
Role of Glycoproteins during Fruit Ripening and Seed Development




Abstract
1. Introduction
2. Different Glycosylation Types
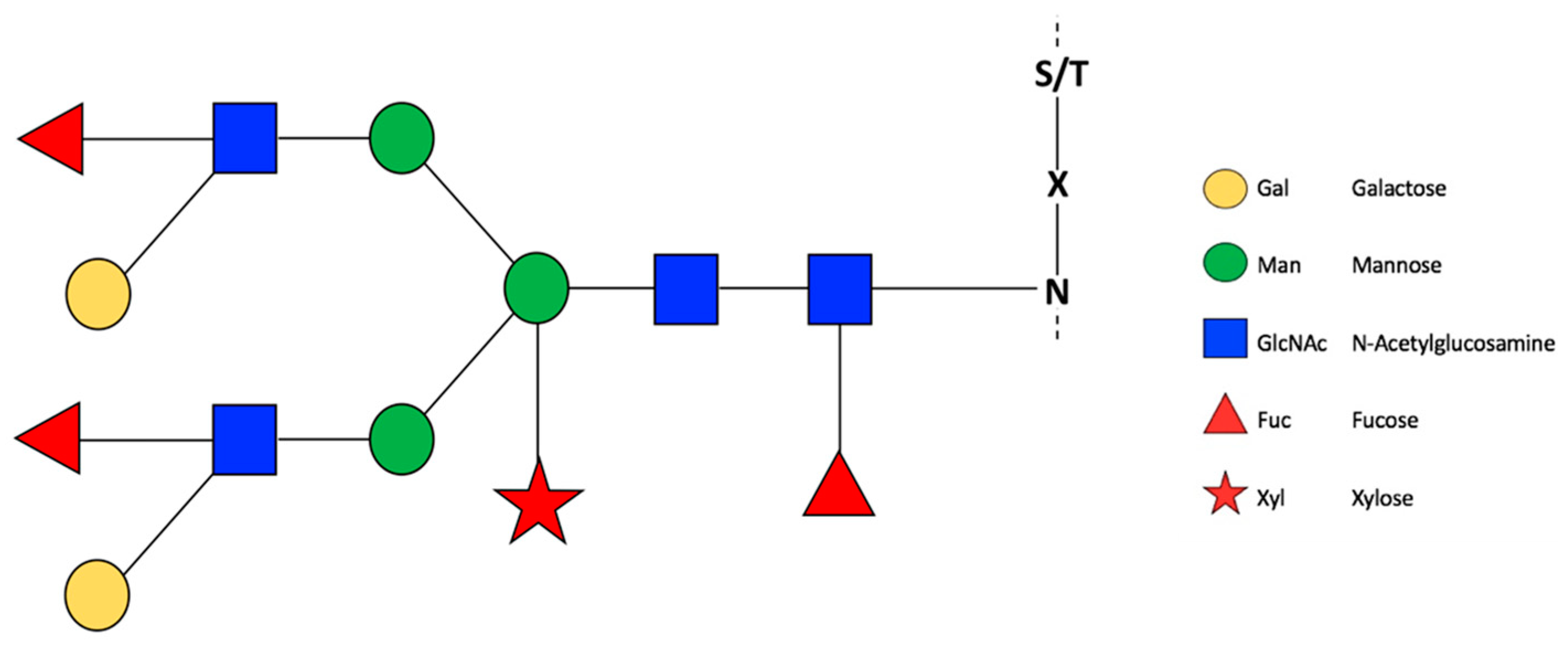
2.1. N-Glycosylations
2.1.1. N-Glycosylation and Plant Development
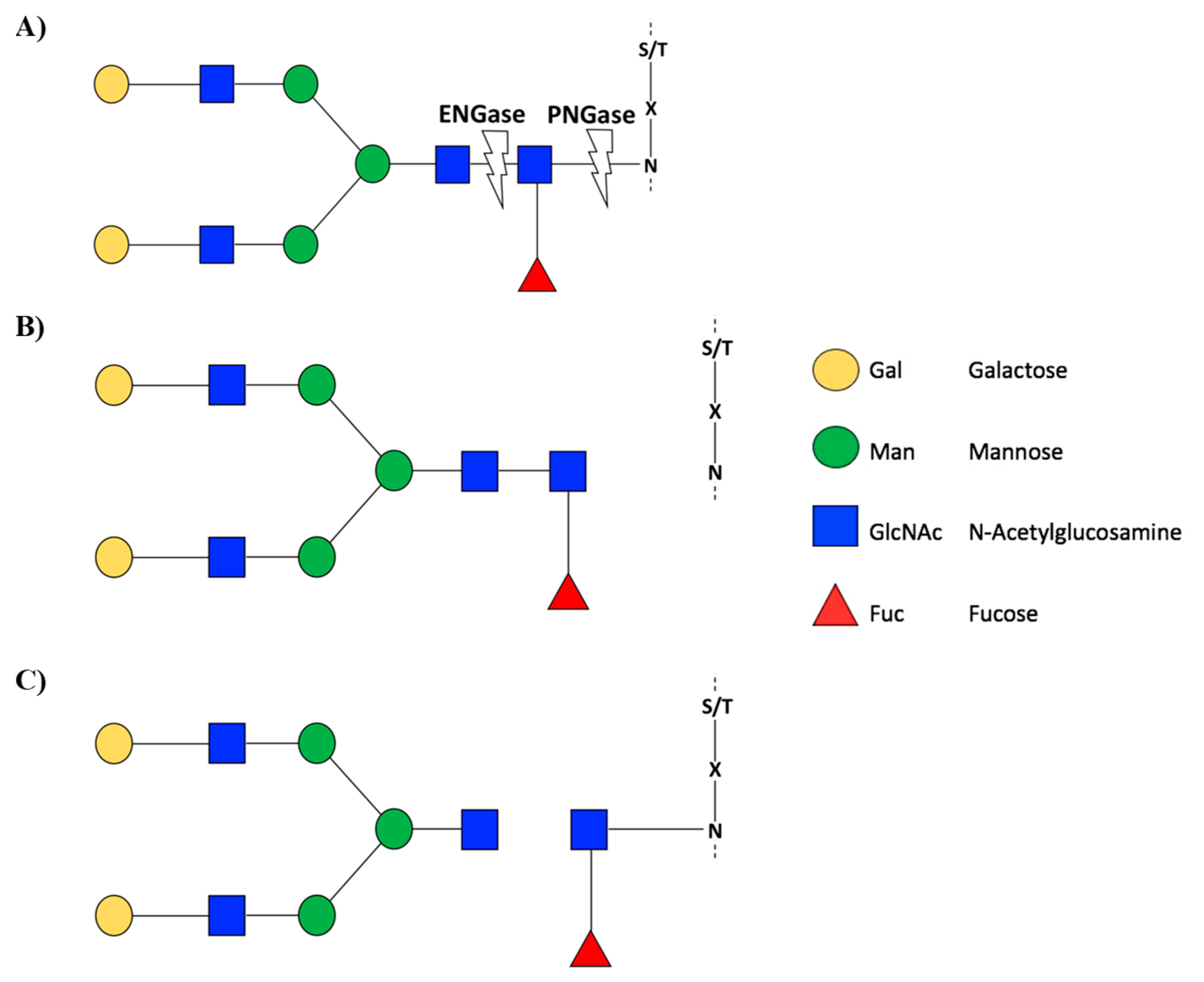
2.1.2. N-Glycosylation and Fruit Ripening
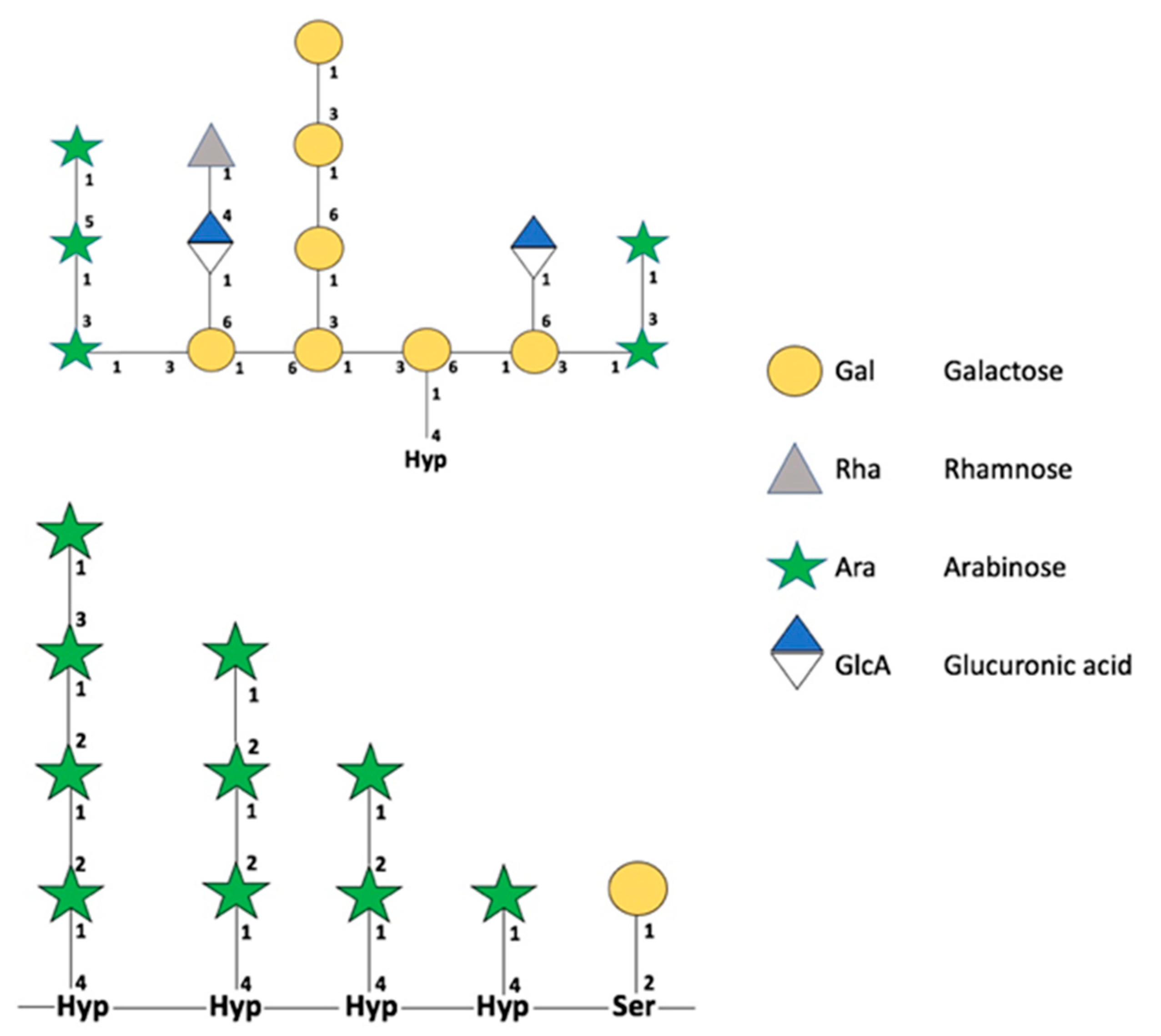
2.2. O-Glycoproteins in Fruits and a World Not Yet Explored
Reports of O-Glycosylations in Plants
3. Experimental Techniques to Identify N- and O-Glycosylations
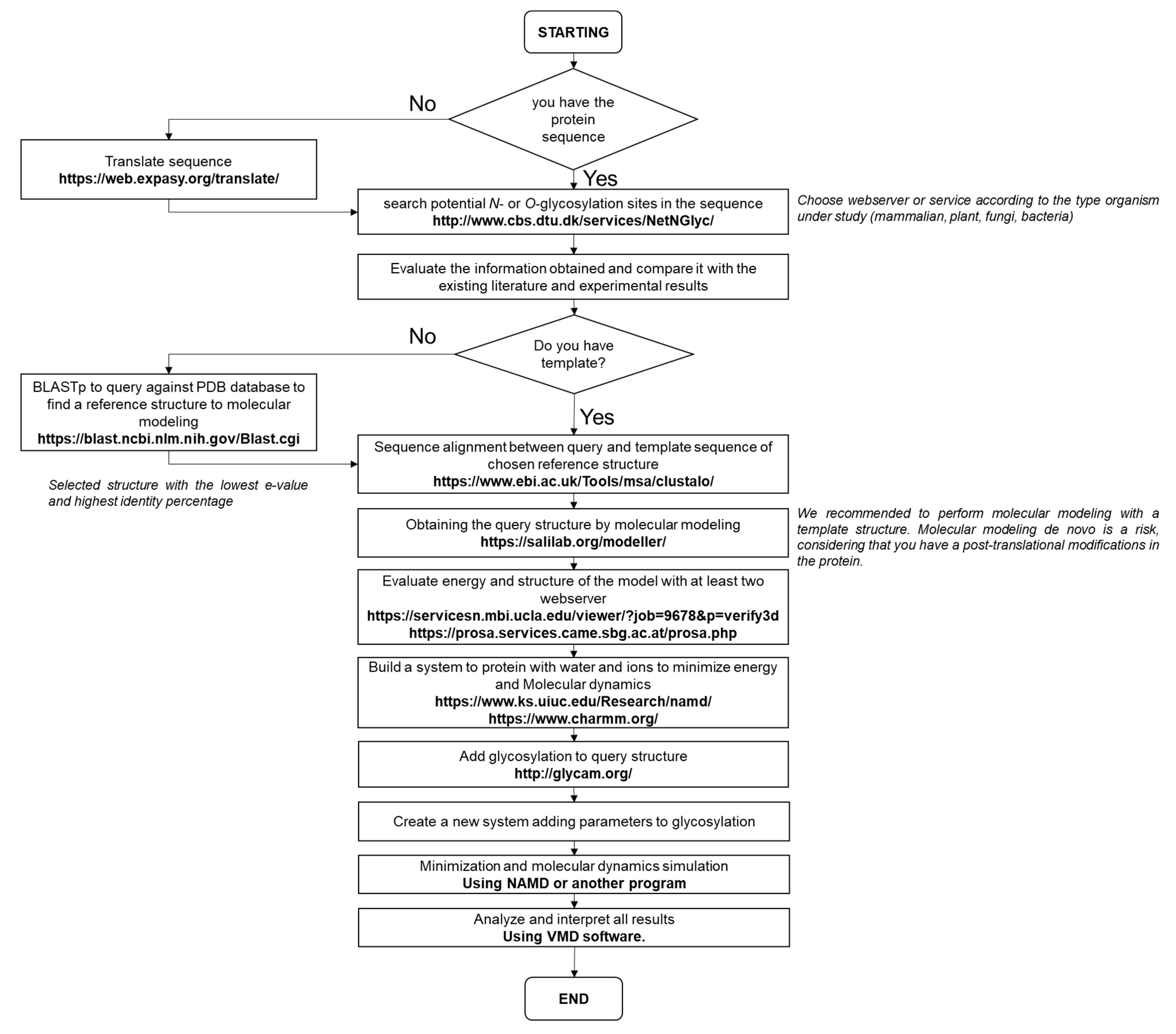
4. Bioinformatics Glycosylations Tools in Plants
5. New Perspectives and Challenges
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, P.; Li, X.; Wang, Y.; Wang, W.; Tian, S.; Qin, G. Redox proteomic analysis reveals the involvement of oxidative post-translational modification in tomato fruit ripening. Postharvest Biol. Technol. 2021, 178, 111556. [Google Scholar] [CrossRef]
- Hebert, D.N.; Lamriben, L.; Powers, E.T.; Kelly, J.W. The intrinsic and extrinsic effects of N-linked glycans on glycoproteostasis. Nat. Chem. Biol. 2014, 10, 902–910. [Google Scholar] [CrossRef]
- Nagashima, Y.; von Schaewen, A.; Koiwa, H. Function of N-glycosylation in plants. Plant Sci. 2018, 274, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Inoue, M.; Maeda, M.; Nakano, R.; Hosoi, K.; Fijiyama, K.; Kimura, Y. Molecular cloning and gene expression analysis of tomato endo-β-N-acetylglucosaminidase, an endoglycosidase involved in the production of high-mannose type free N-glycans during tomato fruit ripening. Biosci. Biotechnol. Biochem. 2009, 73, 461–464. [Google Scholar] [CrossRef]
- Méndez-Yáñez, A.; Beltrán, D.; Campano-Romero, C.; Molinett, S.; Herrera, R.; Moya-León, M.A.; Morales-Quintana, L. Glycosylation is important for FcXTH1 activity as judged by its structural and biochemical characterization. Plant Physiol. Biochem. 2017, 119, 200–210. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, H.; Du, H.; Liu, Z.; Bao, Z.; Shi, Q. Comparative N-glycoproteome analysis provides novel insights into the regulation mechanism in tomato (Solanum lycopersicum L.) During fruit ripening process. Plant Sci. 2020, 293, 110413. [Google Scholar] [CrossRef] [PubMed]
- Pattison, R.J.; Amtmann, A. N-glycan production in the endoplasmic reticulum of plants. Trends Plant Sci. 2009, 14, 92–99. [Google Scholar] [CrossRef]
- Tegl, G.; Nidetzky, B. Leloir glycosyltransferases of natural product C-glycosylation: Structure, mechanism and specificity. Biochem. Soc. Trans. 2020, 48, 1583–1598. [Google Scholar] [CrossRef]
- Gorres, K.L.; Raines, R.T. Prolyl 4-hydroxylase. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 106–124. [Google Scholar] [CrossRef]
- Lannoo, N.; Van Damme, E.J. Review/N-glycans: The making of a varied toolbox. Plant Sci. 2015, 239, 67–83. [Google Scholar] [CrossRef]
- Strasser, R. Plant protein glycosylation. Glycobiology 2016, 26, 926–939. [Google Scholar] [CrossRef]
- Kern, N.R.; Lee, H.S.; Wu, E.L.; Park, S.; Vanommeslaeghe, K.; MacKerell, A.; Klauda, J.B.; Jo, S.; Im, W. Lipid-linked oligosaccharides in membranes sample conformations that facilitate binding to oligosaccharyltransferase. Biophys. J. 2014, 107, 1885–1895. [Google Scholar] [CrossRef]
- Strasser, R. Biological significance of complex N-glycans in plants and their impact on plant physiology. Front. Plant Sci. 2014, 5, 363. [Google Scholar] [CrossRef]
- Montero-Morales, L.; Steinkellner, H. Advanced plant-based glycan engineering. Front. Bioeng. Biotechnol. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Ford, K.L.; Bacic, A.; Heazlewood, J.L. N-linked glycan micro-heterogeneity in glycoproteins of arabidopsis. Mol. Cell. Proteom. 2018, 17, 413–421. [Google Scholar] [CrossRef]
- Fan, F.; Zhang, Q.; Lu, D. Identification of N-glycosylation sites on AtERO1 and AtERO2 using a transient expression system. Biochem. Biophys. Res. Commun. 2020, 533, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Geißler, M.F. Metabolic Engineering of Cannabinoid Biosynthesis in Tobacco. Ph.D. Thesis, Technische Universität Darmstadt, Dramstadt, Germany, 2021. [Google Scholar]
- Li, X.; Cheng, M.; Tang, C.; Zhu, X.; Qi, K.; Zhang, S.; Wu, J.; Peng, W. Identification and function analysis of fascicles-like arabinogalactan rptoein family genes in pear (Pyrus bretschneideri). Plant Syst. Evol. 2021, 307, 48. [Google Scholar] [CrossRef]
- Wenderoth, I.; von Schaewen, A. Isolation and characterization of plant N-acetyl glucosaminyltransferase I (GntI) cDNA sequences. Functional analyses in the arabidopsis cgl mutant and in antisense plants. Plant Physiol. 2000, 123, 1097–1108. [Google Scholar] [CrossRef]
- Limkul, J.; Misaki, R.; Kato, K.; Fujiyama, K. The combination of plant translational enhancers and terminator increase the expression of human glucocerebrosidase in Nicotiana benthamiana plants. Plant Sci. 2015, 240, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.; Stadlmann, J.; Schähs, M.; Stiegler, G.; Quendler, H.; Mach, L.; Glössl, J.; Weterings, K.; Pabst, M.; Steinkellner, H. Generation of glyco-engineered Nicotiana benthamiana for the production of monoclonal antibodies with a homogeneous human-like N-glycan structure. Plant Biotechnol. J. 2008, 6, 392–402. [Google Scholar] [CrossRef]
- Mutanwad, K.V.; Zangl, I.; Lucyshyn, D. Arabidopsis O-fucosyltransferase SPINDLY regulates root hair patterning independently of gibberellin signaling. Development 2020, 147. [Google Scholar] [CrossRef]
- Jiao, Q.-S.; Niu, G.-T.; Wang, F.-F.; Dong, J.-Y.; Chen, T.-S.; Zhou, C.-F.; Hong, Z. N-glycosylation regulates photosynthetic efficiency of Arabidopsis thaliana. Photosynthesis 2020, 58, 72–79. [Google Scholar] [CrossRef]
- Ajayi, O.O.; Held, M.A.; Showalter, A.M. Two β-glucuronosyltransferases involved in the biosynthesis of type II arabinogalactans function in mucilage polysaccharide matrix organization in Arabidopsis thaliana. BMC Plant Biol. 2021, 21, 1–19. [Google Scholar] [CrossRef]
- Kallas, A.M.; Piens, K.; Denman, S.E.; Henriksson, H.; Faldt, J.; Johansson, P.; Brumer, H.; Teeri, T.T. Enzymatic properties of native and deglycosylated hybrid aspen (Populus tremulaxtremuloides) xyloglucan endotransglycosylase 16A expressed in Pichia Pastor. Biochem. J. 2005, 390, 105–113. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bose, S.K.; He, Y.; Howlader, P.; Wang, W.; Yin, H. The N-glycan processing enzymes beta-D-N acetylhexosaminidase are involved in ripening-associated softening in strawberry fruit. J. Food Sci. Technol. 2020, 58, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Bi, Y.; Shrestha, R.; Zhang, Z.; Burlingame, A.; Wang, Z. Nutrient sensing in plants by O-GlcNAcylation and O-Fucosylation. FASEB J. 2021, 35. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Castillo, N.; Lara, J.A.; Arró, M.; Boronat, A.; Ferrer, A.; Altabella, T. Tomato UDP-glucose sterol glycosyltransferases: A family of developmental and stress regulated genes that encode cytosolic and membrane-associated forms of the enzyme. Front. Plant Sci. 2017, 8, 984. [Google Scholar] [CrossRef] [PubMed]
- Trempel, F.; Kajiura, H.; Ranf, S.; Grimmer, J.; Westphal, L.; Zipfel, C.; Scheel, D.; Fujiyama, K.; Lee, J. Altered glycosylation of exported proteins, including surface immune receptors, compromises calcium and downstream signaling responses to microbe-associated molecular patterns in Arabidospis thaliana. BMC Plant Biol. 2016, 16, 31. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Qing, X.; Liao, J.; Zhuo, K. Role of protein glycosylation in host-pathogen interaction. Cells 2020, 9, 1022. [Google Scholar] [CrossRef] [PubMed]
- Kaulfürst-Soboll, H.; Mertens-Beer, M.; Brehler, R.; Albert, M.; von Schaewen, A. Complex n-glycans are important for normal fruit ripening and seed development in tomato. Front. Plant Sci. 2021, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, H.; Gai, J.; Tian, X.; Zhang, X.; Lv, Y.; Jian, Y. Evolution of protein N-glycosylation process in Golgi apparatus which shapes diversity of protein N-glycan structures in plants, animals and fungi. Sci. Rep. 2017, 7, 40301. [Google Scholar] [CrossRef]
- Ghosh, S.; Meli, V.S.; Kumar, A.; Thakur, A.; Chakraborty, N.; Chakraborty, S.; Datta, A. The N-glycan processing enzymes α-mannosidase and β-D-N-acetylhexosaminidase are involved in ripening-associated softening in the non-climateric fruits of capsicum. J. Exp. Bot. 2010, 62, 571–582. [Google Scholar] [CrossRef]
- Meli, V.S.; Ghosh, S.; Prabha, T.N.; Chakraborty, N.; Chakraborty, S.; Datta, A. Enhancement of N-glycan processing enzymes. Proc. Natl. Acad. Sci. USA 2010, 107, 2413–2418. [Google Scholar] [CrossRef]
- Varki, A. Biological roles of glycans. Glycobiology 2016, 27, 3–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tang, H.; Du, H.; Bao, Z.; Shi, Q. Sugar metabolic and N-glycosylated profiles unveil the regulatory mechanism of tomato quality under salt stress. Environ. Exp. Bot. 2020, 177, 104145. [Google Scholar] [CrossRef]
- Faugeron, C.; Mollet, J.-C.; Karamanos, Y.; Morvan, H. Activities of de-N-glycosylation are ubiquitously found in tomato plant. Acta Physiol. Plant. 2006, 28, 557–565. [Google Scholar] [CrossRef]
- Karav, S.; Casaburi, G.; Arslan, A.; Kaplan, M.; Sucu, B.; Frese, S. N-glycans from human milk glycoproteins are selectively released by an infant gut symbiont in vivo. J. Funct. Foods 2019, 61. [Google Scholar] [CrossRef]
- Fairbanks, A.J. The ENGases: Versatile biocatalysts for the production of homogeneous N-linked glycopeptides and glycoproteins. Chem. Soc. Rev. 2017, 46, 5128–5146. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.; Braam, J. In vitro activities of four xyloglucan endotransglycosylases from Arabidopsis. Plant J. 1999, 18, 371–382. [Google Scholar] [CrossRef]
- Vaaje-Kolstad, G.; Farkaš, V.; Hrmova, M.; Fincher, G.B. Xyloglucan xyloglucosyl transferases from barley (Hordeum vulgare L.) bind oligomeric and polymeric xyloglucan molecules in their acceptor binding sites. Biochim. Biophys. Acta Gen. Subj. 2010, 1800, 674–684. [Google Scholar] [CrossRef]
- Torres, C.R.; Hart, G.W. Topography and polypeptide distribution of terminal N-acetylglucosamine residues on the surfaces of intact lymphocytes. Evidence for O-linked GlcNAc. J. Biol. Chem. 1984, 259, 3308–3317. [Google Scholar] [CrossRef]
- Seifert, G.J. On the potential function of type II arabinogalactan O-glycosylation in regulating the fate of plant secretory proteins. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, S.M.; Ricardi, M.M.; Dorosz, J.G.; Fernandez, P.V.; Nadra, A.D.; Pol-Fachin, L.; Egelund, J.; Gille, S.; Harholt, J.; Ciancia, M.; et al. O-glycosylated cell wall proteins are essential in root hair growth. Science 2011, 332, 1401–1403. [Google Scholar] [CrossRef]
- Tan, L.; Varnai, P.; Lamport, D.T.; Yuan, C.; Xu, J.; Qiu, F.; Kieliszewski, M.J. Plant O-hydroxyproline arabinogalactans are composed of repeating trigalactosyl subunits with short bifurcated side chains. J. Biol. Chem. 2010, 285, 24575–24583. [Google Scholar] [CrossRef] [PubMed]
- Nguema-Ona, E.; Vicré-Gibouin, M.; Gotté, M.; Plancot, B.; Lerouge, P.; Bardor, M.; Driouich, A. Cell wall O-glycoproteins and N-glycoproteins: Aspects of biosynthesis and function. Front. Plant Sci. 2014, 5, 499. [Google Scholar] [CrossRef]
- Ruíz-May, E.; Thannhauser, T.W.; Zhang, S.; Rose, J.K.C. Analytical technologies for identification and characterization of the plant N-glycoproteome. Front. Plant Sci. 2012, 3, 150. [Google Scholar] [CrossRef]
- Riley, N.M.; Hebert, A.S.; Westphall, M.S.; Coon, J.J. Capturing site-specific heterogeneity with large-scale N-glycoproteome analysis. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Stavenhagen, K.; Hinneburg, H.; Kolarich, D.; Wuhrer, M. Site-specific N and O-Glycopeptide analysis using an integrated C18-PGC-LC-ESI-QTOF-MS/MS approach. Methods Mol. Biol. 2016, 1503, 109–119. [Google Scholar] [CrossRef]
- An, H.J.; Froehlich, J.W.; Lebrilla, C.B. Determination of glycosylation sites and site-specific heterogeneity in glycoproteins. Curr. Opin. Chem. Biol. 2009, 13, 421–426. [Google Scholar] [CrossRef]
- Wilkinson, H.; Saldova, R. Current methods for the characterization of O-glycans. J. Proteome Res. 2020, 19, 3890–3905. [Google Scholar] [CrossRef]
- Mewono, L.; Nguema-Ona, E.; Gotté, M.; Koroney, A.S.; Gueye, M.F.; Driouich, A.; Vicré-Gibouin, M.; Aboughe-Angone, S. O-glycosylation in plant and mammal cells: The use of chemical inhibitors to understand the biosynthesis and function of O-glycosylated proteins. Plant Sci. Today 2015, 2, 43–51. [Google Scholar]
- Muthu, M.; Chun, S.; Gopal, J.; Anthonydhason, V.; Haga, S.W.; Devadoss, A.J.P.; Oh, J. Insights into bionformatic applications for glycosylation: Instigating an awakening towards applying glycoinformatic resources for cancer diagnosis and therapy. Int. J. Mol. Sci. 2020, 21, 9336. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.; Schloissnig, S. Bioinformatics and molecular modeling in glycobiology. Cell. Mol. Life Sci. 2010, 67, 2749–2772. [Google Scholar] [CrossRef]
- Park, S.; Lee, J.; Qi, Y.; Kern, N.R.; Lee, H.S.; Jo, S.; Joung, I.; Joo, K.; Lee, J.; Im, W. CHARMM-GUI Glycan Modeler for modeling and simulation of carbohydrates and glycoconjugates. Glycobiology 2019, 29, 320–331. [Google Scholar] [CrossRef]
- Turupcu, A.; Diem, M.; Smith, L.J.; Oostenbrink, C. Structural aspects of the O-glycosylation linkage in glycopeptides via MD simulations and comparison with NMR experiments. Phys. Chem. 2019, 20, 1527–1537. [Google Scholar] [CrossRef]
- Johnson, K.L.; Cassin, A.M.; Lonsdale, A.; Bacic, A.; Doblin, M.S.; Schultz, C.J. Pipeline to identify hydroxyproline-rich glycoproteins. Plant Physiol. 2017, 174, 886–903. [Google Scholar] [CrossRef]
- Willems, P.; Horne, A.; Van Parys, T.; Goormachtig, S.; De Smet, I.; Botzki, A.; Van Breusegem, F.; Gevaert, K. The plant PTM viewer, a central resource for exploring plant protein modifications. Plant J. 2019, 99, 752–762. [Google Scholar] [CrossRef]
- Lemmin, T.; Soto, C. Glycosylator: A phyton framework for the rapid modeling of glycans. BMC Bioinform. 2019, 20, 513. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme or Protein Type | Organism | Enzyme | Function | Reference |
|---|---|---|---|---|
| Glycosylated enzymes and proteins | Arabidopsis thaliana | AtERO1 | Identification of N-glycosylation sites of ER oxidoreductin-1. | [16] |
| Arabidopsis thaliana | AtERO2 | Identification of N-glycosylation sites of ER oxidoreductin-2. | [16] | |
| Nicotiana benthamiana | Cannabichromenic acid synthase | Mutagenesis is a glycosylated enzyme to produce tetrahydrocannabinolic acid in tobacco. | [17] | |
| Pyrus bretschneideri | Fasciclin-like arabinogalactan protein family | Identification and function analysis of this family of proteins. | [18] | |
| Transferase | Solanum tuberosum L. Nicotiana tabacum L. Arabidopsis thaliana | N-acetyl glucosaminyltransferase I | Enzyme that begins the complex N-linked glycans. | [19] |
| Nicotiana benthamiana | N-acetyl glucosaminyltransferase I | Glycoengineering to produce human enzyme glucocerebrosidase | [20] | |
| Nicotiana benthamiana | α1,3-fucosyltransferase | Inhibition to produce human-like N-glycan structure. | [21] | |
| Nicotiana benthamiana | β1,2-xylosyltransferase | Inhibition to produce human-like N-glycan structure. | [21] | |
| Arabidopsis thaliana | O-fucosyltransferase | O-glycosylation regulates transitions in developmental stages. | [22] | |
| Arabidopsis thaliana | cgl1-1 | Knockout of N-acetyl glucosaminyltransferase I. | [23] | |
| Arabidopsis thaliana | β-glucuronosyltransferase | Identification of two β-glucuronosyltransferase involved in the biosynthesis of mucilage polysaccharides. | [24] | |
| Arabidopsis thaliana | alg3-3 | Knockout of α1,3-mannosyltransferase | [23] | |
| Transglycosylase | Populus tremula x tremuloides | PtXTH1 | Enzyme that modifies the cellulose-xyloglucan network during wood formation. | [25] |
| Fragaria chiloensis | FcXTH1 | Enzyme that modifies the cellulose-xyloglucan network during fruit ripening. | [5] | |
| Hydrolase | Fragaria x ananassa | β-Hex | Influence of β-Hex in the strawberry ripening | [26] |
| O-glycosylation | Arabidopsis thaliana | O-glycosylation screening in plant peptides | Identification of 262 proteins O-glycosylated with regulatory functions. | [27] |
| Nicotiana benthamiana | Human Granulocyte-Colony Stimulating Factor | Transient expression via agroinfiltration | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendez-Yañez, A.; Ramos, P.; Morales-Quintana, L. Role of Glycoproteins during Fruit Ripening and Seed Development. Cells 2021, 10, 2095. https://doi.org/10.3390/cells10082095
Mendez-Yañez A, Ramos P, Morales-Quintana L. Role of Glycoproteins during Fruit Ripening and Seed Development. Cells. 2021; 10(8):2095. https://doi.org/10.3390/cells10082095
Chicago/Turabian StyleMendez-Yañez, Angela, Patricio Ramos, and Luis Morales-Quintana. 2021. "Role of Glycoproteins during Fruit Ripening and Seed Development" Cells 10, no. 8: 2095. https://doi.org/10.3390/cells10082095
APA StyleMendez-Yañez, A., Ramos, P., & Morales-Quintana, L. (2021). Role of Glycoproteins during Fruit Ripening and Seed Development. Cells, 10(8), 2095. https://doi.org/10.3390/cells10082095