
Derivation of Oligodendrocyte Precursors from Adult Bone Marrow Stromal Cells for Remyelination Therapy


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Bone Marrow Extraction, aMSC Culture and Cryopreservation
2.3. aMSC Proliferation Assay
2.4. Neural Progenitor Derivation
2.5. Differentiation of Neural Progenitors along the OL Lineage and Cryopreservation
2.6. aMSC-Derived OP Proliferation Test
2.7. Isolation and Primary Culture of Rat OPs
2.8. aMSC-Derived OP-Neuron Co-Culture
2.9. Transplantation of aMSC-Derived OPs
2.10. Immunofluorescence
2.11. Sample Preparation and Immunohistochemistry
2.12. Transmisison Electron Microscopy
2.13. Flow Cytometry
2.14. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.15. Statistical Analysis
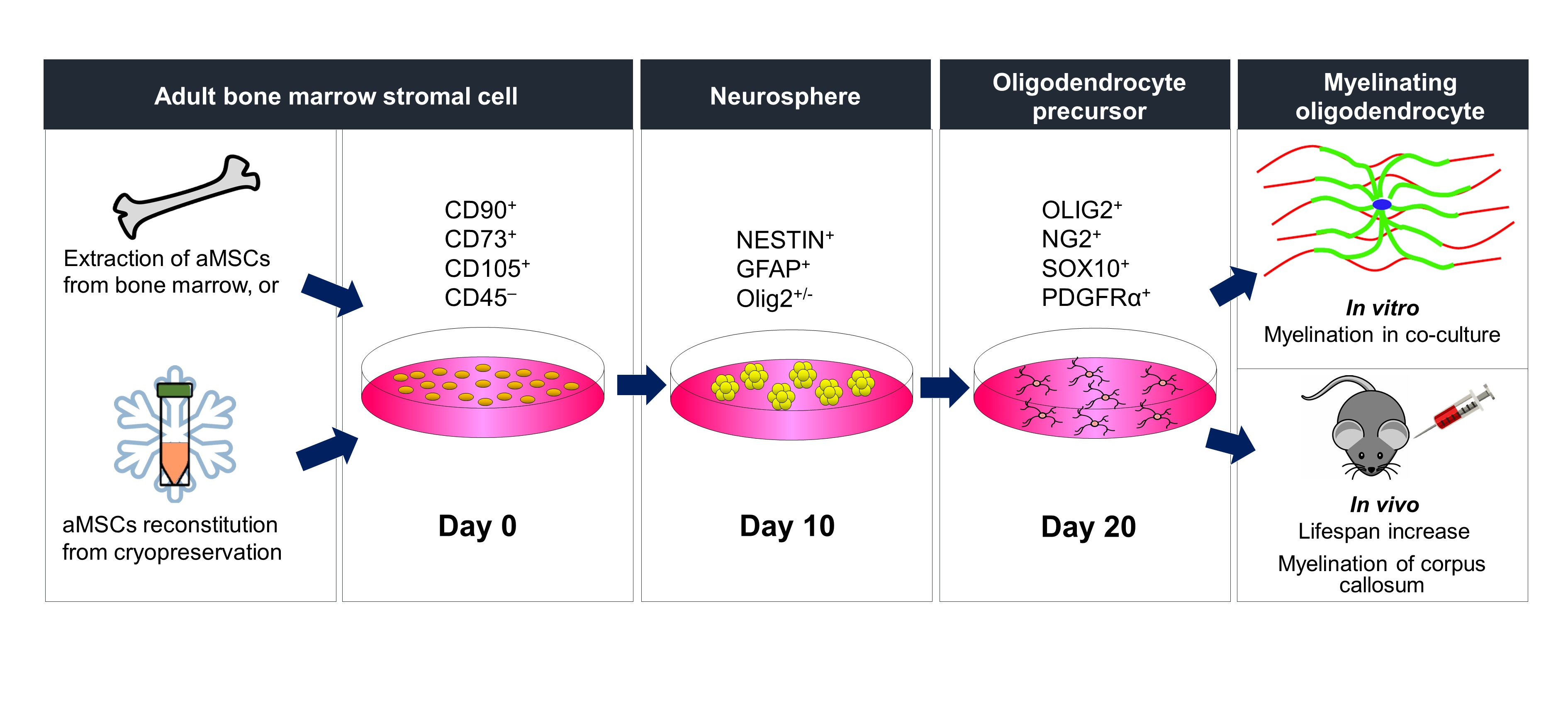
3. Results
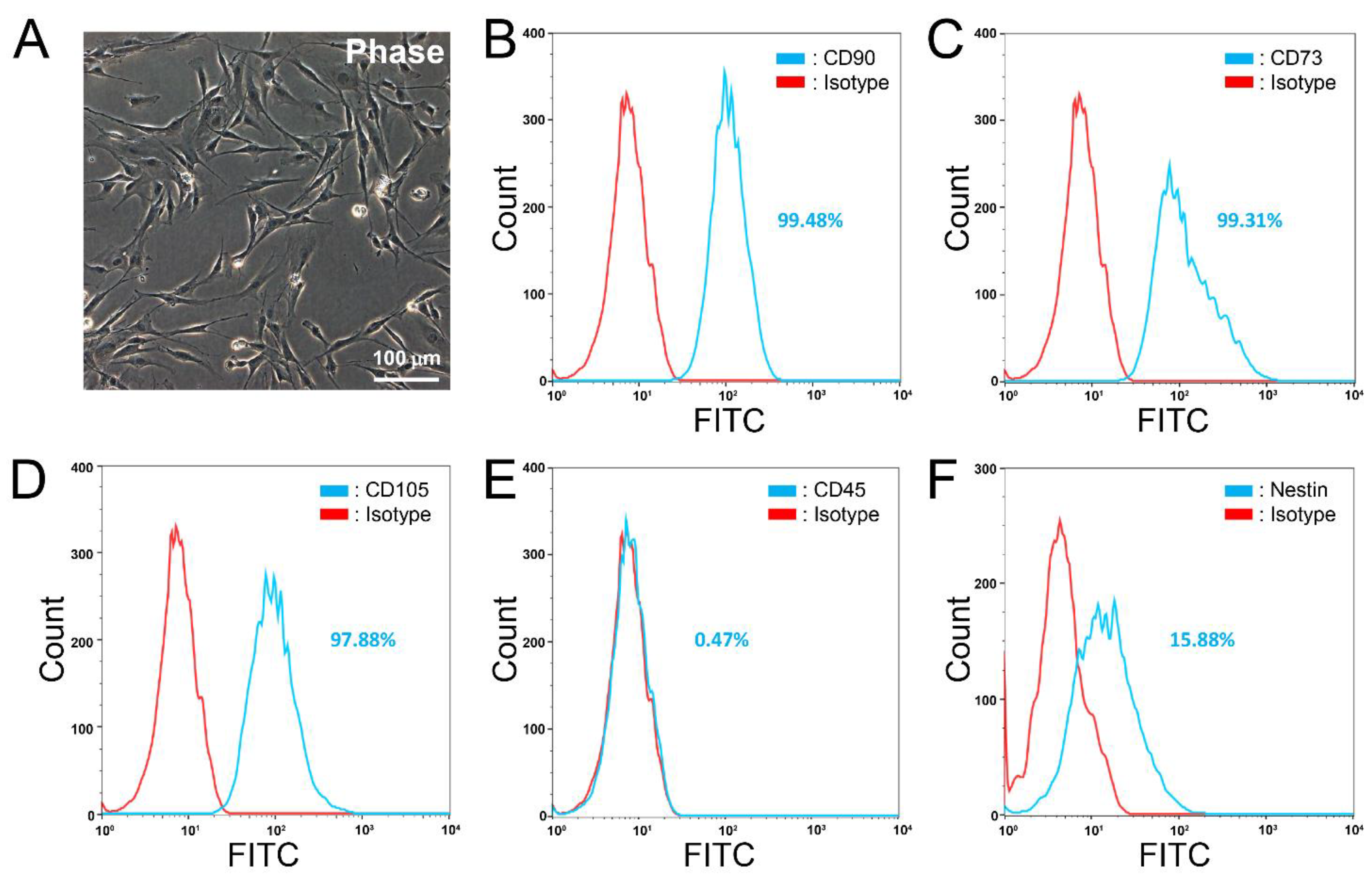
3.1. Characterization of Primary and Cryopreserved aMSCs
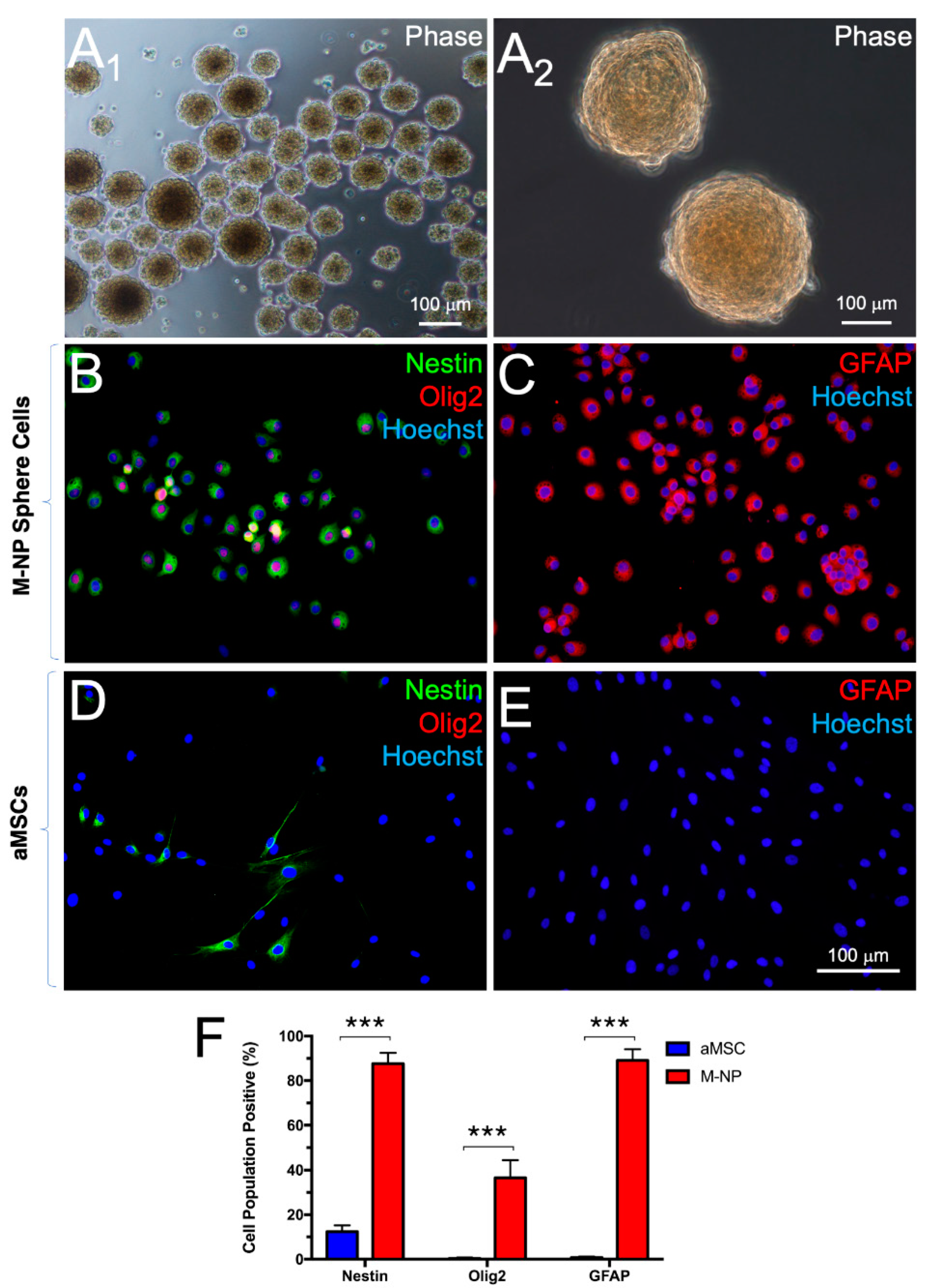
3.2. Derivation of Neural Progenitors through Non-Adherent Culture
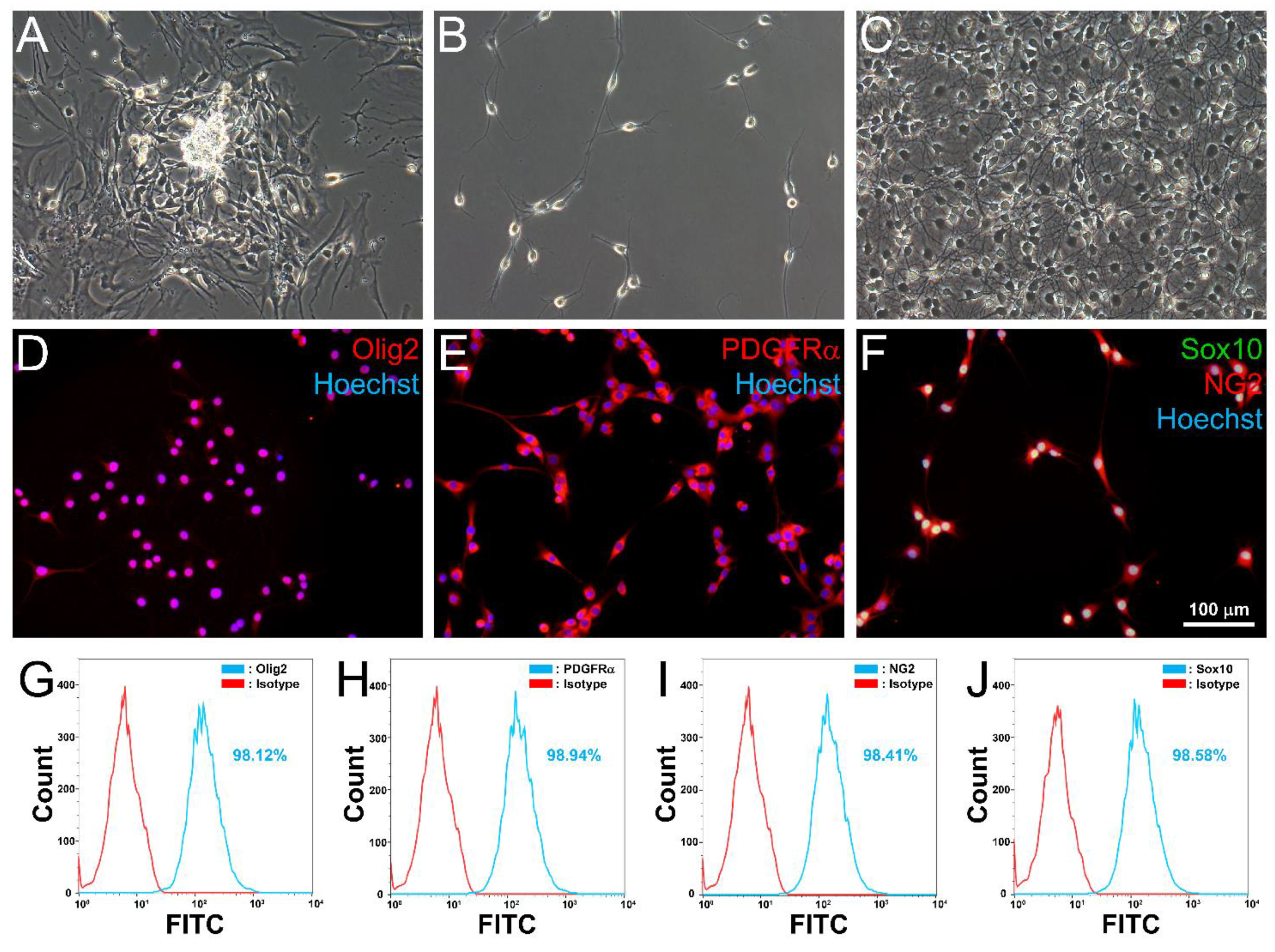
3.3. Differentiation of aMSC-Derived Neural Progenitors along the OL Lineage
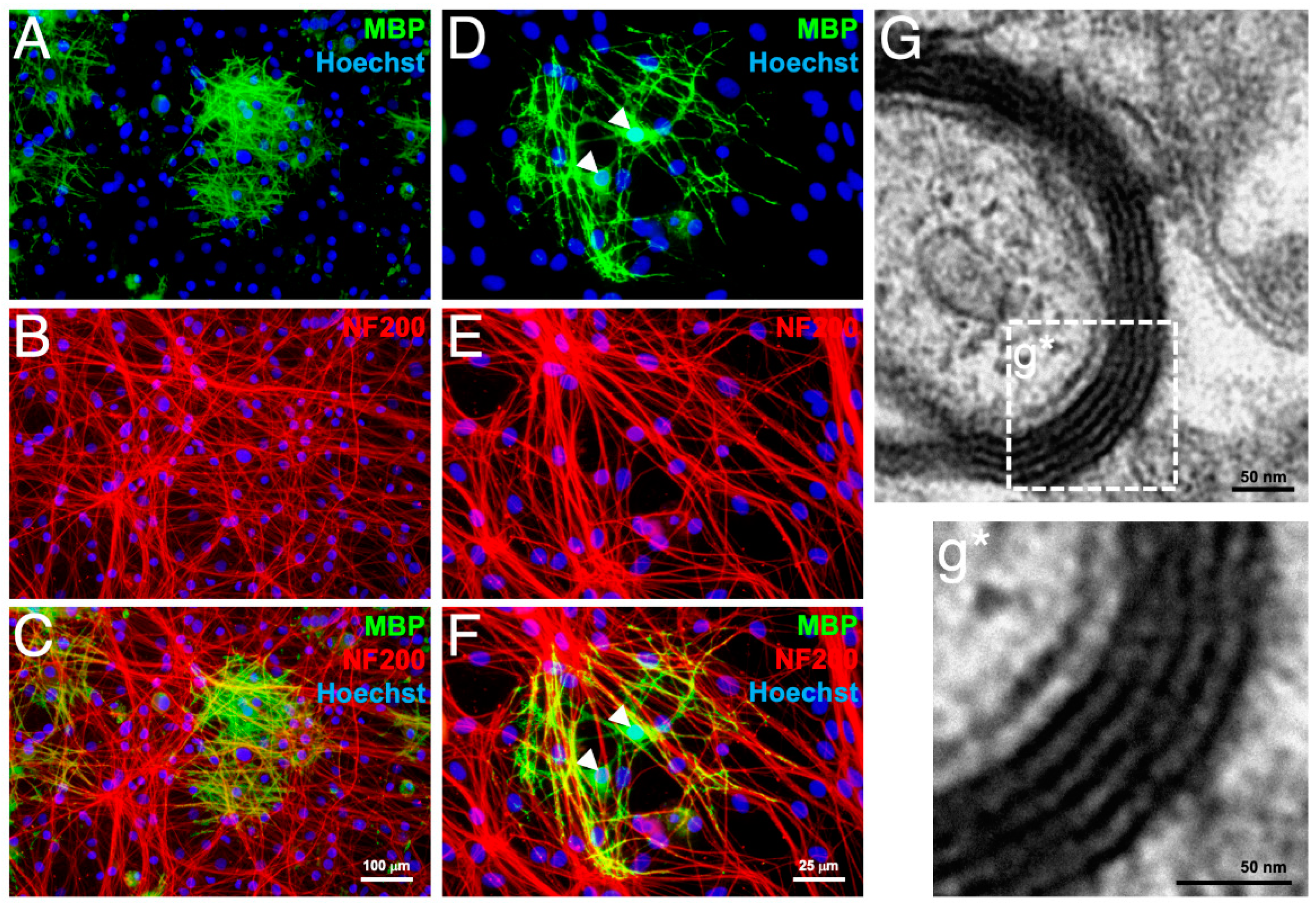
3.4. In Vitro Myelination
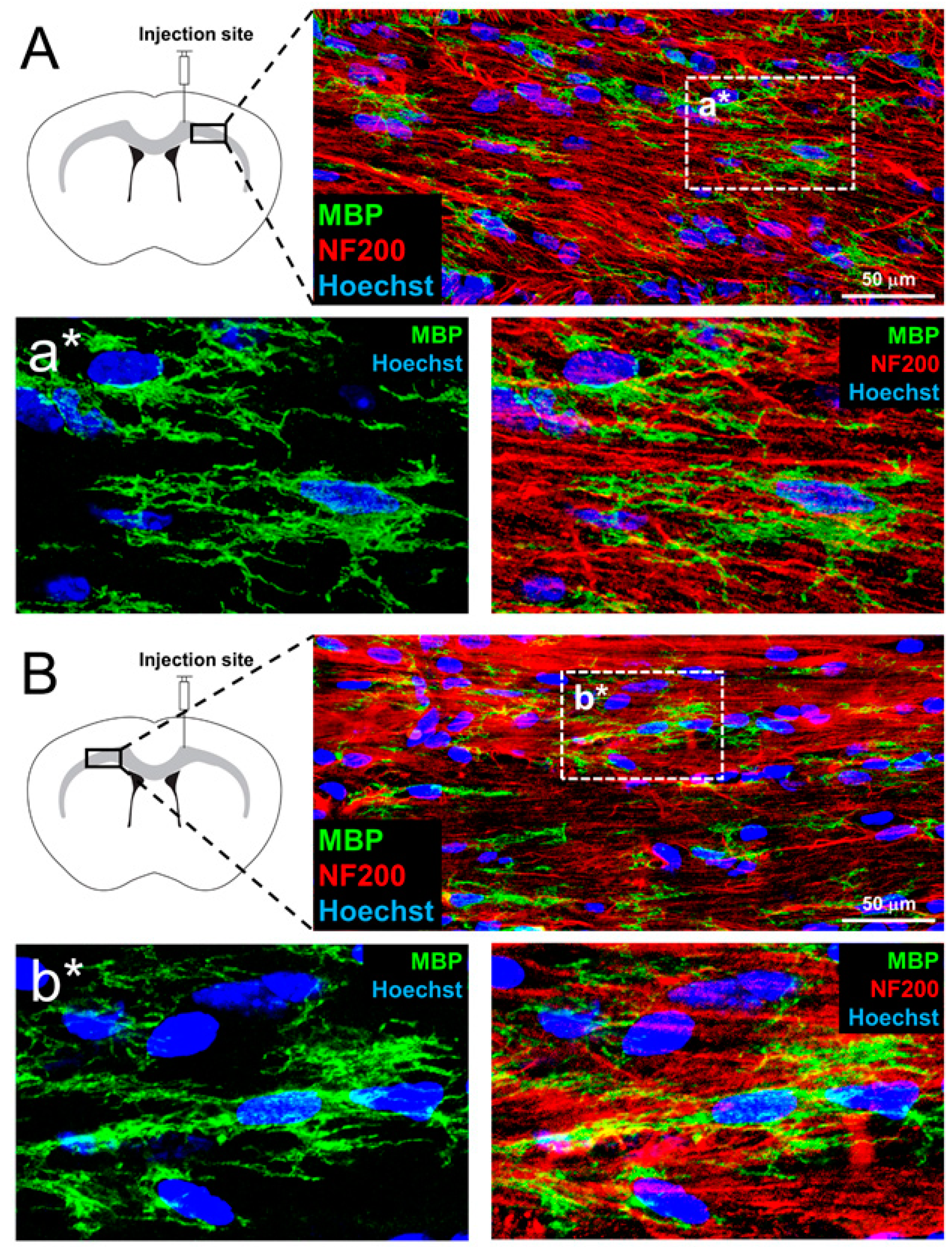
3.5. In Vivo Myelination and Extended Lifespan Follwing Transplantation of OPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Franklin, R.J.M.; ffrench-Constant, C. Remyelination in the CNS: From biology to therapy. Nat. Rev. Neurosci. 2008, 9, 839–855. [Google Scholar] [CrossRef]
- Goldman, S.A. Stem and progenitor cell-based therapy of the central nervous system: Hopes, hype, and wishful thinking. Cell Stem Cell 2016, 18, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Stadelmann, C.; Timmler, S.; Barrantes-Freer, A.; Simons, M. Myelin in the central nervous system: Structure, function and pathology. Physiol. Rev. 2019, 99, 1381–1431. [Google Scholar] [CrossRef]
- Cao, Q.; Xu, X.M.; DeVries, W.H.; Enzmann, G.U.; Ping, P.; Tsoulfas, P.; Wood, P.M.; Bunge, M.B.; Whittemore, S.R. Functional recovery in traumatic spinal cord injury after transplantation of multineurotrophin-expressing glial-restricted precursor cells. J. Neurosci. 2005, 25, 6947–6957. [Google Scholar] [CrossRef] [PubMed]
- Windrem, M.S.; Schanz, S.J.; Guo, M.; Tian, G.F.; Washco, V.; Stanwood, N.; Rasband, M.; Roy, N.S.; Nedergaard, M.; Havton, L.A.; et al. Neonatal chimerization with human glial progenitor cells can both remyelinate and rescue the otherwise lethally hypomyelinated shiverer mouse. Cell Stem Cell 2008, 2, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Sharp, J.; Frame, J.; Siegenthaler, M.; Nistor, G.; Keirstead, H.S. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants improve recovery after cervical spinal cord injury. Stem Cells 2010, 28, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Sim, F.J.; McClain, C.R.; Schanz, S.J.; Protack, T.L.; Windrem, M.S.; Goldman, S.A. CD140a identify a population of highly myelinogenic, migration competent and efficiently engrafting human oligodendrocyte progenitor cells. Nat. Biotech. 2011, 29, 934–941. [Google Scholar] [CrossRef]
- Kalladka, D.; Sinden, J.; Pollock, K.; Haig, C.; McLean, J.; Smith, W.; McConnachie, A.; Santosh, C.; Bath, P.M.; Dunn, L.; et al. Human neural stem cells in patients with chronic ischaemic stroke (PISCES): A phase 1, first-in-man study. Lancet 2016, 388, 787–796. [Google Scholar] [CrossRef]
- Kim, H.J.; Seo, S.W.; Chang, J.W.; Lee, J.I.; Kim, C.H.; Chin, J.; Choi, S.J.; Kwon, H.; Yun, H.J.; Lee, J.M.; et al. Stereotactic brain injection of human umbilical cord blood mesenchymal stem cells in patients with Alzheimer’s disease dementia: A phase 1 clinical trial. Alzheimers Dement. (N. Y.) 2015, 1, 95–102. [Google Scholar] [CrossRef]
- Duma, C.; Kopyov, O.; Kopyov, A.; Berman, M.; Lander, E.; Elam, M.; Arata, M.; Weiland, D.; Cannell, R.; Crawat, C.; et al. Human intracerebroventricular (ICV) injection of autologous, non-engineered, adipose-derived stromal vascular fraction (ADSVF) for neurodegenerative disorders: Results of a 3-year phase 1 study of 113 injections in 31 patients. Mol. Biol. Rep. 2019, 46, 5257–5272. [Google Scholar] [CrossRef]
- Hyun, I. The bioethics of stem cell research and therapy. J. Clin. Investig. 2010, 120, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Lo, B.; Parham, L. Ethical issues in stem cell research. Endocr. Rev. 2009, 30, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Bates, J.; Li, X.; Schanz, S.; Chandler-Militello, D.; Levine, C.; Maherali, N.; Studer, L.; Hochedlinger, K.; Windrem, M.; et al. Human iPSC-derived oligodendrocyte progenitors can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell 2013, 12, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Windrem, M.S.; Osipovitch, M.; Liu, Z.; Bates, J.; Chandler-Militello, D.; Zou, L.; Munir, J.; Schanz, S.; McCoy, K.; Miller, R.H.; et al. Human iPSC glial mouse chimeras reveal glial contributions to schizophrenia. Cell Stem Cell 2017, 21, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, M.; Nevin, Z.S.; Shick, H.E.; Garrison, E.; Clarkson-Paredes, C.; Karl, M.; Clayton, B.L.L.; Factor, D.C.; Allan, K.C.; Barbar, L.; et al. Induction of myelinating oligodendrocytes in human cortical spheroids. Nat. Methods. 2018, 15, 700–706. [Google Scholar] [CrossRef]
- Tsui, Y.P.; Shea, G.K.H.; Chan, Y.S.; Shum, D.K.Y. Derivation of fate committed Schwann cells from adult rat bone marrow-derived neural progenitors. Methods Mol. Biol. 2018, 1739, 137–148. [Google Scholar]
- Shea, G.K.H.; Tai, E.W.Y.; Leung, K.H.Y.; Mung, A.K.L.; Li, M.T.S.; Tsui, A.Y.P.; Tam, A.K.W.; Shum, D.K.Y.; Chan, Y.S. Juxtacrine signalling via Notch and ErbB receptors in the switch to fate commitment of bone marrow-derived Schwann cells. Eur. J. Neurosci. 2020, 52, 3306–3321. [Google Scholar] [CrossRef] [PubMed]
- Mung, K.L.; Tsui, Y.P.; Tai, E.W.Y.; Chan, Y.S.; Shum, D.K.Y.; Shea, G.K.H. Rapid and efficient generation of neural progenitors from adult bone marrow stromal cells by hypoxic preconditioning. Stem Cell Res. Ther. 2016, 7, 146. [Google Scholar] [CrossRef]
- Cai, S.; Tsui, Y.P.; Tam, K.W.; Shea, G.K.H.; Chang, R.S.K.; Ao, Q.; Shum, D.K.Y.; Chan, Y.S. Directed differentiation of human bone marrow stromal cells to fate-committed schwann cells. Stem Cell Rep. 2017, 9, 1097–1108. [Google Scholar] [CrossRef]
- Cai, S.; Han, L.; Ao, Q.; Chan, Y.S.; Shum, D.K.Y. Human induced pluripotent stem cell-derived sensory neurons for fate commitment of bone marrow-derived Schwann cells: Implications for remyelination therapy. Stem Cells Trans. Med. 2016, 6, 1–13. [Google Scholar]
- Ao, Q.; Fung, C.K.; Tsui, Y.P.; Cai, S.; Zuo, H.C.; Chan, Y.S.; Shum, D.K.Y. The regeneration of transected sciatic nerves of adult rats using chitosan nerve conduits seeded with bone marrow stromal cell-derived Schwann cells. Biomaterials 2011, 32, 787–796. [Google Scholar] [CrossRef] [PubMed]
- García-León, J.A.; García-Díaz, B.; Eggermont, K.; Cáceres-Palomo, L.; Neyrinck, K.; Madeiro da Costa, R.; Dávila, J.C.; Baron-Van Evercooren, A.; Gutiérrez, A.; Verfaillie, C.M. Generation of oligodendrocytes and establishment of an all-human myelinating platform from human pluripotent stem cells. Nat. Protoc. 2020, 15, 3716–3744. [Google Scholar] [CrossRef] [PubMed]
- Nolbrant, S.; Heuer, A.; Parmar, M.; Kirkeby, A. Generation of high-purity human ventral midbrain dopaminergic progenitors for in vitro maturation and intracerebral transplantation. Nat. Protoc. 2017, 12, 1962–1979. [Google Scholar] [CrossRef] [PubMed]
- Lendahl, U.; Zimmerman, L.B.; McKay, R.D.G. CNS stem cells express a new class of intermediate filament protein. Cell 1990, 60, 585–595. [Google Scholar] [CrossRef]
- Lu, Q.R.; Sun, T.; Zhu, Z.; Ma, N.; Garcia, M.; Stiles, C.D.; Rowitch, D.H. Common developmental requirement of Olig function indicates a motor neuron/oligodendrocyte connection. Cell 2002, 109, 75–86. [Google Scholar] [CrossRef]
- Zhou, Q.; Anderson, D.J. The bHLH transcription factors Olig2 and Olig1 couple neuronal and glial subtype specification. Cell 2002, 109, 61–73. [Google Scholar] [CrossRef]
- Chen, Y.; Balasubramaniyan, V.; Peng, J.; Hurlock, E.C.; Tallquist, M.; Li, J.; Lu, Q.R. Isolation and culture of rat and mouse oligodendrocyte precursor cells. Nat. Protocol. 2007, 2, 1044–1051. [Google Scholar] [CrossRef]
- Rao, R.C.; Boyd, J.; Padmanabhan, R.; Chenoweth, J.G.; McKay, R.D. Efficient serum-free derivation of oligodendrocyte precursors from neural stem cell-enriched cultures. Stem Cells 2009, 27, 116–125. [Google Scholar] [CrossRef][Green Version]
- Hu, J.G.; Fu, S.L.; Wang, Y.X.; Li, Y.; Jiang, X.Y.; Wang, X.F.; Wang, X.F.; Qiu, M.S.; Lu, P.H.; Xu, X.M. Platelet-derived growth factor-AA mediates oligodendrocyte lineage differentiation through activation of extracellular signal-regulated kinase signaling pathway. Neuroscience 2008, 151, 138–147. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, R.D.; Smith, C.; Behar, T.; Smith, T.; Dubois-Dalcq, M. Distinct effects of bFGF and PDGF on oligodendrocyte progenitor cells. Glia 1993, 7, 45–254. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.R.; Watkins, T.; Cosgaya, J.M.; Zhang, C.; Chen, L.; Reichardt, L.F.; Shooter, E.M.; Barres, B.A. NGF controls axonal receptivity to myelination by Schwann cells or oligodendrocytes. Neuron 2004, 43, 183–191. [Google Scholar] [CrossRef]
- Yang, N.; Zuchero, J.B.; Ahlenius, H.; Marro, S.; Ng, Y.H.; Vierbuchen, T.; Hawkins, J.S.; Geissler, R.; Barres, B.A.; Wernig, M. Generation of oligodendroglial cells by direct lineage conversion. Nat. Biotechnol. 2013, 31, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Milling, S.; Frede, A. Teaching tolerance: Diverse cellular interactions enable healthy maturation. Immunology 2021, 163, 237–238. [Google Scholar]
- Graca, L. The contribution of B cells to transplantation tolerance. J. Clin. Investig. 2020, 130, 3406–3408. [Google Scholar] [CrossRef]
- Meyer, N.; Richter, N.; Fan, Z.; Siemonsmeier, G.; Pivneva, T.; Jordan, P.; Steinhäuser, C.; Semtner, M.; Nolte, C.; Kettenmann, H. Oligodendrocytes in the mouse corpus callosum maintain axonal function by delivery of glucose. Cell Rep. 2018, 22, 2383–2394. [Google Scholar] [CrossRef] [PubMed]
- Mount, C.W.; Yalçın, B.; Cunliffe-Koehler, K.; Sundaresh, S.; Monje, M. Monosynaptic tracing maps brain-wide afferent oligodendrocyte precursor cell connectivity. Elife 2019, 8, e49291. [Google Scholar] [CrossRef]
- De Faria, O., Jr.; Pama, E.A.C.; Evans, K.; Luzhynskaya, A.; Káradóttir, R.T. Neuroglial interactions underpinning myelin plasticity. Dev. Neurobiol. 2018, 78, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Káradóttir, R.T.; Kuo, C.T. Neuronal activity-dependent control of postnatal neurogenesis and gliogenesis. Annu. Rev. Neurosci. 2018, 41, 139–161. [Google Scholar] [CrossRef]
- Jia, W.; Kamen, Y.; Pivonkova, H.; Káradóttir, R.T. Neuronal activity-dependent myelin repair after stroke. Neurosci. Lett. 2019, 703, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Singer, W.; Dietz, A.B.; Zeller, A.D.; Gehrking, T.L.; Schmelzer, J.D.; Schmeichel, A.M.; Gehrking, J.A.; Suarez, M.D.; Sletten, D.M.; Minota Pacheco, K.V.; et al. Intrathecal administration of autologous mesenchymal stem cells in multiple system atrophy. Neurology 2019, 93, e77–e87. [Google Scholar] [CrossRef] [PubMed]
- Bain, B.J. Bone marrow biopsy morbidity and mortality. Br. J. Haematol. 2003, 121, 949–951. [Google Scholar] [CrossRef] [PubMed]
- Malempati, S.; Joshi, S.; Lai, S.; Braner, D.A.V.; Tegtmeyer, K. Bone marrow aspiration and biopsy. N. Engl. J. Med. 2009, 361, e28. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsui, Y.-P.; Lam, G.; Wu, K.L.-K.; Li, M.T.-S.; Tam, K.-W.; Shum, D.K.-Y.; Chan, Y.-S. Derivation of Oligodendrocyte Precursors from Adult Bone Marrow Stromal Cells for Remyelination Therapy. Cells 2021, 10, 2166. https://doi.org/10.3390/cells10082166
Tsui Y-P, Lam G, Wu KL-K, Li MT-S, Tam K-W, Shum DK-Y, Chan Y-S. Derivation of Oligodendrocyte Precursors from Adult Bone Marrow Stromal Cells for Remyelination Therapy. Cells. 2021; 10(8):2166. https://doi.org/10.3390/cells10082166
Chicago/Turabian StyleTsui, Yat-Ping, Guy Lam, Kenneth Lap-Kei Wu, Maximilian Tak-Sui Li, Kin-Wai Tam, Daisy Kwok-Yan Shum, and Ying-Shing Chan. 2021. "Derivation of Oligodendrocyte Precursors from Adult Bone Marrow Stromal Cells for Remyelination Therapy" Cells 10, no. 8: 2166. https://doi.org/10.3390/cells10082166
APA StyleTsui, Y.-P., Lam, G., Wu, K. L.-K., Li, M. T.-S., Tam, K.-W., Shum, D. K.-Y., & Chan, Y.-S. (2021). Derivation of Oligodendrocyte Precursors from Adult Bone Marrow Stromal Cells for Remyelination Therapy. Cells, 10(8), 2166. https://doi.org/10.3390/cells10082166