
Hypothyroidism Intensifies Both Canonic and the De Novo Pathway of Peroxisomal Biogenesis in Rat Brown Adipocytes in a Time-Dependent Manner

 , , , and
, , , and
Abstract
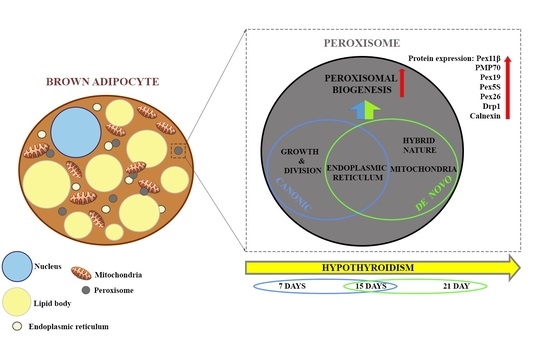
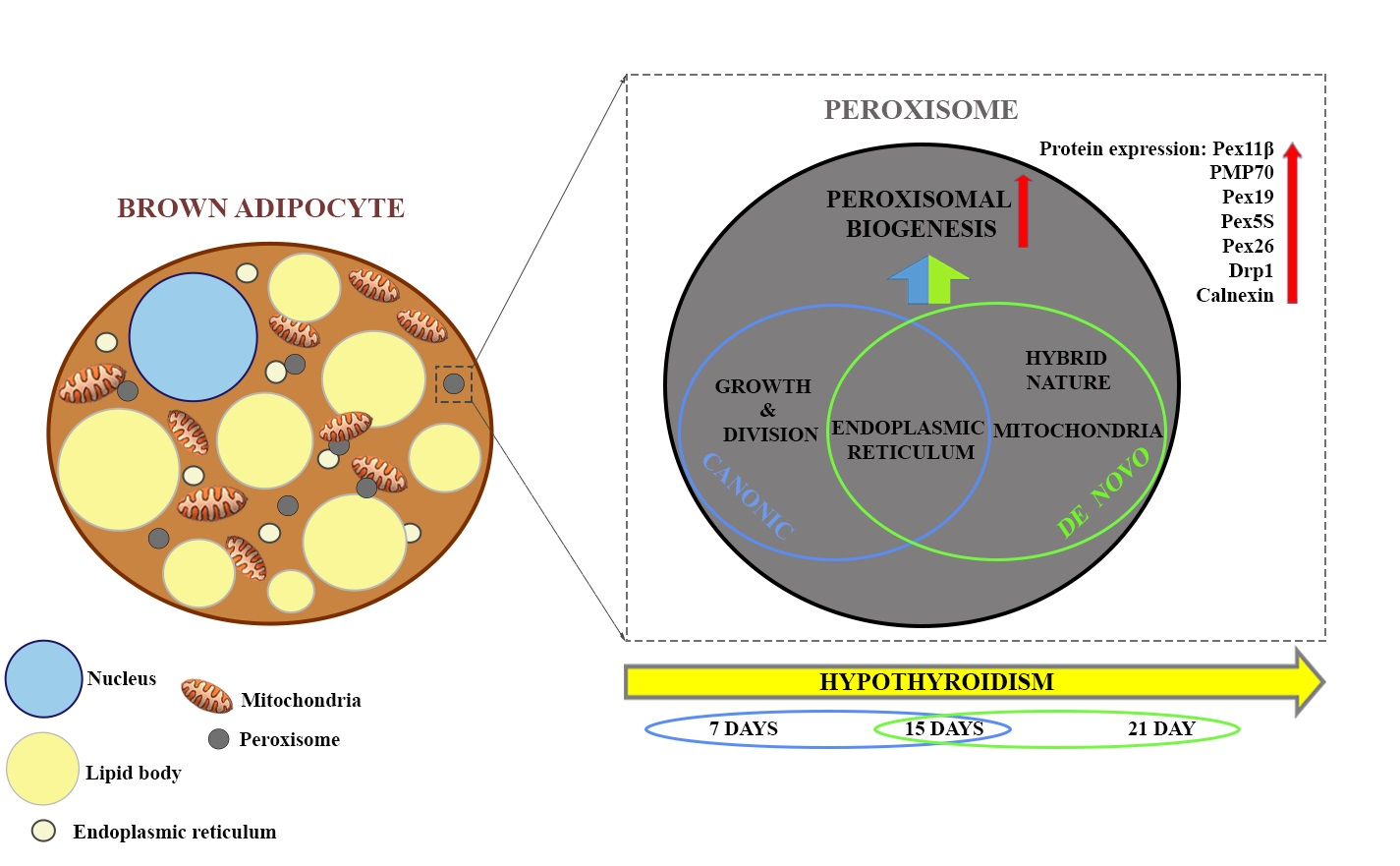{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Transmission Electron Microscopy
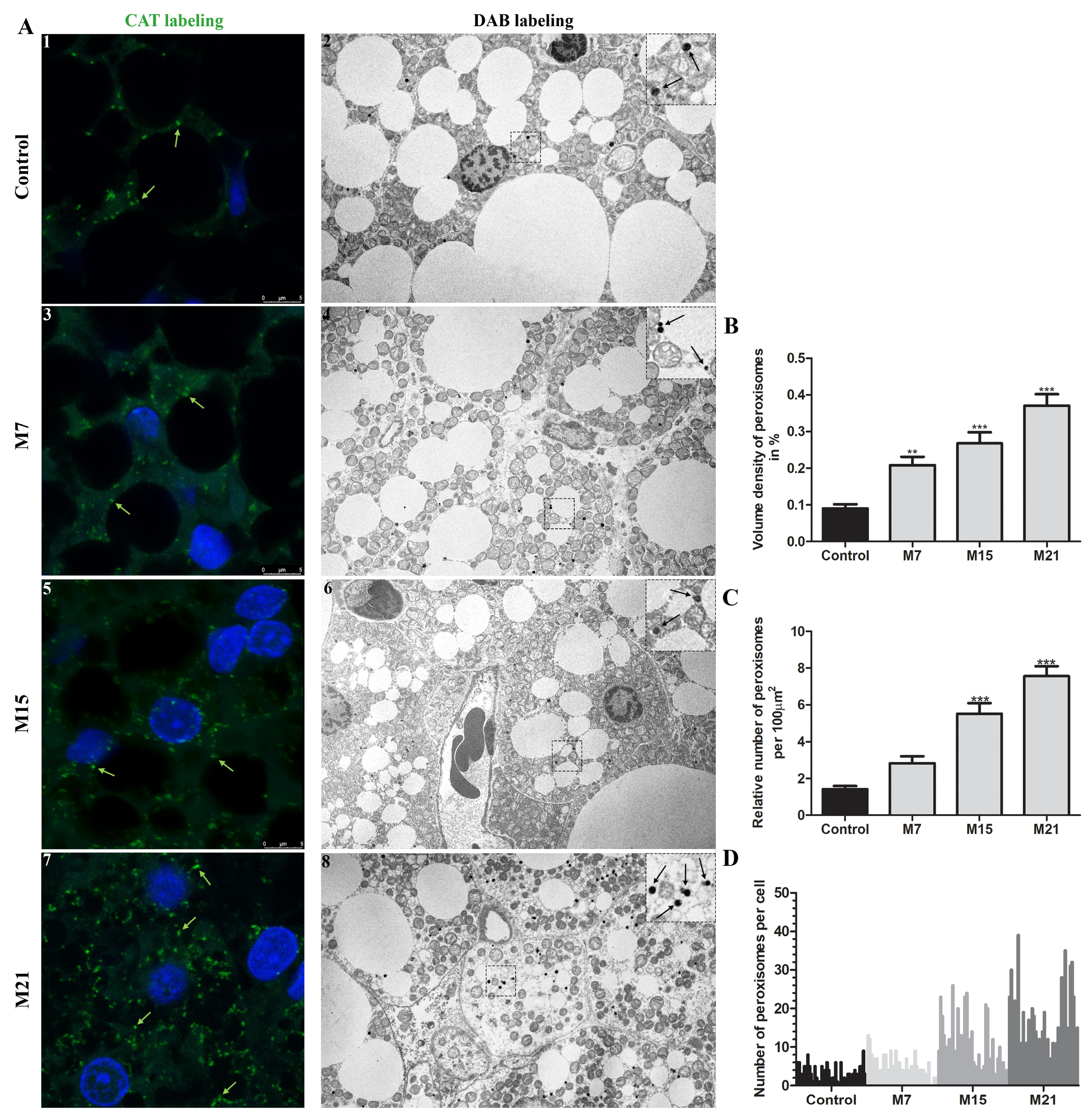
2.3. Stereological and Morphometric Analyses
2.4. Immunofluorescence
2.5. Immunogold
2.6. Western Blotting
2.7. Statistics
3. Results
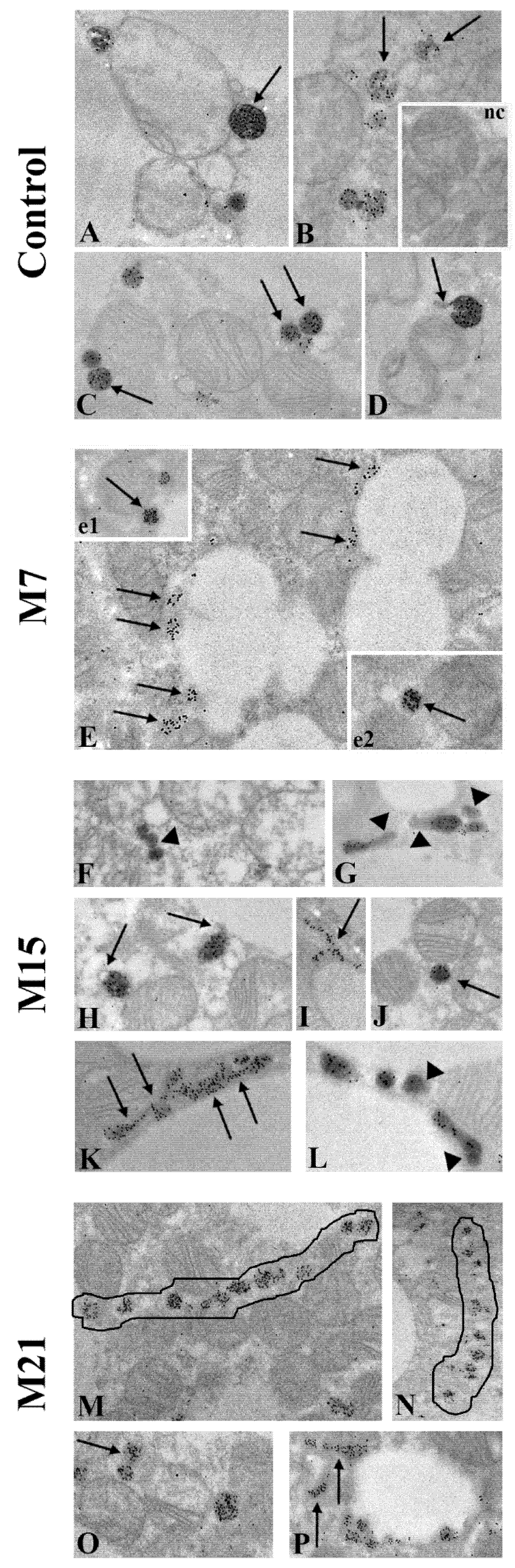
3.1. Hypothyroidism Induces Peroxisomal Proliferation
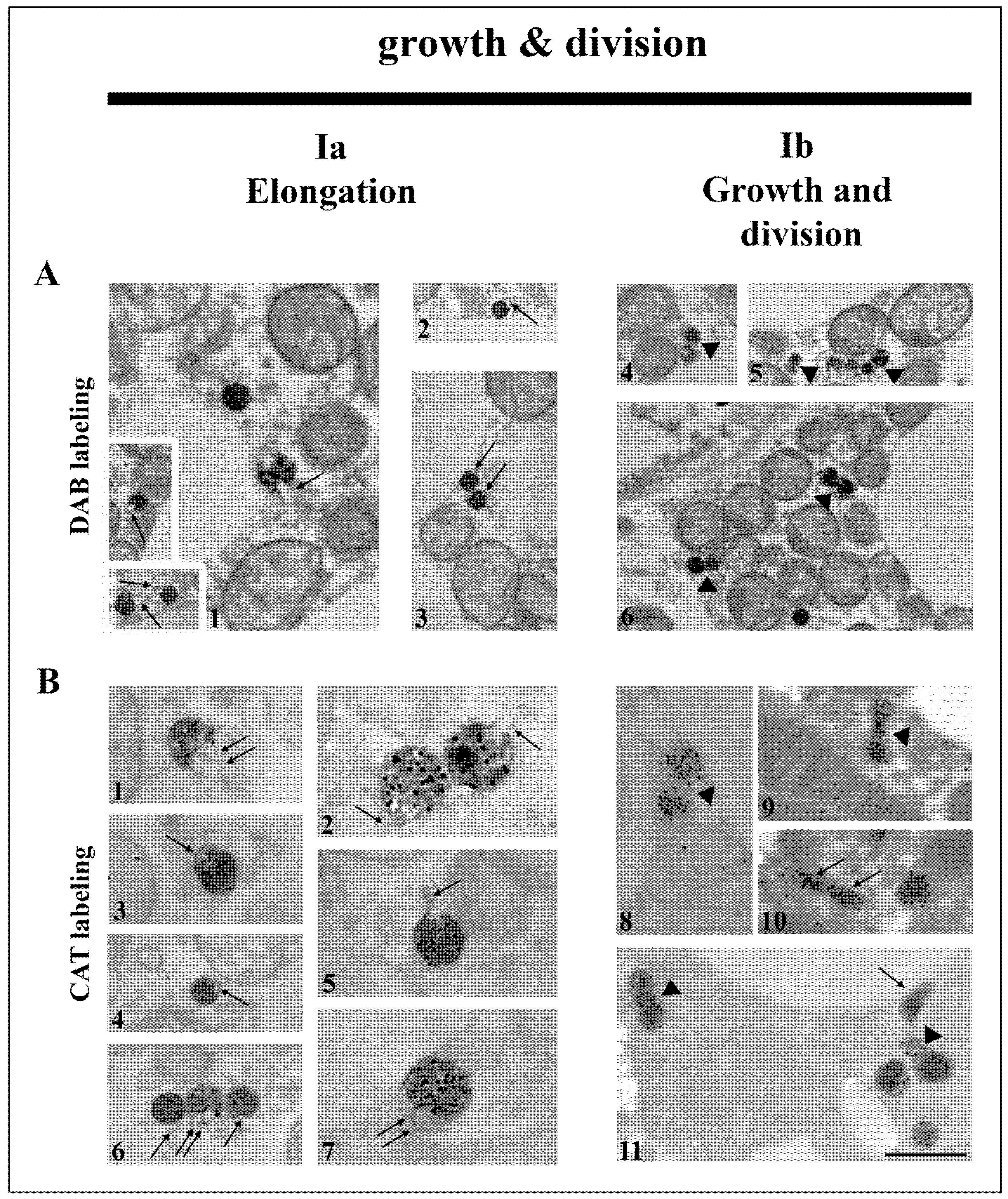
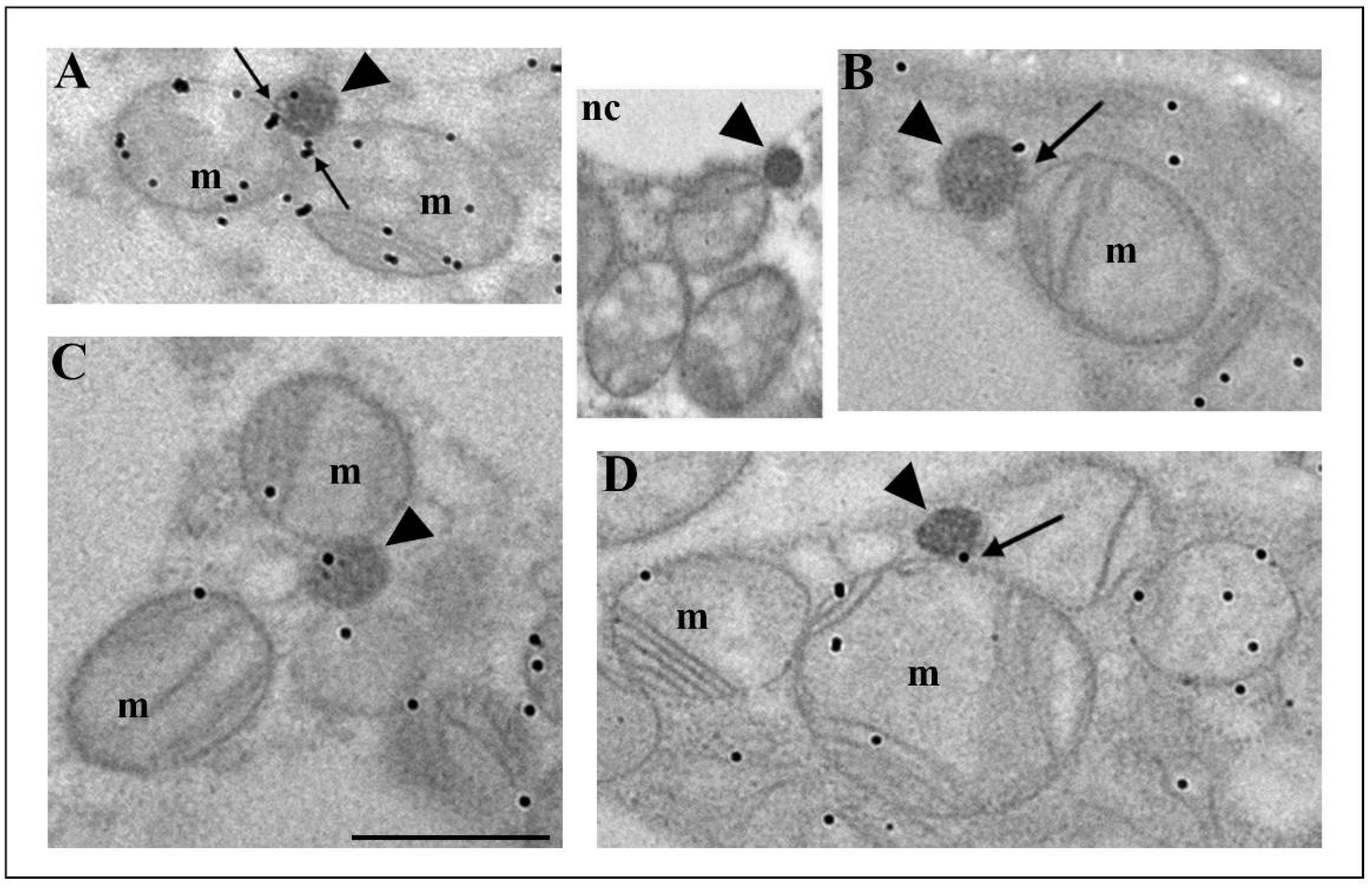
3.2. Hypothyroidism Intensifies Both Canonic and De Novo Pathway of Peroxisomal Biogenesis in a Time-Dependent Manner
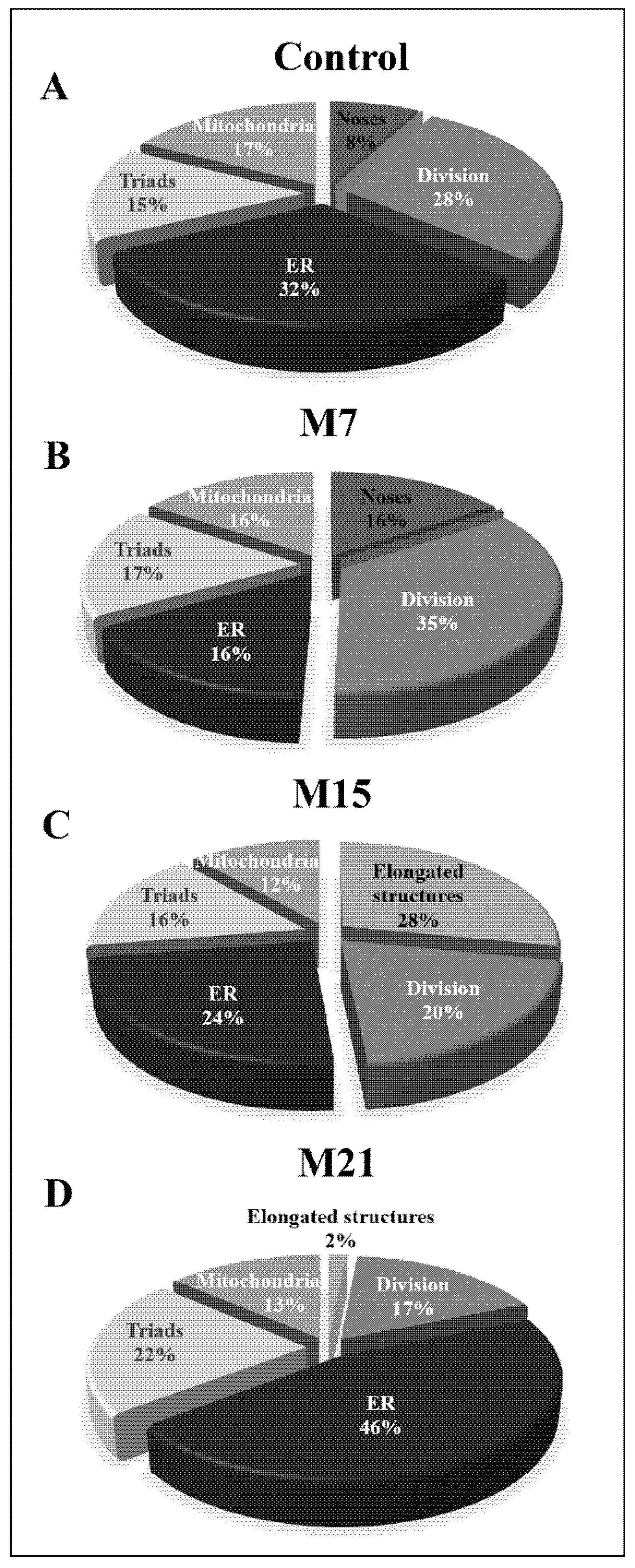
3.3. How Extensive Are the Contributions of ER, Mitochondria, or Hybrid Triads to the Observed Biogenesis Pathway, and Are They Affected over the Time Course of Hypothyroidism?
3.4. The Peroxisomal Maturation Level
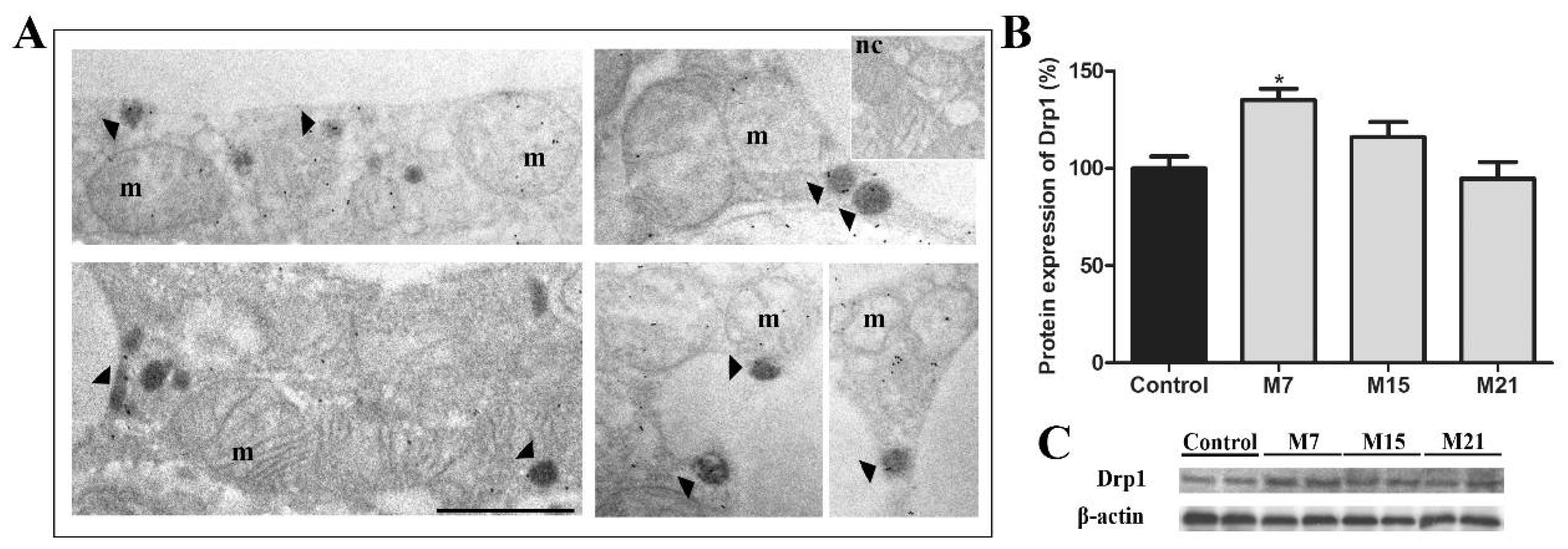
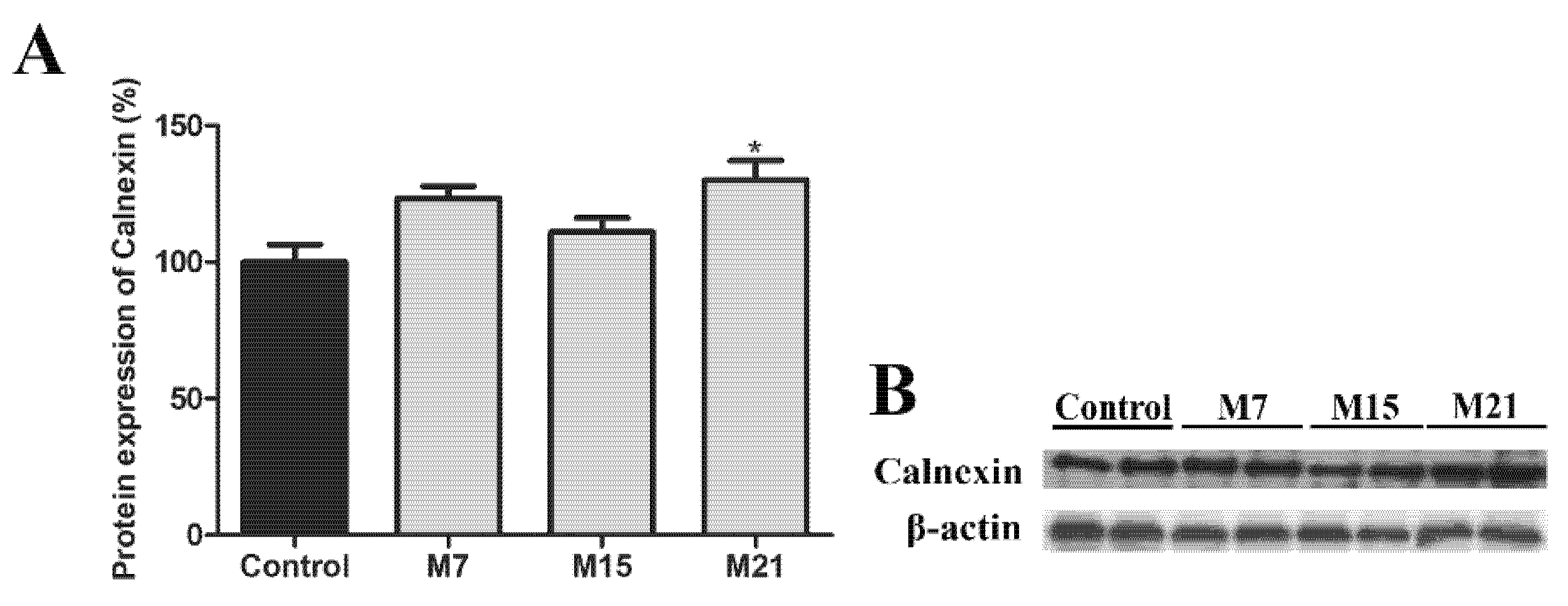
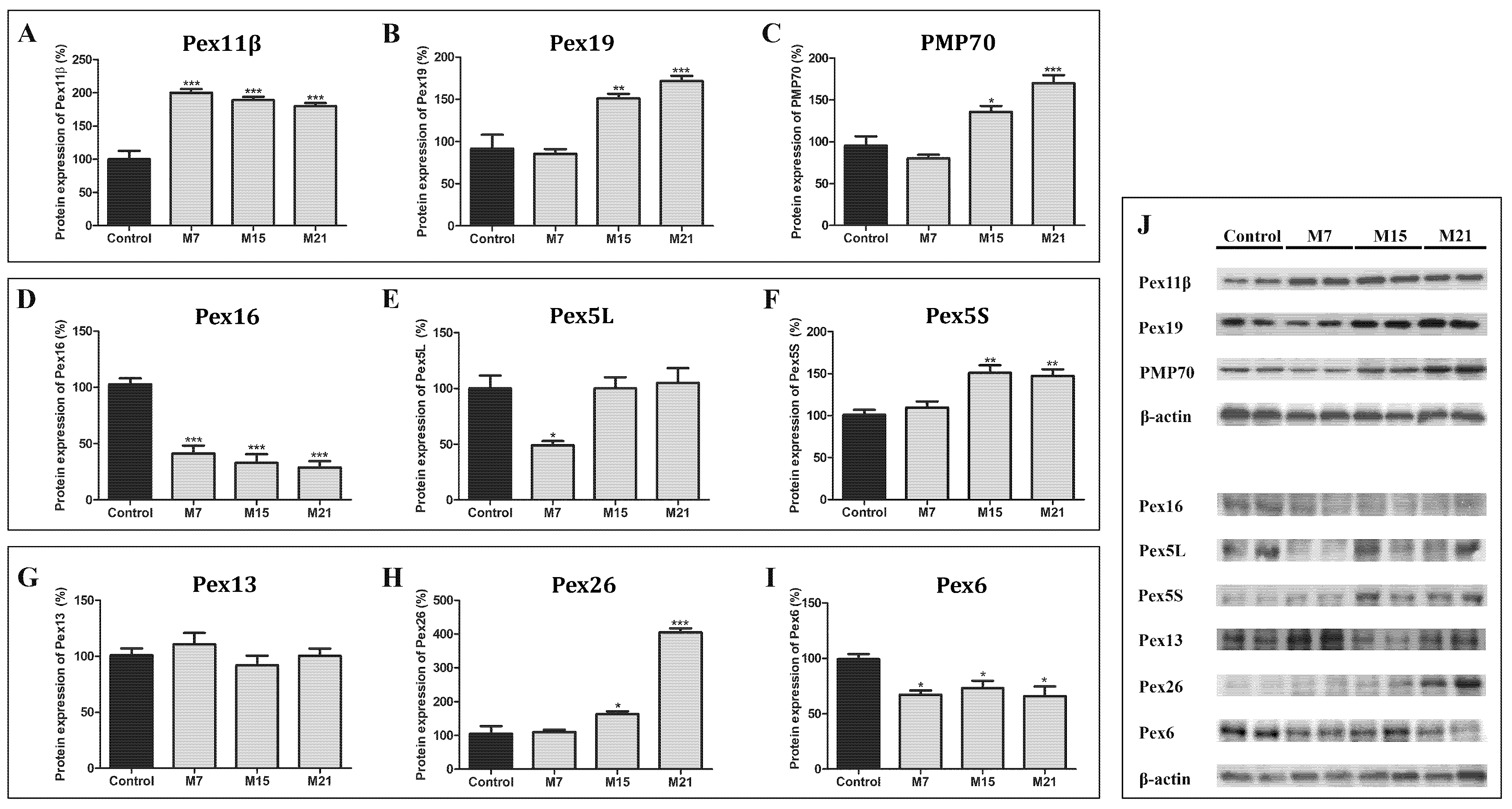
3.5. Peroxisomal Biogenesis in Brown Adipocytes in Hypothyroidism Is Driven by a Specific Set of Peroxins and PMP70
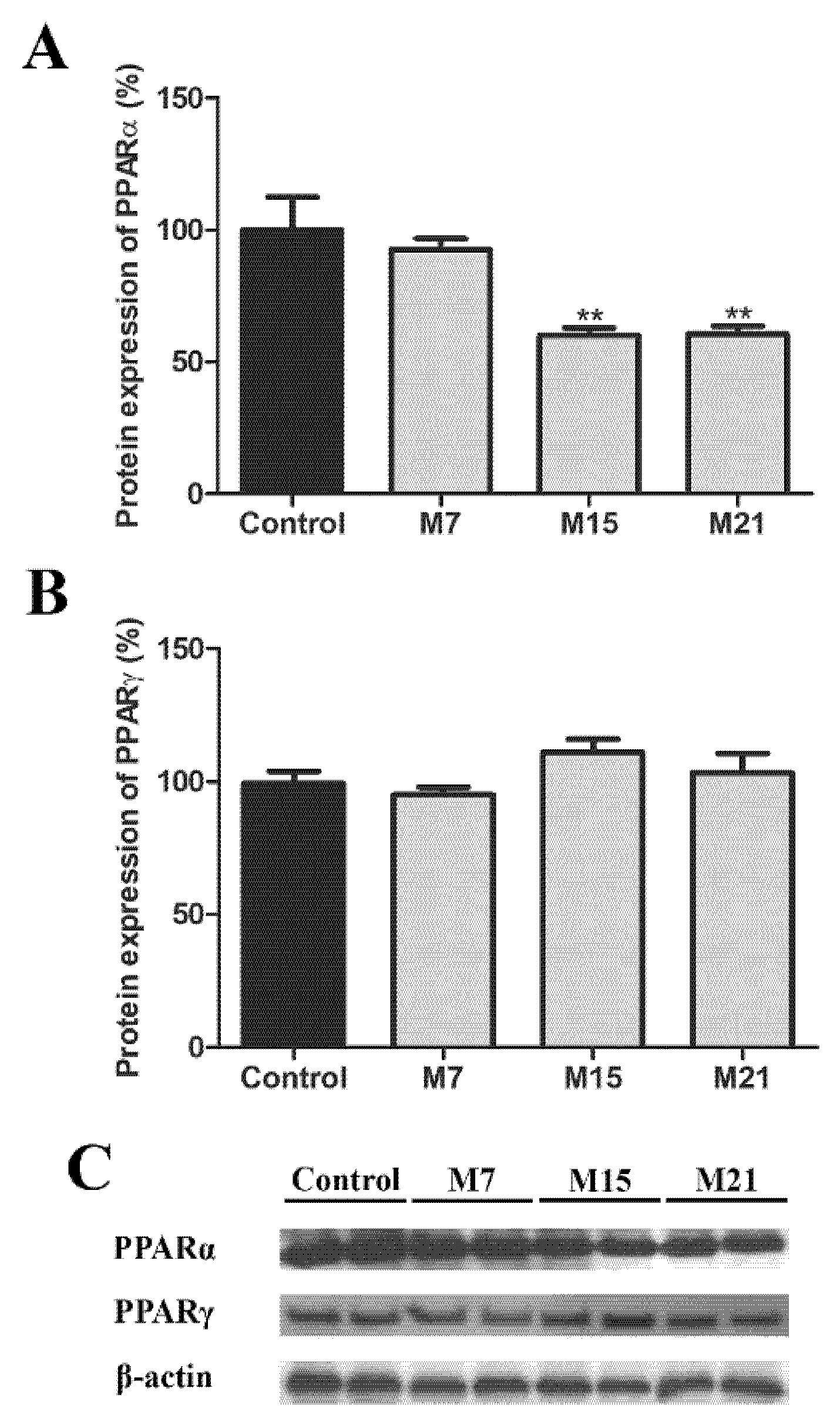
3.6. Transcription Regulation of Peroxisomal Biogenesis in Brown Adipocytes
4. Discussion
4.1. Peroxisomal Biogenesis by Growth and Division in Brown Adipocytes
4.2. Peroxisomal De Novo Biogenesis in Brown Adipocytes
4.3. Peroxisomal Maturation Level
4.4. Transcriptional Regulation of Peroxisomal Biogenesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, J.J.; Aitchison, J.D. Peroxisomes take shape. Nat. Rev. Mol. Cell Biol. 2013, 14, 803–817. [Google Scholar] [CrossRef]
- Lodhi, I.J.; Semenkovich, C.F. Peroxisomes: A nexus for lipid metabolism and cellular signaling. Cell Metab. 2014, 19, 380–392. [Google Scholar] [CrossRef]
- Fujiki, Y.; Okumoto, K.; Mukai, S.; Honsho, M.; Tamura, S. Peroxisome biogenesis in mammalian cells. Front. Physiol. 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Schrader, M.; Costello, J.L.; Godinho, L.F.; Azadi, A.S.; Islinger, M. Proliferation and fission of peroxisomes—An update. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 971–983. [Google Scholar] [CrossRef]
- Agrawal, G.; Subramani, S. De novo peroxisome biogenesis: Evolving concepts and conundrums. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 892–901. [Google Scholar] [CrossRef] [PubMed]
- Neuspiel, M.; Schauss, A.C.; Braschi, E.; Zunino, R.; Rippstein, P.; Rachubinski, R.A.; Andrade-Navarro, M.A.; McBride, H.M. Cargo-Selected Transport from the Mitochondria to Peroxisomes Is Mediated by Vesicular Carriers. Curr. Biol. 2008, 18, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, A.; McBride, H.M. Emerging roles of mitochondria in the evolution, biogenesis, and function of peroxisomes. Front. Physiol. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Sugiura, A.; Mattie, S.; Prudent, J.; Mcbride, H.M. Newly born peroxisomes are a hybrid of mitochondrial and ER-derived pre-peroxisomes. Nature 2017, 542, 251–254. [Google Scholar] [CrossRef]
- Guardiola-Diaz, H.M.; Rehnmark, S.; Usuda, N.; Albrektsen, T.; Feltkamp, D.; Gustafsson, J.Å.; Alexson, S.E.H. Rat peroxisome proliferator-activated receptors and brown adipose tissue function during cold acclimatization. J. Biol. Chem. 1999, 274, 23368–23377. [Google Scholar] [CrossRef]
- Ahlabo, I.; Barnard, T. Observations on peroxisomes in brown adipose tissue of the rat. J. Histochem. Cytochem. 1971, 19, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Pavelka, M.; Goldenberg, H.; Htittinger, M.; Kramar, R. Enzymic and Morphological Studies on Catalase Positive Particles from Brown Fat of Cold Adapted Rats. Histochemistry 1976, 50, 47–55. [Google Scholar] [CrossRef]
- Nedergaard, J.; Alexson, S.; Cannon, B. Cold adaptation in the rat: Increased brown fat peroxisomal β-oxidation relative to maximal mitochondrial oxidative capacity. Am. J. Physiol. Cell Physiol. 1980, 8. [Google Scholar] [CrossRef]
- Petrović, V.; Buzadžić, B.; Korać, A.; Vasilijević, A.; Janković, A.; Korać, B. Free radical equilibrium in interscapular brown adipose tissue: Relationship between metabolic profile and antioxidative defense. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 60–65. [Google Scholar] [CrossRef]
- Cannon, B.; Alexson, S.; Nedergaard, J. Peroxisomal beta-oxidation in brown fat. Ann. N. Y. Acad. Sci. 1982, 386, 40–58. [Google Scholar] [CrossRef]
- Craemer, D. De Secondary alterations of human hepatocellular peroxisomes. J. Inherit. Metab. Dis. 1995, 18, 181–213. [Google Scholar] [CrossRef]
- LeHir, M.; Herzog, V.; Fahimi, H.D. Cytochemical Detection of Catalase with 3,3′-Diaminobenzidine. Histochemistry 1979, 66, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Weibel, E.R.; Stäubli, W.; Gnägi, H.R.; Hess, F.A. Correlated Morphometric and Biochemical Studies on the Liver Cell. J. Cell Biol. 1969, 42, 92–112. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. Anal. Biochem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Lazarow, P.B.; Fujiki, Y. Biogenesis of peroxisomes. Annu. Rev. Cell Biol. 1985, 1, 489–530. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gould, S.J. PEX11 promotes peroxisome division independently of peroxisome metabolism. J. Cell Biol. 2002, 156, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gould, S.J. Membrane Transport Structure Function and Biogenesis: The Dynamin-like GTPase DLP1 is Essential for Peroxisome Division and is Recruited to Peroxisomes in Part by PEX11. J. Biol. Chem. 2003, 278, 17012–17020. [Google Scholar] [CrossRef]
- Thoms, S.; Erdmann, R. Dynamin-related proteins and Pex11 proteins in peroxisome division and proliferation. FEBS J. 2005, 272, 5169–5181. [Google Scholar] [CrossRef]
- Koch, A.; Yoon, Y.; Bonekamp, N.A.; McNiven, M.A.; Michael, S. A Role for Fis1 in Both Mitochondrial and Peroxisomal Fission in Mammalian Cells. Mol. Biol. Cell 2005, 16, 5077–5086. [Google Scholar] [CrossRef]
- Koch, J.; Pranjic, K.; Huber, A.; Ellinger, A.; Hartig, A.; Kragler, F.; Brocard, C. PEX11 family members are membrane elongation factors that coordinate peroxisome proliferation and maintenance. J. Cell Sci. 2010, 123, 3389–3400. [Google Scholar] [CrossRef]
- Delille, H.K.; Agricola, B.; Guimaraes, S.C.; Borta, H.; Lüers, G.H.; Fransen, M.; Schrader, M. Pex11pβ-mediated growth and division of mammalian peroxisomes follows a maturation pathway. J. Cell Sci. 2010, 123, 2750–2762. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Reuber, B.E.; Morrell, J.C.; Jimenez-Sanchez, G.; Obie, C.; Stroh, T.A.; Valle, D.; Schroer, T.A.; Gould, S.J. Expression of PEX11β Mediates Peroxisome Proliferation in the Absence of Extracellular Stimuli. J. Biol. Chem. 1998, 273, 29607–29614. [Google Scholar] [CrossRef]
- Delille, H.K.; Dodt, G.; Schrader, M. Pex11pβ-mediated maturation of peroxisomes. Commun. Integr. Biol. 2011, 4, 51–54. [Google Scholar] [CrossRef]
- Koch, A. Peroxisome elongation and constriction but not fission can occur independently of dynamin-like protein 1. J. Cell Sci. 2004, 117, 3995–4006. [Google Scholar] [CrossRef] [PubMed]
- Motley, A.M.; Hettema, E.H. Yeast peroxisomes multiply by growth and division. J. Cell Biol. 2007, 178, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Van der Zand, A.; Gent, J.; Braakman, I.; Tabak, H.F. Biochemically distinct vesicles from the endoplasmic reticulum fuse to form peroxisomes. Cell 2012, 149, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.K.; Mullen, R.T.; Schumann, U.; Lippincott-Schwartz, J. The origin and maintenance of mammalian peroxisomes involves a de novo PEX16-dependent pathway from the ER. J. Cell Biol. 2006, 173, 521–532. [Google Scholar] [CrossRef]
- Novikoff, P.M.; Novikoff, A.B. Peroxisomes In Absorptive Cells Of Mammalian Small Intestine. J. Cell Biol. 1972, 53, 532–560. [Google Scholar] [CrossRef]
- Titorenko, V.I.; Rachubinski, R.A. Peroxisomal membrane fusion requires two AAA family ATPases, Pex1p and Pex6p. J. Cell Biol. 2000, 150, 881–886. [Google Scholar] [CrossRef]
- Lam, S.K.; Yoda, N.; Schekman, R. A vesicle carrier that mediates peroxisome protein traffic from the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2010, 108, E51–E52. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.; Joshi, S.; Subramani, S. Cell-free sorting of peroxisomal membrane proteins from the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2011, 108, 9113–9118. [Google Scholar] [CrossRef] [PubMed]
- Geuze, H.J.; Murk, J.L.; Stroobants, A.K.; Griffith, J.M.; Kleijmeer, M.J.; Koster, A.J.; Verkleij, A.J.; Distel, B.; Tabak, H.F. Involvement of the Endoplasmic Reticulum in Peroxisome Formation. Mol. Biol. Cell 2003, 14, 2900–2907. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, Y.; Lazarow, P.B. Post-translational import of fatty acyl-CoA oxidase and catalase into peroxisomes of rat liver in vitro. J. Biol. Chem. 1985, 260, 5603–5609. [Google Scholar] [CrossRef]
- Gould, S.J.; Valle, D. Peroxisome biogenesis disorders: Genetics and cell biology. Trends Genet. 2000, 16, 340–345. [Google Scholar] [CrossRef]
- Sacksteder, K.A.; Jones, J.M.; South, S.T.; Li, X.; Liu, Y.; Gould, S.J. PEX19 binds multiple peroxisomal membrane proteins, is predominantly cytoplasmic, and is required for peroxisome membrane synthesis. J. Cell Biol. 2000, 148, 931–944. [Google Scholar] [CrossRef]
- Jones, J.M.; Morrell, J.C.; Gould, S.J. PEX19 is a predominantly cytosolic chaperone and import receptor for class 1 peroxisomal membrane proteins. J. Cell Biol. 2004, 164, 57–67. [Google Scholar] [CrossRef]
- Van Roermund, C.W.T.; Ijlst, L.; Wagemans, T.; Wanders, R.J.A.; Waterham, H.R. A role for the human peroxisomal half-transporter ABCD3 in the oxidation of dicarboxylic acids. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2014, 1841, 563–568. [Google Scholar] [CrossRef]
- Fujiki, Y.; Miyata, N.; Matsumoto, N.; Tamura, S. Dynamic and functional assembly of the AAA peroxins, Pex1p and Pex6p, and their membrane receptor Pex26p involved in shuttling of the PTS1 receptor Pex5p in peroxisome biogenesis. Biochem. Soc. Trans. 2008, 36, 109–113. [Google Scholar] [CrossRef]
- Pieuchot, L.; Jedd, G. Peroxisome Assembly and Functional Diversity in Eukaryotic Microorganisms. Annu. Rev. Microbiol. 2012, 66, 237–263. [Google Scholar] [CrossRef]
- Dodt, G.; Warren, D.; Becker, E.; Rehling, P.; Gould, S.J. Domain Mapping of Human PEX5 Reveals Functional and Structural Similarities to Saccharomyces cerevisiae Pex18p and Pex21p. J. Biol. Chem. 2001, 276, 41769–41781. [Google Scholar] [CrossRef]
- Matsumoto, N.; Tamura, S.; Furuki, S.; Miyata, N.; Moser, A.; Shimozawa, N.; Moser, H.W.; Suzuki, Y.; Kondo, N.; Fujiki, Y. Mutations in Novel Peroxin Gene PEX26 That Cause Peroxisome-Biogenesis Disorders of Complementation Group 8 Provide a Genotype-Phenotype Correlation. Am. J. Hum. Genet. 2003, 73, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Tamura, S.; Yasutake, S.; Matsumoto, N.; Fujiki, Y. Dynamic and functional assembly of the AAA peroxins, Pex1p and Pex6p, and their membrane receptor Pex26p. J. Biol. Chem. 2006, 281, 27693–27704. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, Y.; Nashiro, C.; Miyata, N.; Tamura, S.; Okumoto, K. New insights into dynamic and functional assembly of the AAA peroxins, Pex1p and Pex6p, and their membrane receptor Pex26p in shuttling of PTS1-receptor Pex5p during peroxisome biogenesis ☆. BBA Mol. Cell Res. 2012, 1823, 145–149. [Google Scholar] [CrossRef]
- Weller, S.; Cajigas, I.; Morrell, J.; Obie, C.; Steel, G.; Gould, S.J.; Valle, D. Alternative Splicing Suggests Extended Function of PEX26 in Peroxisome Biogenesis. Am. J. Hum. Genet. 2005, 76, 987–1007. [Google Scholar] [CrossRef] [PubMed]
- Halbach, A. Targeting of the tail-anchored peroxisomal membrane proteins PEX26 and PEX15 occurs through C-terminal PEX19-binding sites. J. Cell Sci. 2006, 119, 2508–2517. [Google Scholar] [CrossRef] [PubMed]
- Yagita, Y.; Hiromasa, T.; Fujiki, Y. Tail-anchored PEX26 targets peroxisomes via a PEX19-dependent and TRC40-independent class I pathway. J. Cell Biol. 2013, 200, 651–666. [Google Scholar] [CrossRef]
- Goldfisher, S.; Moore, C.L.; Johnson, A.B.; Spiro, A.J.; Valsamis, M.P.; Wisniewski, H.K.; Ritch, R.H.; Norton, W.T.; Rapin, I.; Gartner, L.M. Peroxisomal and Mitochondrial Defects in the Cerebro-Hepato-Renal Syndrome. Science 1973, 182, 62–64. [Google Scholar] [CrossRef]
- Messina, A.; Reina, S.; Guarino, F.; Pinto, V. De Biochimica et Biophysica Acta VDAC isoforms in mammals ☆. BBA Biomembr. 2012, 1818, 1466–1476. [Google Scholar] [CrossRef]
- Antonenkov, V.D.; Hiltunen, J.K. Peroxisomal membrane permeability and solute transfer. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 1697–1706. [Google Scholar] [CrossRef]
- Yamamoto, K.; Fahimi, H.D. Three-dimensional reconstruction of a peroxisomal reticulum in regenerating rat liver: Evidence of interconnections between heterogeneous segments. J. Cell Biol. 1987, 105, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Wilcke, M.; Hultenby, K.; Alexson, S.E.H. Novel peroxisomal populations in subcellular fractions from rat liver. J. Biol. Chem. 1995, 270, 6949. [Google Scholar] [CrossRef]
- Luers, G.; Hashimoto, T.; Fahimi, H.D.; Volkl, A. Biogenesis of peroxisomes: Isolation and characterization of two distinct peroxisomal populations from normal and regenerating rat liver. J. Cell Biol. 1993, 121, 1271–1280. [Google Scholar] [CrossRef]
- Schrader, M.; Costello, J.; Godinho, L.F.; Islinger, M. Peroxisome-mitochondria interplay and disease. J. Inherit. Metab. Dis. 2015, 38, 681–702. [Google Scholar] [CrossRef] [PubMed]
- Aleksic, M.; Kalezic, A.; Saso, L.; Jankovic, A.; Korac, B.; Korac, A. The Unity of Redox and Structural Remodeling of Brown Adipose Tissue in Hypothyroidism. Antioxidants 2021, 10, 591. [Google Scholar] [CrossRef]
- Delille, H.K.; Alves, R.; Schrader, M. Biogenesis of peroxisomes and mitochondria: Linked by division. Histochem. Cell Biol. 2009, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, E.; Volkl, A.; Hashimoto, T.; Fahimi, H.D. Biogenesis of peroxisomes: Immunocytochemical investigation of peroxisomal membrane proteins in proliferating rat liver peroxisomes and in catalase-negative membrane loops. J. Cell Biol. 1989, 108, 2221–2231. [Google Scholar] [CrossRef]
- Roels, F.; Cornelis, A. Heterogeneity of catalase staining in human hepatocellular peroxisomes. J. Histochem. Cytochem. 1989, 37, 331–337. [Google Scholar] [CrossRef]
- De Craemer, D.; Kerckaert, I.; Roels, F. Hepatocellular peroxisomes in human alcoholic and drug-induced hepatitis: A quantitative study. Hepatology 1991, 14, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Titorenko, V.I.; Smith, J.J.; Szilard, R.K.; Rachubinski, R.A. Peroxisome Biogenesis in the Yeast Yarrowia lipolytica. Cell Biochem. Biophys. 2000, 32, 21–26. [Google Scholar] [CrossRef]
- Tan, X.; Titorenko, V.I.; van der Klei, I.J.; Sulter, G.J.; Haima, P.; Waterham, H.R.; Evers, M.; Harder, W.; Veenhuis, M.; Cregg, J.M. Characterization of peroxisome-deficient mutants of Hansenula polymorpha. Curr. Genet. 1995, 28, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Veenhuis, M.; Salomons, F.A.; Van Der Klei, I.J. Peroxisome biogenesis and degradation in yeast: A structure/function analysis. Microsc. Res. Tech. 2000, 51, 584–600. [Google Scholar] [CrossRef]
- Berger, J.; Moller, D.E. The Mechanisms of Action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef] [PubMed]
- Schrader, M.; Bonekamp, N.A.; Islinger, M. Fission and proliferation of peroxisomes. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 1343–1357. [Google Scholar] [CrossRef]
- Weng, H.; Endo, K.; Li, J.; Kito, N.; Iwai, N. Induction of Peroxisomes by Butyrate-Producing Probiotics. PLoS ONE 2015, 10, e0117851. [Google Scholar] [CrossRef]
- Bagattin, A.; Hugendubler, L.; Mueller, E. Transcriptional coactivator PGC-1 promotes peroxisomal remodeling and biogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 20376–20381. [Google Scholar] [CrossRef]
- Shen, Y.; Su, Y.; Silva, F.J.; Weller, A.H.; Sostre-Colón, J.; Titchenell, P.M.; Steger, D.J.; Seale, P.; Soccio, R.E. Shared PPARα/γ Target Genes Regulate Brown Adipocyte Thermogenic Function. Cell Rep. 2020, 30, 3079–3091.e5. [Google Scholar] [CrossRef]
- Hofer, D.C.; Pessentheiner, A.R.; Pelzmann, H.J.; Schlager, S.; Madreiter-Sokolowski, C.T.; Kolb, D.; Eichmann, T.O.; Rechberger, G.; Bilban, M.; Graier, W.F.; et al. Critical role of the peroxisomal protein PEX16 in white adipocyte development and lipid homeostasis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 358–368. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleksic, M.; Golic, I.; Kalezic, A.; Jankovic, A.; Korac, B.; Korac, A. Hypothyroidism Intensifies Both Canonic and the De Novo Pathway of Peroxisomal Biogenesis in Rat Brown Adipocytes in a Time-Dependent Manner. Cells 2021, 10, 2248. https://doi.org/10.3390/cells10092248
Aleksic M, Golic I, Kalezic A, Jankovic A, Korac B, Korac A. Hypothyroidism Intensifies Both Canonic and the De Novo Pathway of Peroxisomal Biogenesis in Rat Brown Adipocytes in a Time-Dependent Manner. Cells. 2021; 10(9):2248. https://doi.org/10.3390/cells10092248
Chicago/Turabian StyleAleksic, Marija, Igor Golic, Andjelika Kalezic, Aleksandra Jankovic, Bato Korac, and Aleksandra Korac. 2021. "Hypothyroidism Intensifies Both Canonic and the De Novo Pathway of Peroxisomal Biogenesis in Rat Brown Adipocytes in a Time-Dependent Manner" Cells 10, no. 9: 2248. https://doi.org/10.3390/cells10092248
APA StyleAleksic, M., Golic, I., Kalezic, A., Jankovic, A., Korac, B., & Korac, A. (2021). Hypothyroidism Intensifies Both Canonic and the De Novo Pathway of Peroxisomal Biogenesis in Rat Brown Adipocytes in a Time-Dependent Manner. Cells, 10(9), 2248. https://doi.org/10.3390/cells10092248