
Protective Role of Recombinant Human Thrombomodulin in Diabetes Mellitus

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Groups
2.3. Diabetes Status Evaluation
2.4. Histological Study
2.5. Apoptosis Evaluation in Pancreatic Islets
2.6. Cell Culture
2.7. Apoptosis Assay
2.8. Akt Activation
2.9. Analysis of Spleen Cells
2.10. Statistical Analysis
3. Results
3.1. Treatment with rhTM Improved Glucose Intolerance
3.2. Increased Insulin-Producing Cells in Diabetic Mice Treated with rhTM
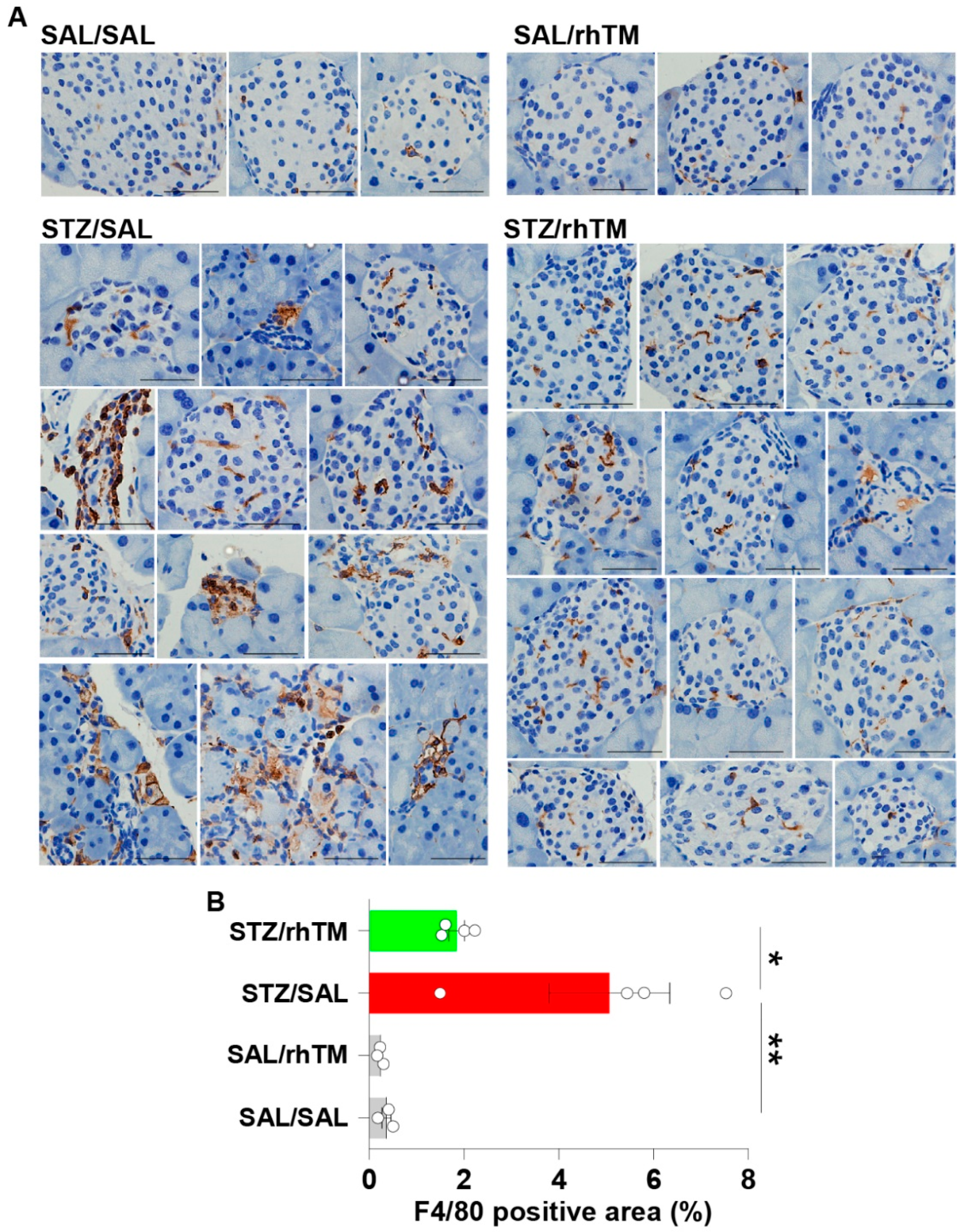
3.3. Decreased Islet Infiltration of Macrophages in Diabetic Mice Treated with rhTM
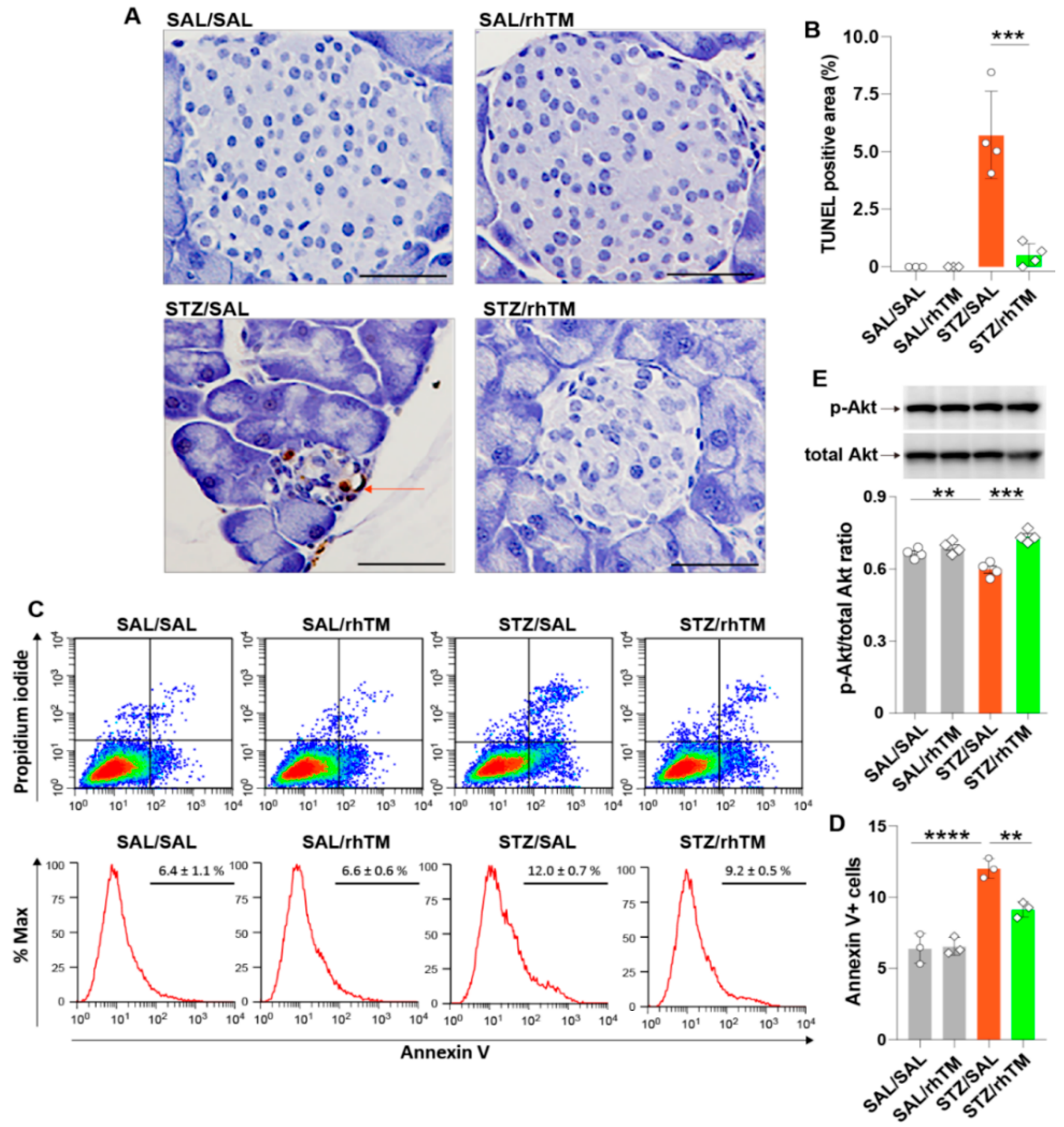
3.4. rhTM Activated the Akt Pathway and Inhibited Apoptosis of Islet β-Cells
3.5. rhTM Regulated the Immune Response Under Diabetic Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Collaborators, G.B.D.R.F. Global, regional, and national comparative risk assessment of 79 behavioural, environmental and occupational, and metabolic risks or clusters of risks, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1659–1724. [Google Scholar]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Harding, J.L.; Pavkov, M.E.; Magliano, D.J.; Shaw, J.E.; Gregg, E.W. Global trends in diabetes complications: A review of current evidence. Diabetologia 2019, 62, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Vanhoutte, P.M. Macro- and microvascular endothelial dysfunction in diabetes. J. Diabetes 2017, 9, 434–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strain, W.D.; Paldanius, P.M. Diabetes, cardiovascular disease and the microcirculation. Cardiovasc. Diabetol. 2018, 17, 57. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2014, 37 (Suppl. 1), S81–S90. [Google Scholar] [CrossRef] [Green Version]
- DiMeglio, L.A.; Evans-Molina, C.; Oram, R.A. Type 1 diabetes. Lancet 2018, 391, 2449–2462. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martin, C. Pathophysiology of Type 2 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 46–57. [Google Scholar] [CrossRef]
- Quan, W.; Jo, E.K.; Lee, M.S. Role of pancreatic beta-cell death and inflammation in diabetes. Diabetes Obes. Metab. 2013, 15 (Suppl. 3), 141–151. [Google Scholar] [CrossRef]
- Rorsman, P.; Ashcroft, F.M. Pancreatic beta-Cell Electrical Activity and Insulin Secretion: Of Mice and Men. Physiol. Rev. 2018, 98, 117–214. [Google Scholar] [CrossRef]
- Katsiki, N.; Ferrannini, E.; Mantzoros, C. New American Diabetes Association (ADA)/European Association for the Study of Diabetes (EASD) guidelines for the pharmacotherapy of type 2 diabetes: Placing them into a practicing physician’s perspective. Metabolism 2020, 107, 154218. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M. American Diabetes Association issues new guidelines for type 1 diabetes. BMJ 2014, 348, g4119. [Google Scholar] [CrossRef]
- Loghmani, H.; Conway, E.M. Exploring traditional and nontraditional roles for thrombomodulin. Blood 2018, 132, 148–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, T.; Taguchi, O.; Toda, M.; Ruiz, D.B.; Bernabe, P.G.; D’Alessandro-Gabazza, C.N.; Miyake, Y.; Kobayashi, T.; Aoki, S.; Chiba, F.; et al. Inhibition of allergic bronchial asthma by thrombomodulin is mediated by dendritic cells. Am. J. Respir. Crit. Care Med. 2011, 183, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Gabazza, E.C.; Taguchi, O.; Kamada, H.; Hayashi, T.; Adachi, Y.; Suzuki, K. Progress in the understanding of protease-activated receptors. Int. J. Hematol. 2004, 79, 117–122. [Google Scholar] [CrossRef]
- Roeen, Z.; Toda, M.; D’Alessandro-Gabazza, C.N.; Onishi, M.; Kobayashi, T.; Yasuma, T.; Urawa, M.; Taguchi, O.; Gabazza, E.C. Thrombomodulin inhibits the activation of eosinophils and mast cells. Cell Immunol. 2015, 293, 34–40. [Google Scholar] [CrossRef]
- Toda, M.; Shao, Z.; Yamaguchi, K.D.; Takagi, T.; D’Alessandro-Gabazza, C.N.; Taguchi, O.; Salamon, H.; Leung, L.L.; Gabazza, E.C.; Morser, J. Differential gene expression in thrombomodulin (TM; CD141)(+) and TM(-) dendritic cell subsets. PLoS ONE 2013, 8, e72392. [Google Scholar]
- Yang, S.M.; Ka, S.M.; Wu, H.L.; Yeh, Y.C.; Kuo, C.H.; Hua, K.F.; Shi, G.Y.; Hung, Y.J.; Hsiao, F.C.; Yang, S.S.; et al. Thrombomodulin domain 1 ameliorates diabetic nephropathy in mice via anti-NF-kappaB/NLRP3 inflammasome-mediated inflammation, enhancement of NRF2 antioxidant activity and inhibition of apoptosis. Diabetologia 2014, 57, 424–434. [Google Scholar] [CrossRef] [PubMed]
- Sharfuddin, A.A.; Sandoval, R.M.; Berg, D.T.; McDougal, G.E.; Campos, S.B.; Phillips, C.L.; Jones, B.E.; Gupta, A.; Grinnell, B.W.; Molitoris, B.A. Soluble thrombomodulin protects ischemic kidneys. J. Am. Soc. Nephrol. 2009, 20, 524–534. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Vinnikov, I.; Shahzad, K.; Bock, F.; Ranjan, S.; Wolter, J.; Kashif, M.; Oh, J.; Bierhaus, A.; Nawroth, P.; et al. The lectin-like domain of thrombomodulin ameliorates diabetic glomerulopathy via complement inhibition. Thromb. Haemost. 2012, 108, 1141–1153. [Google Scholar]
- Fujiwara, K.; Kobayashi, T.; Fujimoto, H.; Nakahara, H.; D’Alessandro-Gabazza, C.N.; Hinneh, J.A.; Takahashi, Y.; Yasuma, T.; Nishihama, K.; Toda, M.; et al. Inhibition of Cell Apoptosis and Amelioration of Pulmonary Fibrosis by Thrombomodulin. Am. J. Pathol. 2017, 187, 2312–2322. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, Y.; Ri, J.; Sakai, K.; Niki, K.; Funauchi, M.; Matsumura, I. Protective Effects of Recombinant Human Soluble Thrombomodulin on Lipopolysaccharide-Induced Acute Kidney Injury. Int. J. Mol. Sci. 2020, 21, 2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suyama, K.; Kawasaki, Y.; Miyazaki, K.; Kanno, S.; Ono, A.; Ohara, S.; Sato, M.; Hosoya, M. The efficacy of recombinant human soluble thrombomodulin for the treatment of shiga toxin-associated hemolytic uremic syndrome model mice. Nephrol. Dial. Transplant. 2015, 30, 969–977. [Google Scholar] [CrossRef] [Green Version]
- Takeshita, A.; Yasuma, T.; Nishihama, K.; D’Alessandro-Gabazza, C.N.; Toda, M.; Totoki, T.; Okano, Y.; Uchida, A.; Inoue, R.; Qin, L.; et al. Thrombomodulin ameliorates transforming growth factor-beta1-mediated chronic kidney disease via the G-protein coupled receptor 15/Akt signal pathway. Kidney Int. 2020, 98, 1179–1192. [Google Scholar] [CrossRef]
- Akatsuka, M.; Masuda, Y.; Tatsumi, H.; Sonoda, T. The effect of recombinant human soluble thrombomodulin on renal function and mortality in septic disseminated intravascular coagulation patients with acute kidney injury: A retrospective study. J. Intensive Care 2020, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Maruyama, I.; Shimazaki, S.; Yamamoto, Y.; Aikawa, N.; Ohno, R.; Hirayama, A.; Matsuda, T.; Asakura, H.; Nakashima, M.; et al. Efficacy and safety of recombinant human soluble thrombomodulin (ART-123) in disseminated intravascular coagulation: Results of a phase III, randomized, double-blind clinical trial. J. Thromb. Haemost. 2007, 5, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Katayama, S.; Nunomiya, S.; Koyama, K.; Wada, M.; Koinuma, T.; Goto, Y.; Tonai, K.; Shima, J. Markers of acute kidney injury in patients with sepsis: The role of soluble thrombomodulin. Crit. Care 2017, 21, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuma, T.; Yano, Y.; D’Alessandro-Gabazza, C.N.; Toda, M.; Gil-Bernabe, P.; Kobayashi, T.; Nishihama, K.; Hinneh, J.A.; Mifuji-Moroka, R.; Roeen, Z.; et al. Amelioration of Diabetes by Protein S. Diabetes 2016, 65, 1940. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro-Gabazza, C.N.; Kobayashi, T.; Yasuma, T.; Toda, M.; Kim, H.; Fujimoto, H.; Hataji, O.; Takeshita, A.; Nishihama, K.; Okano, T.; et al. A Staphylococcus pro-apoptotic peptide induces acute exacerbation of pulmonary fibrosis. Nat. Commun. 2020, 11, 1539. [Google Scholar] [CrossRef]
- Sramek, J.; Nemcova-Furstova, V.; Kovar, J. Molecular Mechanisms of Apoptosis Induction and Its Regulation by Fatty Acids in Pancreatic beta-Cells. Int. J. Mol. Sci. 2021, 22, 4285. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, K.; Nagai, R. Islet inflammation in type 2 diabetes and physiology. J. Clin. Invest. 2017, 127, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Powers, A.C. Type 1 diabetes mellitus: Much progress, many opportunities. J. Clin. Investig. 2021, 131, e142242. [Google Scholar] [CrossRef] [PubMed]
- Donath, M.Y. Targeting inflammation in the treatment of type 2 diabetes. Diabetes Obes. Metab. 2013, 15 (Suppl. 3), 193–196. [Google Scholar] [CrossRef] [PubMed]
- Honzawa, N.; Fujimoto, K. The Plasticity of Pancreatic beta-Cells. Metabolites 2021, 11, 218. [Google Scholar] [CrossRef]
- In’t Veld, P. Insulitis in human type 1 diabetes: A comparison between patients and animal models. Semin. Immunopathol. 2014, 36, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Herzog, C.; Lorenz, A.; Gillmann, H.J.; Chowdhury, A.; Larmann, J.; Harendza, T.; Echtermeyer, F.; Muller, M.; Schmitz, M.; Stypmann, J.; et al. Thrombomodulin’s lectin-like domain reduces myocardial damage by interfering with HMGB1-mediated TLR2 signalling. Cardiovasc. Res. 2014, 101, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Hirao, S.; Minakata, K.; Masumoto, H.; Yamazaki, K.; Ikeda, T.; Minatoya, K.; Sakata, R. Recombinant human soluble thrombomodulin prevents acute lung injury in a rat cardiopulmonary bypass model. J. Thorac. Cardiovasc. Surg. 2017, 154, 1973–1983. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Wang, X.; Nishioka, C.; Honda, G.; Yokoyama, A.; Zeng, L.; Xu, K.; Ikezoe, T. G-protein coupled receptor 15 mediates angiogenesis and cytoprotective function of thrombomodulin. Sci. Rep. 2017, 7, 692. [Google Scholar] [CrossRef]
- Takada, S.; Miyashita, T.; Yamamoto, Y.; Kanou, S.; Munesue, S.; Ohbatake, Y.; Nakanuma, S.; Okamoto, K.; Sakai, S.; Kinoshita, J.; et al. Soluble Thrombomodulin Attenuates Endothelial Cell Damage in Hepatic Sinusoidal Obstruction Syndrome. In Vivo 2018, 32, 1409–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanemura, A.; Kuriyama, N.; Azumi, Y.; Ohsawa, I.; Kishiwada, M.; Mizuno, S.; Usui, M.; Sakurai, H.; Tabata, M.; Isaji, S. Thrombomodulin administration attenuates ischemia-reperfusion injury of the remnant liver after 70% hepatectomy in rats: Simulated model of small-for-size graft in living donor liver transplantation. Transplant. Proc. 2014, 46, 1107–1111. [Google Scholar] [CrossRef]
- Brooks-Worrell, B.M.; Boyko, E.J.; Palmer, J.P. Impact of islet autoimmunity on the progressive beta-cell functional decline in type 2 diabetes. Diabetes Care 2014, 37, 3286–3293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchetti, P. Islet inflammation in type 2 diabetes. Diabetologia 2016, 59, 668–672. [Google Scholar] [CrossRef]
- King, A.J. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. 2021, 1, e78. [Google Scholar] [CrossRef]
- Ikezoe, T.; Yang, J.; Nishioka, C.; Yokoyama, A. Thrombomodulin alleviates murine GVHD in association with an increase in the proportion of regulatory T cells in the spleen. Bone Marrow Transplant. 2015, 50, 113–120. [Google Scholar] [CrossRef]
- Lindley, S.; Dayan, C.M.; Bishop, A.; Roep, B.O.; Peakman, M.; Tree, T.I. Defective suppressor function in CD4(+)CD25(+) T-cells from patients with type 1 diabetes. Diabetes 2005, 54, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Iwashita, Y.; Zhang, E.; Maruyama, J.; Yokochi, A.; Yamada, Y.; Sawada, H.; Mitani, Y.; Imai, H.; Suzuki, K.; Maruyama, K. Thrombomodulin protects against lung damage created by high level of oxygen with large tidal volume mechanical ventilation in rats. J. Intensive Care 2014, 2, 57. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Saka, R.; Takama, Y.; Ueno, T.; Tazuke, Y.; Okuyama, H. Recombinant human soluble thrombomodulin reduces the severity and incidence of necrotizing enterocolitis in a newborn rat model. Surg. Today 2019, 49, 971–976. [Google Scholar] [CrossRef]
- Pathak, R.; Wang, J.; Garg, S.; Aykin-Burns, N.; Petersen, K.U.; Hauer-Jensen, M. Recombinant Thrombomodulin (Solulin) Ameliorates Early Intestinal Radiation Toxicity in a Preclinical Rat Model. Radiat. Res 2016, 186, 112–120. [Google Scholar] [CrossRef]
- Rajashekhar, G.; Gupta, A.; Marin, A.; Friedrich, J.; Willuweit, A.; Berg, D.T.; Cramer, M.S.; Sandusky, G.E.; Sutton, T.A.; Basile, D.P.; et al. Soluble thrombomodulin reduces inflammation and prevents microalbuminuria induced by chronic endothelial activation in transgenic mice. Am. J. Physiol. Renal. Physiol. 2012, 302, F703–F712. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Terai, Y.; Daimon, A.; Nunode, M.; Nagayasu, Y.; Okamoto, A.; Fujita, D.; Hayashi, M.; Ohmichi, M. Recombinant human soluble thrombomodulin as an anticoagulation therapy improves recurrent miscarriage and fetal growth restriction due to placental insufficiency—The leading cause of preeclampsia. Placenta 2018, 65, 1–6. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okano, Y.; Takeshita, A.; Yasuma, T.; Toda, M.; Nishihama, K.; Fridman D’Alessandro, V.; Inoue, C.; D’Alessandro-Gabazza, C.N.; Kobayashi, T.; Yano, Y.; et al. Protective Role of Recombinant Human Thrombomodulin in Diabetes Mellitus. Cells 2021, 10, 2237. https://doi.org/10.3390/cells10092237
Okano Y, Takeshita A, Yasuma T, Toda M, Nishihama K, Fridman D’Alessandro V, Inoue C, D’Alessandro-Gabazza CN, Kobayashi T, Yano Y, et al. Protective Role of Recombinant Human Thrombomodulin in Diabetes Mellitus. Cells. 2021; 10(9):2237. https://doi.org/10.3390/cells10092237
Chicago/Turabian StyleOkano, Yuko, Atsuro Takeshita, Taro Yasuma, Masaaki Toda, Kota Nishihama, Valeria Fridman D’Alessandro, Chisa Inoue, Corina N. D’Alessandro-Gabazza, Tetsu Kobayashi, Yutaka Yano, and et al. 2021. "Protective Role of Recombinant Human Thrombomodulin in Diabetes Mellitus" Cells 10, no. 9: 2237. https://doi.org/10.3390/cells10092237
APA StyleOkano, Y., Takeshita, A., Yasuma, T., Toda, M., Nishihama, K., Fridman D’Alessandro, V., Inoue, C., D’Alessandro-Gabazza, C. N., Kobayashi, T., Yano, Y., & Gabazza, E. C. (2021). Protective Role of Recombinant Human Thrombomodulin in Diabetes Mellitus. Cells, 10(9), 2237. https://doi.org/10.3390/cells10092237