
Eosinophils in the Gastrointestinal Tract: Key Contributors to Neuro-Immune Crosstalk and Potential Implications in Disorders of Brain-Gut Interaction

 , , , , and
, , , , and
Abstract
:1. Introduction
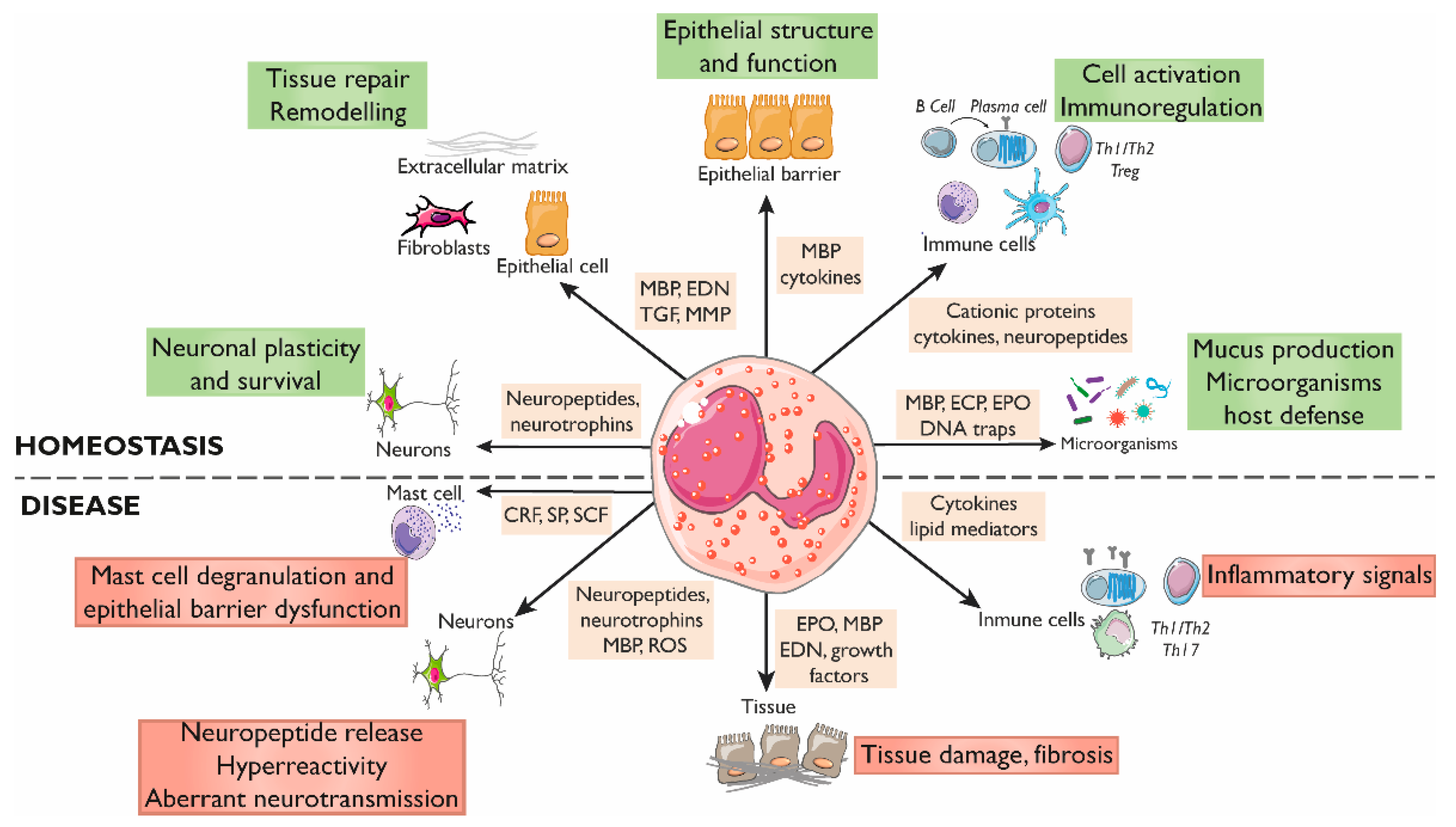
2. Eosinophils in the Gastrointestinal Tract
2.1. Eosinophils Regulate the Intestinal Barrier Function
2.2. Eosinophils Modulate Intestinal Immune Responses
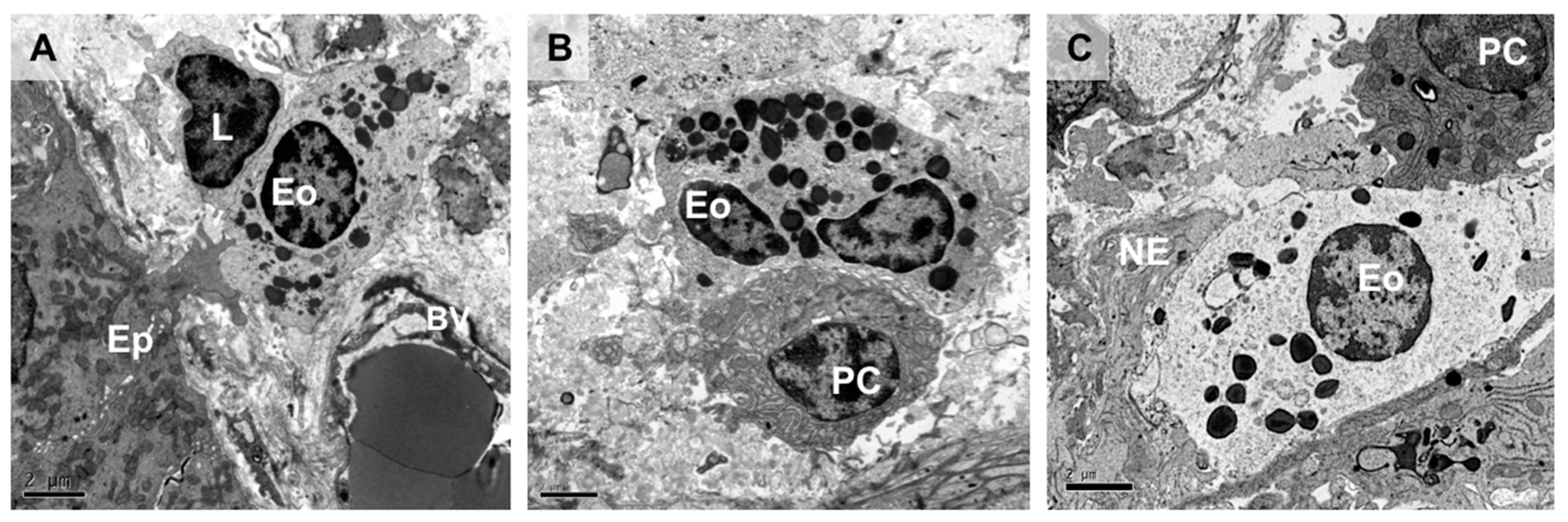
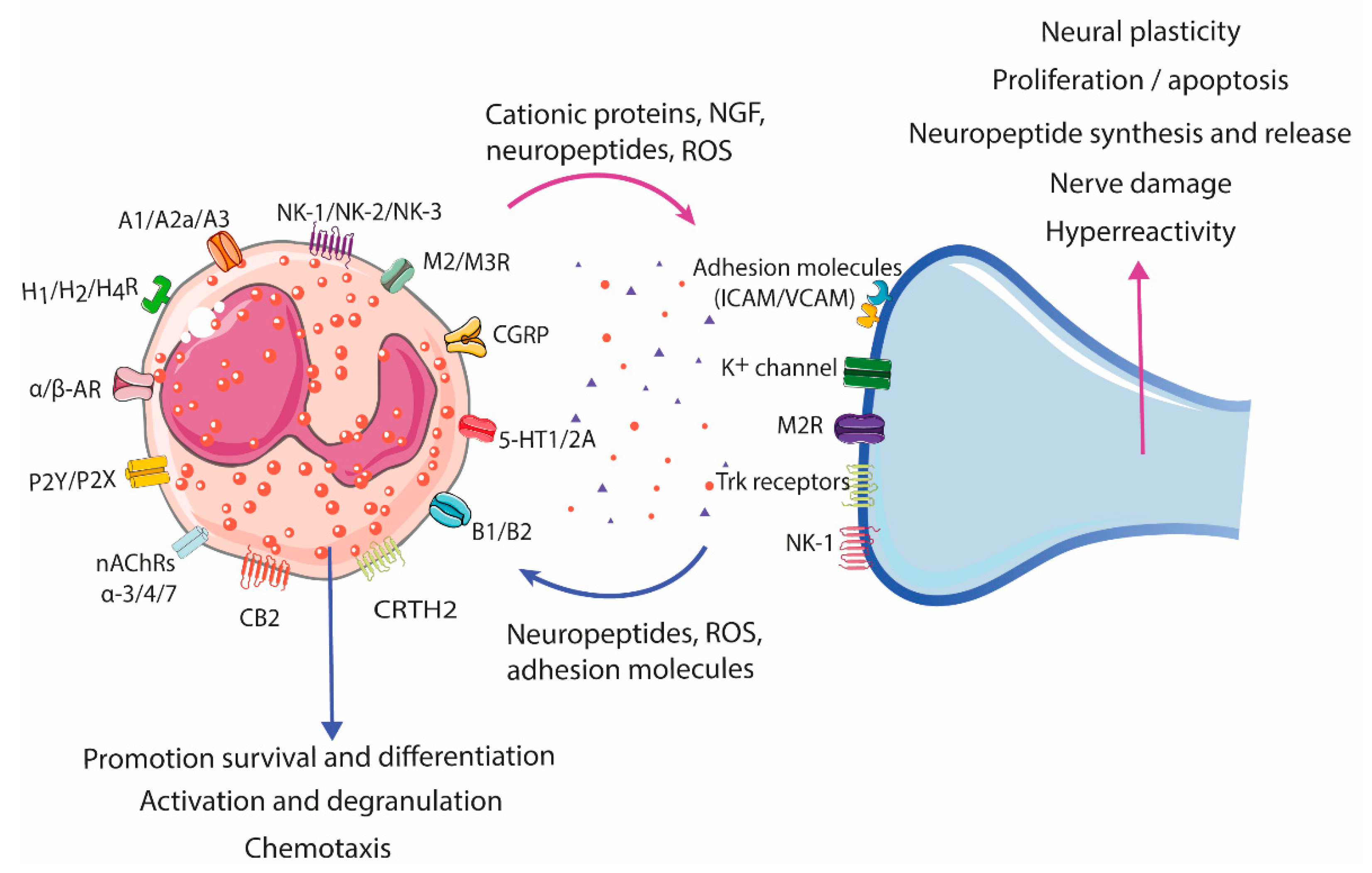
2.3. Eosinophil–Neuron Interactions
2.3.1. Innervation of the Gastrointestinal Tract
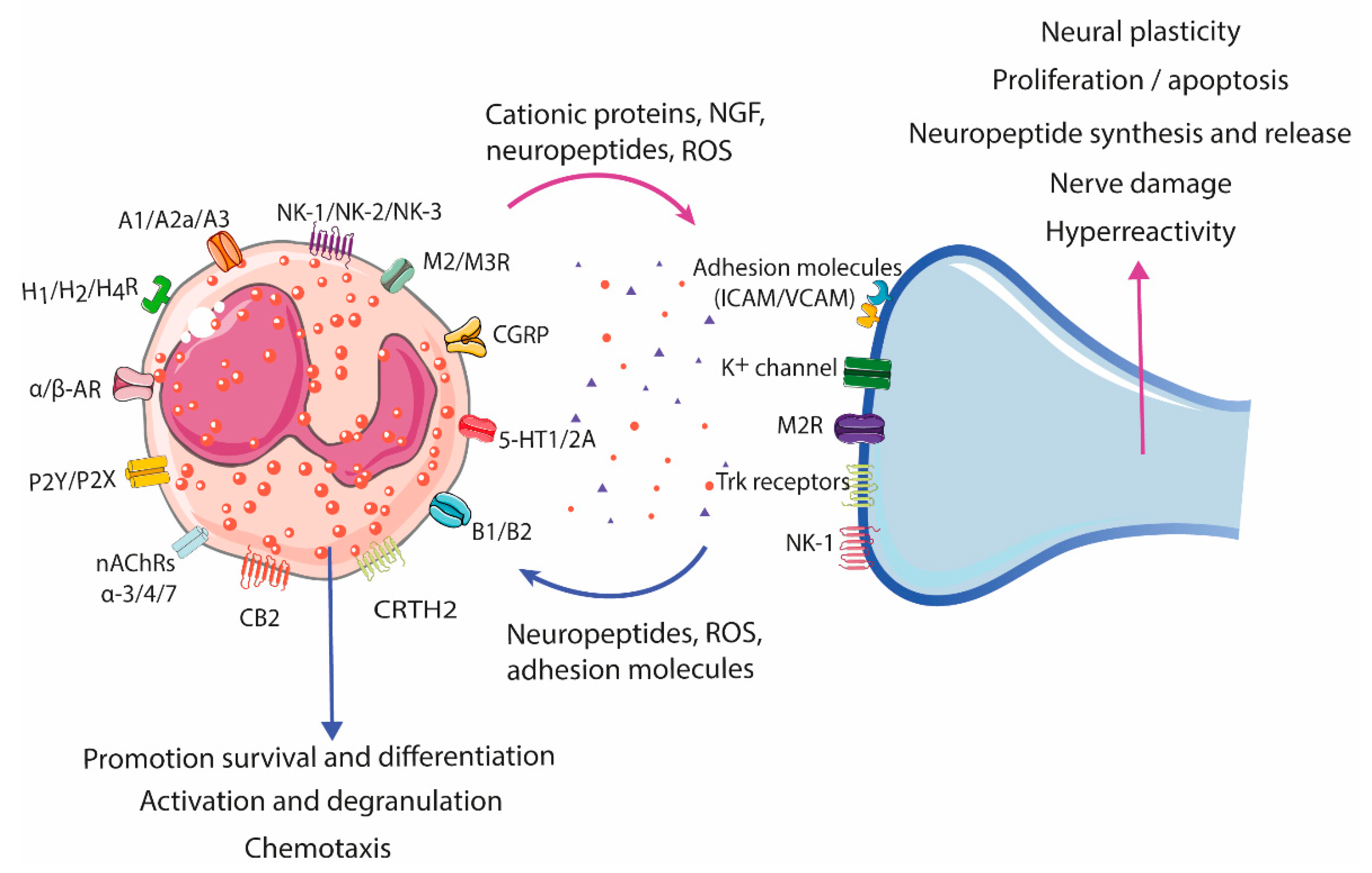
2.3.2. Neural-Induced Recruitment and Activation of Eosinophils by Extrinsic Nerves

{kind=link}
{kind=link}
{kind=link}
| Receptor 1 | Effect | References |
|---|---|---|
| Adenosine receptors A1, A2a, and A3 | Activation (A1), NADPH oxidase activity regulation (A2a), pro- or anti-inflammatory response (A3) | [64,65] |
| Adrenergic receptors (α1, α2), β1, β2, and β3 | Inhibition of NADPH oxidase, degranulation, chemotaxis, adhesion molecules, membrane lipid metabolism (β2). Inhibition of ICAM adhesion, ROS production, and EDN degranulation in IL-5, LTD4, and CXCL10 primed eosinophils | [64,66,67,68] |
| Bradykinin receptor B1, B2 | Proliferation, migration, and increase in intracellular Ca2+ levels, generation of lipid bodies and decreased eosinophil cell count in allergic airway inflammation (B1) Implications for eosinophil accumulation (B2) | [64,69,70,71] |
| Cannabinoid receptor CB2 | Chemotaxis, ICAM adhesion, increased eotaxin-2-primed CD11b expression, increased ROS production | [72,73,74] |
| Calcitonin gene-related peptide (CGRP) receptor | Increased migration | [60,64] |
| Histamine receptors H1R, H2R, H4R | Inhibition of ROS, EPO release, and chemotaxis (H2R) Priming of chemotaxis to eotaxin and adhesion to endothelium (H4R) | [75,76,77,78] |
| Muscarinic receptors M2, M3 | Stimulate production and release of CRF | [79] |
| Nicotinic acetylcholine receptors (nAChRs) α-3, -α4, and α-7 | Decrease infiltration into the lungs and airways Down-regulate eosinophil function in vitro | [80] |
| Purinergic receptors 2 P2Y and P2X family | Chemotaxis, induction of ROS production, CD11b upregulation, calcium mobilization, production of cytokines and ECP, induce release of EDN, EPO, and inflammatory factors | [81,82,83,84] |
| Serotonin receptor 5-HT1 (A, B, E), 5-HT2A | Migration(5-HT2A); effects on rolling and changes in shape of eosinophils | [85,86] |
| Tachykinin receptor NK1, NK2, and NK3 | Induction of the expression (NK1) and secretion (NK2) of CRF Increment of ROS production and thromboxane and degranulation of eosinophils | [64,87,88,89] |
| Vasointestinal peptide associated receptor CRTH2 | Chemokinesis or chemotaxis | [64,90,91] |
2.3.3. Neural-Induced Recruitment and Activation of Eosinophils by the ENS
2.3.4. Eosinophil-Dependent Neuroplasticity
3. Emerging Role of Eosinophils in Disorders of Brain–Gut Interaction
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rothenberg, M.E.; Hogan, S.P. The eosinophil. Annu. Rev. Immunol. 2006, 24, 147–174. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.J.; Jacobsen, E.A.; McGarry, M.P.; Schleimer, R.P.; Lee, N.A. Eosinophils in Health and Disease: The LIAR Hypothesis: Eosinophils and the LIAR Hypothesis. Clin. Exp. Allergy 2010, 40, 563–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, E.A.; Helmers, R.A.; Lee, J.J.; Lee, N.A. The Expanding Role(s) of Eosinophils in Health and Disease. Blood 2012, 120, 3882–3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurtner, A.; Gonzalez-Perez, I.; Arnold, I.C. Intestinal Eosinophils, Homeostasis and Response to Bacterial Intrusion. In Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2021; Volume 43, pp. 295–306. [Google Scholar] [CrossRef]
- Drake, M.G.; Lebold, K.M.; Roth-Carter, Q.R.; Pincus, A.B.; Blum, E.D.; Proskocil, B.J.; Jacoby, D.B.; Fryer, A.D.; Nie, Z. Eosinophil and Airway Nerve Interactions in Asthma. J. Leukoc. Biol. 2018, 104, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Chu, V.T.; Beller, A.; Rausch, S.; Strandmark, J.; Zänker, M.; Arbach, O.; Kruglov, A.; Berek, C. Eosinophils Promote Generation and Maintenance of Immunoglobulin-A-Expressing Plasma Cells and Contribute to Gut Immune Homeostasis. Immunity 2014, 40, 582–593. [Google Scholar] [CrossRef] [Green Version]
- O’Shea, K.M.; Aceves, S.S.; Dellon, E.S.; Gupta, S.K.; Spergel, J.M.; Furuta, G.T.; Rothenberg, M.E. Pathophysiology of Eosinophilic Esophagitis. Gastroenterology 2018, 154, 333–345. [Google Scholar] [CrossRef]
- Wechsler, M.E.; Munitz, A.; Ackerman, S.J.; Drake, M.G.; Jackson, D.J.; Wardlaw, A.J.; Dougan, S.K.; Berdnikovs, S.; Schleich, F.; Matucci, A.; et al. Eosinophils in Health and Disease: A State-of-the-Art Review. Mayo Clin. Proc. 2021, 96, 2694–2707. [Google Scholar] [CrossRef]
- Veiga-Fernandes, H.; Mucida, D. Neuro-Immune Interactions at Barrier Surfaces. Cell 2016, 165, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, A.; Yang, D.; Vella, M.; Chiu, I.M. The Intestinal Neuro-Immune Axis: Crosstalk between Neurons, Immune Cells, and Microbes. Mucosal Immunol. 2021, 14, 555–565. [Google Scholar] [CrossRef]
- Blanchard, C.; Rothenberg, M.E. Chapter 3 Biology of the Eosinophil. In Advances in Immunology; Elsevier: Amsterdam, The Netherlands, 2009; Volume 101, pp. 81–121. ISBN 978-0-12-374793-8. [Google Scholar]
- Mishra, A.; Hogan, S.P.; Lee, J.J.; Foster, P.S.; Rothenberg, M.E. Fundamental Signals That Regulate Eosinophil Homing to the Gastrointestinal Tract. J. Clin. Investig. 1999, 103, 1719–1727. [Google Scholar] [CrossRef] [Green Version]
- Masterson, J.C.; Menard-Katcher, C.; Larsen, L.D.; Furuta, G.T.; Spencer, L.A. Heterogeneity of Intestinal Tissue Eosinophils: Potential Considerations for Next-Generation Eosinophil-Targeting Strategies. Cells 2021, 10, 426. [Google Scholar] [CrossRef] [PubMed]
- Mesnil, C.; Raulier, S.; Paulissen, G.; Xiao, X.; Birrell, M.A.; Pirottin, D.; Janss, T.; Starkl, P.; Ramery, E.; Henket, M.; et al. Lung-Resident Eosinophils Represent a Distinct Regulatory Eosinophil Subset. J. Clin. Investig. 2016, 126, 3279–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdala Valencia, H.; Loffredo, L.F.; Misharin, A.V.; Berdnikovs, S. Phenotypic Plasticity and Targeting of Siglec-F high CD11c low Eosinophils to the Airway in a Murine Model of Asthma. Allergy 2016, 71, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Weller, P.F.; Spencer, L.A. Functions of Tissue-Resident Eosinophils. Nat. Rev. Immunol. 2017, 17, 746–760. [Google Scholar] [CrossRef] [PubMed]
- Brandt, E.B.; Zimmermann, N.; Muntel, E.E.; Yamada, Y.; Pope, S.M.; Mishra, A.; Hogan, S.P.; Rothenberg, M.E. The Alpha4bbeta7-Integrin Is Dynamically Expressed on Murine Eosinophils and Involved in Eosinophil Trafficking to the Intestine. Clin. Exp. Allergy 2006, 36, 543–553. [Google Scholar] [CrossRef]
- Forbes, E. ICAM-1-Dependent Pathways Regulate Colonic Eosinophilic Inflammation. J. Leukoc. Biol. 2006, 80, 330–341. [Google Scholar] [CrossRef]
- Carlens, J.; Wahl, B.; Ballmaier, M.; Bulfone-Paus, S.; Förster, R.; Pabst, O. Common γ-Chain-Dependent Signals Confer Selective Survival of Eosinophils in the Murine Small Intestine. J. Immunol. 2009, 183, 5600–5607. [Google Scholar] [CrossRef] [Green Version]
- Xenakis, J.J.; Howard, E.D.; Smith, K.M.; Olbrich, C.L.; Huang, Y.; Anketell, D.; Maldonado, S.; Cornwell, E.W.; Spencer, L.A. Resident Intestinal Eosinophils Constitutively Express Antigen Presentation Markers, and Include Two Phenotypically Distinct Subsets of Eosinophils. Immunology 2018, 154, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Chojnacki, A.; Wojcik, K.; Petri, B.; Aulakh, G.; Jacobsen, E.A.; LeSuer, W.E.; Colarusso, P.; Patel, K.D. Intravital Imaging Allows Real-Time Characterization of Tissue Resident Eosinophils. Commun. Biol. 2019, 2, 181. [Google Scholar] [CrossRef]
- Wouters, M.M.; Vicario, M.; Santos, J. The Role of Mast Cells in Functional GI Disorders. Gut 2016, 65, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Davoine, F.; Lacy, P. Eosinophil Cytokines, Chemokines, and Growth Factors: Emerging Roles in Immunity. Front. Immunol. 2014, 5, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loktionov, A. Eosinophils in the Gastrointestinal Tract and Their Role in the Pathogenesis of Major Colorectal Disorders. World J. Gastroenterol. 2019, 25, 3503–3526. [Google Scholar] [CrossRef] [PubMed]
- Coakley, G.; Wang, H.; Harris, N.L. Intestinal Eosinophils: Multifaceted Roles in Tissue Homeostasis and Disease. Semin. Immunopathol. 2021, 43, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Furuta, G.T.; Nieuwenhuis, E.E.S.; Karhausen, J.; Gleich, G.; Blumberg, R.S.; Lee, J.J.; Ackerman, S.J. Eosinophils Alter Colonic Epithelial Barrier Function: Role for Major Basic Protein. Am. J. Physiol.-Gastrointest. Liver Physiol. 2005, 289, G890–G897. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, D.B.; Ueki, I.F.; Widdicombe, J.H.; Loegering, D.A.; Gleich, G.J.; Nadel, J.A. Effect of Human Eosinophil Major Basic Protein on Ion Transport in Dog Tracheal Epithelium. Am. Rev. Respir. Dis. 1988, 137, 13–16. [Google Scholar] [CrossRef]
- White, S.R.; Sigrist, K.S.; Spaethe, S.M. Prostaglandin Secretion by Guinea Pig Tracheal Epithelial Cells Caused by Eosinophil Major Basic Protein. Am. J. Physiol.-Lung Cell. Mol. Physiol. 1993, 265, L234–L242. [Google Scholar] [CrossRef]
- Johnson, A.M.F.; Costanzo, A.; Gareau, M.G.; Armando, A.M.; Quehenberger, O.; Jameson, J.M.; Olefsky, J.M. High Fat Diet Causes Depletion of Intestinal Eosinophils Associated with Intestinal Permeability. PLoS ONE 2015, 10, e0122195. [Google Scholar] [CrossRef] [Green Version]
- Furuta, G.T.; Katzka, D.A. Eosinophilic Esophagitis. N. Engl. J. Med. 2015, 373, 1640–1648. [Google Scholar] [CrossRef] [Green Version]
- Honda, K.; Chihara, J. Eosinophil Activation by Eotaxin--Eotaxin Primes the Production of Reactive Oxygen Species from Eosinophils. Allergy 1999, 54, 1262–1269. [Google Scholar] [CrossRef]
- Choe, M.M.; Sporn, P.H.S.; Swartz, M.A. An in vitro Airway Wall Model of Remodeling. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2003, 285, L427–L433. [Google Scholar] [CrossRef] [Green Version]
- Sherrill, J.D.; Kc, K.; Wu, D.; Djukic, Z.; Caldwell, J.M.; Stucke, E.M.; Kemme, K.A.; Costello, M.S.; Mingler, M.K.; Blanchard, C.; et al. Desmoglein-1 Regulates Esophageal Epithelial Barrier Function and Immune Responses in Eosinophilic Esophagitis. Mucosal Immunol. 2014, 7, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Racca, F.; Pellegatta, G.; Cataldo, G.; Vespa, E.; Carlani, E.; Pelaia, C.; Paoletti, G.; Messina, M.R.; Nappi, E.; Canonica, G.W.; et al. Type 2 Inflammation in Eosinophilic Esophagitis: From Pathophysiology to Therapeutic Targets. Front. Physiol. 2021, 12, 815842. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Wen, T.; Mingler, M.K.; Caldwell, J.M.; Wang, Y.H.; Chaplin, D.D.; Lee, E.H.; Jang, M.H.; Woo, S.Y.; Seoh, J.Y.; et al. IL-1β in Eosinophil-Mediated Small Intestinal Homeostasis and IgA Production. Mucosal Immunol. 2015, 8, 930–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohn, L.; Homer, R.J.; MacLeod, H.; Mohrs, M.; Brombacher, F.; Bottomly, K. Th2-Induced Airway Mucus Production Is Dependent on IL-4Ralpha, but Not on Eosinophils. J. Immunol. 1999, 162, 6178–6183. [Google Scholar]
- Burgel, P.-R.; Lazarus, S.C.; Tam, D.C.-W.; Ueki, I.F.; Atabai, K.; Birch, M.; Nadel, J.A. Human Eosinophils Induce Mucin Production in Airway Epithelial Cells Via Epidermal Growth Factor Receptor Activation. J. Immunol. 2001, 167, 5948–5954. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.; Xu, F.; Zhang, G.; Wang, S.; Xu, W. CCR3 Monoclonal Antibody Inhibits Airway Eosinophilic Inflammation and Mucus Overproduction in a Mouse Model of Asthma. Acta Pharmacol. Sin. 2006, 27, 1594–1599. [Google Scholar] [CrossRef]
- Yousefi, S.; Gold, J.A.; Andina, N.; Lee, J.J.; Kelly, A.M.; Kozlowski, E.; Schmid, I.; Straumann, A.; Reichenbach, J.; Gleich, G.J.; et al. Catapult-like Release of Mitochondrial DNA by Eosinophils Contributes to Antibacterial Defense. Nat. Med. 2008, 14, 949–953. [Google Scholar] [CrossRef]
- Mukherjee, M.; Lacy, P.; Ueki, S. Eosinophil Extracellular Traps and Inflammatory Pathologies—Untangling the Web! Front. Immunol. 2018, 9, 2763. [Google Scholar] [CrossRef]
- Puxeddu, I.; Ribatti, D.; Crivellato, E.; Levi-Schaffer, F. Mast Cells and Eosinophils: A Novel Link between Inflammation and Angiogenesis in Allergic Diseases. J. Allergy Clin. Immunol. 2005, 116, 531–536. [Google Scholar] [CrossRef]
- Gomes, I.; Mathur, S.K.; Espenshade, B.M.; Mori, Y.; Varga, J.; Ackerman, S.J. Eosinophil-Fibroblast Interactions Induce Fibroblast IL-6 Secretion and Extracellular Matrix Gene Expression: Implications in Fibrogenesis. J. Allergy Clin. Immunol. 2005, 116, 796–804. [Google Scholar] [CrossRef]
- Temkin, V.; Aingorn, H.; Puxeddu, I.; Goldshmidt, O.; Zcharia, E.; Gleich, G.J.; Vlodavsky, I.; Levi-Schaffer, F. Eosinophil Major Basic Protein: First Identified Natural Heparanase-Inhibiting Protein. J. Allergy Clin. Immunol. 2004, 113, 703–709. [Google Scholar] [CrossRef] [PubMed]
- Shinkai, K.; Mohrs, M.; Locksley, R.M. Helper T Cells Regulate Type-2 Innate Immunity in vivo. Nature 2002, 420, 825–829. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Angkasekwinai, P.; Lu, N.; Voo, K.S.; Arima, K.; Hanabuchi, S.; Hippe, A.; Corrigan, C.J.; Dong, C.; Homey, B.; et al. IL-25 Augments Type 2 Immune Responses by Enhancing the Expansion and Functions of TSLP-DC–Activated Th2 Memory Cells. J. Exp. Med. 2007, 204, 1837–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, L.A.; Szela, C.T.; Perez, S.A.C.; Kirchhoffer, C.L.; Neves, J.S.; Radke, A.L.; Weller, P.F. Human Eosinophils Constitutively Express Multiple Th1, Th2, and Immunoregulatory Cytokines That Are Secreted Rapidly and Differentially. J. Leukoc. Biol. 2008, 85, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Akuthota, P.; Wang, H.B.; Spencer, L.A.; Weller, P.F. Immunoregulatory Roles of Eosinophils: A New Look at a Familiar Cell. Clin. Exp. Allergy 2008, 38, 1254–1263. [Google Scholar] [CrossRef] [PubMed]
- Farhan, R.K.; Vickers, M.A.; Ghaemmaghami, A.M.; Hall, A.M.; Barker, R.N.; Walsh, G.M. Effective Antigen Presentation to Helper T Cells by Human Eosinophils. Immunology 2016, 149, 413–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, K.M.; Rahman, R.S.; Spencer, L.A. Humoral Immunity Provides Resident Intestinal Eosinophils Access to Luminal Antigen via Eosinophil-Expressed Low-Affinity Fcγ Receptors. J. Immunol. 2016, 197, 3716–3724. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.K.; Jimenez-Saiz, R.; Verschoor, C.P.; Walker, T.D.; Goncharova, S.; Llop-Guevara, A.; Shen, P.; Gordon, M.E.; Barra, N.G.; Bassett, J.D.; et al. Indigenous Enteric Eosinophils Control DCs to Initiate a Primary Th2 Immune Response in vivo. J. Exp. Med. 2014, 211, 1657–1672. [Google Scholar] [CrossRef]
- Wang, H.-B.; Weller, P.F. Pivotal Advance: Eosinophils Mediate Early Alum Adjuvant-Elicited B Cell Priming and IgM Production. J. Leukoc. Biol. 2008, 83, 817–821. [Google Scholar] [CrossRef] [Green Version]
- Shakoory, B.; Fitzgerald, S.M.; Lee, S.A.; Chi, D.S.; Krishnaswamy, G. The Role of Human Mast Cell-Derived Cytokines in Eosinophil Biology. J. Interferon Cytokine Res. 2004, 24, 271–281. [Google Scholar] [CrossRef]
- Galdiero, M.R.; Varricchi, G.; Seaf, M.; Marone, G.; Levi-Schaffer, F.; Marone, G. Bidirectional Mast Cell–Eosinophil Interactions in Inflammatory Disorders and Cancer. Front. Med. 2017, 4, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niranjan, R.; Mavi, P.; Rayapudi, M.; Dynda, S.; Mishra, A. Pathogenic Role of Mast Cells in Experimental Eosinophilic Esophagitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G1087–G1094. [Google Scholar] [CrossRef] [PubMed]
- Hirota, C.L.; McKay, D.M. Cholinergic Regulation of Epithelial Ion Transport in the Mammalian Intestine. Br. J. Pharmacol. 2006, 149, 463–479. [Google Scholar] [CrossRef]
- Brierley, S.M.; Linden, D.R. Neuroplasticity and Dysfunction after Gastrointestinal Inflammation. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.J.; ten Hove, A.S.; Vervoordeldonk, M.J.; Luyer, M.D.; de Jonge, W.J. Neuroimmune Interactions in the Gut and Their Significance for Intestinal Immunity. Cells 2019, 8, 670. [Google Scholar] [CrossRef] [Green Version]
- Chou, D.L.; Daugherty, B.L.; McKenna, E.K.; Hsu, W.M.; Tyler, N.K.; Plopper, C.G.; Hyde, D.M.; Schelegle, E.S.; Gershwin, L.J.; Miller, L.A. Chronic Aeroallergen during Infancy Enhances Eotaxin-3 Expression in Airway Epithelium and Nerves. Am. J. Respir. Cell Mol. Biol. 2005, 33, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Fryer, A.D. Neuronal Eotaxin and the Effects of Ccr3 Antagonist on Airway Hyperreactivity and M2 Receptor Dysfunction. J. Clin. Investig. 2005, 116, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Numao, T.; Agrawal, D.K. Neuropeptides Modulate Human Eosinophil Chemotaxis. J. Immunol. 1992, 149, 3309–3315. [Google Scholar]
- Jacoby, D.B.; Costello, R.M.; Fryer, A.D. Eosinophil Recruitment to the Airway Nerves. J. Allergy Clin. Immunol. 2001, 107, 211–218. [Google Scholar] [CrossRef]
- Dunzendorfer, S.; Meierhofer, C.; Wiedermann, C.J. Signaling in Neuropeptide-Induced Migration of Human Eosinophils. J. Leukoc. Biol. 1998, 64, 828–834. [Google Scholar] [CrossRef]
- Kingham, P.J.; McLean, W.G.; Walsh, M.-T.; Fryer, A.D.; Gleich, G.J.; Costello, R.W. Effects of Eosinophils on Nerve Cell Morphology and Development: The Role of Reactive Oxygen Species and P38 MAP Kinase. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2003, 285, L915–L924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giembycz, M.A.; Lindsay, M.A. Pharmacology of the Eosinophil. Pharmacol. Rev. 1999, 51, 213–340. [Google Scholar] [PubMed]
- Brown, R.A.; Spina, D.; Page, C.P. Adenosine Receptors and Asthma: Adenosine Receptors and Asthma. Br. J. Pharmacol. 2008, 153, S446–S456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kankaanranta, H.; Parkkonen, J.; Ilmarinen-Salo, P.; Giembycz, M.A.; Moilanen, E. Salbutamol Delays Human Eosinophil Apoptosis via a CAMP-Dependent Mechanism. Pulm. Pharmacol. Ther. 2011, 24, 394–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, T.; Nakagome, K.; Kobayashi, T.; Ueda, Y.; Soma, T.; Nakamoto, H.; Nagata, M. Effect of Beta2-Adrenergic Agonists on Eosinophil Adhesion, Superoxide Anion Generation, and Degranulation. Allergol. Int. 2015, 64, S46–S53. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Huang, S.; Li, F.; Wu, M.; He, J.; Xue, Y.; Fu, T.; Yu, R.; Chen, X.; Wang, Y.; et al. Sympathetic Nerves Positively Regulate Eosinophil-Driven Allergic Conjunctivitis via A1-Adrenergic Receptor Signaling. Am. J. Pathol. 2020, 190, 1298–1308. [Google Scholar] [CrossRef]
- Vasquez-Pinto, L.M.C.; Nantel, F.; Sirois, P.; Jancar, S. Bradykinin B1 Receptor Antagonist R954 Inhibits Eosinophil Activation/Proliferation/Migration and Increases TGF-β and VEGF in a Murine Model of Asthma. Neuropeptides 2010, 44, 107–113. [Google Scholar] [CrossRef]
- Eric, J.; Bkaily, G.; Bkaily, G.B.; Volkov, L.; Gabra, B.H.; Sirois, P. Des-Arg9-Bradykinin Increases Intracellular Ca2+ in Bronchoalveolar Eosinophils from Ovalbumin-Sensitized and -Challenged Mice. Eur. J. Pharmacol. 2003, 475, 129–137. [Google Scholar] [CrossRef]
- Silva, A.R.; Larangeira, A.P.; Pacheco, P.; Calixto, J.B.; Henriques, M.G.M.O.; Bozza, P.T.; Castro-Faria-Neto, H.C. Bradykinin Down-Regulates LPS-Induced Eosinophil Accumulation in the Pleural Cavity of Mice through Type 2-Kinin Receptor Activation: A Role for Prostaglandins: Bradykinin Modulates Eosinophil Recruitment by LPS. Br. J. Pharmacol. 1999, 127, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, S.; Oka, S.; Gokoh, M.; Sugiura, T. Chemotaxis of Human Peripheral Blood Eosinophils to 2-Arachidonoylglycerol: Comparison with Other Eosinophil Chemoattractants. Int. Arch. Allergy Immunol. 2006, 140, 3–7. [Google Scholar] [CrossRef]
- Oka, S.; Ikeda, S.; Kishimoto, S.; Gokoh, M.; Yanagimoto, S.; Waku, K.; Sugiura, T. 2-Arachidonoylglycerol, an Endogenous Cannabinoid Receptor Ligand, Induces the Migration of EoL-1 Human Eosinophilic Leukemia Cells and Human Peripheral Blood Eosinophils. J. Leukoc. Biol. 2004, 76, 1002–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frei, R.B.; Luschnig, P.; Parzmair, G.P.; Peinhaupt, M.; Schranz, S.; Fauland, A.; Wheelock, C.E.; Heinemann, A.; Sturm, E.M. Cannabinoid Receptor 2 Augments Eosinophil Responsiveness and Aggravates Allergen-induced Pulmonary Inflammation in Mice. Allergy 2016, 71, 944–956. [Google Scholar] [CrossRef] [PubMed]
- Reher, T.M.; Brunskole, I.; Neumann, D.; Seifert, R. Evidence for Ligand-Specific Conformations of the Histamine H2-Receptor in Human Eosinophils and Neutrophils. Biochem. Pharmacol. 2012, 84, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- Ezeamuzie, C.I.; Philips, E. Histamine H 2 Receptors Mediate the Inhibitory Effect of Histamine on Human Eosinophil Degranulation: Histamine H 2 Receptors and Eosinophil Degranulation. Br. J. Pharmacol. 2000, 131, 482–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckland, K.F.; Williams, T.J.; Conroy, D.M. Histamine Induces Cytoskeletal Changes in Human Eosinophils via the H 4 Receptor: Histamine H 4 Receptors and Human Eosinophil Responses. Br. J. Pharmacol. 2003, 140, 1117–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merves, J.; Chandramouleeswaran, P.M.; Benitez, A.J.; Muir, A.B.; Lee, A.J.; Lim, D.M.; Dods, K.; Mehta, I.; Ruchelli, E.D.; Nakagawa, H.; et al. Altered Esophageal Histamine Receptor Expression in Eosinophilic Esophagitis (EoE): Implications on Disease Pathogenesis. PLoS ONE 2015, 10, e0114831. [Google Scholar] [CrossRef] [PubMed]
- Wallon, C.; Persborn, M.; Jönsson, M.; Wang, A.; Phan, V.; Lampinen, M.; Vicario, M.; Santos, J.; Sherman, P.M.; Carlson, M.; et al. Eosinophils Express Muscarinic Receptors and Corticotropin-Releasing Factor to Disrupt the Mucosal Barrier in Ulcerative Colitis. Gastroenterology 2011, 140, 1597–1607. [Google Scholar] [CrossRef]
- Blanchet, M.-R.; Langlois, A.; Israël-Assayag, E.; Beaulieu, M.-J.; Ferland, C.; Laviolette, M.; Cormier, Y. Modulation of Eosinophil Activation in vitro by a Nicotinic Receptor Agonist. J. Leukoc. Biol. 2007, 81, 1245–1251. [Google Scholar] [CrossRef]
- Müller, T.; Robaye, B.; Vieira, R.P.; Ferrari, D.; Grimm, M.; Jakob, T.; Martin, S.F.; Di Virgilio, F.; Boeynaems, J.-M.; Virchow, J.C.; et al. The Purinergic Receptor P2Y2 Receptor Mediates Chemotaxis of Dendritic Cells and Eosinophils in Allergic Lung Inflammation: P2Y2 Receptors and Asthma. Allergy 2010, 65, 1545–1553. [Google Scholar] [CrossRef]
- Muniz, V.S.; Baptista-dos-Reis, R.; Benjamim, C.F.; Mata-Santos, H.A.; Pyrrho, A.S.; Strauch, M.A.; Melo, P.A.; Vicentino, A.R.R.; Silva-Paiva, J.; Bandeira-Melo, C.; et al. Purinergic P2Y12 Receptor Activation in Eosinophils and the Schistosomal Host Response. PLoS ONE 2015, 10, e0139805. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Soma, T.; Noguchi, T.; Nakagome, K.; Nakamoto, H.; Kita, H.; Nagata, M. ATP Drives Eosinophil Effector Responses through P2 Purinergic Receptors. Allergol. Int. 2015, 64, S30–S36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberto, A.V.P.; Faria, R.X.; de Menezes, J.R.L.; Surrage, A.; da Rocha, N.C.; Ferreira, L.G.B.; Frutuoso, V.d.S.; Martins, M.A.; Alves, L.A. Role of P2 Receptors as Modulators of Rat Eosinophil Recruitment in Allergic Inflammation. PLoS ONE 2016, 11, e0145392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehme, S.A.; Lio, F.M.; Sikora, L.; Pandit, T.S.; Lavrador, K.; Rao, S.P.; Sriramarao, P. Cutting Edge: Serotonin Is a Chemotactic Factor for Eosinophils and Functions Additively with Eotaxin. J. Immunol. 2004, 173, 3599–3603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, B.N.; Ha, S.G.; Bahaie, N.S.; Hosseinkhani, M.R.; Ge, X.N.; Blumenthal, M.N.; Rao, S.P.; Sriramarao, P. Regulation of Serotonin-Induced Trafficking and Migration of Eosinophils. PLoS ONE 2013, 8, e54840. [Google Scholar] [CrossRef] [Green Version]
- Smyth, C.M.; Akasheh, N.; Woods, S.; Kay, E.; Morgan, R.K.; Thornton, M.A.; O’Grady, A.; Cummins, R.; Sheils, O.; Smyth, P.; et al. Activated Eosinophils in Association with Enteric Nerves in Inflammatory Bowel Disease. PLoS ONE 2013, 8, e64216. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, I.; Nakagawa, N.; Yamazaki, H.; Kimura, A.; Tomioka, H.; Yoshida, S. Mechanism for Substance P-Induced Activation of Human Neutrophils and Eosinophils. Regul. Pept. 1993, 46, 228–230. [Google Scholar] [CrossRef]
- Kroegel, C.; Giembycz, M.A.; Barnes, P.J. Characterization of Eosinophil Cell Activation by Peptides. Differential Effects of Substance P, Melittin, and FMET-Leu-Phe. J. Immunol. 1990, 145, 2581–2587. [Google Scholar]
- El-Shazly, A.E.; Begon, D.Y.; Kustermans, G.; Arafa, M.; Dortu, E.; Henket, M.; Lefebvre, P.P.; Louis, R.; Delvenne, P. Novel Association between Vasoactive Intestinal Peptide and CRTH2 Receptor in Recruiting Eosinophils. J. Biol. Chem. 2013, 288, 1374–1384. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.K.; Manohar, M.; Venkateshaiah, S.U.; Mishra, A. Neuroendocrine cells derived chemokine vasoactive intestinal polypeptide (VIP) in allergic diseases. Cytokine Growth Factor Rev. 2017, 38, 37–48. [Google Scholar] [CrossRef]
- Sawatzky, D.A.; Kingham, P.J.; Court, E.; Kumaravel, B.; Fryer, A.D.; Jacoby, D.B.; McLean, W.G.; Costello, R.W. Eosinophil Adhesion to Cholinergic Nerves via ICAM-1 and VCAM-1 and Associated Eosinophil Degranulation. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2002, 282, L1279–L1288. [Google Scholar] [CrossRef] [Green Version]
- Nie, Z.; Nelson, C.S.; Jacoby, D.B.; Fryer, A.D. Expression and Regulation of Intercellular Adhesion Molecule-1 on Airway Parasympathetic Nerves. J. Allergy Clin. Immunol. 2007, 119, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.-T.; Curran, D.R.; Kingham, P.J.; Morgan, R.K.; Durcan, N.; Gleich, G.J.; McLean, W.G.; Costello, R.W. Effect of Eosinophil Adhesion on Intracellular Signaling in Cholinergic Nerve Cells. Am. J. Respir. Cell Mol. Biol. 2004, 30, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Kingham, P.J.; McLean, W.G.; Sawatzky, D.A.; Walsh, M.T.; Costello, R.W. Adhesion-Dependent Interactions between Eosinophils and Cholinergic Nerves. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2002, 282, L1229–L1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor-Clark, T.E.; Undem, B.J. Sensing Pulmonary Oxidative Stress by Lung Vagal Afferents. Respir. Physiol. Neurobiol. 2011, 178, 406–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, C.M.; Belmonte, K.E.; Costello, R.W.; Jacoby, D.B.; Gleich, G.J.; Fryer, A.D. Substance P-Induced Airway Hyperreactivity Is Mediated by Neuronal M 2 Receptor Dysfunction. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2000, 279, L477–L486. [Google Scholar] [CrossRef] [Green Version]
- Raap, M.; Rüdrich, U.; Ständer, S.; Gehring, M.; Kapp, A.; Raap, U. Substance P Activates Human Eosinophils. Exp. Dermatol. 2015, 24, 557–559. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, L.M.; Fitzpatrick, E.; Baird, A.W.; Campion, D.P. Eosinophil–Nerve Interactions and Neuronal Plasticity in Rat Gut Associated Lymphoid Tissue (GALT) in Response to Enteric Parasitism. J. Neuroimmunol. 2008, 197, 1–9. [Google Scholar] [CrossRef]
- Zheng, P.-Y.; Feng, B.-S.; Oluwole, C.; Struiksma, S.; Chen, X.; Li, P.; Tang, S.-G.; Yang, P.-C. Psychological Stress Induces Eosinophils to Produce Corticotrophin Releasing Hormone in the Intestine. Gut 2009, 58, 1473–1479. [Google Scholar] [CrossRef]
- Kiank, C.; Taché, Y.; Larauche, M. Stress-Related Modulation of Inflammation in Experimental Models of Bowel Disease and Post-Infectious Irritable Bowel Syndrome: Role of Corticotropin-Releasing Factor Receptors. Brain Behav. Immun. 2010, 24, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Salvo-Romero, E.; Martínez, C.; Lobo, B.; Rodiño-Janeiro, B.K.; Pigrau, M.; Sánchez-Chardi, A.D.; González-Castro, A.M.; Fortea, M.; Pardo-Camacho, C.; Nieto, A.; et al. Overexpression of Corticotropin-Releasing Factor in Intestinal Mucosal Eosinophils Is Associated with Clinical Severity in Diarrhea-Predominant Irritable Bowel Syndrome. Sci. Rep. 2020, 10, 20706. [Google Scholar] [CrossRef]
- Neunlist, M.; Toumi, F.; Oreschkova, T.; Denis, M.; Leborgne, J.; Laboisse, C.L.; Galmiche, J.P.; Jarry, A. Human ENS regulates the intestinal epithelial barrier permeability and a tight junction-associated protein ZO-1 via VIPergic pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G1028–G1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metwali, A.; Blum, A.M.; Ferraris, L.; Klein, J.S.; Fiocchi, C.; Weinstock, J.V. Eosinophils within the healthy or inflamed human intestine produce substance P and vasoactive intestinal peptide. J. Neuroimmunol. 1994, 52, 69–78. [Google Scholar] [CrossRef]
- Carr, M.J.; Undem, B.J. Inflammation-Induced Plasticity of the Afferent Innervation of the Airways. Environ. Health Perspect. 2001, 109, 567–571. [Google Scholar] [CrossRef] [PubMed]
- Johansson, O.; Liang, Y.; Marcusson, J.A.; Reimert, C.M. Eosinophil Cationic Protein- and Eosinophil-Derived Neurotoxin/Eosinophil Protein X-Immunoreactive Eosinophils in Prurigo Nodularis. Arch. Dermatol. Res. 2000, 292, 371–378. [Google Scholar] [CrossRef]
- Zhang, H.; Verkman, A.S. Eosinophil Pathogenicity Mechanisms and Therapeutics in Neuromyelitis Optica. J. Clin. Investig. 2013, 123, 2306–2316. [Google Scholar] [CrossRef]
- Hogan, S.P.; Mishra, A.; Brandt, E.B.; Royalty, M.P.; Pope, S.M.; Zimmermann, N.; Foster, P.S.; Rothenberg, M.E. A Pathological Function for Eotaxin and Eosinophils in Eosinophilic Gastrointestinal Inflammation. Nat. Immunol. 2001, 2, 353–360. [Google Scholar] [CrossRef]
- Singh, U.R.; Malhotra, A.; Bhatia, A. Eosinophils, Mast Cells, Nerves and Ganglion Cells in Appendicitis. Indian J. Surg. 2008, 70, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Jacoby, D.B.; Gleich, G.J.; Fryer, A.D. Human Eosinophil Major Basic Protein Is an Endogenous Allosteric Antagonist at the Inhibitory Muscarinic M2 Receptor. J. Clin. Investig. 1993, 91, 1314–1318. [Google Scholar] [CrossRef] [Green Version]
- Morgan, R.K.; Kingham, P.J.; Walsh, M.T.; Curran, D.C.; Durcan, N.; McLean, W.G.; Costello, R.W. Eosinophil Adhesion to Cholinergic IMR-32 Cells Protects against Induced Neuronal Apoptosis. J. Immunol. 2004, 173, 5963–5970. [Google Scholar] [CrossRef] [Green Version]
- Morgan, R.K.; Costello, R.W.; Durcan, N.; Kingham, P.J.; Gleich, G.J.; McLean, W.G.; Walsh, M.-T. Diverse Effects of Eosinophil Cationic Granule Proteins on IMR-32 Nerve Cell Signaling and Survival. Am. J. Respir. Cell Mol. Biol. 2005, 33, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, C.; Bessissow, T.; Desmet, A.-S.; Vanheel, H.; Tack, J.; Vanden Berghe, P. Evidence for Neuronal and Structural Changes in Submucous Ganglia of Patients with Functional Dyspepsia. Am. J. Gastroenterol. 2015, 110, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Jung, H.-K.; Lee, K.E.; Mun, Y.-C.; Park, S. Degranulated Eosinophils Contain More Fine Nerve Fibers in the Duodenal Mucosa of Patients with Functional Dyspepsia. J. Neurogastroenterol. Motil. 2019, 25, 212–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperber, A.D.; Bangdiwala, S.I.; Drossman, D.A.; Ghoshal, U.C.; Simren, M.; Tack, J.; Whitehead, W.E.; Dumitrascu, D.L.; Fang, X.; Fukudo, S.; et al. Worldwide Prevalence and Burden of Functional Gastrointestinal Disorders, Results of Rome Foundation Global Study. Gastroenterology 2021, 160, 99–114. [Google Scholar] [CrossRef] [PubMed]
- Drossman, D.A.; Hasler, W.L. Rome IV—Functional GI Disorders: Disorders of Gut-Brain Interaction. Gastroenterology 2016, 150, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Talley, N.J.; Ford, A.C. Functional Dyspepsia. N. Engl. J. Med. 2015, 373, 1853–1863. [Google Scholar] [CrossRef] [Green Version]
- Ford, A.C.; Lacy, B.E.; Talley, N.J. Irritable Bowel Syndrome. N. Engl. J. Med. 2017, 376, 2566–2578. [Google Scholar] [CrossRef] [Green Version]
- Beeckmans, D.; Farré, R.; Riethorst, D.; Keita, Å.V.; Augustijns, P.; Söderholm, J.D.; Vanuytsel, T.; Vanheel, H.; Tack, J. Relationship between Bile Salts, Bacterial Translocation, and Duodenal Mucosal Integrity in Functional Dyspepsia. Neurogastroenterol. Motil. 2020, 32, e13788. [Google Scholar] [CrossRef]
- Vanheel, H.; Carbone, F.; Valvekens, L.; Simren, M.; Tornblom, H.; Vanuytsel, T.; Van Oudenhove, L.; Tack, J. Pathophysiological Abnormalities in Functional Dyspepsia Subgroups According to the Rome III Criteria. Am. J. Gastroenterol. 2017, 112, 132–140. [Google Scholar] [CrossRef]
- Vanheel, H.; Vicario, M.; Vanuytsel, T.; Van Oudenhove, L.; Martinez, C.; Keita, Å.V.; Pardon, N.; Santos, J.; Söderholm, J.D.; Tack, J.; et al. Impaired Duodenal Mucosal Integrity and Low-Grade Inflammation in Functional Dyspepsia. Gut 2014, 63, 262–271. [Google Scholar] [CrossRef]
- Vanheel, H.; Vicario, M.; Boesmans, W.; Vanuytsel, T.; Salvo-Romero, E.; Tack, J.; Farré, R. Activation of Eosinophils and Mast Cells in Functional Dyspepsia: An Ultrastructural Evaluation. Sci. Rep. 2018, 8, 5383. [Google Scholar] [CrossRef]
- Nojkov, B.; Zhou, S.-Y.; Dolan, R.D.; Davis, E.M.; Appelman, H.D.; Guo, X.; Jackson, K.; Sturm, M.B.; Wang, T.D.; Owyang, C.; et al. Evidence of Duodenal Epithelial Barrier Impairment and Increased Pyroptosis in Patients with Functional Dyspepsia on Confocal Laser Endomicroscopy and “Ex Vivo” Mucosa Analysis. Am. J. Gastroenterol. 2020, 115, 1891–1901. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.P.; O’Brien, D.R.; Sharma, M.; Smyrk, T.C.; Graham, R.P.; Grover, M.; Bharucha, A.E. Duodenal Mucosal Barrier in Functional Dyspepsia. Clin. Gastroenterol. Hepatol. 2021, 20, 1019–1028.e3. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Hara, K.; Fikree, A.; Siddiqi, S.; Woodland, P.; Masamune, A.; Aziz, Q.; Sifrim, D.; Yazaki, E. Patients with Dyspepsia Have Impaired Mucosal Integrity Both in the Duodenum and Jejunum: in vivo Assessment of Small Bowel Mucosal Integrity Using Baseline Impedance. J. Gastroenterol. 2020, 55, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deiteren, A.; de Wit, A.; van der Linden, L.; De Man, J.G.; Pelckmans, P.A.; De Winter, B.Y. Irritable Bowel Syndrome and Visceral Hypersensitivity: Risk Factors and Pathophysiological Mechanisms. Acta Gastroenterol. Belg. 2016, 79, 29–38. [Google Scholar]
- Martínez, C.; Lobo, B.; Pigrau, M.; Ramos, L.; González-Castro, A.M.; Alonso, C.; Guilarte, M.; Guilá, M.; de Torres, I.; Azpiroz, F.; et al. Diarrhoea-Predominant Irritable Bowel Syndrome: An Organic Disorder with Structural Abnormalities in the Jejunal Epithelial Barrier. Gut 2013, 62, 1160–1168. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.; Schurman, J.V.; Colombo, J.M.; Friesen, C.A. The Relationship between Mucosal Inflammatory Cells, Specific Symptoms, and Psychological Functioning in Youth with Irritable Bowel Syndrome. Sci. Rep. 2020, 10, 11988. [Google Scholar] [CrossRef]
- Dunn, J.L.M.; Rothenberg, M.E. 2021 Year in Review: Spotlight on Eosinophils. J. Allergy Clin. Immunol. 2022, 149, 517–524. [Google Scholar] [CrossRef]
- Dellon, E.S.; Simon, D.; Wechsler, M.E. The Potential Role of Biologics as First-Line Therapy in Eosinophilic Disorders. The J. Allergy Clin. Immunol. Pract. 2022, 10, 1169–1176. [Google Scholar] [CrossRef]
- Dellon, E.S.; Peterson, K.A.; Murray, J.A.; Falk, G.W.; Gonsalves, N.; Chehade, M.; Genta, R.M.; Leung, J.; Khoury, P.; Klion, A.D.; et al. Anti–Siglec-8 Antibody for Eosinophilic Gastritis and Duodenitis. N. Engl. J. Med. 2020, 383, 1624–1634. [Google Scholar] [CrossRef]

| Type of Molecule | Class | Function | Mediator |
|---|---|---|---|
| Cationic Proteins | Preformed | Host Defense/Barrier Function Homeostasis | ECP, EDN, EPO, MBP |
| Cytokines and growth factors | Preformed and de novo synthesis | Adaptive immunity | Th1: IFN-γ, IL-2, IL-12 Th2: IL-4, IL-5, IL-13, IL-9, IL-25 Th17: IL-17A, IL-17F Treg: IL-10, TGF-β |
| Innate immunity | GM-CSF, IL-3, IL-4, IL-5, IL-13, SCF | ||
| B cell class-switch and plasma cell maintenance | APRIL, IL-4, IL-6 | ||
| Tissue remodeling and repair | TGF-α, TGF-β, IL-1β, IL-13, PDGF-B, VEGF | ||
| Chemokines | Preformed and de novo synthesis | Recruitment of innate and adaptive immune cells | CCL3, CCL5, CCL6, CCL7, CCL8, CCL11, CCL13, CCL17, CCL22, CXCL9, CXCL10, IL-8 |
| Lipid mediators | De novo synthesis | Pro-inflammatory | LTC4, PAF, PGE, PGF1, TxA2 |
| Resolution inflammation | PD1, RvE3 | ||
| Neuropeptides and neurotrophines | Preformed and de novo synthesis | Nerve function, survival, and development | CGRP, CRF, NGF, NT-3, SP, VIP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvo-Romero, E.; Rodiño-Janeiro, B.K.; Albert-Bayo, M.; Lobo, B.; Santos, J.; Farré, R.; Martinez, C.; Vicario, M. Eosinophils in the Gastrointestinal Tract: Key Contributors to Neuro-Immune Crosstalk and Potential Implications in Disorders of Brain-Gut Interaction. Cells 2022, 11, 1644. https://doi.org/10.3390/cells11101644
Salvo-Romero E, Rodiño-Janeiro BK, Albert-Bayo M, Lobo B, Santos J, Farré R, Martinez C, Vicario M. Eosinophils in the Gastrointestinal Tract: Key Contributors to Neuro-Immune Crosstalk and Potential Implications in Disorders of Brain-Gut Interaction. Cells. 2022; 11(10):1644. https://doi.org/10.3390/cells11101644
Chicago/Turabian StyleSalvo-Romero, Eloísa, Bruno K. Rodiño-Janeiro, Mercé Albert-Bayo, Beatriz Lobo, Javier Santos, Ricard Farré, Cristina Martinez, and María Vicario. 2022. "Eosinophils in the Gastrointestinal Tract: Key Contributors to Neuro-Immune Crosstalk and Potential Implications in Disorders of Brain-Gut Interaction" Cells 11, no. 10: 1644. https://doi.org/10.3390/cells11101644
APA StyleSalvo-Romero, E., Rodiño-Janeiro, B. K., Albert-Bayo, M., Lobo, B., Santos, J., Farré, R., Martinez, C., & Vicario, M. (2022). Eosinophils in the Gastrointestinal Tract: Key Contributors to Neuro-Immune Crosstalk and Potential Implications in Disorders of Brain-Gut Interaction. Cells, 11(10), 1644. https://doi.org/10.3390/cells11101644