Abstract
Advances in screening methods and pharmacological treatments are increasing the life expectancy of cancer patients. During recent decades, the community of long-term disease-free cancer survivors (LCS) has grown exponentially, raising the issues related to cancer follow-up. Cancer relapse and other cancer-related diseases, as well as lifestyle, influence cancer survival. Recently, the regulatory role of microRNAs (miRNAs) in gene expression and their involvement in human diseases, including cancer, has been identified. Extracellular circulating miRNAs (ECmiRNAs) have been found in biological fluids and specific ECmiRNAs have been associated with cancer development and progression or with a therapy response. Here, we focus on the pivotal role of ECmiRNAs as biomarkers in cancer diagnosis and prognosis. Then, we discuss the relevance of ECmiRNAs expression in cancer survivors for the identification of specific ECmiRNAs profiles as potential tools to assess cancer outcome and to control LCS follow-up.
1. Introduction
The International Agency for Research on Cancer (IARC) recently described cancer as a globally spreading disease involving 20% of the worldwide population and causing high mortality, ranging from 12% in men to 9% in women [1]. Cancer is identified as the second leading cause of death, but improvements in early tumor diagnosis combined with new therapeutic treatments are boosting patient survival [2]. Since 2015 in the European Union, total cancer mortality rates have resulted in a 5-year mortality decline of about 5% in men and 4% in women [3].
Cancer survivors or long-term disease-free cancer survivors (LCS) are a growing community of cancer patients with a life expectancy of several years after the diagnosis. According to the IARC evaluation of December 2020, more than 50 million people worldwide are alive 5 years after cancer is diagnosed [4]. The survival rate after cancer diagnosis is not only related to cancer recurrence and to a healthy lifestyle, but also to the development of diseases deriving from cancer treatments. Therefore, risk factors associated with the neoplastic diseases influence cancer survival [5,6], as described in a recent study showing an increased risk of cardiovascular diseases in survivors of different cancer types [7].
MicroRNAs (miRNAs) have been identified as biomarkers for human disorders, including cancer [8]. These 18–23 nucleotides-long RNA sequences regulate cellular gene expression influencing several biological processes, such as cell proliferation, mitochondrial functions and organ development [9,10]. After annealing to complementary mRNA targets, miRNAs interfere with mRNAs translation or promote their degradation [11]. Since their regulatory role, alterations of intracellular miRNAs have been associated to several diseases other than cancers, including cardiovascular illnesses [12]. Moreover, miRNAs have been identified extracellularly in many biological fluids (i.e., blood, urine, tears) [13,14]. The association of extracellular circulating miRNAs (ECmiRNAs) with extracellular vesicles (EVs), high density lipoproteins (HDLs) and proteins, protects them from degradation and ensures the ECmiRNAs stability in RNAse-rich biological fluids, such as blood [15]. Stable ECmiRNAs act as mediators in cell-to-cell communication, modulating the biological processes of neighboring cells by gene expression regulation, such as the establishment of a favorable niche around the primary tumor, within the tumor microenvironment [16]. ECmiRNAs expression has been suggested as a biomarker in pathological conditions: for example, decreased levels of miR-17-5p and miR-20a were observed in the plasma of breast cancer patients with cardiotoxicity in comparison to cancer patients without cardiotoxicity, arising a potential diagnostic role in cardiovascular diseases [17].
miRNAs can be potential diagnostic and prognostic tools to improve cancer outcome. Investigations on novel highly informative and specific biological molecules (ECmiRNAs) can lead to early diagnosis of cancer recurrence and improve cancer risk stratification in LCS [18]. Moreover, ECmiRNAs can give feedback to adopt the most appropriate pharmacological treatment and to assess therapy response in order to achieve a precision drug therapy and, therefore, to enhance cancer survival.
2. miRNAs’ Characteristics and Functions
2.1. miRNAs Biogenesis and Gene Regulation
miRNAs are small noncoding RNAs involved in the regulation of gene expression interacting with the target mRNAs [19]. Starting from a longer nucleotide sequence (several hundreds of nucleotides) named primary miRNA (pri-miRNA), subsequent processes lead to a mature single strand miRNA capable of regulating biological functions [19]. In the nucleus, the pri-miRNAs are transcribed mostly by nonprotein coding genes (about 50%) and intragenic regions, mainly introns, while relatively few transcripts result from non-coding exons [20]. The generated pri-miRNA has a characteristic hairpin structure which is recognized and processed by the microprocessor complex [21]. The double-stranded RNA (dsRNA) binding protein DGCR8 (DGR8 microprocessor complex subunit, also named Pasha) and the RNAse III enzyme DROSHA (drosha ribonuclease III), components of the microprocessor complex, cleave the pri-miRNA, generating a 70–120 nucleotides-long precursor miRNA (pre-miRNA) with an overhang of two nucleotides at the 3′ end [22]. Within the pre-miRNA duplex, the two miRNAs strands are differently named as 5p or 3p in accordance with their direction: 5p- and 3p- strands originate from the 5′ and the 3′ ends, respectively [23]. The resulting pre-miRNA is translocated by the nuclear transport receptor protein Exportin 5 (XPO5)/GTP-binding nuclear protein Ran (RanGTP) complex into the cytoplasm, where the final steps of the canonical pathway of miRNA biogenesis can occur [24]. In the cytoplasm, the terminal loop of the pre-miRNA is removed by the ribonuclease III Dicer1 (DICER1) together with the kinase EIF2AK2 (eukaryotic translation initiation factor 2 alpha kinase 2) and the dsRNA-binding proteins TARBP and PRKRA (TAR (HIV-1) RNA binding protein and protein activator of interferon induced protein kinase EIF2AK2, respectively), obtaining a 18–23 nucleotide-long mature miRNA duplex [25]. On the other hand, noncanonical miRNAs biogenesis occurs independently from DROSHA/DGCR8 and DICER1 cleavages [26]. At the end of both canonical and noncanonical miRNAs biogenesis pathways, mature single strand miRNAs regulating the gene expression are generated.
Within the mature miRNA duplex, the two strands are distinguished as a guide strand or passenger strand in accordance with their association, or lack of, with Argonaute RISC complex (AGO) proteins in the assembly of an RNA-induced silencing complex loading complex (RISC) [27]. Generally, the guide strand is loaded into the RISC-loading complex, while the passenger strand is usually degraded [28]. However, the degradation of the passenger strand does not always occur, and both the two mature single strand miRNAs can be loaded into AGO proteins regulating gene expression. For example, miR-574-5p and miR-574-3p act oppositely in gastric cancer: a 5p-strand promotes cancer progression, while a 3p-strand suppresses it and is therefore related to a better cancer prognosis [29].
Once the mature miRNA guide strand is loaded into an AGO protein (AGO1-4 in humans) [30], the gene expression regulation can occur in presence of both the other RISC-loading complex components and the mRNA target [31,32]. miRNAs anneal to a complementary sequence on the 5′ or 3′ UTR of their mRNA targets, named miRNA Response Element (MRE) [33]. The degree of their engagement influences the mRNA fate, which can be degraded or its translation can be repressed [34,35].
Evidences showed miRNAs as pleiotropic and redundant factors in the regulation of biological processes. Indeed, a single mRNA may be targeted by multiple miRNAs, and one miRNA can bind to different mRNAs [36].
2.2. Extracellular miRNAs as Cell Messengers
Several studies observed the presence of several various miRNAs in extracellular fluids such as blood, urine and cell culture medium [15]. The increasing number of studies analysing extracellular circulating miRNAs (ECmiRNAs) suggest the existence of systems preserving miRNAs from their potential extracellular degradation. Plasma ECmiRNAs are remarkably stable when stored at room temperature, subjected to different pH or multiple freeze–thaw cycles, in comparison to synthetic miRNAs and mRNAs [37,38]. The encapsulation and transport of ECmiRNAs by extracellular vesicles was first hypothesized as a protecting mechanism [39,40]. Moreover, two different contemporary studies observed ECmiRNAs from plasma or cell culture medium mainly associated to AGO proteins (90%) [41,42]. Currently, circulating ECmiRNAs can be found within extracellular vesicles (EVs) or bound to proteins. Exosomes, microvesicles and apoptotic bodies are the main EVs enwrapping ECmiRNAs [43,44]. Differences in dimension, composition and biosynthesis distinguish the various EVs [45]: (i) exosomes are 30–100 nm in diameter vesicles that originate by internal budding of early endosomes [46]; (ii) microvesicles are both heterogeneous in size (i.e., from 100 to 1000 nm in diameter) and composition, since they originate by outward budding or fission of plasma membrane [47]; (iii) apoptotic bodies are larger vesicles of 1–5 µm in diameter released by dying cells [48]. Among the non-packaged ECmiRNAs, AGO2 and HDL protect them from the extracellular degradation [42,49]. Circulating miRNAs identified in cerebrospinal fluid were mostly immunoprecipitated by anti-AGO2 antibodies, suggesting their predominant association with AGO proteins [50]. A potential diagnostic role of this complex was hypothesized due to the variation of AGO2/ECmiRNAs concentration under pathological conditions, such as colorectal cancer [50].
The stability of ECmiRNAs in the extracellular fluids proposes the existence of their key roles in gene expression regulation among cells, rather than resulting as byproducts of cellular activities [19]. Furthermore, the release of ECmiRNAs is a controlled process. Ceramide is a sphingolipid on plasma membrane involved in the secretion of exosomal miRNAs. Inhibition of the sphingomyelin phosphodiesterase 2 (SMPD2) enzyme in HEK293 cells caused reduced ceramide biosynthesis, resulting in decreased exosomal miRNAs [51]. Recently, the GGAG and GGUC conserved sequences (EXO and extra-seed EXO [hEXO] sequence, respectively) at the 3′ end of exosomal miRNAs have been identified and related to the ECmiRNAs secretion mechanism [52,53]. Evidence showed the interaction among the recognized miRNAs motif and specific cellular proteins. The sumoylated form of HNRNPA2B1 (heterogeneous nuclear ribonucleoprotein A2/B1) recognized the GGAG motif on miR-198 but not the mutated sequence in T cells [54], while the GGUC sequence was bound by the SYNCRIP protein (synaptotagmin-binding cytoplasmic RNA-interacting protein) in hepatocytes [55].
As a consequence of the intracellular concentrations of both miRNAs and their targets, a passive cellular mechanism for ECmiRNAs release has been hypothesized [56]. According to this thesis, miRNAs containing vesicles are secreted when miRNAs concentration raises at a higher level than its targeted mRNA, in order to maintain a stable intracellular miRNA:mRNA ratio [57].
3. miRNAs in Cancer
Early and specific predictors of tumor development and progression are key elements to achieve constant monitoring in LCS patients. miRNAs have been identified as good potential screening markers for different cancer types and, therefore, for related drug treatments [58]. Their eligibility also resides in noninvasive and nonharmful tests (i.e., blood draws), that can be conceivably repeated limitlessly.
3.1. Aspects of Intracellular miRNAs
miRNAs regulate at least 30% of all human genes influencing most of the biological processes, including cell fate differentiation, proliferation and host-viral infection [59]. Alteration in miRNAs biogenesis, as well as in miRNAs expression levels and functions, have been related to several human cancers [60]. Decreased human HCT-116 cell (human colorectal carcinoma cells) proliferation was observed as a consequence of the knockout of DROSHA and DICER1. Western blot and sequencing analyses showed miRNAs expression reductions of 96.5 and 96% in the absence of DROSHA and DICER1, respectively [61].
The quick malignant cellular proliferation and expansion requires continuous availability of intermediates and metabolites. Cancer cells are capable of switching their miRNA expression in order to support their metabolic activities, forcing tumor cells to perform anaerobic glycolysis instead of aerobic respiration (Warburg effect) [62]. Different glycolytic enzymes are targeted by the miR-200 family: miR-122; miR-17/92 cluster; miR-15a/16-1; miR-29; miR-326 and miR-133 to support cancer cell metabolic needs, proliferation and metastasis [63]. The PGI/AMF is one of the first glycolytic enzymes, which also induces the expression of ZEB1/ZEB2 (zinc finger E-Box-binding homeobox 1/zinc finger E-Box-binding homeobox 2), thus enhancing the epithelial-to-mesenchymal transition (EMT). In MDA-MB-231 breast cancer cells, the repression of miR-200 family (miR-200a, miR-200b, and miR-200c), as a consequence of PGI/AMF (phosphoglucose isomerase/autocrine motility factor) overexpression, resulted in increased metastasis [64].
According to their role within cancer development and progression, miRNAs can be recognized as tumor-promoting miRNAs (oncomiRs), metastasis-promoting miRNAs (metastamiRs) and tumor-suppressor miRNAs [65]. The tumorigenic role of miRNAs is also influenced by the cellular context, since they can act either as tumor suppressors or as oncomiRs [66].
The PIK3/AKT/MTOR (phosphatidylinositol-4,5-bisphosphate 3-kinase/AKT serine-threonine kinase/mechanistic target of rapamycin kinase) is one of the most activated cell signaling routes in cancer [67]. Loss of function mutations of phosphatase and tensin homolog (PTEN), a negative regulator of PIK3/AKT/MTOR [68], increase PIK3/AKT/MTOR pathway activation and are associated with cancer development [69]. Gene expression analyses by RNA-seq and miRNAs microarray in biopsy or surgery tissue from 217 colorectal carcinoma patients observed the association between miRNAs and genes involving a PIK3/AKT signaling pathway, such as miR-590-5p, miR-106b and miR-93 acting as PTEN suppressors [70]. In SUM149 breast cancer cells, miR-181c inhibited the PTEN protein expression, targeting its 3′UTR mRNA, and promoted breast cancer proliferation [71]. Upregulation of miR-181a (a member of the miR-181 cluster) in Jurkat T-ALL cells (T-acute lymphoblastic leukaemia) reduced EGR1 (early growth response one) level, inducing G1/S cell-cycle progression and cell proliferation [72].
The same miRNAs may have different targets within diverse cancer types. MiR-105 was upregulated in triple-negative breast cancer tissue promoting chemoresistance, stemness and metastasis, acting on a WNT/β-catenin pathway [73]. In hepatocellular carcinoma, miR-105 downregulation activated the PIK3/AKT signaling, enhancing cancer proliferation both in vitro and in vivo [74].
Specific groups of miRNAs (clusters), and their relative expression levels may disclose a set of information with a diagnostic and prognostic role [12,16]. The first evidence of miRNAs’ involvement in human cancer dates back to 2002, when miR-15a and miR-16-1 dysregulation was associated to B-cell chronic lymphocytic leukemia (B-CLL) [75]. The 13q14 chromosome deletion, present in over 50% of B-CLL cases, induces the loss of the tumor suppressor miR-15a/miR-16-1 cluster (miR-15/16). Downregulation of these miRNAs promotes overexpression of several oncogenes including BCL2 (BCL2 apoptosis regulator), inhibiting apoptosis of tumor cells and enhancing cell proliferation [76,77,78]. Moreover, deletions or loss of functions (mutations) of the miR-15/16 were reported in a variety of tumors other than CLL, such as melanoma, colorectal and prostate cancer [79].
3.2. Extracellular miRNAs in Cancer
Recent studies have highlighted the functional role of ECmiRNAs, that after secretion are delivered towards target cells where they can induce regulatory effects. Therefore, since their role in cell-to-cell communication, ECmiRNAs have been identified as biomarkers for several human pathologies, including cancer.
The involvement of ECmiRNAs in tumorigenesis was hypothesized as a consequence of the higher number of released exosomes by cancer cells compared to normal controls [80]. In 24 h, the B42 breast cancer cell line released approximately 53 × 108 exosomes per million cells, whereas nonpathological human mammary epithelial cells (HMEC B42) released 5 × 108 exosomes per million cells [81].
Exosomes derived from EMT-transformed HCT-116 cells and containing miR-128-3p downregulated FOXO4 (forkhead box O4) and activated TGF-β/SMAD (transforming growth factor- β/SMAD family) and JAK (Janus kinase)/STAT3 signaling pathways in normal HCT-116 cells, inducing the EMT process [82]. The level of six ECmiRNAs (miR-19b-3p; miR-21-5p; miR-221-3p; miR-409-3p; miR-425-5p and miR-584-5p) were increased in the plasma of lung adenocarcinoma (LA) patients in comparison to healthy subjects, suggesting their potential role for LA diagnosis [83].
Colorectal cancer growth was also related to the regulation of the immune system through miR-222-EVs secretion. Decreased release of miR-222 microvesicles from implanted SW480 human colon cancer cells in mice resulted in reduced colorectal cancer growth and higher CD3+ cells infiltration of cancer tissue, suggesting a reduced immune escape [84].
Further studies investigated the messenger role of ECmiRNAs in cell-to-cell communication, highlighting their relevance within the tumor microenvironment [16,85]. Exosomal miR-21, secreted by human bronchial epithelial cancer cells (HBE), increased VEGF (vascular endothelial growth factor) in human umbilical vein endothelial cells (HUVEC), promoting angiogenesis [86]. Exosomes containing miR-105 can be released by metastatic breast cancer cells to destroy the vascular endothelium barrier targeting the tight junction protein 1 (TJP1) in endothelial cells (recipient cells) and promoting cancer metastasis [87].
Cancer therapy induces changes in the ECmiRNAs pattern of tumor cells and can become a readout of treatment efficacy. Analysis of blood samples from 68 metastatic colorectal cancer patients showed a decreased miR-126 level in the combined treatment of chemotherapy and bevacizumab [88]. Higher miR-128b level in peripheral blood was related to good prednisolone response and prognosis in children with ALL [89].
Summary of the described miRNAs involved in cancer metabolic pathways and tumorigenic processes are listed in Table 1 and Table 2.

Table 1.
Examples of representative miRNAs involved in cancer cell lines development, progression and therapy response.
Table 2.
Examples of representative miRNAs from samples of cancer patients involved in tumor development, progression and therapy response.
4. Cancer Survival
When it was defined in 1985, the term “cancer survivor” referred to a cancer patient who went from a phase of “acute survivorship” (i.e., cancer diagnosis and primary treatments) to “permanent survivorship” (i.e., cancer remission or cancer as a chronic disease), experiencing a phase of “extended survivorship” (i.e., physical and psychological consequences following cancer treatment) [90]. Then, the American National Coalition of Cancer Survivorship (NCCS) identified the LCS as a person other than a patient, recognizing his/her needs (e.g., psychological, legal, medical) from the moment of the cancer diagnosis through the balance of life [91]. Moreover, the National Cancer Survivorship Initiative in the UK includes “patients undergoing primary treatments, in remission, with active or advanced disease and cured” in the LCS concept [92]. In Italy, survivors are generally defined as patients free of cancer disease and therapy for at least 5 years [93]. Within the LCS group, some researchers describe as “cured” survivors, the cancer survivors with a death rate comparable to general populations [94,95]. It should be taken into account that some patients reject the term “survivor” judging it as offensive [96]. Therefore, actually LCS do not correspond to a unique definition [97].
Overcoming the definition issue, the percentage of LCS among cancer patients is increasing due to early diagnosis and new advanced therapies. According to recent research, more than 68% of approximately 17 million of American LCS had a cancer history of 5 years or more, and 18% (i.e., more than 3 million) were 20-year survivors in 2019 [98]. The same research has foreseen an increase in LCS numbers to more than 22.1 million by 2030 [98].
Several factors affect the cancer survivorship, such as type and stage of cancer at diagnosis, cancer therapy and lifestyle. It is also known that cancer treatments, such as chemotherapy, lead to several side effects due to their nonspecific action targeting both cancer and normal cells [99]. Other than an increased risk of mortality, cancer therapy may worsen the quality of life [100]. For example, higher incidences of obesity, diabetes or fractures associated with osteoporosis were observed in prostate cancer survivors treated with androgen deprivation therapy (ADT) [101].
Hence, LCS life expectancy is a multifactorial condition in which patient care and lifestyle contribute to survival regardless of the tumor characteristics (Figure 1) [102,103].
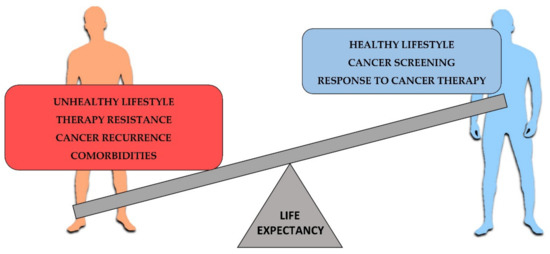
Figure 1.
Factors affecting the cancer life expectancy. On the left, most relevant negative factors for cancer survival. On the right, positive factors contributing to increase the cancer life expectancy.
5. miRNAs as Biomarkers in Cancer Survivors
Other than advances in pharmacological treatments, an earlier cancer diagnosis, accompanied by the most appropriate therapy, can improve the life expectancy of cancer patients [104]. Several studies showed the high sensitivity and specificity of miRNAs in cancer diagnosis [105], highlighting their potential in reducing tumor mortality as a consequence of earlier cancer detection. Recent studies evaluated the ECmiRNAs expression and correlated it to the cancer prognosis, focusing on long-term survival.
5.1. miRNAs for Cancer Follow-Up and Monitoring of the Therapy
Since ECmiRNAs profiles have been recognized as representative of miRNAs patterns expressed by different tumors and are capable of distinguishing their different development stages, they may act as a useful tool to predict cancer relapse or progression. Among ECmiRNAs profiles, the analysis of more than 2000 serum samples of patients with or without lung cancer, miR-1268b and miR-6075 showed 99% of both sensitivity and specificity in the detection of lung cancer [106]. Distinct ECmiRNAs patterns were detected among hepatocellular carcinoma patients and noncancerous subjects during disease progression [107].
For example, in chronic hepatitis B patients, upregulated miR-122-5p, miR-125b-5p, miR-885-5p, miR-100-5p and miR-148a-3p were found, while miR-34a-5p was identified as a biomarker for cirrhosis. Finally, downregulation of miR-424-5p and miR-101-3p and upregulation of miR-128, miR-139-5p, miR-382-5p and miR-410 were associated with larger HCC (hepatocellular carcinoma) tumor size and invasion, as well as a worse survival outcome [107].
Evaluation of ECmiRNAs patterns in breast cancer patients with more than 15 years of follow-up displayed a cancer survival potentially influenced by the expression of the five most relevant miRNAs (miR-29c, miR-143-3p, miR-187-3p, miR-205-5p and miR-210). In particular, decreased levels of both miR-210 and miR-29c were associated with a 10-year life expectancy, suggesting a better cancer prognosis [108].
Specific miRNAs patterns have been related to therapy response, thus contributing to assess its efficacy and therefore, cancer prognosis. The signature of four serum ECmiRNAs (miR-940, miR-451a, miR-16-5p and miR-17-3p) was related to the trastuzumab treatment response in 386 ERBB2+ (erb-b2 receptor tyrosine kinase 2-positive)-metastatic breast cancer patients, compared to 179 chemotherapy-treated patients. The increased expression of these four miRNAs was associated with trastuzumab resistance [109]. Analysis of miRNAs in the serum of 97 castration-resistant prostate cancer showed high levels of miR-200b, miR-429, miR-200a, miR-21, miR-200c, miR-375, miR-132, miR-20a and low levels of miR-590-5p in patients with shorter survival. Moreover, after cancer therapy with docetaxel, decreased or unchanged levels of miR-20a, miR-222, miR-20b, miR-132 or miR-25 were associated with poor prognosis. Therefore, these miRNAs patterns were suggested as potential biomarkers for docetaxel resistance and poor prognosis in prostate cancer patients [110].
Diet contributes to cancer survival and influences the ECmiRNAs pattern. In a very recent study, 42 EVs-miRNAs (among the 798 studied) were differently expressed in the plasma of overweight breast cancer survivors treated with the Mediterranean diet for eight weeks: 36 miRNAs were upregulated and six miRNAs were downregulated. The identified miRNAs were associated with different cellular pathways involved in breast cancer progression (e.g., miR-122-5p and let-7a-5p, engaged in tumor metastasis), energy and glucose metabolisms (e.g., miR-329-3p and miR-216-5p, altered in obesity), thus suggesting a positive influence of the diet [111].
ECmiRNAs have been also associated with cancer relapse prediction. qRT-PCR analysis showed a higher expression of miR-194 and miR-146-3p in the serum of patients with prostate cancer recurrence compared to nonrecurrent patients, following radical prostatectomy [112]. In another study, increased circulating miR-17, miR-20a, miR-20b and miR-106a (all members of the oncogenic cluster of the miR-17 family) in blood samples predicted a high risk of prostate cancer relapse [113]. In biliary tract cancer, the relapse after radical surgery was disclosed by six specific ECmiRNAs (miR-1225-3p, miR-1234-3p, miR-1260b and miR-1470, miR-6834-3p and miR-6875-5p). Upregulation of miR-1225-3p, miR-1234-3p, miR-1260b and miR-1470 and downregulation of miR-6834-3p and miR-6875-5p were observed in the serum samples of patients with cancer relapse. The best prognostic information with 84.6% of sensitivity, 100% of specificity and 90.0% of accuracy, was obtained using a set of three randomly selected miRNAs among the six analysed [114].
5.2. miRNAs to Monitor Cancer Therapy Side Effects
Other than ECmiRNAs patterns related to cancer follow-up, circulating miRNAs have been also associated with cancer therapy side effects, such as cardiotoxicity, that influence patients’ life expectancy.
At least 50% of cancer chemotherapy drugs, including doxorubicin, anthracycline and cisplatin, are known to induce oxidative stress [115]. Oxidative stress condition changes EVs and ECmiRNAs release to mediate cell–cell communication in order to counteract or alternatively boost the inflammation [116,117]. MiR-1, miR-133b, miR-146a, miR-208a, miR-208b and miR-423-5p (cardiac functions related miRNAs) were observed in plasma of 56 doxorubicin-treated breast cancer patients after each cycle of treatment. Among them, miR-1 was recognized as the best biomarker for doxorubicin-induced cardiotoxicity showing its significant increased expression in comparison to the other miRNAs, and driving a better evaluation of cardiac injuries rather than the cardiac troponin I (cTnI), in patients with cardiotoxicity [118,119]. Upregulation of miR-126-3p, miR-199a-3p, miR-423-5p and miR-34a-5p (markers of congestive heart failure [CHF]) was detected in the plasma of primary breast cancer patients treated with anthracycline-based chemotherapy that developed CHF, suggesting an impairment of heart functions as consequence of cancer therapy [120].
ECmiRNAs identified as biomarkers for cancer follow-up are listed in Table 3.
Table 3.
Examples of miRNAs as cancer follow-up biomarkers.
6. Conclusions
Changes in miRNA expression levels alter biological processes, such as cellular proliferation and metabolism. Evaluation of ECmiRNAs in biological fluids, such as blood or urine, could represent a potential routine test, based on miRNAs quick analysis, noninvasive and nonhazardous characteristics for the patients. Therefore, ECmiRNAs analysis appears conceivably endlessly repeatable, with the possibility of multiple samplings over time that are important especially in the follow-up [121]. Furthermore, since specific circulating miRNAs should reflect the miRNAs profiles within the parental tumor [53], they might provide information on the tumor stages and its modifications more rapidly than other techniques. Currently, some cancers, such as lung cancer, have low life expectancies due to a lack of distinctive symptoms that causes a late diagnosis [98]. A specific ECmiRNAs profile could be informative for cancer diagnosis and follow-up, but could also suggest the appropriate treatment to block tumor growth and to monitor therapy side effects. Recent studies evaluated miRNAs themselves as a potential cancer therapy. For example, TG221 transgenic mice treated with miR-199a-3p mimic displayed a reduction in liver cancer size and a fewer number of nodules compared to control animals [122].
It should be taken into account that ECmiRNAs expression changes as a consequence of personal characteristics, such as lifestyle (i.e., diet and physical activity) [123,124]. Limitations of ECmiRNAs analyses may be related to availability of clinical samples and amount of miRNAs content, RNA extraction methods, miRNA database errors and unstandardized statistical analyses [125,126].
Identification of biomarkers in cancer survivors is a very innovative research topic and ECmiRNAs are gaining an emerging role within it. The identification of ECmiRNAs patterns, related to specific cancer stages and progression phases, could allow us to prevent cancer relapse and to monitor therapy effects.
Knowledge of cancer characteristics, as described by ECmiRNAs, may orient toward the best individual treatment, fitting within the idea of a personalized precision medicine.
Author Contributions
I.C., L.M.N.: idealization, intellectual input; I.C., C.S. and C.B.: literature search and writing initial version of the manuscript; G.V. and E.C.: manuscript editing; L.M.N.: manuscript editing and supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by “Local Research grants” from University of Ferrara, Italy (FIR and FAR to L.M.N. and FAR to C.S.) and by Associazione Onconauti (L.M.N.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to Antonio Frassoldati (Head Oncology Unit, University Hospital St. Anna, Ferrara, Italy), Antonio Maestri (Head Medical Oncology Department, Santa Maria della Scaletta Hospital, Imola, Italy) and Angela Passaro (Head of Metabolic Center, Internal Medicine Unit, Azienda Ospedaliero, Universitaria S. Anna, Ferrara, Italy) for helpful discussion.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Hulvat, M.C. Cancer Incidence and Trends. Surg. Clin. N. Am. 2020, 100, 469–481. [Google Scholar] [CrossRef]
- Carioli, G.; Bertuccio, P.; Boffetta, P.; Levi, F.; La Vecchia, C.; Negri, E.; Malvezzi, M. European cancer mortality predictions for the year 2020 with a focus on prostate cancer. Ann. Oncol. 2020, 31, 650–658. [Google Scholar] [CrossRef] [PubMed]
- IARC (International Agency for Research on Cancer). GLOBOCAN 2020: New Global Cancer Data. Available online: https://gco.iarc.fr/ (accessed on 3 February 2021).
- Koene, R.J.; Prizment, A.E.; Blaes, A.; Konety, S.H. Shared Risk Factors in Cardiovascular Disease and Cancer. Circulation 2016, 133, 1104–1114. [Google Scholar] [CrossRef] [Green Version]
- Zamorano, J.L.; Lancellotti, P.; Munoz, D.; Aboyans, V.; Asteggiano, R.; Galderisi, M.; Habib, G.; Lenihan, D.J.; Lip, G.Y.H.; Lyon, A.R.; et al. 2016 ESC Position Paper on cancer treatments and cardiovascular toxicity developed under the auspices of the ESC Committee for Practice Guidelines The Task Force for cancer treatments and cardiovascular toxicity of the European Society of Cardiology (ESC). Eur. J. Heart Fail. 2017, 19, 9–42. [Google Scholar] [CrossRef] [PubMed]
- Strongman, H.; Gadd, S.; Matthews, A.; Mansfield, K.E.; Stanway, S.; Lyon, A.R.; dos-Santos-Silva, I.; Smeeth, L.; Bhaskaran, K. Medium and long-term risks of specific cardiovascular diseases in survivors of 20 adult cancers: A population-based cohort study using multiple linked UK electronic health records databases. Lancet 2019, 394, 1041–1054. [Google Scholar] [CrossRef] [Green Version]
- Lan, H.Y.; Lu, H.Q.; Wang, X.; Jin, H.C. MicroRNAs as Potential Biomarkers in Cancer: Opportunities and Challenges. Biomed Res. Int. 2015, 2015, 125094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaskaran, M.; Mohan, M. MicroRNAs: History, Biogenesis, and Their Evolving Role in Animal Development and Disease. Vet. Pathol. 2014, 51, 759–774. [Google Scholar] [CrossRef] [Green Version]
- Duarte, F.V.; Palmeira, C.M.; Rolo, A.P. The Emerging Role of MitomiRs in the Pathophysiology of Human Disease. Adv. Exp. Med. Biol. 2015, 888, 123–154. [Google Scholar] [CrossRef] [PubMed]
- Eulalio, A.; Huntzinger, E.; Izaurralde, E. Getting to the root of miRNA-Mediated gene silencing. Cell 2008, 132, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Paul, P.; Chakraborty, A.; Sarkar, D.; Langthasa, M.; Rahman, M.; Singha, R.K.S.; Malakar, A.K.; Chakraborty, S. Interplay between miRNAs and human diseases. J. Cell. Physiol. 2018, 233, 2007–2018. [Google Scholar] [CrossRef]
- Van den Brande, S.; Gijbels, M.; Wynant, N.; Santos, D.; Mingels, L.; Gansemans, Y.; Van Nieuwerburgh, F.; Vanden Broeck, J. The presence of extracellular microRNAs in the media of cultured Drosophila cells. Sci. Rep. 2018, 8, 17312. [Google Scholar] [CrossRef]
- Weber, J.A.; Baxter, D.H.; Zhang, S.L.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Sohel, M.H. Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges. Achiev. Life Sci. 2016, 10, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Conti, I.; Varano, G.; Simioni, C.; Laface, I.; Milani, D.; Rimondi, E.; Neri, L.M. miRNAs as Influencers of Cell-Cell Communication in Tumor Microenvironment. Cells 2020, 9, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, X.T.; Chang, F.Y.; Wang, Z.F.; Jiang, W.Y. Correlation of circulating pro-angiogenic miRNAs with cardiotoxicity induced by epirubicin/cyclophosphamide followed by docetaxel in patients with breast cancer. Cancer Biomark 2018, 23, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global Cancer Statistics. Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Saini, H.K.; Griffiths-Jones, S.; Enright, A.J. Genomic analysis of human microRNA transcripts. Proc. Natl. Acad. Sci. USA 2007, 104, 17719–17724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denli, A.M.; Tops, B.B.J.; Plasterk, R.H.A.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yu, J.F.; Yu, H.; Zhao, Y.; Chen, S.J.; Xu, C.Q.; Chen, F. Evolutionary and Expression Analysis of miR-#-5p and miR-#-3p at the miRNAs/isomiRs Levels. Biomed Res. Int. 2015, 2015, 168358. [Google Scholar] [CrossRef] [PubMed]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.; Yoneda, Y.; Tsukihara, T. A High-Resolution Structure of the Pre-microRNA Nuclear Export Machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef]
- Zhang, H.D.; Kolb, F.A.; Jaskiewicz, L.; Westhof, E.; Filipowicz, W. Single processing center models for human dicer and bacterial RNase III. Cell 2004, 118, 57–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pong, S.K.; Gullerova, M. Noncanonical functions of microRNA pathway enzymes—Drosha, DGCR8, Dicer and Ago proteins. FEBS Lett. 2018, 592, 2973–2986. [Google Scholar] [CrossRef] [Green Version]
- Kawamata, T.; Tomari, Y. Making RISC. Trends Biochem. Sci. 2010, 35, 368–376. [Google Scholar] [CrossRef]
- Noland, C.L.; Doudna, J.A. Multiple sensors ensure guide strand selection in human RNAi pathways. RNA 2013, 19, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Y.; Pi, J.N.; Zou, D.L.; Wang, X.S.; Xu, J.Y.; Yu, S.; Zhang, T.; Li, F.; Zhang, X.X.; Zhao, H.L.; et al. microRNA arm-imbalance in part from complementary targets mediated decay promotes gastric cancer progression. Nat. Commun. 2019, 10, 4397. [Google Scholar] [CrossRef] [Green Version]
- Yoda, M.; Kawamata, T.; Paroo, Z.; Ye, X.C.; Iwasaki, S.; Liu, Q.H.; Tomari, Y. ATP-dependent human RISC assembly pathways. Nat. Struct. Mol. Biol. 2010, 17, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, D.S.; Hutvagner, G.; Du, T.; Xu, Z.S.; Aronin, N.; Zamore, P.D. Asymmetry in the assembly of the RNAi enzyme complex. Cell 2003, 115, 199–208. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Jing, Y.; Zhang, H.Y.; Niu, Y.H.; Liu, C.; Wang, J.; Zen, K.; Zhang, C.Y.; Li, D.H. Comprehensive Evolutionary Analysis of the Major RNA-Induced Silencing Complex Members. Sci. Rep. 2018, 8, 14189. [Google Scholar] [CrossRef]
- Jo, M.H.; Shin, S.; Jung, S.R.; Kim, E.; Song, J.J.; Hohng, S. Human Argonaute 2 Has Diverse Reaction Pathways on Target RNAs. Mol. Cell 2015, 59, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Christie, M.; Boland, A.; Huntzinger, E.; Weichenrieder, O.; Izaurralde, E. Structure of the PAN3 Pseudokinase Reveals the Basis for Interactions with the PAN2 Deadenylase and the GW182 Proteins. Mol. Cell 2013, 51, 360–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiriakidou, M.; Tan, G.S.; Lamprinaki, S.; De Planell-Saguer, M.; Nelson, P.T.; Mourelatos, Z. An mRNA m(7)G cap binding-like motif within human Ago2 represses translation. Cell 2007, 129, 1141–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebert, M.S.; Sharp, P.A. Roles for MicroRNAs in Conferring Robustness to Biological Processes. Cell 2012, 149, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [Green Version]
- Jung, M.; Schaefer, A.; Steiner, I.; Kempkensteffen, C.; Stephan, C.; Erbersdobler, A.; Jung, K. Robust MicroRNA Stability in Degraded RNA Preparations from Human Tissue and Cell Samples. Clin. Chem. 2010, 56, 998–1006. [Google Scholar] [CrossRef] [Green Version]
- Hunter, M.P.; Ismail, N.; Zhang, X.L.; Aguda, B.D.; Lee, E.J.; Yu, L.B.; Xiao, T.; Schafer, J.; Lee, M.L.T.; Schmittgen, T.D.; et al. Detection of microRNA Expression in Human Peripheral Blood Microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenen-Stass, A.M.L.; Pauwels, M.J.; Hanson, B.; Perez, C.M.; Conceicao, M.; Wood, M.J.A.; Mager, I.; Roberts, T.C. Extracellular microRNAs exhibit sequence-dependent stability and cellular release kinetics. RNA Biol. 2019, 16, 696–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef]
- Crescitelli, R.; Lasser, C.; Szabo, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzas, E.I.; Lotvall, J. Distinct RNA profiles in subpopulations of extracellular vesicles: Apoptotic bodies, microvesicles and exosomes. J. Extracell. Vesicles 2013, 2, 20677. [Google Scholar] [CrossRef]
- Than, U.T.T.; Guanzon, D.; Broadbent, J.A.; Leavesley, D.I.; Salomon, C.; Parker, T.J. Differential Expression of Keratinocyte-Derived Extracellular Vesicle Mirnas Discriminate Exosomes From Apoptotic Bodies and Microvesicles. Front. Endocrinol. 2018, 9, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Yanez-Mo, M.; Siljander, P.R.M.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thery, C.; Ostrowski, M.; Segura, E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009, 9, 581–593. [Google Scholar] [CrossRef]
- Turiak, L.; Misjak, P.; Szabo, T.G.; Aradi, B.; Paloczi, K.; Ozohanics, O.; Drahos, L.; Kittel, A.; Falus, A.; Buzas, E.I.; et al. Proteomic characterization of thymocyte-derived microvesicles and apoptotic bodies in BALB/c mice. J. Proteomics 2011, 74, 2025–2033. [Google Scholar] [CrossRef]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Fuji, T.; Umeda, Y.; Nyuya, L.A.; Taniguchi, L.F.; Kawai, T.; Yasui, K.; Toshima, T.; Yoshida, K.; Fujiwara, T.; Goel, A.; et al. Detection of circulating microRNAs with Ago2 complexes to monitor the tumor dynamics of colorectal cancer patients during chemotherapy. Int. J. Cancer 2019, 144, 2169–2180. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory Mechanisms and Intercellular Transfer of MicroRNAs in Living Cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, L.; Giurato, G.; Cicchini, C.; Montaldo, C.; Mancone, C.; Tarallo, R.; Battistelli, C.; Alonzi, T.; Weisz, A.; Tripodi, M. The RNA-Binding Protein SYNCRIP Is a Component of the Hepatocyte Exosomal Machinery Controlling MicroRNA Sorting. Cell Rep. 2016, 17, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Bhome, R.; Del Vecchio, F.; Lee, G.H.; Bullock, M.D.; Primrose, J.N.; Sayan, A.E.; Mirnezami, A.H. Exosomal microRNAs (exomiRs): Small molecules with a big role in cancer. Cancer Lett. 2018, 420, 228–235. [Google Scholar] [CrossRef]
- Villarroya-Beltri, C.; Gutierrez-Vazquez, C.; Sanchez-Cabo, F.; Perez-Hernandez, D.; Vazquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sanchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [Green Version]
- Hobor, F.; Dallmann, A.; Ball, N.J.; Cicchini, C.; Battistelli, C.; Ogrodowicz, R.W.; Christodoulou, E.; Martin, S.R.; Castello, A.; Tripodi, M.; et al. A cryptic RNA-binding domain mediates Syncrip recognition and exosomal partitioning of miRNA targets. Nat. Commun. 2018, 9, 831. [Google Scholar] [CrossRef]
- Bell, E.; Taylor, M.A. Functional Roles for Exosomal MicroRNAs in the Tumour Microenvironment. Comput Struct. Biotec. 2017, 15, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Squadrito, M.L.; Baer, C.; Burdet, F.; Maderna, C.; Gilfillan, G.D.; Lyle, R.; Ibberson, M.; De Palma, M. Endogenous RNAs Modulate MicroRNA Sorting to Exosomes and Transfer to Acceptor Cells. Cell Rep. 2014, 8, 1432–1446. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [Green Version]
- Iorio, M.V.; Croce, C.M. microRNA involvement in human cancer. Carcinogenesis 2012, 33, 1126–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwivedi, S.; Purohit, P.; Sharma, P. MicroRNAs and Diseases: Promising Biomarkers for Diagnosis and Therapeutics. Indian J. Clin. Biochem. 2019, 34, 243–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.K.; Kim, B.; Kim, V.N. Re-evaluation of the roles of DROSHA, Exportin 5, and DICER in microRNA biogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E1881–E1889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azizi, M.I.H.N.; Othman, I.; Naidu, R. The Role of MicroRNAs in Lung Cancer Metabolism. Cancers 2021, 13, 1716. [Google Scholar] [CrossRef]
- Mirzaei, H.; Hamblin, M.R. Regulation of Glycolysis by Non-coding RNAs in Cancer: Switching on the Warburg Effect. Mol. Ther. Oncolytics 2020, 19, 218–239. [Google Scholar] [CrossRef]
- Ahmad, A.; Aboukameel, A.; Kong, D.J.; Wang, Z.W.; Sethi, S.; Chen, W.; Sarkar, F.H.; Raz, A. Phosphoglucose Isomerase/Autocrine Motility Factor Mediates Epithelial-Mesenchymal Transition Regulated by miR-200 in Breast Cancer Cells. Cancer Res. 2011, 71, 3400–3409. [Google Scholar] [CrossRef] [Green Version]
- Gambari, R.; Brognara, E.; Spandidos, D.A.; Fabbri, E. Targeting oncomiRNAs and mimicking tumor suppressor miRNAs: New trends in the development of miRNA therapeutic strategies in oncology. Int. J. Oncol. 2016, 49, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, Z.Y.; Chen, F.Y.; Hu, T.; Peng, W.; Gu, Q.; Sun, Y.M. The Diverse Oncogenic and Tumor Suppressor Roles of microRNA-105 in Cancer. Front. Oncol. 2019, 9, 518. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, R.A. Oncogenes and the Molecular-Biology of Cancer. J. Cell Biol. 1983, 97, 1661–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simioni, C.; Martelli, A.M.; Zauli, G.; Melloni, E.; Neri, L.M. Targeting mTOR in Acute Lymphoblastic Leukemia. Cells 2019, 8, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.S.; Salmena, L.; Pandolfi, P.P. The functions and regulation of the PTEN tumour suppressor. Nat. Rev. Mol. Cell Bio. 2012, 13, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.L.; Mullany, L.E.; Sakoda, L.C.; Wolff, R.K.; Stevens, J.R.; Samowitz, W.S.; Herrick, J.S. The PI3K/AKT signaling pathway: Associations of miRNAs with dysregulated gene expression in colorectal cancer. Mol. Carcinogen. 2018, 57, 243–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.L.; Zhang, J.H. miR-181c promotes proliferation via suppressing PTEN expression in inflammatory breast cancer. Int. J. Oncol. 2015, 46, 2011–2020. [Google Scholar] [CrossRef] [Green Version]
- Verduci, L.; Azzalin, G.; Gioiosa, S.; Carissimi, C.; Laudadio, I.; Fulci, V.; Macino, G. microRNA-181a enhances cell proliferation in acute lymphoblastic leukemia by targeting EGR1. Leuk. Res. 2015, 39, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.Y.; Liang, J.L.; Kuo, Y.L.; Lee, H.H.; Calkins, M.J.; Chang, H.T.; Lin, F.C.; Chen, Y.C.; Hsu, T.I.; Hsiao, M.; et al. miR-105/93-3p promotes chemoresistance and circulating miR-105/93-3p acts as a diagnostic biomarker for triple negative breast cancer. Breast Cancer Res. 2017, 19, 1–14. [Google Scholar] [CrossRef]
- Shen, G.; Rong, X.X.; Zhao, J.B.; Yang, X.W.; Li, H.B.; Jiang, H.; Zhou, Q.; Ji, T.X.; Huang, S.C.; Zhang, J.; et al. MicroRNA-105 suppresses cell proliferation and inhibits PI3K/AKT signaling in human hepatocellular carcinoma. Carcinogenesis 2014, 35, 2748–2755. [Google Scholar] [CrossRef] [Green Version]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [Green Version]
- Aqeilan, R.I.; Calin, G.A.; Croce, C.M. miR-15a and miR-16-1 in cancer: Discovery, function and future perspectives. Cell Death Differ. 2010, 17, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pekarsky, Y.; Croce, C.M. Role of miR-15/16 in CLL. Cell Death Differ. 2015, 22, 6–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calin, G.A.; Cimmino, A.; Fabbri, M.; Ferracin, M.; Wojcik, S.E.; Shimizu, M.; Taccioli, C.; Zanesi, N.; Garzon, R.; Aqeilan, R.I.; et al. MiR-15a and miR-16-1 cluster functions in human leukemia. Proc. Natl. Acad. Sci. USA 2008, 105, 5166–5171. [Google Scholar] [CrossRef] [Green Version]
- Pekarsky, Y.; Balatti, V.; Croce, C.M. BCL2 and miR-15/16: From gene discovery to treatment. Cell Death Differ. 2018, 25, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Ingenito, F.; Roscigno, G.; Affinito, A.; Nuzzo, S.; Scognamiglio, I.; Quintavalle, C.; Condorelli, G. The Role of Exo-miRNAs in Cancer: A Focus on Therapeutic and Diagnostic Applications. Int. J. Mol. Sci. 2019, 20, 4687. [Google Scholar] [CrossRef] [Green Version]
- Riches, A.; Campbell, E.; Borger, E.; Powis, S. Regulation of exosome release from mammary epithelial and breast cancer cells—A new regulatory pathway. Eur. J. Cancer 2014, 50, 1025–1034. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, X.; Shi, D.D.; Xiang, Z.X.; Wang, S.Y.; Yang, C.G.; Liu, Q.; Huang, S.H.; Fang, Y.; Zhang, W.S.; et al. Exosomal miR-128-3p Promotes Epithelial-to-Mesenchymal Transition in Colorectal Cancer Cells by Targeting FOXO4 via TGF-beta/SMAD and JAK/STAT3 Signaling. Front. Cell Dev. Biol. 2021, 9, 568738. [Google Scholar] [CrossRef]
- Zhou, X.; Wen, W.; Shan, X.; Zhu, W.; Xu, J.; Guo, R.H.; Cheng, W.F.; Wang, F.; Qi, L.W.; Chen, Y.; et al. A six-microRNA panel in plasma was identified as a potential biomarker for lung adenocarcinoma diagnosis. Oncotarget 2017, 8, 6513–6525. [Google Scholar] [CrossRef]
- Li, S.Q.; Yan, G.Q.; Yue, M.; Wang, L. Extracellular vesicles-derived microRNA-222 promotes immune escape via interacting with ATF3 to regulate AKT1 transcription in colorectal cancer. BMC Cancer 2021, 21, 349. [Google Scholar] [CrossRef]
- Schwarzenbach, H.; Gahan, P.B. MicroRNA Shuttle from Cell-To-Cell by Exosomes and Its Impact in Cancer. Non-Coding RNA 2019, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Luo, F.; Wang, B.R.; Li, H.Q.; Xu, Y.; Liu, X.L.; Shi, L.; Lu, X.L.; Xu, W.C.; Lu, L.; et al. STAT3-regulated exosomal miR-21 promotes angiogenesis and is involved in neoplastic processes of transformed human bronchial epithelial cells. Cancer Lett. 2016, 370, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.Y.; Fong, M.Y.; Min, Y.F.; Somlo, G.; Liu, L.; Palomares, M.R.; Yu, Y.; Chow, A.; O’Connor, S.T.F.; Chin, A.R.; et al. Cancer-Secreted miR-105 Destroys Vascular Endothelial Barriers to Promote Metastasis. Cancer Cell 2014, 25, 501–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, T.F.; Carlsen, A.L.; Heegaard, N.H.H.; Sorensen, F.B.; Jakobsen, A. Changes in circulating microRNA-126 during treatment with chemotherapy and bevacizumab predicts treatment response in patients with metastatic colorectal cancer. Br. J. Cancer 2015, 112, 624–629. [Google Scholar] [CrossRef]
- Nemes, K.; Csoka, M.; Nagy, N.; Mark, A.; Varadi, Z.; Danko, T.; Kovacs, G.; Kopper, L.; Sebestyen, A. Expression of Certain Leukemia/Lymphoma Related microRNAs and its Correlation with Prognosis in Childhood Acute Lymphoblastic Leukemia. Pathol. Oncol. Res. 2015, 21, 597–604. [Google Scholar] [CrossRef]
- Mullan, F. Seasons of Survival—Reflections of a Physician with Cancer. N. Engl. J. Med. 1985, 313, 270–273. [Google Scholar] [CrossRef]
- NCCS (National Coalition of Cancer Survivorship). About NCCS. Available online: https://canceradvocacy.org/ (accessed on 15 March 2021).
- Richards, M.; Corner, J.; Maher, J. The National Cancer Survivorship Initiative: New and emerging evidence on the ongoing needs of cancer survivors. Br. J. Cancer 2011, 105, S1–S4. [Google Scholar] [CrossRef]
- Simonelli, C.; Annunziata, M.A.; Chimienti, E.; Berretta, M.; Tirelli, U. Cancer survivorship: A challenge for the European oncologists. Ann. Oncol. 2008, 19, 1216–1217. [Google Scholar] [CrossRef] [PubMed]
- Dal Maso, L.; Guzzinati, S.; Buzzoni, C.; Capocaccia, R.; Serraino, D.; Caldarella, A.; Dei Tos, A.P.; Falcini, F.; Autelitano, M.; Masanotti, G.; et al. Long-term survival, prevalence, and cure of cancer: A population-based estimation for 818,902 Italian patients and 26 cancer types. Ann. Oncol. 2014, 25, 2251–2260. [Google Scholar] [CrossRef]
- Dal Maso, L.; Panato, C.; Guzzinati, S.; Serraino, D.; Francisci, S.; Botta, L.; Capocaccia, R.; Tavilla, A.; Gigli, A.; Crocetti, E.; et al. Prognosis and cure of long-term cancer survivors: A population-based estimation. Cancer Med. 2019, 8, 4497–4507. [Google Scholar] [CrossRef] [PubMed]
- Berry, L.L.; Davis, S.W.; Flynn, A.G.; Landercasper, J.; Deming, K.A. Is it time to reconsider the term “cancer survivor”? J. Psychosoc. Oncol. 2019, 37, 413–426. [Google Scholar] [CrossRef]
- Marzorati, C.; Riva, S.; Pravettoni, G. Who Is a Cancer Survivor? A Systematic Review of Published Definitions. J. Cancer Educ. 2017, 32, 228–237. [Google Scholar] [CrossRef]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [Green Version]
- Berry, D.A.; Cronin, K.A.; Plevritis, S.K.; Fryback, D.G.; Clarke, L.; Zelen, M.; Mandelblatt, J.S.; Yakovlev, A.Y.; Habbema, J.D.F.; Feuer, E.J.; et al. Effect of screening and adjuvant therapy on mortality from breast cancer. N. Engl. J. Med. 2005, 353, 1784–1792. [Google Scholar] [CrossRef] [PubMed]
- Gegechkori, N.; Haines, L.; Lin, J.J. Long-Term and Latent Side Effects of Specific Cancer Types. Med. Clin. N. Am. 2017, 101, 1053–1073. [Google Scholar] [CrossRef]
- Saylor, P.J.; Smith, M.R. Metabolic Complications of Androgen Deprivation Therapy for Prostate Cancer. J. Urol. 2009, 181, 1998–2006. [Google Scholar] [CrossRef]
- Morgan, M.A. Cancer Survivorship: History, Quality-of-Life Issues, and the Evolving Multidisciplinary Approach to Implementation of Cancer Survivorship Care Plans. Oncol. Nurs. Forum 2009, 36, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Fidler-Benaoudia, M.; Keegan, T.H.; Hipp, H.S.; Jemal, A.; Siegel, R.L. Cancer statistics for adolescents and young adults, 2020. Cancer J. Clin. 2020, 70, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Buccafusca, G.; Proserpio, I.; Tralongo, A.C.; Giuliano, S.R.; Tralongo, P. Early colorectal cancer: Diagnosis, treatment and survivorship care. Crit. Rev. Oncol. Hemat. 2019, 136, 20–30. [Google Scholar] [CrossRef]
- Kanwal, R.; Plaga, A.R.; Liu, X.Q.; Shukla, G.C.; Gupta, S. MicroRNAs in prostate cancer: Functional role as biomarkers. Cancer Lett. 2017, 407, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Asakura, K.; Kadota, T.; Matsuzaki, J.; Yoshida, Y.; Yamamoto, Y.; Nakagawa, K.; Takizawa, S.; Aoki, Y.; Nakamura, E.; Miura, J.; et al. A miRNA-based diagnostic model predicts resectable lung cancer in humans with high accuracy. Commun. Biol. 2020, 3, 134. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Wong, Y.S.; Goh, B.K.P.; Chan, C.; Cheow, P.C.; Chow, P.K.H.; Lim, T.K.H.; Goh, G.B.B.; Krishnamoorthy, T.L.; Kumar, R.; et al. Circulating microRNAs as Potential Diagnostic and Prognostic Biomarkers in Hepatocellular Carcinoma. Sci. Rep. 2019, 9, 10464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natarajan, L.; Pu, M.Y.; Davies, S.R.; Vickery, T.L.; Nelson, S.H.; Pittman, E.; Parker, B.A.; Ellis, M.J.; Flatt, S.W.; Mardis, E.R.; et al. miRNAs and Long-term Breast Cancer Survival: Evidence from the WHEL Study. Cancer Epidem. Biomar. 2019, 28, 1525–1533. [Google Scholar] [CrossRef]
- Li, H.P.; Liu, J.; Chen, J.N.; Wang, H.Y.; Yang, L.B.; Chen, F.; Fan, S.T.; Wang, J.; Shao, B.; Yin, D.; et al. A serum microRNA signature predicts trastuzumab benefit in HER2-positive metastatic breast cancer patients. Nat. Commun. 2018, 9, 1614. [Google Scholar] [CrossRef]
- Lin, H.M.; Castillo, L.; Mahon, K.L.; Chiam, K.; Lee, B.Y.; Nguyen, Q.; Boyer, M.J.; Stockler, M.R.; Pavlakis, N.; Marx, G.; et al. Circulating microRNAs are associated with docetaxel chemotherapy outcome in castration-resistant prostate cancer. Br. J. Cancer 2014, 110, 2462–2471. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Cho, Y.E.; Cho, A.R.; Choi, W.J.; Yun, S.; Park, H.; Kim, H.S.; Cashion, A.K.; Gill, J.; Lee, H.; et al. The Possible Influence of Mediterranean Diet on Extracellular Vesicle miRNA Expression in Breast Cancer Survivors. Cancers 2020, 12, 1355. [Google Scholar] [CrossRef]
- Selth, L.A.; Townley, S.L.; Bert, A.G.; Stricker, P.D.; Sutherland, P.D.; Horvath, L.G.; Goodall, G.J.; Butler, L.M.; Tilley, W.D.; BioResour, A.P.C. Circulating microRNAs predict biochemical recurrence in prostate cancer patients. Br. J. Cancer 2013, 109, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Hoey, C.; Ahmed, M.; Ghiam, A.F.; Vesprini, D.; Huang, X.; Commisso, K.; Commisso, A.; Ray, J.; Fokas, E.; Loblaw, D.A.; et al. Circulating miRNAs as non-invasive biomarkers to predict aggressive prostate cancer after radical prostatectomy. J. Transl. Med. 2019, 17, 173. [Google Scholar] [CrossRef]
- Akazawa, Y.; Mizuno, S.; Fujinami, N.; Suzuki, T.; Yoshioka, Y.; Ochiya, T.; Nakamoto, Y.; Nakatsura, T. Usefulness of serum microRNA as a predictive marker of recurrence and prognosis in biliary tract cancer after radical surgery. Sci. Rep. 2019, 9, 5925. [Google Scholar] [CrossRef]
- Chen, Y.; Jungsuwadee, P.; Vore, M.; Butterfield, D.A.; Clair, D.K.S. Collateral damage in cancer chemotherapy—Oxidative stress in nontargeted tissues. Mol. Interv. 2007, 7, 147–156. [Google Scholar] [CrossRef]
- Yarana, C.; St Clair, D.K. Chemotherapy-Induced Tissue Injury: An Insight into the Role of Extracellular Vesicles-Mediated Oxidative Stress Responses. Antioxidants 2017, 6, 75. [Google Scholar] [CrossRef] [PubMed]
- Yarana, C.; Carroll, D.; Chen, J.; Chaiswing, L.; Zhao, Y.M.; Noel, T.; Alstott, M.; Bae, Y.; Dressler, E.V.; Moscow, J.A.; et al. Extracellular Vesicles Released by Cardiomyocytes in a Doxorubicin-Induced Cardiac Injury Mouse Model Contain Protein Biomarkers of Early Cardiac Injury. Clin. Cancer Res. 2018, 24, 1644–1653. [Google Scholar] [CrossRef] [Green Version]
- Rigaud, V.O.C.; Ferreira, L.R.P.; Ayub-Ferreira, S.M.; Avila, M.S.; Brandao, S.M.G.; Cruz, F.D.; Santos, M.H.H.; Cruz, C.B.B.V.; Alves, M.S.L.; Issa, V.S.; et al. Circulating miR-1 as a potential biomarker of doxorubicin-induced cardiotoxicity in breast cancer patients. Oncotarget 2017, 8, 6994–7002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggeri, C.; Gioffre, S.; Achilli, F.; Colombo, G.I.; D’Alessandra, Y. Role of microRNAs in doxorubicin-induced cardiotoxicity: An overview of preclinical models and cancer patients. Heart Fail Rev. 2018, 23, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Freres, P.; Bouznad, N.; Servais, L.; Josse, C.; Wenric, S.; Poncin, A.; Thiry, J.; Moonen, M.; Oury, C.; Lancellotti, P.; et al. Variations of circulating cardiac biomarkers during and after anthracycline-containing chemotherapy in breast cancer patients. BMC Cancer 2018, 18, 102. [Google Scholar] [CrossRef] [PubMed]
- Perez, D.D.; Martinez, A.R.; Palomo, A.O.; Urena, M.D.; Puche, J.L.G.; Remacho, A.R.; Hernandez, J.E.; Acosta, J.A.L.; Sanchez, F.G.O.; Serrano, M.J. Extracellular vesicle-miRNAs as liquid biopsy biomarkers for disease identification and prognosis in metastatic colorectal cancer patients. Sci. Rep. 2020, 10, 3974. [Google Scholar] [CrossRef] [Green Version]
- Callegari, E.; D’Abundo, L.; Guerriero, P.; Simioni, C.; Elamin, B.K.; Russo, M.; Cani, A.; Bassi, C.; Zagatti, B.; Giacomelli, L.; et al. miR-199a-3p Modulates MTOR and PAK4 Pathways and Inhibits Tumor Growth in a Hepatocellular Carcinoma Transgenic Mouse Model. Mol. Ther. Nucl. Acids 2018, 11, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Dufresne, S.; Rebillard, A.; Muti, P.; Friedenreich, C.M.; Brenner, D.R. A Review of Physical Activity and Circulating miRNA Expression: Implications in Cancer Risk and Progression. Cancer Epidem. Biomar. 2018, 27, 11–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kura, B.; Parikh, M.; Slezak, J.; Pierce, G.N. The Influence of Diet on MicroRNAs that Impact Cardiovascular Disease. Molecules 2019, 24, 1509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafson, D.; Tyryshkin, K.; Renwick, N. microRNA-guided diagnostics in clinical samples. Best Pract. Res. Clin. Endocrinol. 2016, 30, 563–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozawa, P.M.M.; Jucoski, T.S.; Vieira, E.; Carvalho, T.M.; Malheiros, D.; Ribeiro, E.M.D.F. Liquid biopsy for breast cancer using extracellular vesicles and cell-free microRNAs as biomarkers. Transl. Res. 2020, 223, 40–60. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).