
Differences in a Single Extracellular Residue Underlie Adhesive Functions of Two Zebrafish Aqp0s


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell-to-Cell Adhesion Assay
2.3. Statistical Analysis
2.4. Western Blot Analysis
2.5. Mutagenesis
3. Results
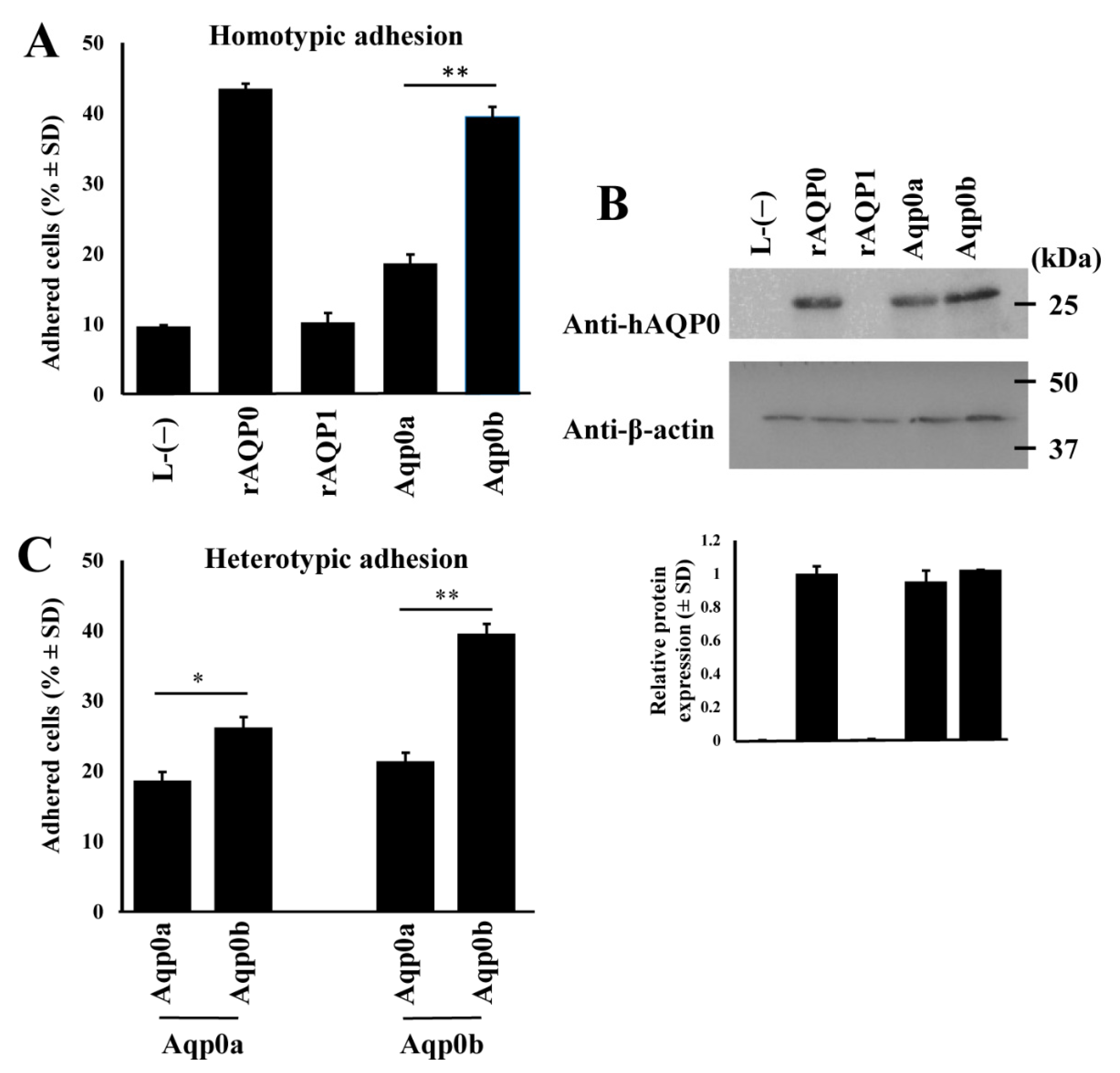
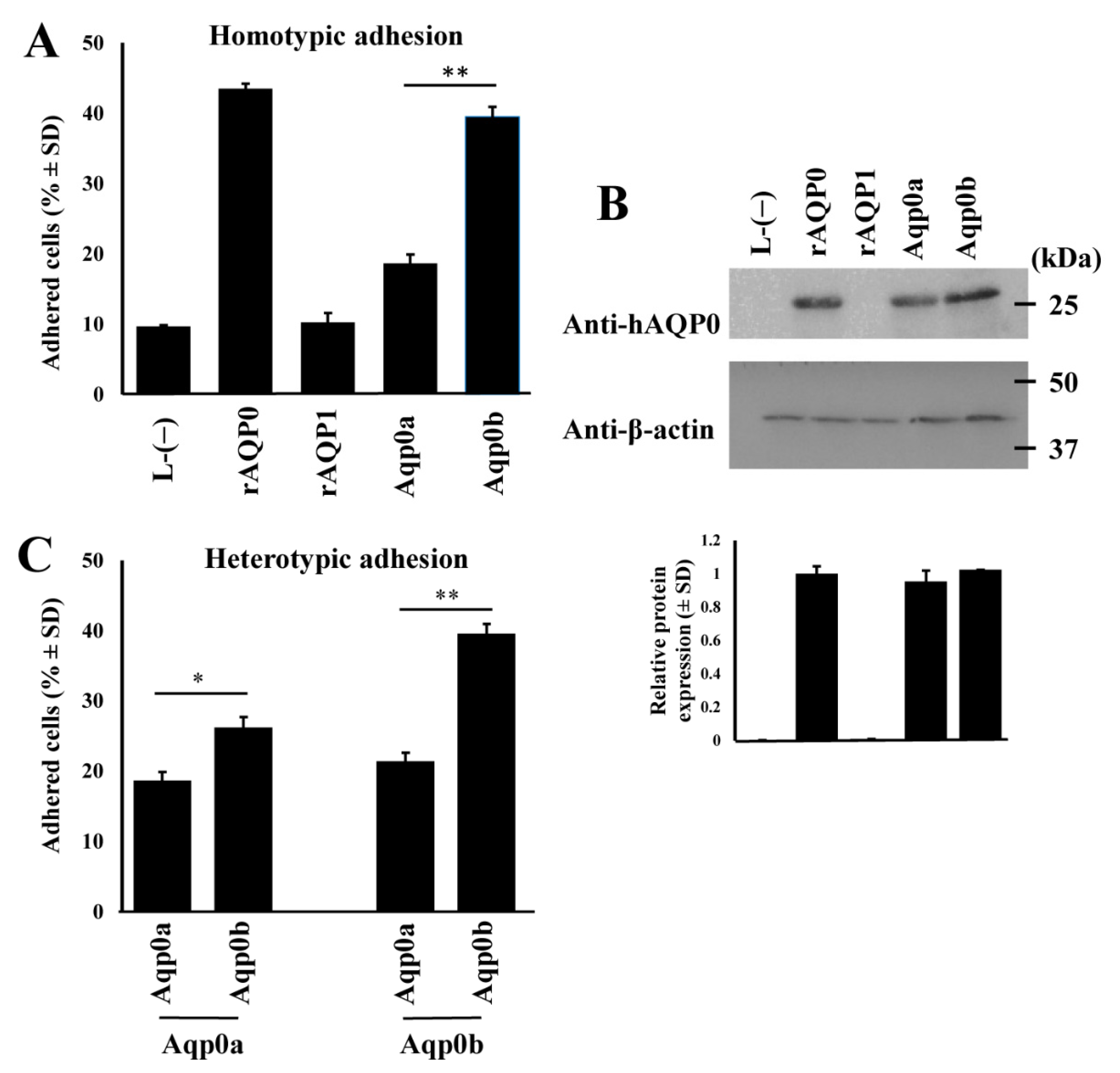
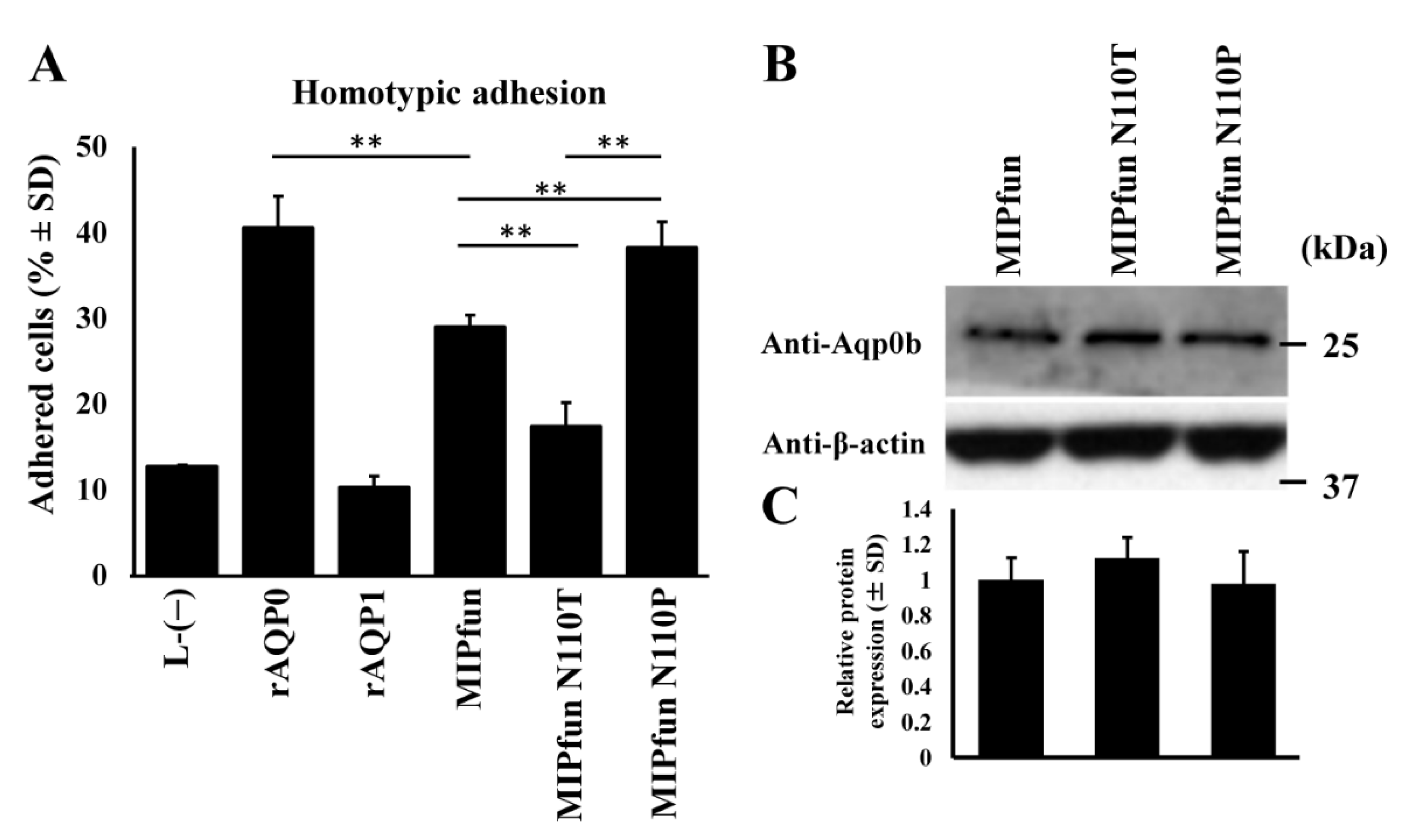
3.1. Zebrafish Aqp0b Has Adhesive Properties
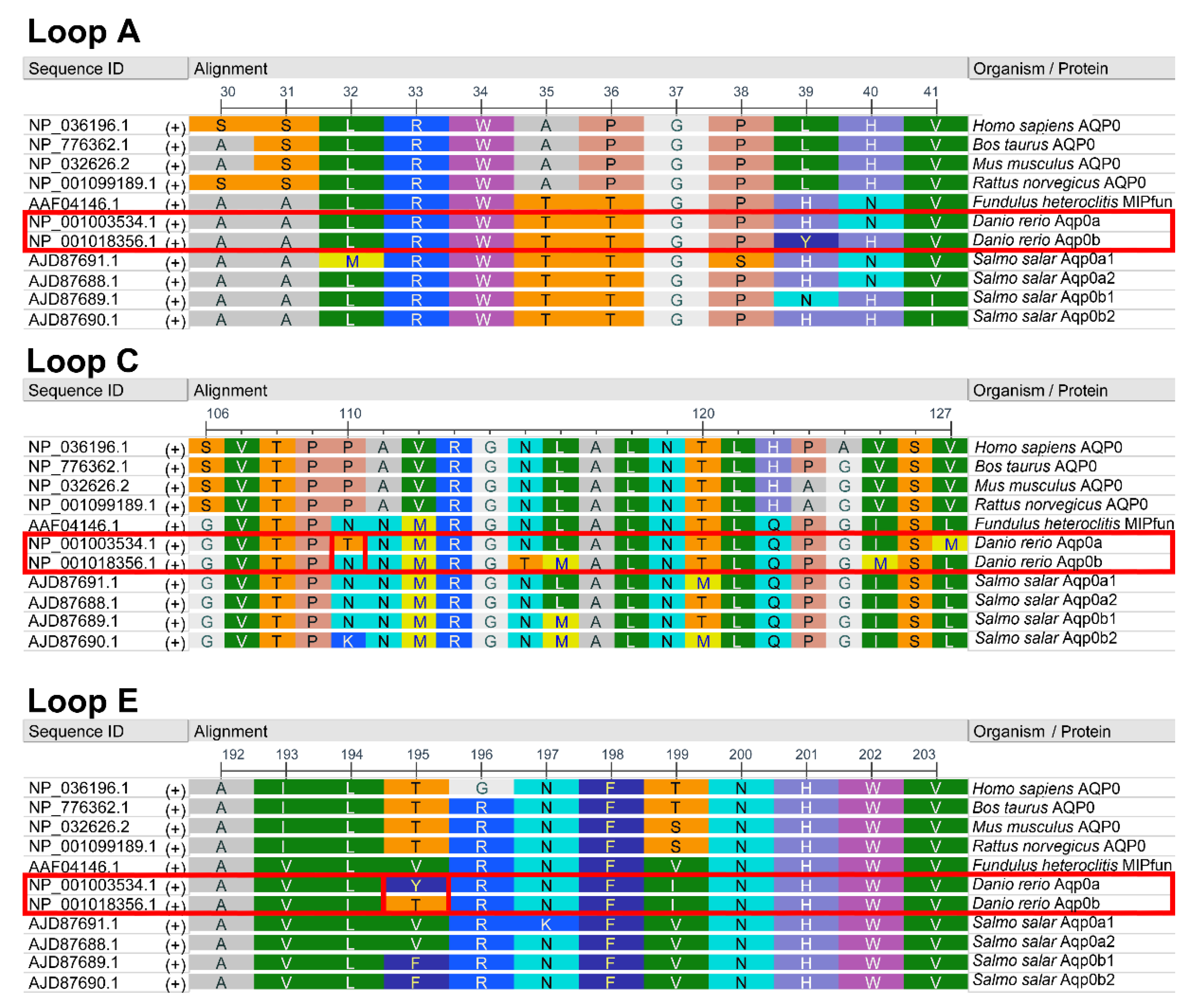
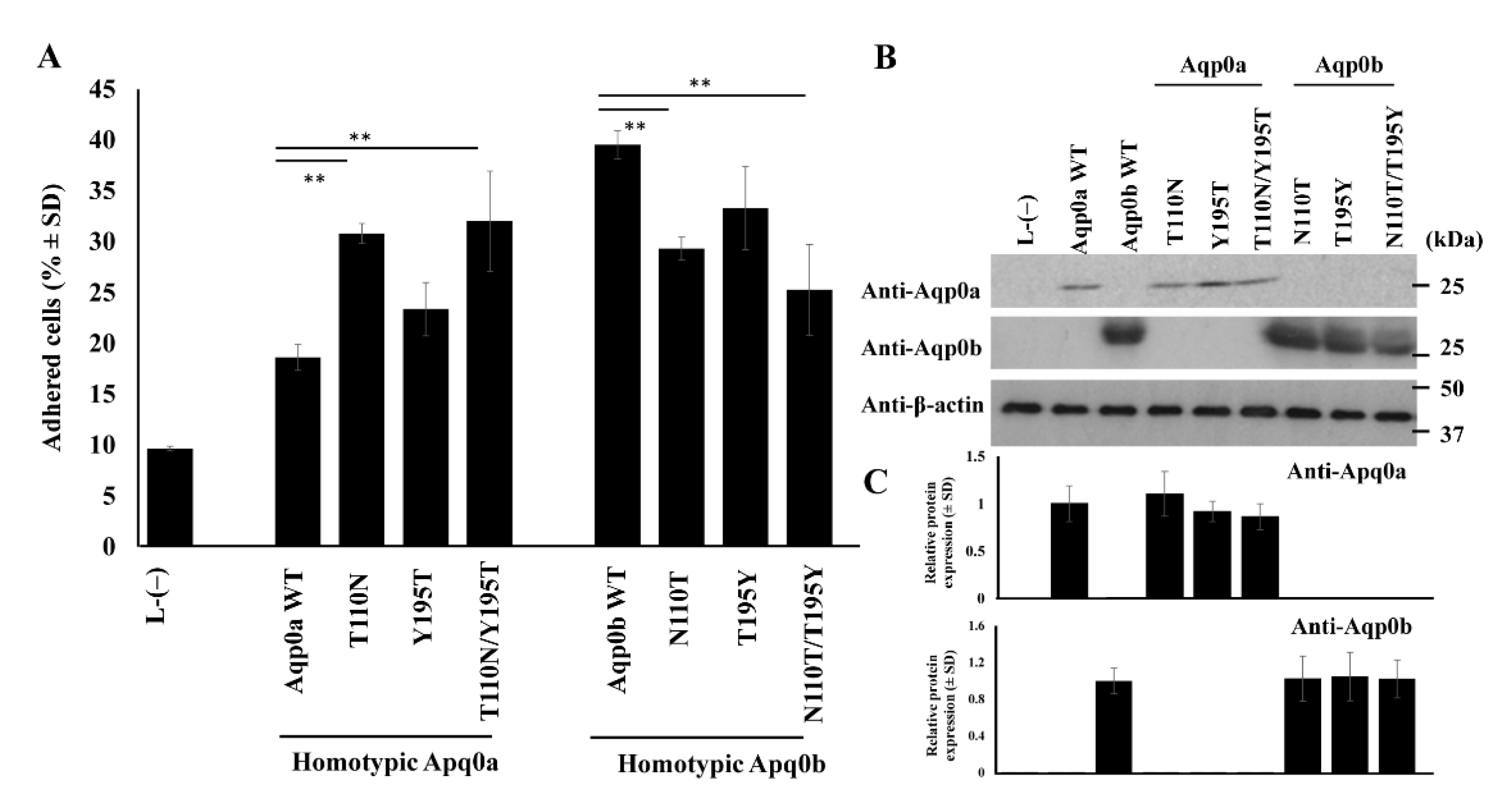
3.2. Residues 110 and 195 Differ between Aqp0a and Aqp0b
3.3. N110 Is Key in Aqp0b-Mediated Adhesion
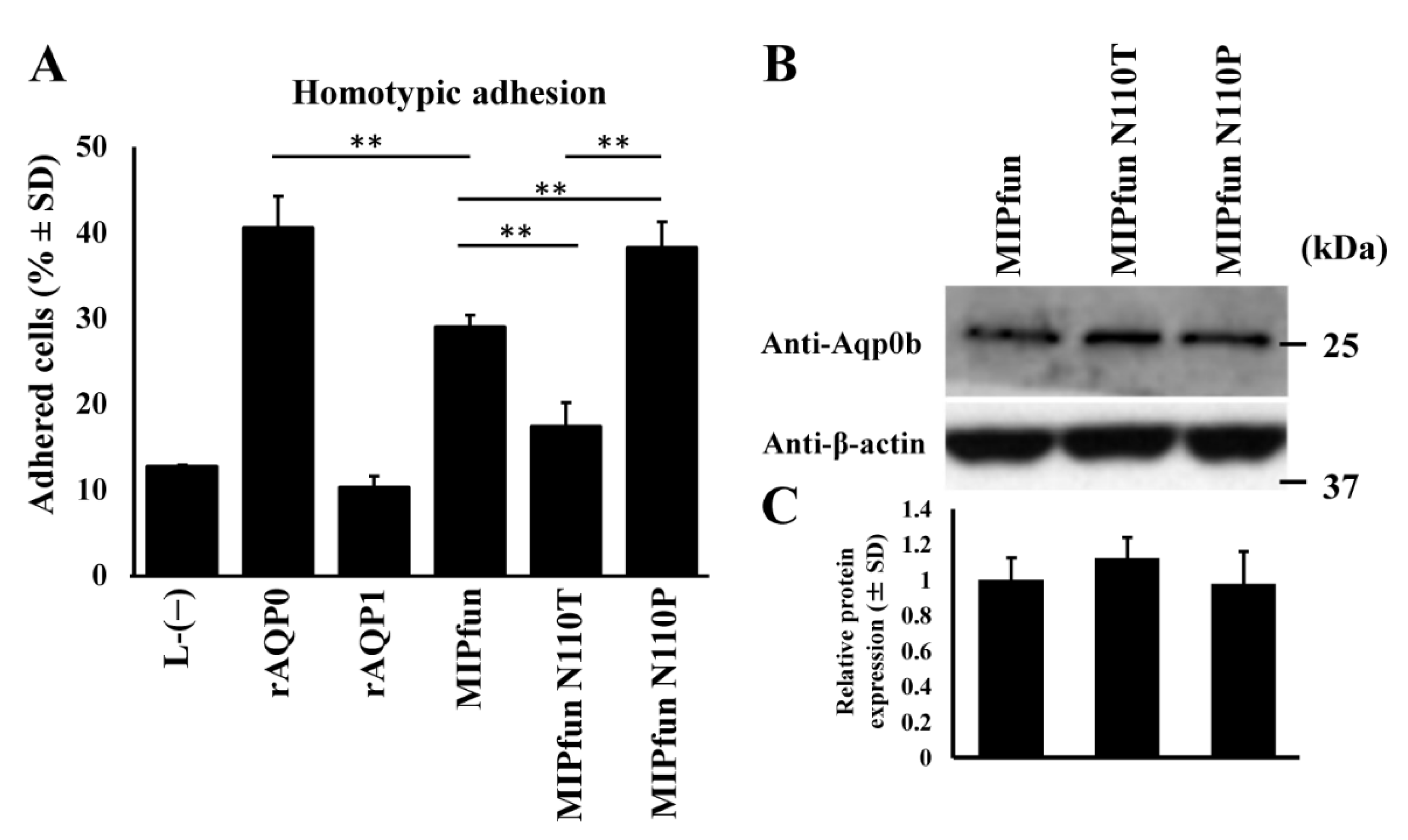
3.4. MIPfun and Aqp0b Have Similar Adhesive Properties
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shiels, A.; Bennett, T.M.; Hejtmancik, J.F. Fielding Cat-Map: Putting Cataract on the Map. Mol. Vis. 2010, 16, 2007. [Google Scholar] [PubMed]
- Liu, J.; Xu, J.; Gu, S.; Nicholson, B.J.; Jiang, J.X. Aquaporin 0 Enhances Gap Junction Coupling via Its Cell Adhesion Function and Interaction with Connexin 50. J. Cell Sci. 2011, 124, 198–206. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, Y.; Oka, M.; Furuki, K.; Mitsuishi, A.; Nakashima, E.; Takehana, M. The Effect of the Interaction between Aquaporin 0 (AQP0) and the Filensin Tail Region on AQP0 Water Permeability. Mol. Vis. 2011, 17, 3191–3199. [Google Scholar] [PubMed]
- Rose, K.M.L.; Gourdie, R.G.; Prescott, A.R.; Quinlan, R.A.; Crouch, R.K.; Schey, K.L. The C Terminus of Lens Aquaporin 0 Interacts with the Cytoskeletal Proteins Filensin and CP49. Invest. Ophthalmol. Vis. Sci. 2006, 47, 1562. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.-K.; Harding, C.V. Square Arrays and Their Role in Ridge Formation in Human Lens Fibers. J. Ultrastruct. Res. 1984, 86, 228–245. [Google Scholar] [CrossRef]
- Zampighi, G.A.; Hall, J.E.; Ehring, G.R.; Simon, S.A. The Structural Organization and Protein Composition of Lens Fiber Junctions. J. Cell Biol. 1989, 108, 2255–2275. [Google Scholar] [CrossRef]
- Zampighi, G.A.; Eskandari, S.; Hall, J.E.; Zampighi, L.; Kreman, M. Micro-Domains of AQP0 in Lens Equatorial Fibers. Exp. Eye Res. 2002, 75, 505–519. [Google Scholar] [CrossRef]
- Kumari, S.S.; Varadaraj, K. Intact AQP0 Performs Cell-to-Cell Adhesion. Biochem. Biophys. Res. Commun. 2009, 390, 1034–1039. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.S.; Gandhi, J.; Mustehsan, M.H.; Eren, S.; Varadaraj, K. Functional Characterization of an AQP0 Missense Mutation, R33C, That Causes Dominant Congenital Lens Cataract, Reveals Impaired Cell-to-Cell Adhesion. Exp. Eye Res. 2013, 116, 371–385. [Google Scholar] [CrossRef] [Green Version]
- Michea, L.F.; de la Fuente, M.; Lagos, N. Lens Major Intrinsic Protein (MIP) Promotes Adhesion When Reconstituted into Large Unilamellar Liposomes. Biochemistry 1994, 33, 7663–7669. [Google Scholar] [CrossRef]
- Varadaraj, K.; Kumari, S.S. Molecular Mechanism of Aquaporin 0-Induced Fiber Cell to Fiber Cell Adhesion in the Eye Lens. Biochem. Biophys. Res. Commun. 2018, 506, 284–289. [Google Scholar] [CrossRef]
- Gonen, T.; Sliz, P.; Kistler, J.; Cheng, Y.; Walz, T. Aquaporin-0 Membrane Junctions Reveal the Structure of a Closed Water Pore. Nature 2004, 429, 5. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, Y.; Oka, M.; Funakoshi-Tago, M.; Tamura, H.; Takehana, M. The Extracellular C-Loop Domain Plays an Important Role in the Cell Adhesion Function of Aquaporin 0. Curr. Eye Res. 2017, 42, 617–624. [Google Scholar] [CrossRef]
- Chauvigné, F.; Zapater, C.; Stavang, J.A.; Taranger, G.L.; Cerdà, J.; Finn, R.N. The PH Sensitivity of Aqp0 Channels in Tetraploid and Diploid Teleosts. Faseb J. 2015, 29, 2172–2184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauvigné, F.; Fjelldal, P.G.; Cerdà, J.; Finn, R.N. Auto-Adhesion Potential of Extraocular Aqp0 during Teleost Development. PLoS ONE 2016, 11, e0154592. [Google Scholar] [CrossRef] [Green Version]
- Clemens, D.M.; Nemeth-Cahalan, K.L.; Trinh, L.; Zhang, T.; Schilling, T.F.; Hall, J.E. In Vivo Analysis of Aquaporin 0 Function in Zebrafish: Permeability Regulation Is Required for Lens Transparency. Investig. Ophthalmol. Vis. Sci. 2013, 54, 5136–5143. [Google Scholar] [CrossRef] [PubMed]
- Froger, A.; Clemens, D.; Kalman, K.; Németh-Cahalan, K.L.; Schilling, T.F.; Hall, J.E. Two Distinct Aquaporin 0s Required for Development and Transparency of the Zebrafish Lens. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6582–6592. [Google Scholar] [CrossRef] [PubMed]
- Vorontsova, I.; Gehring, I.; Hall, J.E.; Schilling, T.F. Aqp0a Regulates Suture Stability in the Zebrafish Lens. Investig. Ophthalmol. Vis. Sci. 2018, 59, 2869–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vihtelic, T.S.; Fadool, J.M.; Gao, J.; Thornton, K.A.; Hyde, D.R.; Wistow, G. Expressed Sequence Tag Analysis of Zebrafish Eye Tissues for NEIBank. Mol. Vis. 2005, 11, 1083–1100. [Google Scholar]
- Wang, K.; Vorontsova, I.; Hoshino, M.; Uesugi, K.; Yagi, N.; Hall, J.E.; Schilling, T.F.; Pierscionek, B.K. Aquaporins Have Regional Functions in Development of Refractive Index in the Zebrafish Eye Lens. Investig. Ophthalmol. Vis. Sci. 2021, 62, 23. [Google Scholar] [CrossRef]
- Hu, Z.; Shi, W.; Riquelme, M.A.; Shi, Q.; Biswas, S.; Lo, W.-K.; White, T.W.; Gu, S.; Jiang, J.X. Connexin 50 Functions as an Adhesive Molecule and Promotes Lens Cell Differentiation. Sci. Rep. 2017, 7, 5298. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Gokhin, D.S.; Nowak, R.B.; Fowler, V.M. Sequential Application of Glass Coverslips to Assess the Compressive Stiffness of the Mouse Lens: Strain and Morphometric Analyses. J. Vis. Exp. 2016, 111, e53986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collery, R.F.; Veth, K.N.; Dubis, A.M.; Carroll, J.; Link, B.A. Rapid, Accurate, and Non-Invasive Measurement of Zebrafish Axial Length and Other Eye Dimensions Using SD-OCT Allows Longitudinal Analysis of Myopia and Emmetropization. PLoS ONE 2014, 9, e110699. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Protein | C-Loop | Auto-Adhesive | Reference |
|---|---|---|---|
| HsAQP0 BtAQP0 MsAQP0 RtAQP0 MIPfun SsAqp0a1 SsAqp0a2 SsAqp0b1 SsAqp0b2 DrAqp0a DrAqp0b | PPAVR PPAVR PPAVR PPAVR PNNMR PNNMR PNNMR PNNMR PKNMR PTNMR PNNMR | Yes ? Yes Yes Yes Yes Yes Yes Yes No Yes | (Kumari and Varadaraj, 2009) (Varadaraj and Kumari, 2018) (Nakazawa et al., 2017) This study (Chauvigné et al., 2016) (Chauvigné et al., 2016) (Chauvigné et al., 2016) (Chauvigné et al., 2016) This study This study |
| HsAQP1 106–110 | SSLTG | No | (Kumari and Varadaraj, 2009) |
| DrAqp08b | SSE | No | (Chauvigné et al., 2016) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vorontsova, I.; Hall, J.E.; Schilling, T.F.; Nagai, N.; Nakazawa, Y. Differences in a Single Extracellular Residue Underlie Adhesive Functions of Two Zebrafish Aqp0s. Cells 2021, 10, 2005. https://doi.org/10.3390/cells10082005
Vorontsova I, Hall JE, Schilling TF, Nagai N, Nakazawa Y. Differences in a Single Extracellular Residue Underlie Adhesive Functions of Two Zebrafish Aqp0s. Cells. 2021; 10(8):2005. https://doi.org/10.3390/cells10082005
Chicago/Turabian StyleVorontsova, Irene, James E. Hall, Thomas F. Schilling, Noriaki Nagai, and Yosuke Nakazawa. 2021. "Differences in a Single Extracellular Residue Underlie Adhesive Functions of Two Zebrafish Aqp0s" Cells 10, no. 8: 2005. https://doi.org/10.3390/cells10082005
APA StyleVorontsova, I., Hall, J. E., Schilling, T. F., Nagai, N., & Nakazawa, Y. (2021). Differences in a Single Extracellular Residue Underlie Adhesive Functions of Two Zebrafish Aqp0s. Cells, 10(8), 2005. https://doi.org/10.3390/cells10082005