prostate Cancer Patients Leveraging Multiple Gene Expression Profiles in Tumor Microenvironment. Front. Oncol. 2021, 11, 632571. [Google Scholar] [CrossRef]
- Reis, P.P.; Waldron, L.; Perez-Ordonez, B.; Pintilie, M.; Galloni, N.N.; Xuan, Y.; Cervigne, N.K.; Warner, G.C.; Makitie, A.A.; Simpson, C.; et al. A gene signature in histologically normal surgical margins is predictive of oral carcinoma recurrence. BMC Cancer 2011, 11, 437. [Google Scholar] [CrossRef] [Green Version]
- Dadiani, M.; Bossel Ben-Moshe, N.; Paluch-Shimon, S.; Perry, G.; Balint, N.; Marin, I.; Pavlovski, A.; Morzaev, D.; Kahana-Edwin, S.; Yosepovich, A.; et al. Tumor Evolution Inferred by Patterns of microRNA Expression through the Course of Disease, Therapy, and Recurrence in Breast Cancer. Clin. Cancer Res. 2016, 22, 3651–3662. [Google Scholar] [CrossRef] [Green Version]
- Ganci, F.; Sacconi, A.; Manciocco, V.; Covello, R.; Benevolo, M.; Rollo, F.; Strano, S.; Valsoni, S.; Bicciato, S.; Spriano, G.; et al. Altered peritumoral microRNA expression predicts head and neck cancer patients with a high risk of recurrence. Mod. Pathol. 2017, 30, 1387–1401. [Google Scholar] [CrossRef] [Green Version]
- Van Wijnen, A.J.; Bagheri, L.; Badreldin, A.A.; Larson, A.N.; Dudakovic, A.; Thaler, R.; Paradise, C.R.; Wu, Z. Biological functions of chromobox (CBX) proteins in stem cell self-renewal, lineage-commitment, cancer and development. Bone 2021, 143, 115659. [Google Scholar] [CrossRef]
- Yang, S.; Liu, W.; Li, M.; Wen, J.; Zhu, M.; Xu, S. Insulin-Like Growth Factor-1 Modulates Polycomb Cbx8 Expression and Inhibits Colon Cancer Cell Apoptosis. Cell Biochem. Biophys. 2015, 71, 1503–1507. [Google Scholar] [CrossRef]
- Tang, J.; Wang, G.; Zhang, M.; Li, F.Y.; Sang, Y.; Wang, B.; Hu, K.; Wu, Y.; Luo, R.; Liao, D.; et al. Paradoxical role of CBX8 in proliferation and metastasis of colorectal cancer. Oncotarget 2014, 5, 10778–10790. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Kang, M.; Zhang, B.; Meng, F.; Song, J.; Kaneko, H.; Shimamoto, F.; Tang, B. m(6)A modification-mediated CBX8 induction regulates stemness and chemosensitivity of colon cancer via upregulation of LGR5. Mol. Cancer 2019, 18, 185. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.; Tian, Y.; Liao, Y.; Li, Z.; Yu, S.; Su, H.; Zhong, F.; Yuan, G.; Wang, Y.; Yu, H.; et al. CBX8 exhibits oncogenic properties and serves as a prognostic factor in hepatocellular carcinoma. Cell Death Dis. 2019, 10, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, C.Y.; Sun, Z.; Mullokandov, G.; Bosch, A.; Qadeer, Z.A.; Cihan, E.; Rapp, Z.; Parsons, R.; Aguirre-Ghiso, J.A.; Farias, E.F.; et al. Cbx8 Acts Non-canonically with Wdr5 to Promote Mammary Tumorigenesis. Cell Rep. 2016, 16, 472–486. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Tang, T.; Li, C.; Liu, X.; Zhou, L. CBX8 and CD96 Are Important Prognostic Biomarkers of Colorectal Cancer. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 7820–7827. [Google Scholar] [CrossRef]
- Li, Q.; Pan, Y.; Cao, Z.; Zhao, S. Comprehensive Analysis of Prognostic Value and Immune Infiltration of Chromobox Family Members in Colorectal Cancer. Front. Oncol. 2020, 10, 582667. [Google Scholar] [CrossRef]
- Aran, D.; Camarda, R.; Odegaard, J.; Paik, H.; Oskotsky, B.; Krings, G.; Goga, A.; Sirota, M.; Butte, A.J. Comprehensive analysis of normal adjacent to tumor transcriptomes. Nat. Commun. 2017, 8, 1077. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Research, N.; Kandoth, C.; Schultz, N.; Cherniack, A.D.; Akbani, R.; Liu, Y.; Shen, H.; Robertson, A.G.; Pashtan, I.; Shen, R.; et al. Integrated genomic characterization of endometrial carcinoma. Nature 2013, 497, 67–73. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas (TCGA) Research Network. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Cancer Genome Atlas Research Network. Comprehensive molecular characterization of urothelial bladder carcinoma. Nature 2014, 507, 315–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.C.Y.; Diakos, C.I.; Chan, D.L.H.; Engel, A.; Pavlakis, N.; Gill, A.; Clarke, S.J. A Longitudinal Investigation of Inflammatory Markers in Colorectal Cancer Patients Perioperatively Demonstrates Benefit in Serial Remeasurement. Ann. Surg. 2018, 267, 1119–1125. [Google Scholar] [CrossRef]
- Yasui, K.; Shida, D.; Nakamura, Y.; Ahiko, Y.; Tsukamoto, S.; Kanemitsu, Y. Postoperative, but not preoperative, inflammation-based prognostic markers are prognostic factors in stage III colorectal cancer patients. Br. J. Cancer 2021, 124, 933–941. [Google Scholar] [CrossRef]
- Ng, L.; Foo, D.C.; Wong, C.K.; Man, A.T.; Lo, O.S.; Law, W.L. Repurposing DPP-4 Inhibitors for Colorectal Cancer: A Retrospective and Single Center Study. Cancers 2021, 13, 3588. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Lin, G.; Ye, M.; Tong, D.; Zhao, J.; Zhu, D.; Yu, Q.; Zhang, W.; Li, W. Decreased mean platelet volume predicts poor prognosis in metastatic colorectal cancer patients treated with first-line chemotherapy: Results from mCRC biomarker study. BMC Cancer 2019, 19, 15. [Google Scholar] [CrossRef]
- Lin, J.K.; Lin, C.C.; Yang, S.H.; Wang, H.S.; Jiang, J.K.; Lan, Y.T.; Lin, T.C.; Li, A.F.; Chen, W.S.; Chang, S.C. Early postoperative CEA level is a better prognostic indicator than is preoperative CEA level in predicting prognosis of patients with curable colorectal cancer. Int. J. Colorectal Dis. 2011, 26, 1135–1141. [Google Scholar] [CrossRef]
- You, W.; Yan, L.; Cai, Z.; Xie, L.; Sheng, N.; Wang, G.; Wu, X.; Wang, Z. Clinical Significances of Positive Postoperative Serum CEA and Post-preoperative CEA Increment in Stage II and III Colorectal Cancer: A Multicenter Retrospective Study. Front. Oncol. 2020, 10, 671. [Google Scholar] [CrossRef]
- Bhatti, I.; Patel, M.; Dennison, A.R.; Thomas, M.W.; Garcea, G. Utility of postoperative CEA for surveillance of recurrence after resection of primary colorectal cancer. Int. J. Surg. 2015, 16, 123–128. [Google Scholar] [CrossRef]
- Konishi, T.; Shimada, Y.; Hsu, M.; Tufts, L.; Jimenez-Rodriguez, R.; Cercek, A.; Yaeger, R.; Saltz, L.; Smith, J.J.; Nash, G.M.; et al. Association of Preoperative and Postoperative Serum Carcinoembryonic Antigen and Colon Cancer Outcome. JAMA Oncol. 2018, 4, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, T.; Ginzac, A.; Molnar, I.; Bailly, S.; Durando, X.; Mahammedi, H. Eosinophil counts as a relevant prognostic marker for response to nivolumab in the management of renal cell carcinoma: A retrospective study. Cancer Med. 2021, 10, 6705–6713. [Google Scholar] [CrossRef]
- Lin, C.H.; Chou, W.C.; Wu, Y.Y.; Lin, C.Y.; Chang, K.P.; Liao, C.T.; Ho, T.Y.; Yeh, C.M.; Liu, C.J.; Hung, S.P.; et al. Prognostic significance of dynamic changes in lymphocyte-to-monocyte ratio in patients with head and neck cancer treated with radiotherapy: Results from a large cohort study. Radiother. Oncol. 2021, 154, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Slaughter, D.P.; Southwick, H.W.; Smejkal, W. Field cancerization in oral stratified squamous epithelium; clinical implications of multicentric origin. Cancer 1953, 6, 963–968. [Google Scholar] [CrossRef]
- Graham, K.; Ge, X.; de Las Morenas, A.; Tripathi, A.; Rosenberg, C.L. Gene expression profiles of estrogen receptor-positive and estrogen receptor-negative breast cancers are detectable in histologically normal breast epithelium. Clin. Cancer Res. 2011, 17, 236–246. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Wang, Y.; Zhang, C.; Chen, M.Y. Upregulated CBX8 Promotes Cancer Metastasis via the WNK2/MMP2 Pathway. Mol. Ther. Oncolytics 2020, 19, 188–196. [Google Scholar] [CrossRef]
- Wang, S.; Denton, K.E.; Hobbs, K.F.; Weaver, T.; McFarlane, J.M.B.; Connelly, K.E.; Gignac, M.C.; Milosevich, N.; Hof, F.; Paci, I.; et al. Optimization of Ligands Using Focused DNA-Encoded Libraries To Develop a Selective, Cell-Permeable CBX8 Chromodomain Inhibitor. ACS Chem. Biol. 2020, 15, 112–131. [Google Scholar] [CrossRef]
- Han, S.; Yang, W.; Zong, S.; Li, H.; Liu, S.; Li, W.; Shi, Q.; Hou, F. Clinicopathological, prognostic and predictive value of CD166 expression in colorectal cancer: A meta-analysis. Oncotarget 2017, 8, 64373–64384. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Chen, Y.; Wei, L.; Wu, S.; Shen, K.; Liu, C.; Dong, Y.; Zhao, Y.; Zhang, Y.; Zhang, C.; et al. ABHD5 inhibits YAP-induced c-Met overexpression and colon cancer cell stemness via suppressing YAP methylation. Nat. Commun. 2021, 12, 6711. [Google Scholar] [CrossRef]
- Long, J.; Yin, Y.; Guo, H.; Li, S.; Sun, Y.; Zeng, C.; Zhu, W. The mechanisms and clinical significance of PDCD4 in colorectal cancer. Gene 2019, 680, 59–64. [Google Scholar] [CrossRef]
- Bai, Y.; Shang, Q.; Zhao, H.; Pan, Z.; Guo, C.; Zhang, L.; Wang, Q. Pdcd4 restrains the self-renewal and white-to-beige transdifferentiation of adipose-derived stem cells. Cell Death Dis. 2016, 7, e2169. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.D.; Ba, H.; Chen, J.; Zhang, M.; Li, P. Celastrus orbiculatus Extract Reduces Stemness of Gastric Cancer Stem Cells by Targeting PDCD4 and EIF3H. Integr. Cancer Ther. 2021, 20, 15347354211058168. [Google Scholar] [CrossRef]
- Liu, D.; Ke, J.; Liu, Y.; Rao, H.; Tang, Z.; Liu, Y.; Zhang, Z.; You, L.; Luo, X.; Sun, Z.; et al. The interaction between PDCD4 and YB1 is critical for cervical cancer stemness and cisplatin resistance. Mol. Carcinog. 2021, 60, 813–825. [Google Scholar] [CrossRef]
- Moore, P.A.; Shah, K.; Yang, Y.; Alderson, R.; Roberts, P.; Long, V.; Liu, D.; Li, J.C.; Burke, S.; Ciccarone, V.; et al. Development of MGD007, a gpA33 x CD3-Bispecific DART Protein for T-Cell Immunotherapy of Metastatic Colorectal Cancer. Mol. Cancer Ther. 2018, 17, 1761–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Guo, H.F.; Xu, H.; Cheung, N.V. Development of a Tetravalent Anti-GPA33/Anti-CD3 Bispecific Antibody for Colorectal Cancers. Mol. Cancer Ther. 2018, 17, 2164–2175. [Google Scholar] [CrossRef] [Green Version]
- Elbadawy, M.; Usui, T.; Yamawaki, H.; Sasaki, K. Emerging Roles of C-Myc in Cancer Stem Cell-Related Signaling and Resistance to Cancer Chemotherapy: A Potential Therapeutic Target Against Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 2340. [Google Scholar] [CrossRef] [Green Version]
- Schaub, F.X.; Dhankani, V.; Berger, A.C.; Trivedi, M.; Richardson, A.B.; Shaw, R.; Zhao, W.; Zhang, X.; Ventura, A.; Liu, Y.; et al. Pan-cancer Alterations of the MYC Oncogene and Its Proximal Network across the Cancer Genome Atlas. Cell Syst. 2018, 6, 282–300.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blancato, J.; Singh, B.; Liu, A.; Liao, D.J.; Dickson, R.B. Correlation of amplification and overexpression of the c-myc oncogene in high-grade breast cancer: FISH, in situ hybridisation and immunohistochemical analyses. Br. J. Cancer 2004, 90, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Meyer, N.; Penn, L.Z. Reflecting on 25 years with MYC. Nat. Rev. Cancer 2008, 8, 976–990. [Google Scholar] [CrossRef]
- Strippoli, A.; Cocomazzi, A.; Basso, M.; Cenci, T.; Ricci, R.; Pierconti, F.; Cassano, A.; Fiorentino, V.; Barone, C.; Bria, E.; et al. c-MYC Expression Is a Possible Keystone in the Colorectal Cancer Resistance to EGFR Inhibitors. Cancers 2020, 12, 638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Wang, G.; Hao, D.; Liu, X.; Wang, D.; Ning, N.; Li, X. Aberrant regulation of the LIN28A/LIN28B and let-7 loop in human malignant tumors and its effects on the hallmarks of cancer. Mol. Cancer 2015, 14, 125. [Google Scholar] [CrossRef] [Green Version]
- Cacciola, N.A.; Calabrese, C.; Malapelle, U.; Pellino, G.; De Stefano, A.; Sepe, R.; Sgariglia, R.; Quintavalle, C.; Federico, A.; Bianco, A.; et al. UbcH10 expression can predict prognosis and sensitivity to the antineoplastic treatment for colorectal cancer patients. Mol. Carcinog. 2016, 55, 793–807. [Google Scholar] [CrossRef]
- Liu, X.; Ma, F.; Liu, C.; Zhu, K.; Li, W.; Xu, Y.; Li, G.; Niu, Z.; Liu, J.; Chen, D.; et al. UBE2O promotes the proliferation, EMT and stemness properties of breast cancer cells through the UBE2O/AMPKalpha2/mTORC1-MYC positive feedback loop. Cell Death Dis. 2020, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Milosevich, N.; Wilson, C.R.; Brown, T.M.; Alpsoy, A.; Wang, S.; Connelly, K.E.; Sinclair, K.A.D.; Ponio, F.R.; Hof, R.; Dykhuizen, E.C.; et al. Polycomb Paralog Chromodomain Inhibitors Active against Both CBX6 and CBX8*. ChemMedChem 2021, 16, 3027–3034. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.L.; Bsteh, D.; Hart, B.; Si, Y.; Weaver, T.M.; Pribitzer, C.; Lau, R.; Soni, S.; Ogana, H.; Rectenwald, J.M.; et al. Reprogramming CBX8-PRC1 function with a positive allosteric modulator. Cell Chem. Biol. 2021, 29, 555–571.e11. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Category | Number of Cases | Tumor CBX8 | p-Value | NAT CBX8 | p-Value |
|---|---|---|---|---|---|---|
| Gender | Male | 52 | 0.000592 | NS | 0.000251 | NS |
| Female | 43 | 0.000526 | 0.000260 | |||
| Age | ≤70 | 36 | 0.000543 | NS | 0.000249 | NS |
| >70 | 59 | 0.000564 | 0.000260 | |||
| Location | Colon | 63 | 0.000575 | NS | 0.000292 | 0.004 |
| Rectum | 32 | 0.000520 | 0.000184 | |||
| Tumor Size | <5 cm | 54 | 0.000546 | NS | 0.000237 | NS |
| ≥5 cm | 41 | 0.000569 | 0.000281 | |||
| TNM Staging | I to II | 39 | 0.000584 | NS | 0.000208 | 0.026 |
| III to IV | 56 | 0.000537 | 0.000289 | |||
| Tumor invasion | T1 to T3 | 79 | 0.000578 | NS | 0.000239 | 0.035 |
| T4 | 16 | 0.000449 | 0.000339 | |||
| Lymph node metastasis | Absent | 48 | 0.000544 | NS | 0.000219 | 0.034 |
| Present | 47 | 0.000569 | 0.000294 | |||
| Distant Metastasis | Present | 73 | 0.000572 | NS | 0.000236 | 0.044 |
| Absent | 22 | 0.000506 | 0.000321 |
| Tumor | NAT | |||
|---|---|---|---|---|
| Correlation Coefficient (R) | p-Value | Correlation Coefficient (R) | p-Value | |
| Positive correlation | ||||
| LGR5 | 0.12 | 0.02 | 0.24 | 0.095 * |
| BMI1 | 0.13 | 0.031 | NS | |
| OCT4 | 0.16 | 0.0017 | NS | |
| MYC | 0.23 | <0.0001 | 0.3 | 0.0063 |
| USP36 | 0.46 | <0.0001 | NS | |
| RPTOR | 0.41 | <0.0001 | NS | |
| UBE2O | 0.49 | <0.0001 | 0.22 | 0.045 |
| JMJD6 | 0.44 | <0.0001 | NS | |
| SLCO4A1 | 0.38 | <0.0001 | NS | |
| UTP18 | 0.42 | <0.0001 | 0.28 | 0.08 * |
| E2F6 | 0.42 | <0.0001 | NS | |
| CD44 | NS | 0.29 | 0.062 * | |
| Inverse correlation | ||||
| CD166 | −0.13 | 0.01 | NS | |
| ABHD5 | −0.21 | 0.00034 | −0.3 | 0.053 * |
| PDCD4 | −0.24 | <0.0001 | NS | |
| GPA33 | −0.28 | <0.0001 | NS | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, L.; Li, H.-S.; Man, A.T.-K.; Chow, A.K.-M.; Foo, D.C.-C.; Lo, O.S.-H.; Pang, R.W.-C.; Law, W.-L. High Expression of a Cancer Stemness-Related Gene, Chromobox 8 (CBX8), in Normal Tissue Adjacent to the Tumor (NAT) Is Associated with Poor Prognosis of Colorectal Cancer Patients. Cells 2022, 11, 1852. https://doi.org/10.3390/cells11111852
Ng L, Li H-S, Man AT-K, Chow AK-M, Foo DC-C, Lo OS-H, Pang RW-C, Law W-L. High Expression of a Cancer Stemness-Related Gene, Chromobox 8 (CBX8), in Normal Tissue Adjacent to the Tumor (NAT) Is Associated with Poor Prognosis of Colorectal Cancer Patients. Cells. 2022; 11(11):1852. https://doi.org/10.3390/cells11111852
Chicago/Turabian StyleNg, Lui, Hung-Sing Li, Abraham Tak-Ka Man, Ariel Ka-Man Chow, Dominic Chi-Chung Foo, Oswens Siu-Hung Lo, Roberta Wen-Chi Pang, and Wai-Lun Law. 2022. "High Expression of a Cancer Stemness-Related Gene, Chromobox 8 (CBX8), in Normal Tissue Adjacent to the Tumor (NAT) Is Associated with Poor Prognosis of Colorectal Cancer Patients" Cells 11, no. 11: 1852. https://doi.org/10.3390/cells11111852
APA StyleNg, L., Li, H.-S., Man, A. T.-K., Chow, A. K.-M., Foo, D. C.-C., Lo, O. S.-H., Pang, R. W.-C., & Law, W.-L. (2022). High Expression of a Cancer Stemness-Related Gene, Chromobox 8 (CBX8), in Normal Tissue Adjacent to the Tumor (NAT) Is Associated with Poor Prognosis of Colorectal Cancer Patients. Cells, 11(11), 1852. https://doi.org/10.3390/cells11111852