
Poloxamer 407 Induces Hypertriglyceridemia but Decreases Atherosclerosis in Ldlr−/− Mice

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and P407 Injection
2.2. Plasma Lipoprotein Profiling
2.3. Antibodies and FACS Analysis of Circulating Monocytes
2.4. Cell Culture
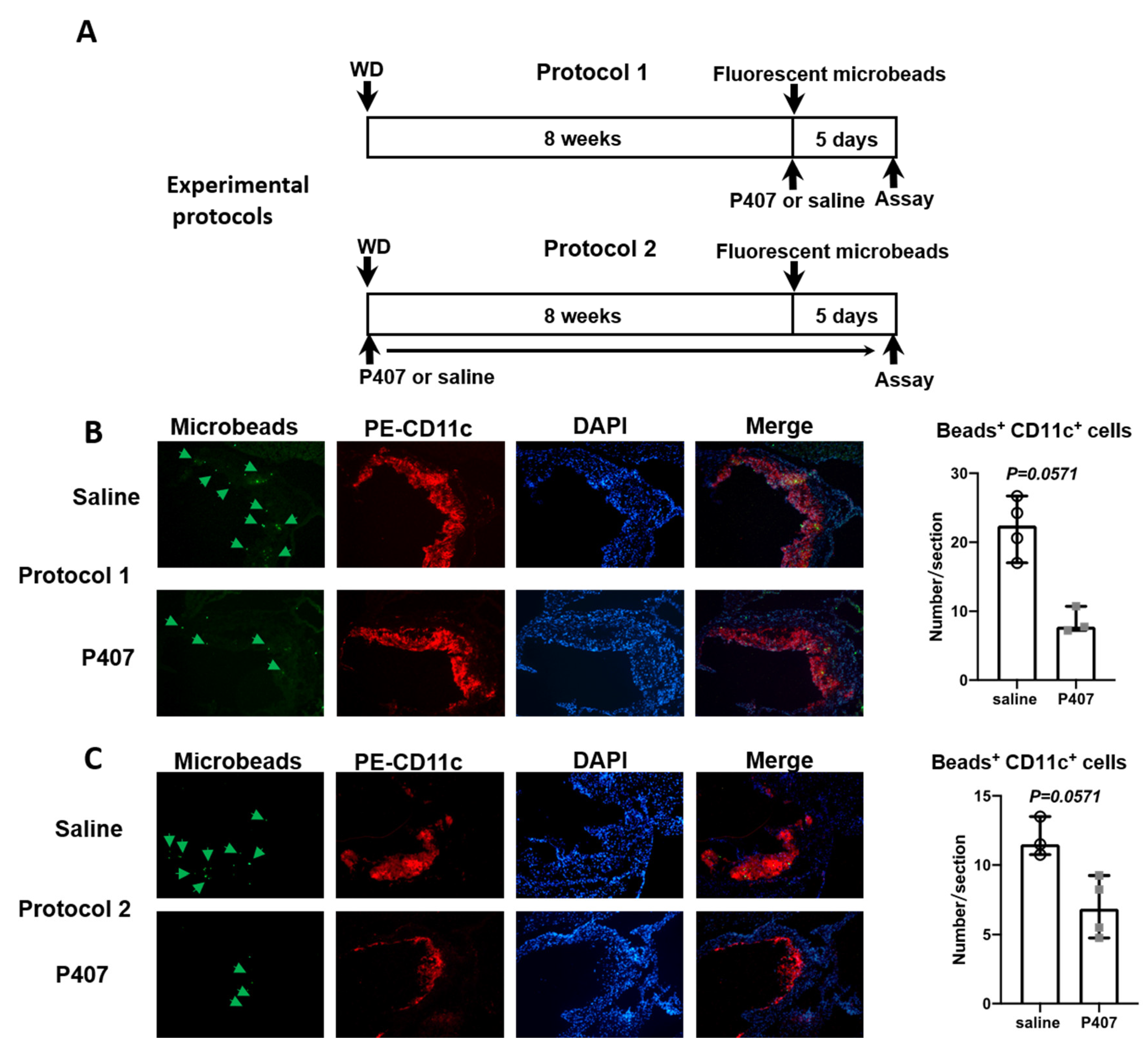
2.5. Analysis of Monocyte Trafficking into Atherosclerotic Lesions
2.6. Analysis of Atherosclerotic Lesions
2.7. Analysis of CD11c mRNA in the Liver
2.8. Statistical Analysis
3. Results
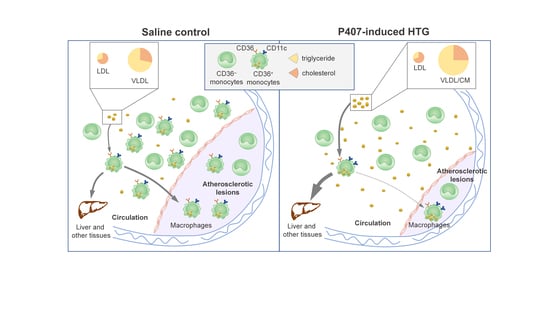
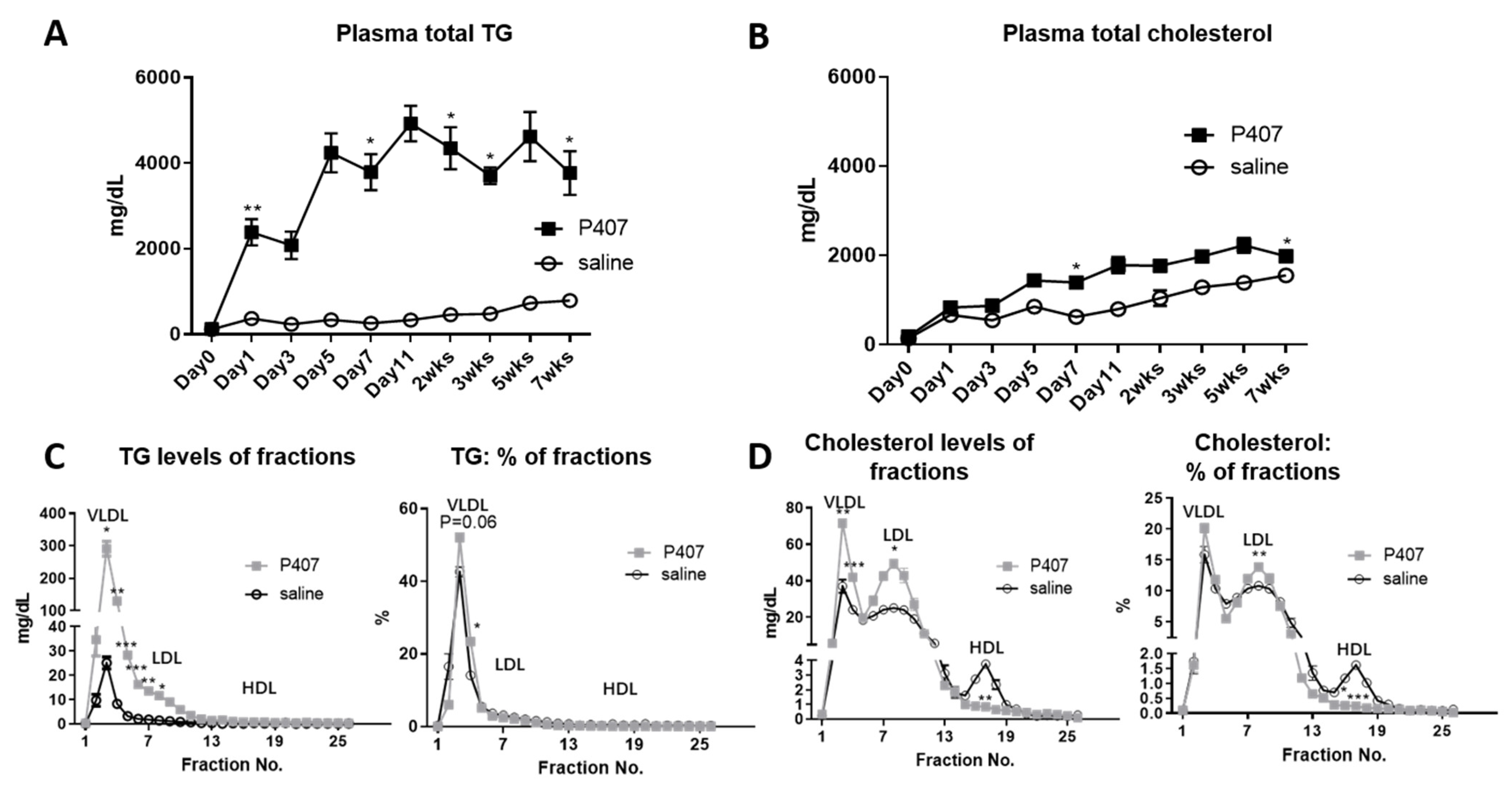
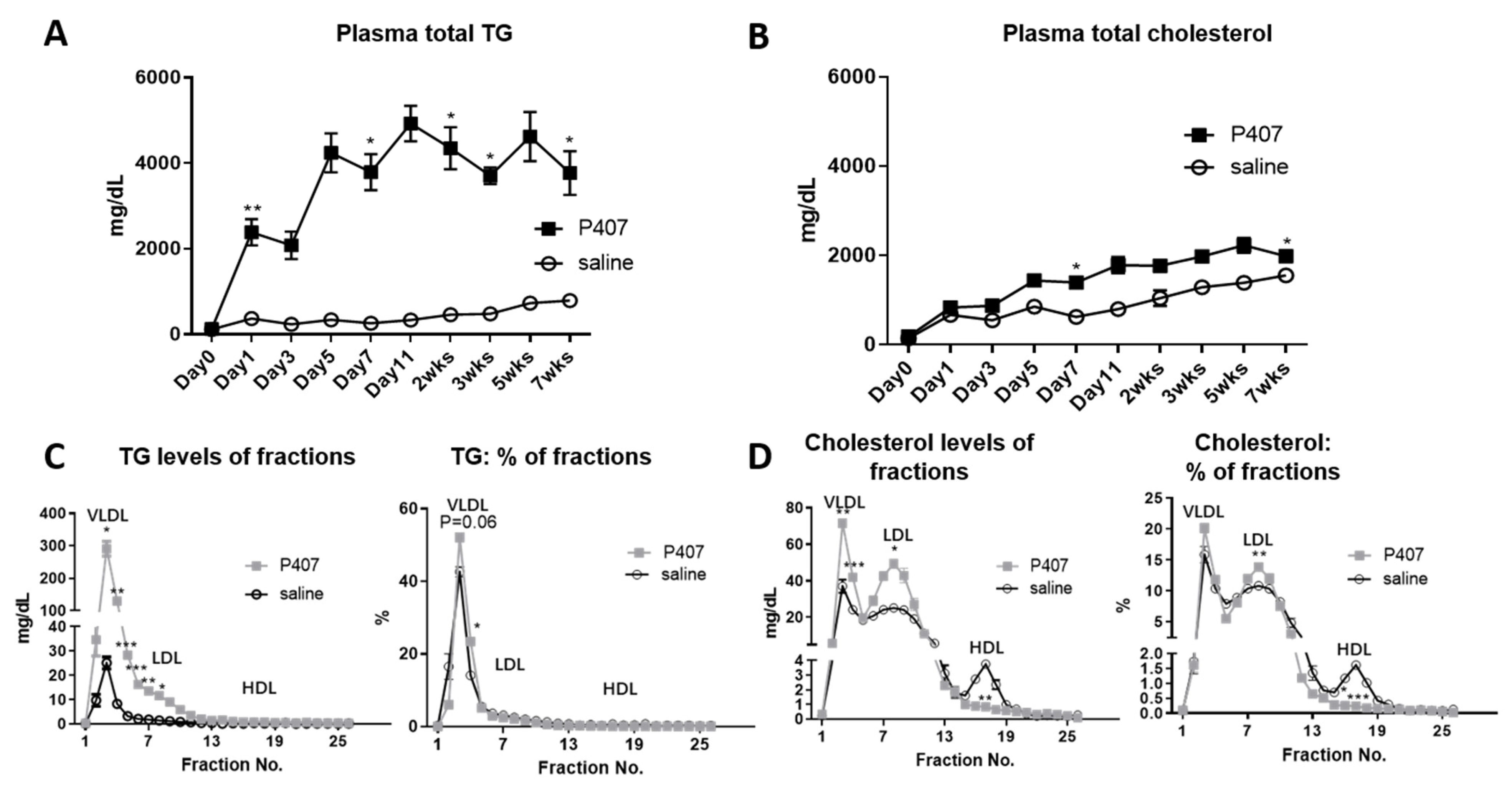
3.1. P407 Induced Rapid and Persistent Severe HTG in Ldlr−/− Mice Fed WD
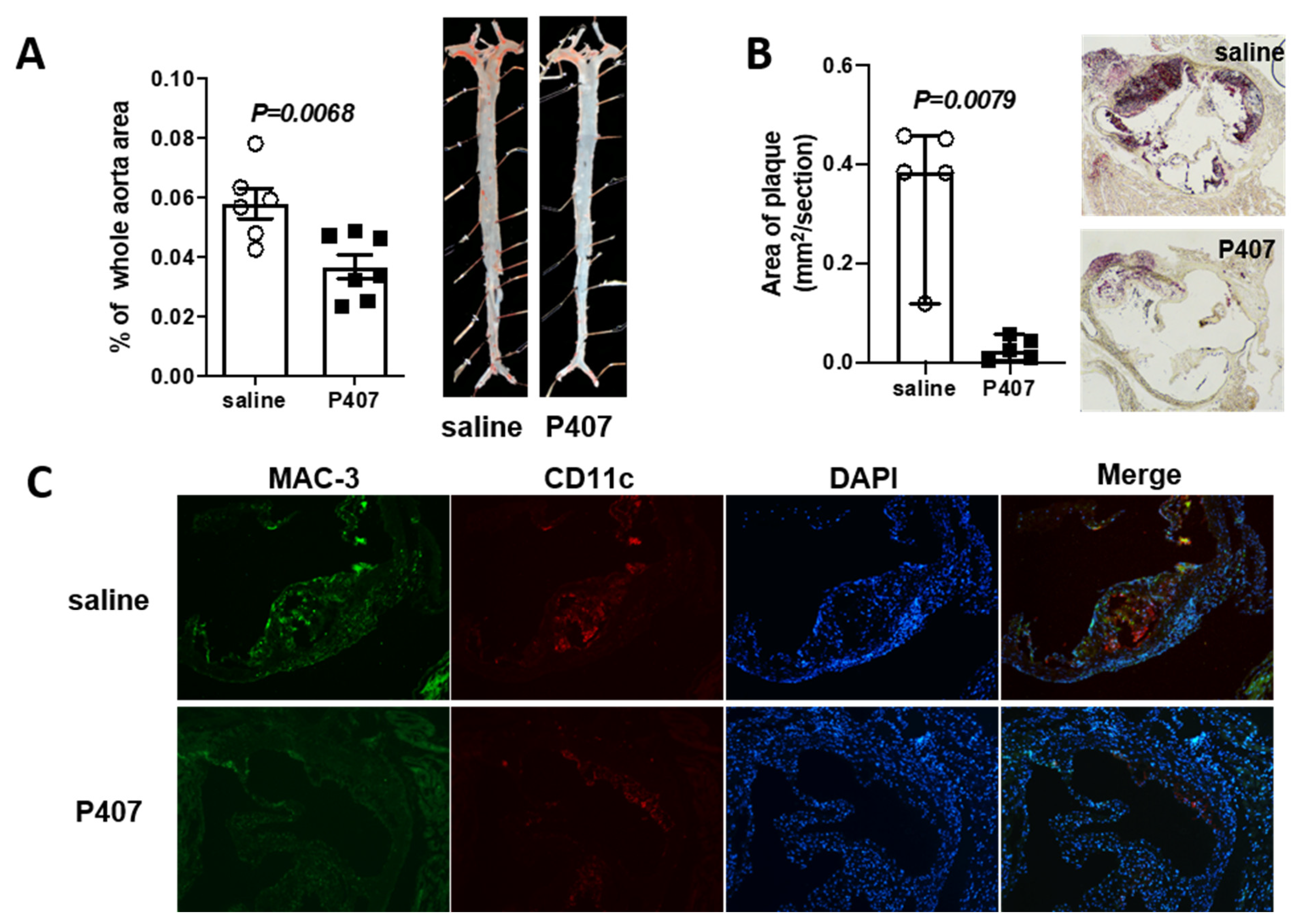
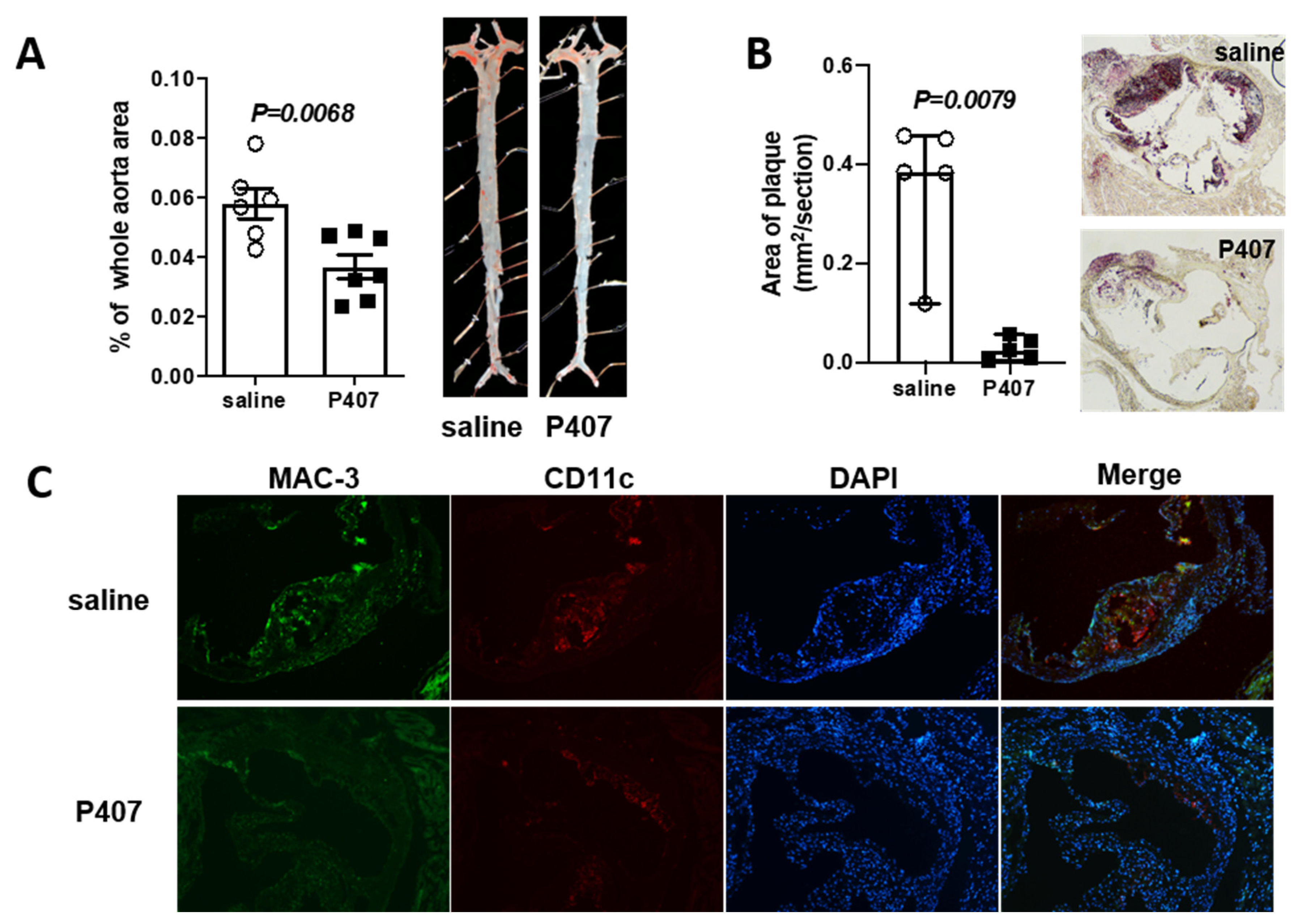
3.2. Ldlr−/− Mice with P407 Injection Had Reduced Atherosclerosis
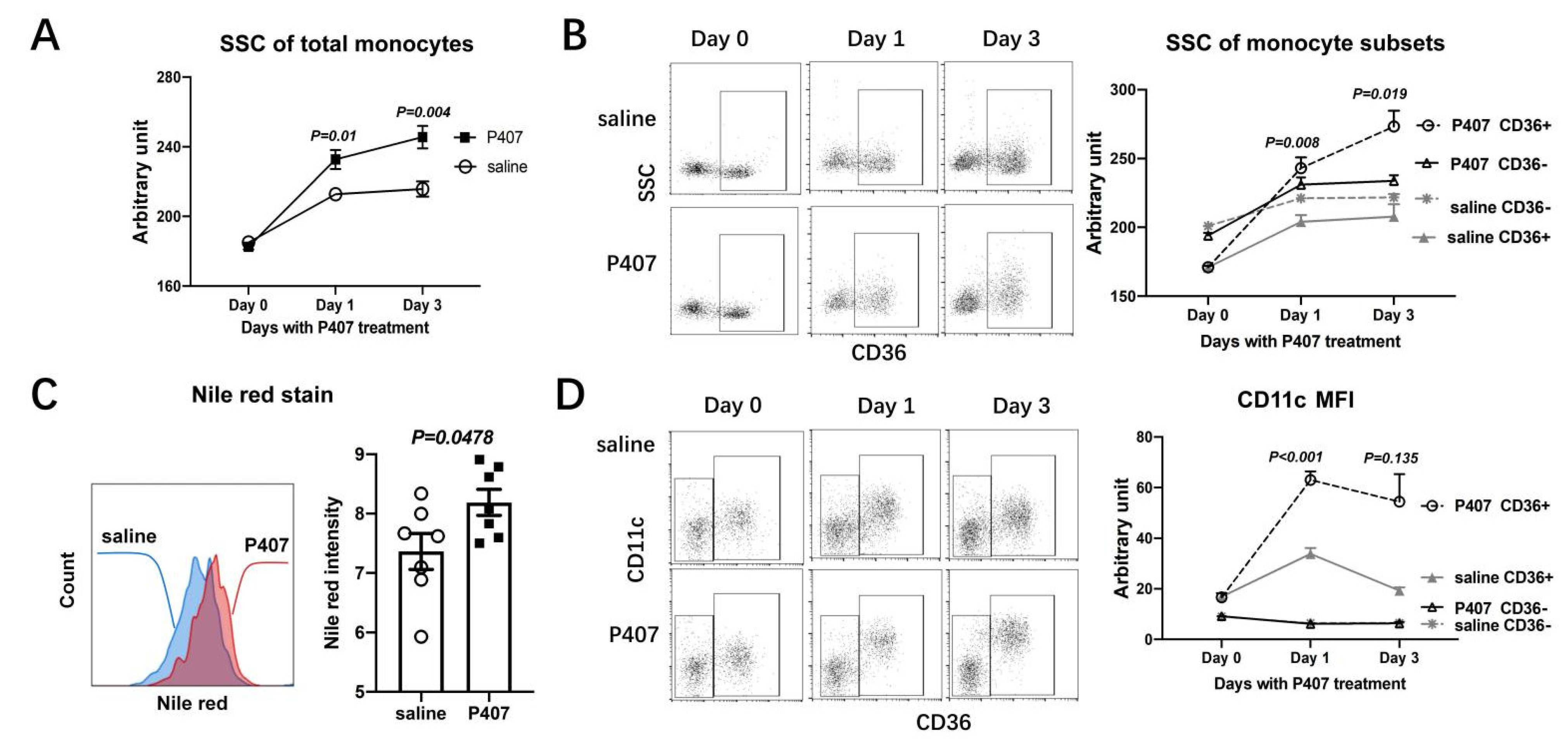
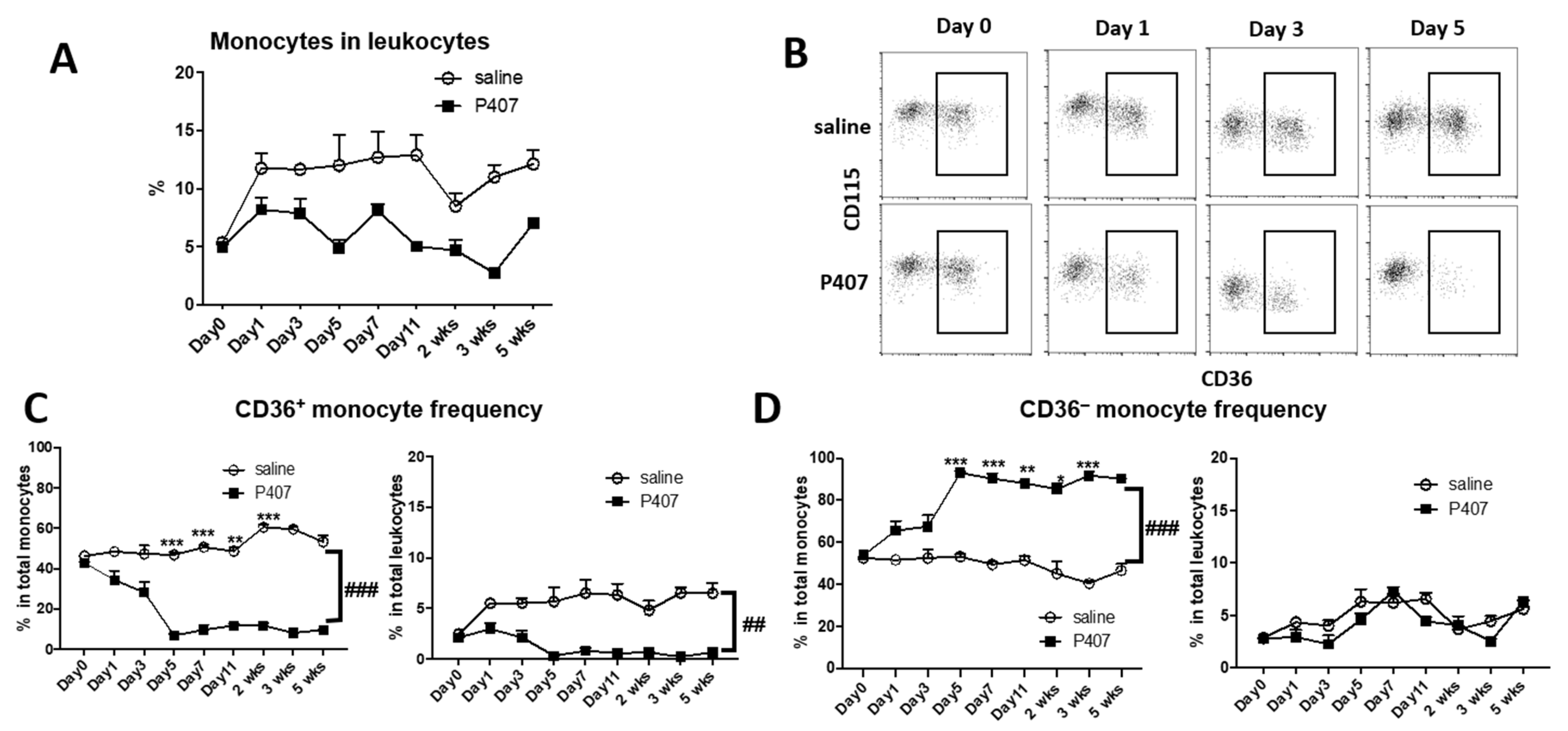
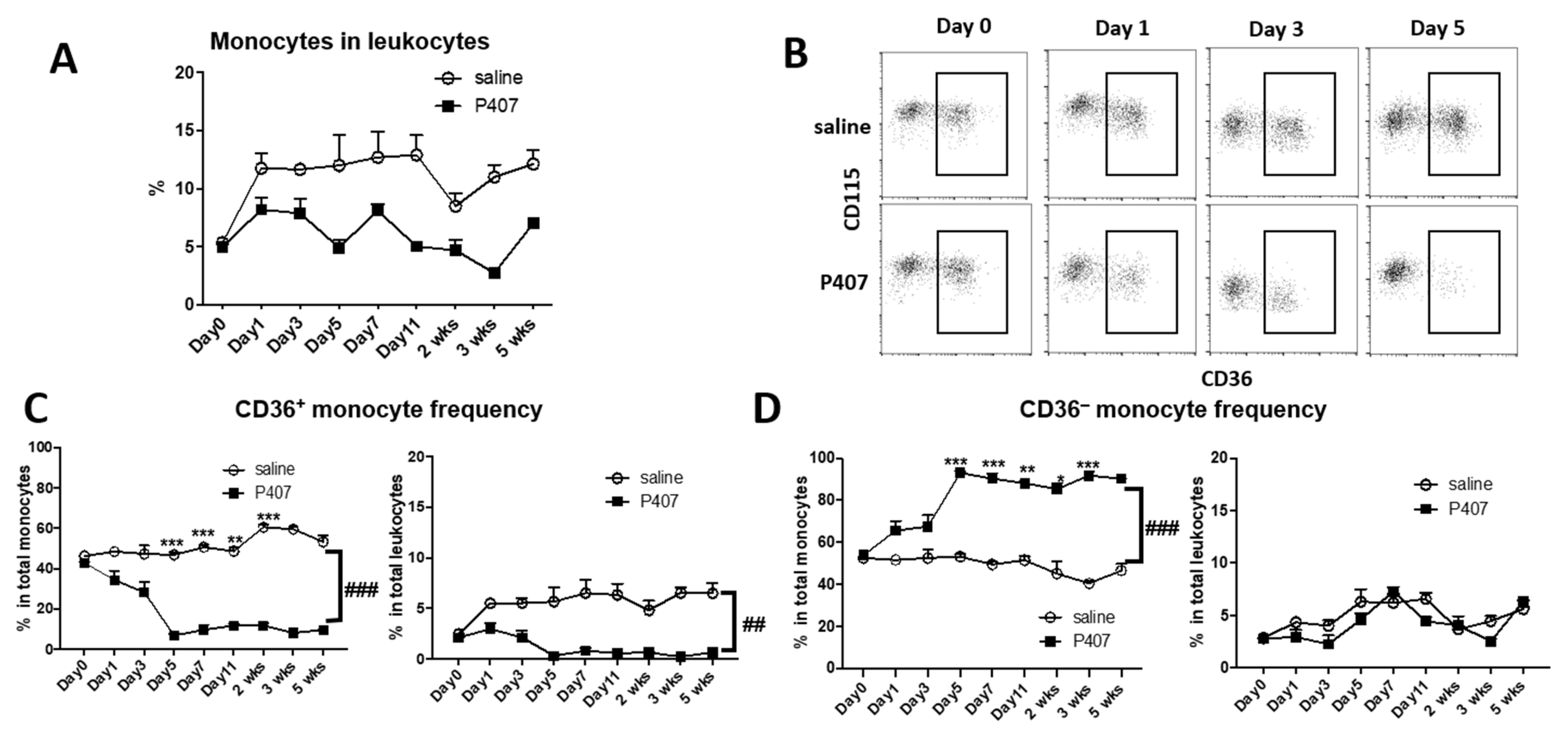
3.3. P407-Induced HTG Rapidly Increased Lipid Accumulation and CD11c Levels in CD36+ Monocytes in Ldlr−/− Mice
3.4. P407 Severely Reduced the Proportion of Circulating CD36+ Monocytes Early and Persistently in Ldlr−/− Mice
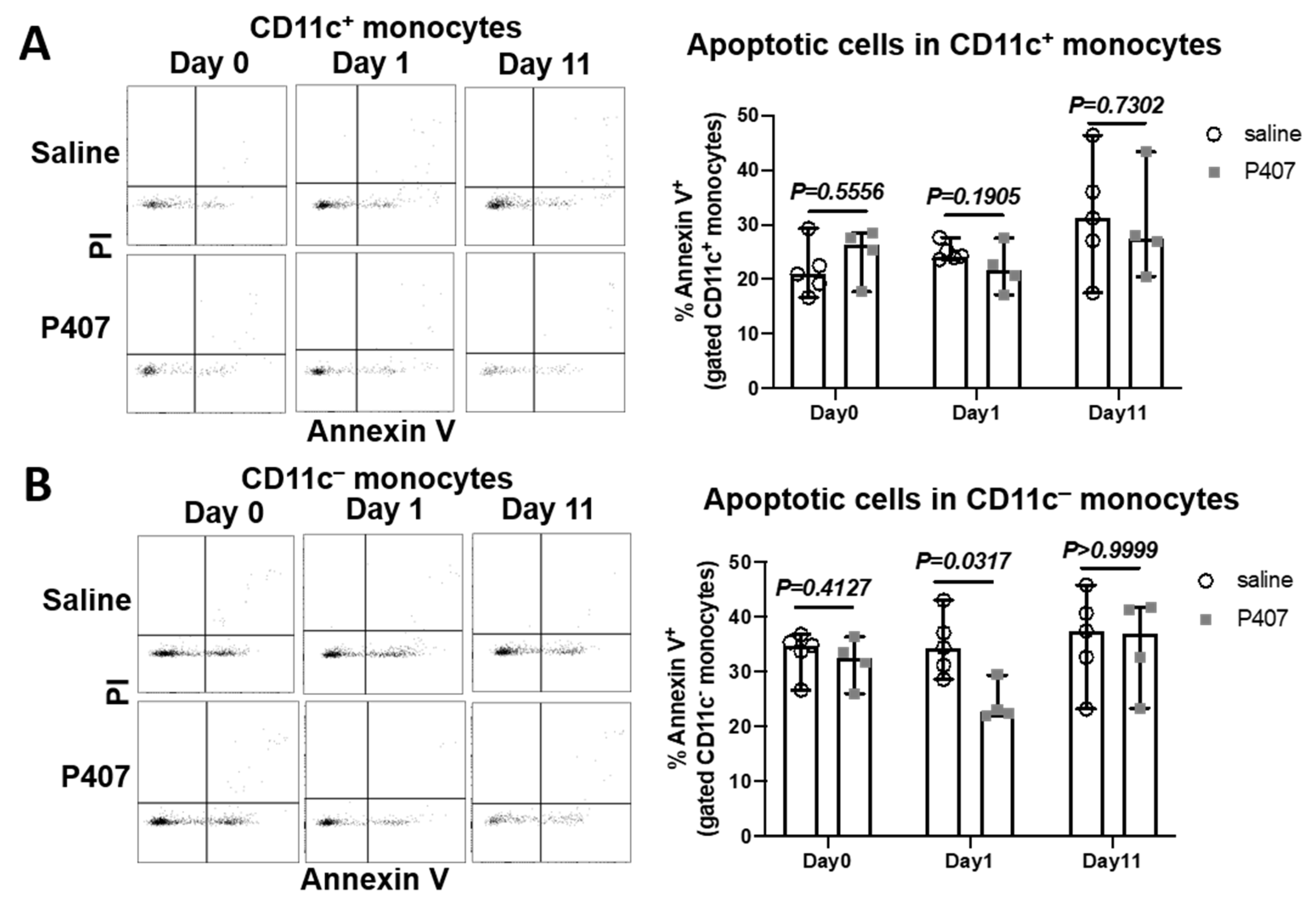
3.5. P407 Did Not Increase Monocyte Apoptosis in Ldlr−/− Mice
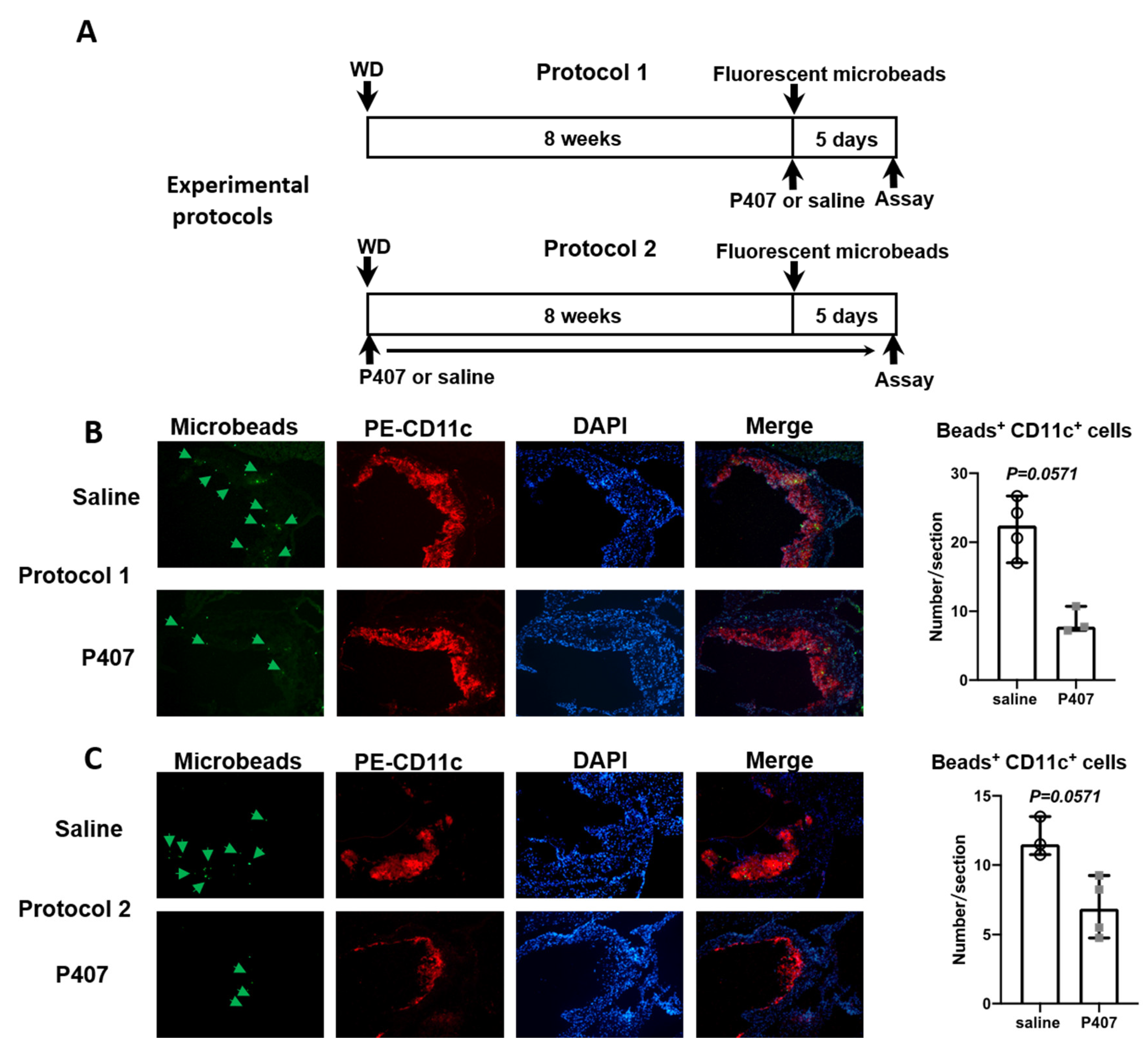
3.6. Infiltration of CD36+ Monocytes into Atherosclerotic Lesions in Ldlr−/− Mice with P407 Injection Was Reduced Early and Persistently
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Nonstandard Abbreviations and Acronyms
| ASCVD | atherosclerotic cardiovascular disease |
| BCA | brachiocephalic artery |
| CM | chylomicrons |
| HTG | hypertriglyceridemia |
| LDL | low-density lipoprotein |
| LDL-C | low-density lipoprotein cholesterol |
| LDL-TG | low-density lipoprotein–triglyceride |
| LPL | lipoprotein lipase |
| P407 | poloxamer 407 |
| PI | propidium iodide |
| SEC | size-exclusion chromatography |
| SSC | side scatter |
| TG | triglyceride |
| VLDL | very-low-density lipoprotein |
| WD | Western high-fat high-cholesterol diet |
| WT | wild-type |
References
- Ginsberg, H.N.; Packard, C.J.; Chapman, M.J.; Boren, J.; Aguilar-Salinas, C.A.; Averna, M.; Ference, B.A.; Gaudet, D.; Hegele, R.A.; Kersten, S.; et al. Triglyceride-rich lipoproteins and their remnants: Metabolic insights, role in atherosclerotic cardiovascular disease, and emerging therapeutic strategies—A consensus statement from the European Atherosclerosis Society. Eur. Heart J. 2021, 42, 4791–4806. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Wu, H. Inflammatory Links between Hypertriglyceridemia and Atherogenesis. Curr. Atheroscler. Rep. 2022, 24, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Reiner, Z. Hypertriglyceridaemia and risk of coronary artery disease. Nat. Rev. Cardiol. 2017, 14, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Gill, P.K.; Dron, J.S.; Hegele, R.A. Genetics of hypertriglyceridemia and atherosclerosis. Curr. Opin. Cardiol. 2021, 36, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Laufs, U.; Parhofer, K.G.; Ginsberg, H.N.; Hegele, R.A. Clinical review on triglycerides. Eur. Heart J. 2020, 41, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawler, P.R.; Kotrri, G.; Koh, M.; Goodman, S.G.; Farkouh, M.E.; Lee, D.S.; Austin, P.C.; Udell, J.A.; Ko, D.T. Real-world risk of cardiovascular outcomes associated with hypertriglyceridaemia among individuals with atherosclerotic cardiovascular disease and potential eligibility for emerging therapies. Eur. Heart J. 2020, 41, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Toth, P.P.; Philip, S.; Hull, M.; Granowitz, C. Association of Elevated Triglycerides With Increased Cardiovascular Risk and Direct Costs in Statin-Treated Patients. Mayo Clin. Proc. 2019, 94, 1670–1680. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Gower, R.M.; Wang, H.; Perrard, X.Y.; Ma, R.; Bullard, D.C.; Burns, A.R.; Paul, A.; Smith, C.W.; Simon, S.I.; et al. Functional role of CD11c+ monocytes in atherogenesis associated with hypercholesterolemia. Circulation 2009, 119, 2708–2717. [Google Scholar] [CrossRef] [Green Version]
- Swirski, F.K.; Libby, P.; Aikawa, E.; Alcaide, P.; Luscinskas, F.W.; Weissleder, R.; Pittet, M.J. Ly-6Chi monocytes dominate hypercholesterolemia-associated monocytosis and give rise to macrophages in atheromata. J. Clin. Investig. 2007, 117, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Tacke, F.; Alvarez, D.; Kaplan, T.J.; Jakubzick, C.; Spanbroek, R.; Llodra, J.; Garin, A.; Liu, J.; Mack, M.; van Rooijen, N.; et al. Monocyte subsets differentially employ CCR2, CCR5, and CX3CR1 to accumulate within atherosclerotic plaques. J. Clin. Investig. 2007, 117, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Combadière, C.; Potteaux, S.; Rodero, M.; Simon, T.; Pezard, A.; Esposito, B.; Merval, R.; Proudfoot, A.; Tedgui, A.; Mallat, Z. Combined inhibition of CCL2, CX3CR1, and CCR5 abrogates Ly6Chi and Ly6Clo monocytosis and almost abolishes atherosclerosis in hypercholesterolemic mice. Circulation 2008, 117, 1649–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Dai Perrard, X.; Perrard, J.L.; Yang, D.; Xiao, X.; Teng, B.B.; Simon, S.I.; Ballantyne, C.M.; Wu, H. Foamy Monocytes Form Early and Contribute to Nascent Atherosclerosis in Mice With Hypercholesterolemia. Arter. Thromb. Vasc. Biol. 2015, 35, 1787–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dresel, H.A.; Via, D.P.; Stohr, M.; Elchner, U.; Gnasso, A.; Postiglione, A.; Blin, N.; Augustin, J.; Schettler, G. Observations on leukocytes from patients with severe familial hypercholesterolemia. Arteriosclerosis 1986, 6, 259–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernelot Moens, S.J.; Neele, A.E.; Kroon, J.; van der Valk, F.M.; Van den Bossche, J.; Hoeksema, M.A.; Hoogeveen, R.M.; Schnitzler, J.G.; Baccara-Dinet, M.T.; Manvelian, G.; et al. PCSK9 monoclonal antibodies reverse the pro-inflammatory profile of monocytes in familial hypercholesterolaemia. Eur. Heart J. 2017, 38, 1584–1593. [Google Scholar] [CrossRef]
- Varela, L.M.; Ortega, A.; Bermudez, B.; Lopez, S.; Pacheco, Y.M.; Villar, J.; Abia, R.; Muriana, F.J. A high-fat meal promotes lipid-load and apolipoprotein B-48 receptor transcriptional activity in circulating monocytes. Am. J. Clin. Nutr. 2011, 93, 918–925. [Google Scholar] [CrossRef]
- Gower, R.M.; Wu, H.; Foster, G.A.; Devaraj, S.; Jialal, I.; Ballantyne, C.M.; Knowlton, A.A.; Simon, S.I. CD11c/CD18 expression is upregulated on blood monocytes during hypertriglyceridemia and enhances adhesion to vascular cell adhesion molecule-1. Arter. Thromb. Vasc. Biol. 2011, 31, 160–166. [Google Scholar] [CrossRef] [Green Version]
- den Hartigh, L.J.; Connolly-Rohrbach, J.E.; Fore, S.; Huser, T.R.; Rutledge, J.C. Fatty acids from very low-density lipoprotein lipolysis products induce lipid droplet accumulation in human monocytes. J. Immunol. 2010, 184, 3927–3936. [Google Scholar] [CrossRef] [Green Version]
- Bernelot Moens, S.J.; Verweij, S.L.; Schnitzler, J.G.; Stiekema, L.C.A.; Bos, M.; Langsted, A.; Kuijk, C.; Bekkering, S.; Voermans, C.; Verberne, H.J.; et al. Remnant Cholesterol Elicits Arterial Wall Inflammation and a Multilevel Cellular Immune Response in Humans. Arter. Thromb. Vasc. Biol. 2017, 37, 969–975. [Google Scholar] [CrossRef] [Green Version]
- Dai Perrard, X.Y.; Lian, Z.; Bobotas, G.; Dicklin, M.R.; Maki, K.C.; Wu, H. Effects of n-3 fatty acid treatment on monocyte phenotypes in humans with hypertriglyceridemia. J. Clin. Lipidol. 2017, 11, 1361–1371. [Google Scholar] [CrossRef]
- Khan, I.M.; Pokharel, Y.; Dadu, R.T.; Lewis, D.E.; Hoogeveen, R.C.; Wu, H.; Ballantyne, C.M. Postprandial Monocyte Activation in Individuals With Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2016, 101, 4195–4204. [Google Scholar] [CrossRef] [Green Version]
- Saja, M.F.; Baudino, L.; Jackson, W.D.; Cook, H.T.; Malik, T.H.; Fossati-Jimack, L.; Ruseva, M.; Pickering, M.C.; Woollard, K.J.; Botto, M. Triglyceride-Rich Lipoproteins Modulate the Distribution and Extravasation of Ly6C/Gr1(low) Monocytes. Cell Rep. 2015, 12, 1802–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, T.P. The P-407-induced murine model of dose-controlled hyperlipidemia and atherosclerosis: A review of findings to date. J. Cardiovasc. Pharm. 2004, 43, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Robinet, P.; Milewicz, D.M.; Cassis, L.A.; Leeper, N.J.; Lu, H.S.; Smith, J.D. Consideration of Sex Differences in Design and Reporting of Experimental Arterial Pathology Studies-Statement From ATVB Council. Arter. Thromb. Vasc. Biol. 2018, 38, 292–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourassa, P.A.; Milos, P.M.; Gaynor, B.J.; Breslow, J.L.; Aiello, R.J. Estrogen reduces atherosclerotic lesion development in apolipoprotein E-deficient mice. Proc. Natl. Acad. Sci. USA 1996, 93, 10022–10027. [Google Scholar] [CrossRef] [Green Version]
- Palmisano, B.T.; Zhu, L.; Stafford, J.M. Role of Estrogens in the Regulation of Liver Lipid Metabolism. Adv. Exp. Med. Biol. 2017, 1043, 227–256. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Yamauchi, J.; Lee, S.; Zhang, T.; Gong, Z.; Muzumdar, R.; Qu, S.; Dong, H.H. APOC3 Protein Is Not a Predisposing Factor for Fat-induced Nonalcoholic Fatty Liver Disease in Mice. J. Biol. Chem. 2017, 292, 3692–3705. [Google Scholar] [CrossRef] [Green Version]
- Lian, Z.; Perrard, X.D.; Peng, X.; Raya, J.L.; Hernandez, A.A.; Johnson, C.G.; Lagor, W.R.; Pownall, H.J.; Hoogeveen, R.C.; Simon, S.I.; et al. Replacing Saturated Fat With Unsaturated Fat in Western Diet Reduces Foamy Monocytes and Atherosclerosis in Male Ldlr(-/-) Mice. Arter. Thromb. Vasc. Biol. 2020, 40, 72–85. [Google Scholar] [CrossRef]
- Feig, J.E.; Parathath, S.; Rong, J.X.; Mick, S.L.; Vengrenyuk, Y.; Grauer, L.; Young, S.G.; Fisher, E.A. Reversal of hyperlipidemia with a genetic switch favorably affects the content and inflammatory state of macrophages in atherosclerotic plaques. Circulation 2011, 123, 989–998. [Google Scholar] [CrossRef] [Green Version]
- Daugherty, A.; Tall, A.R.; Daemen, M.; Falk, E.; Fisher, E.A.; García-Cardeña, G.; Lusis, A.J.; Owens, A.P., 3rd; Rosenfeld, M.E.; Virmani, R. Recommendation on Design, Execution, and Reporting of Animal Atherosclerosis Studies: A Scientific Statement From the American Heart Association. Arter. Thromb. Vasc. Biol. 2017, 37, e131–e157. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Fang, P.; Yu, D.; Zhang, L.; Zhang, D.; Jiang, X.; Yang, W.Y.; Bottiglieri, T.; Kunapuli, S.P.; Yu, J.; et al. Chronic Kidney Disease Induces Inflammatory CD40+ Monocyte Differentiation via Homocysteine Elevation and DNA Hypomethylation. Circ. Res. 2016, 119, 1226–1241. [Google Scholar] [CrossRef] [Green Version]
- Korolenko, T.A.; Johnston, T.P.; Tuzikov, F.V.; Tuzikova, N.A.; Pupyshev, A.B.; Spiridonov, V.K.; Goncharova, N.V.; Maiborodin, I.V.; Zhukova, N.A. Early-stage atherosclerosis in poloxamer 407-induced hyperlipidemic mice: Pathological features and changes in the lipid composition of serum lipoprotein fractions and subfractions. Lipids Health Dis. 2016, 15, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, T.P. Poloxamer 407 as a general lipase inhibitor: Its implications in lipid metabolism and atheroma formation in C57BL/6 mice. J. Pharm. Pharmacol. 2010, 62, 1807–1812. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Ishida, T.; Johnston, T.P.; Yasuda, T.; Ueyama, T.; Kojima, Y.; Kundu, R.K.; Quertermous, T.; Ishikawa, Y.; Hirata, K. Role of endothelial lipase in plasma HDL levels in a murine model of hypertriglyceridemia. J. Atheroscler. Thromb. 2009, 16, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Zernecke, A.; Winkels, H.; Cochain, C.; Williams, J.W.; Wolf, D.; Soehnlein, O.; Robbins, C.S.; Monaco, C.; Park, I.; McNamara, C.A.; et al. Meta-Analysis of Leukocyte Diversity in Atherosclerotic Mouse Aortas. Circ. Res. 2020, 127, 402–426. [Google Scholar] [CrossRef] [PubMed]
- Robbins, C.S.; Hilgendorf, I.; Weber, G.F.; Theurl, I.; Iwamoto, Y.; Figueiredo, J.L.; Gorbatov, R.; Sukhova, G.K.; Gerhardt, L.M.; Smyth, D.; et al. Local proliferation dominates lesional macrophage accumulation in atherosclerosis. Nat. Med. 2013, 19, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Yagyu, H.; Tazoe, F.; Nagashima, S.; Ohshiro, T.; Okada, K.; Osuga, J.; Goldberg, I.J.; Ishibashi, S. Macrophage lipoprotein lipase modulates the development of atherosclerosis but not adiposity. J. Lipid Res. 2013, 54, 1124–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masucci-Magoulas, L.; Goldberg, I.J.; Bisgaier, C.L.; Serajuddin, H.; Francone, O.L.; Breslow, J.L.; Tall, A.R. A mouse model with features of familial combined hyperlipidemia. Science 1997, 275, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qi, R.; Xian, X.; Yang, F.; Blackstein, M.; Deng, X.; Fan, J.; Ross, C.; Karasinska, J.; Hayden, M.R.; et al. Spontaneous atherosclerosis in aged lipoprotein lipase-deficient mice with severe hypertriglyceridemia on a normal chow diet. Circ. Res. 2008, 102, 250–256. [Google Scholar] [CrossRef]
- Zhang, D.; Jiang, X.; Fang, P.; Yan, Y.; Song, J.; Gupta, S.; Schafer, A.I.; Durante, W.; Kruger, W.D.; Yang, X.; et al. Hyperhomocysteinemia promotes inflammatory monocyte generation and accelerates atherosclerosis in transgenic cystathionine beta-synthase-deficient mice. Circulation 2009, 120, 1893–1902. [Google Scholar] [CrossRef]
- Clemente, C.; Rius, C.; Alonso-Herranz, L.; Martin-Alonso, M.; Pollan, A.; Camafeita, E.; Martinez, F.; Mota, R.A.; Nunez, V.; Rodriguez, C.; et al. MT4-MMP deficiency increases patrolling monocyte recruitment to early lesions and accelerates atherosclerosis. Nat. Commun. 2018, 9, 910. [Google Scholar] [CrossRef] [Green Version]
- Joffre, J.; Potteaux, S.; Zeboudj, L.; Loyer, X.; Boufenzer, A.; Laurans, L.; Esposito, B.; Vandestienne, M.; de Jager, S.C.; Henique, C.; et al. Genetic and Pharmacological Inhibition of TREM-1 Limits the Development of Experimental Atherosclerosis. J. Am. Coll. Cardiol. 2016, 68, 2776–2793. [Google Scholar] [CrossRef] [PubMed]
- Distel, E.; Barrett, T.J.; Chung, K.; Girgis, N.M.; Parathath, S.; Essau, C.C.; Murphy, A.J.; Moore, K.J.; Fisher, E.A. miR33 inhibition overcomes deleterious effects of diabetes mellitus on atherosclerosis plaque regression in mice. Circ. Res. 2014, 115, 759–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, T.J.; Distel, E.; Murphy, A.J.; Hu, J.; Garshick, M.S.; Ogando, Y.; Liu, J.; Vaisar, T.; Heinecke, J.W.; Berger, J.S.; et al. Apolipoprotein AI) Promotes Atherosclerosis Regression in Diabetic Mice by Suppressing Myelopoiesis and Plaque Inflammation. Circulation 2019, 140, 1170–1184. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.L.; Zhu, X.; Rong, S.; Shewale, S.; Seo, J.; Boudyguina, E.; Gebre, A.K.; Alexander-Miller, M.A.; Parks, J.S. Omega-3 fatty acids ameliorate atherosclerosis by favorably altering monocyte subsets and limiting monocyte recruitment to aortic lesions. Arter. Thromb. Vasc. Biol. 2012, 32, 2122–2130. [Google Scholar] [CrossRef] [Green Version]
- Foster, G.A.; Xu, L.; Chidambaram, A.A.; Soderberg, S.R.; Armstrong, E.J.; Wu, H.; Simon, S.I. CD11c/CD18 Signals Very Late Antigen-4 Activation To Initiate Foamy Monocyte Recruitment during the Onset of Hypercholesterolemia. J. Immunol. 2015, 195, 5380–5392. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.R.; Josefs, T.; Scerbo, D.; Gumaste, N.; Hu, Y.; Huggins, L.A.; Barrett, T.J.; Chiang, S.S.; Grossman, J.; Bagdasarov, S.; et al. Role of LpL (Lipoprotein Lipase) in Macrophage Polarization In Vitro and In Vivo. Arter. Thromb. Vasc. Biol. 2019, 39, 1967–1985. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, X.; Lian, Z.; Perrard, X.-Y.D.; Xiao, Y.; Ni, J.; O’Brien, V.; Dong, H.; Pownall, H.J.; Ballantyne, C.M.; Wu, H. Poloxamer 407 Induces Hypertriglyceridemia but Decreases Atherosclerosis in Ldlr−/− Mice. Cells 2022, 11, 1795. https://doi.org/10.3390/cells11111795
Peng X, Lian Z, Perrard X-YD, Xiao Y, Ni J, O’Brien V, Dong H, Pownall HJ, Ballantyne CM, Wu H. Poloxamer 407 Induces Hypertriglyceridemia but Decreases Atherosclerosis in Ldlr−/− Mice. Cells. 2022; 11(11):1795. https://doi.org/10.3390/cells11111795
Chicago/Turabian StylePeng, Xueying, Zeqin Lian, Xiao-Yuan Dai Perrard, Yunjie Xiao, Jing Ni, Veronica O’Brien, Henry Dong, Henry J. Pownall, Christie M. Ballantyne, and Huaizhu Wu. 2022. "Poloxamer 407 Induces Hypertriglyceridemia but Decreases Atherosclerosis in Ldlr−/− Mice" Cells 11, no. 11: 1795. https://doi.org/10.3390/cells11111795
APA StylePeng, X., Lian, Z., Perrard, X.-Y. D., Xiao, Y., Ni, J., O’Brien, V., Dong, H., Pownall, H. J., Ballantyne, C. M., & Wu, H. (2022). Poloxamer 407 Induces Hypertriglyceridemia but Decreases Atherosclerosis in Ldlr−/− Mice. Cells, 11(11), 1795. https://doi.org/10.3390/cells11111795