
ZC3HC1 Is a Novel Inherent Component of the Nuclear Basket, Resident in a State of Reciprocal Dependence with TPR



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Antibodies
2.2. Animal Tissues and Primary Cells
2.3. Scanning Electron Microscopy and Immuno-SEM
2.4. High-Pressure Freezing, Freeze-Substitution, and Post-Embedding Immuno-TEM
2.5. Cell Culture
2.6. Synchronisation, Transfection, and Post-transcriptional Gene Silencing of Cultured Cells
2.7. Immunofluorescence Microscopy
2.8. Cell Fractionation and Egg Extracts
2.9. Immunoprecipitation, Phosphatase Treatment, and Immunoblotting
2.10. ZC3HC1 Gene Disruption in Human Cell Lines
3. Results
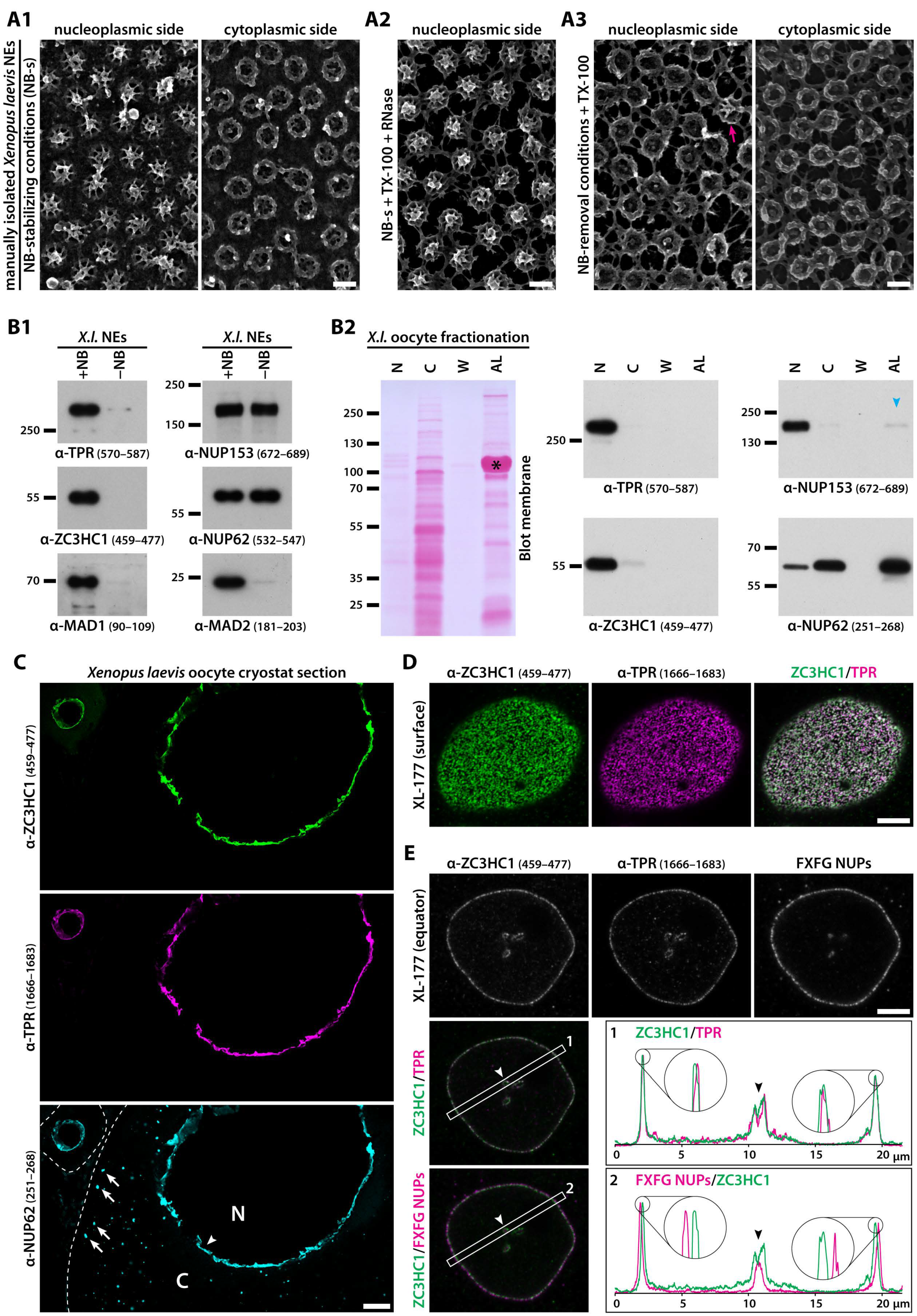
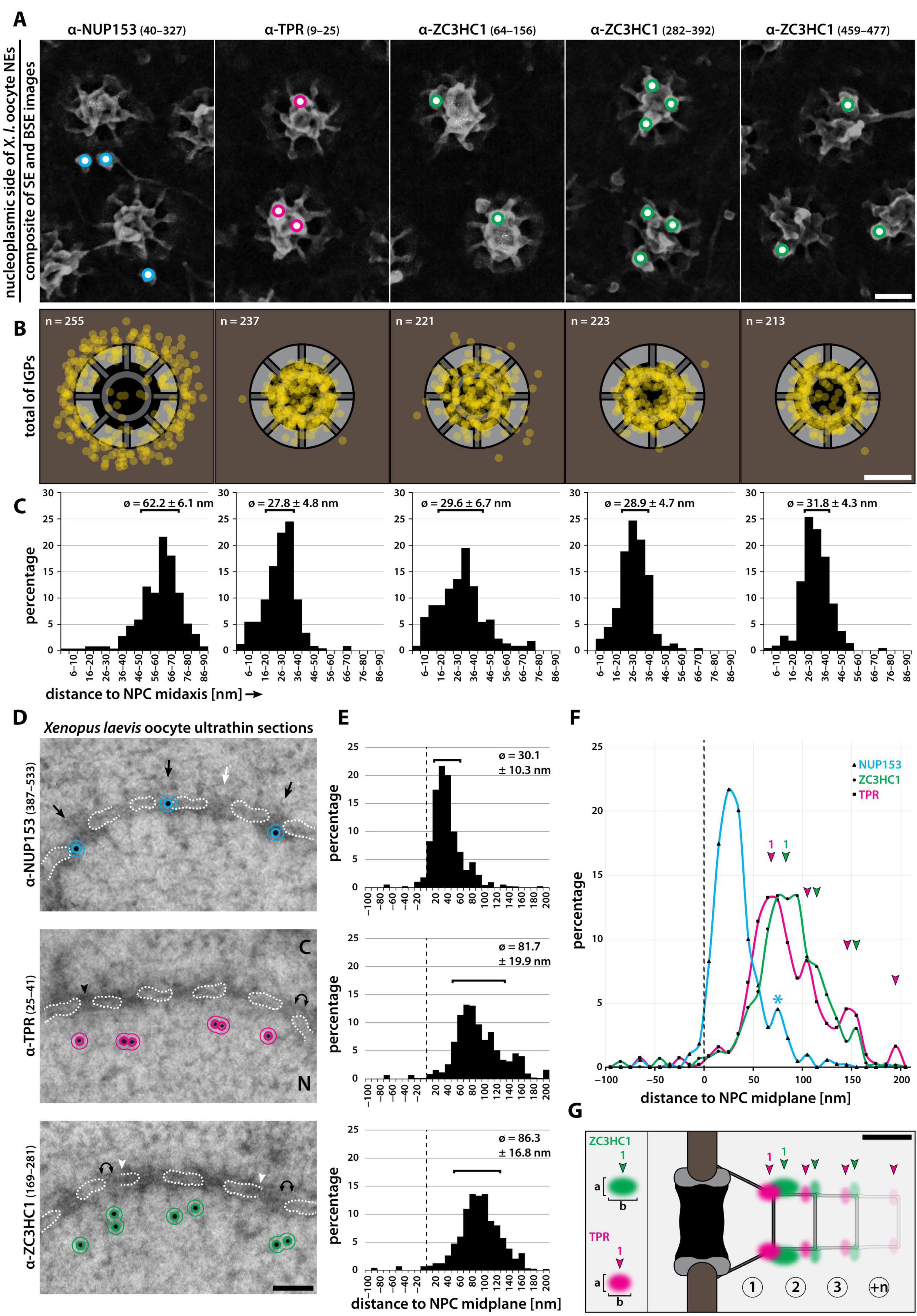
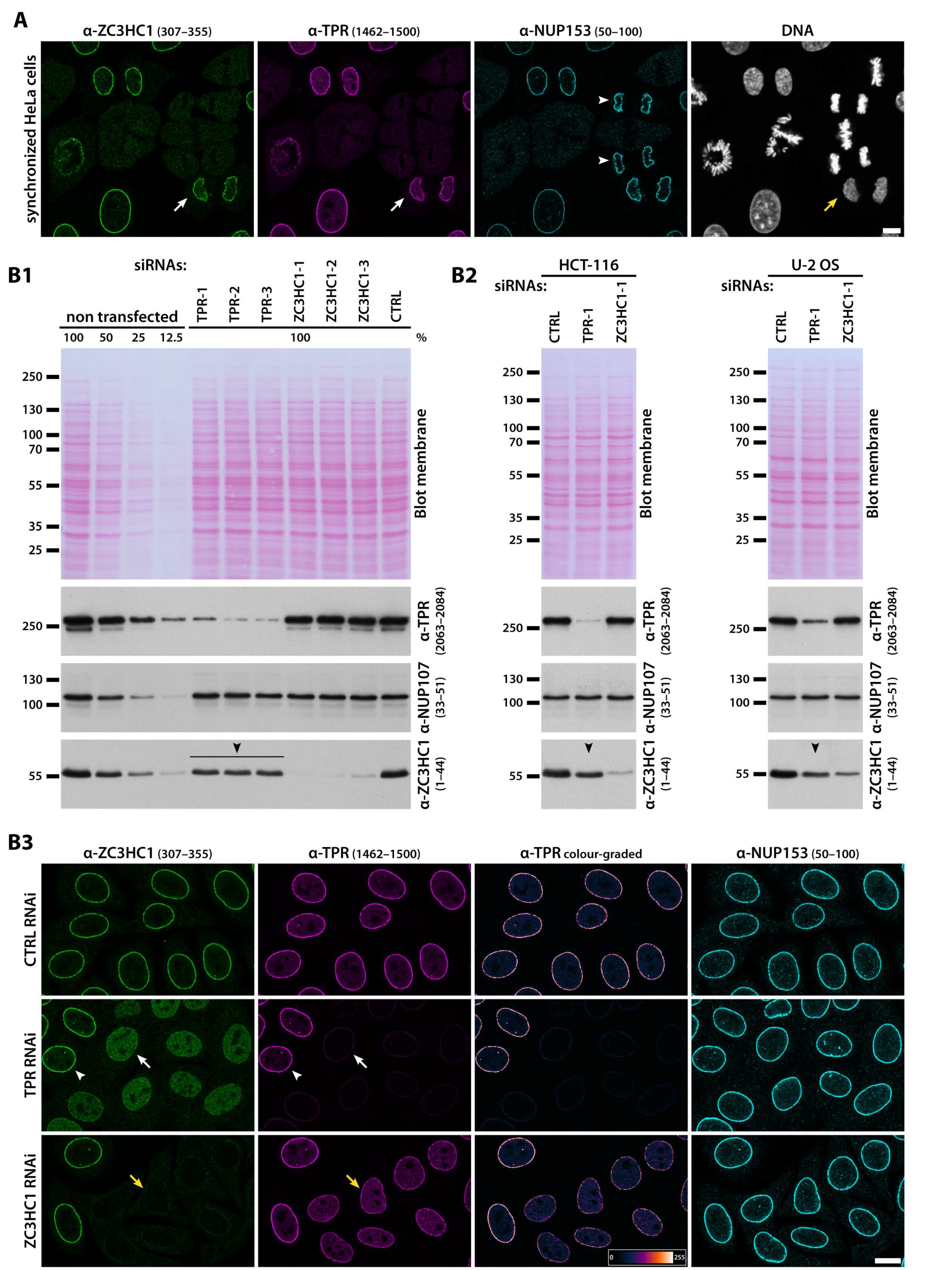
3.1. ZC3HC1 Is an NB-Resident Protein
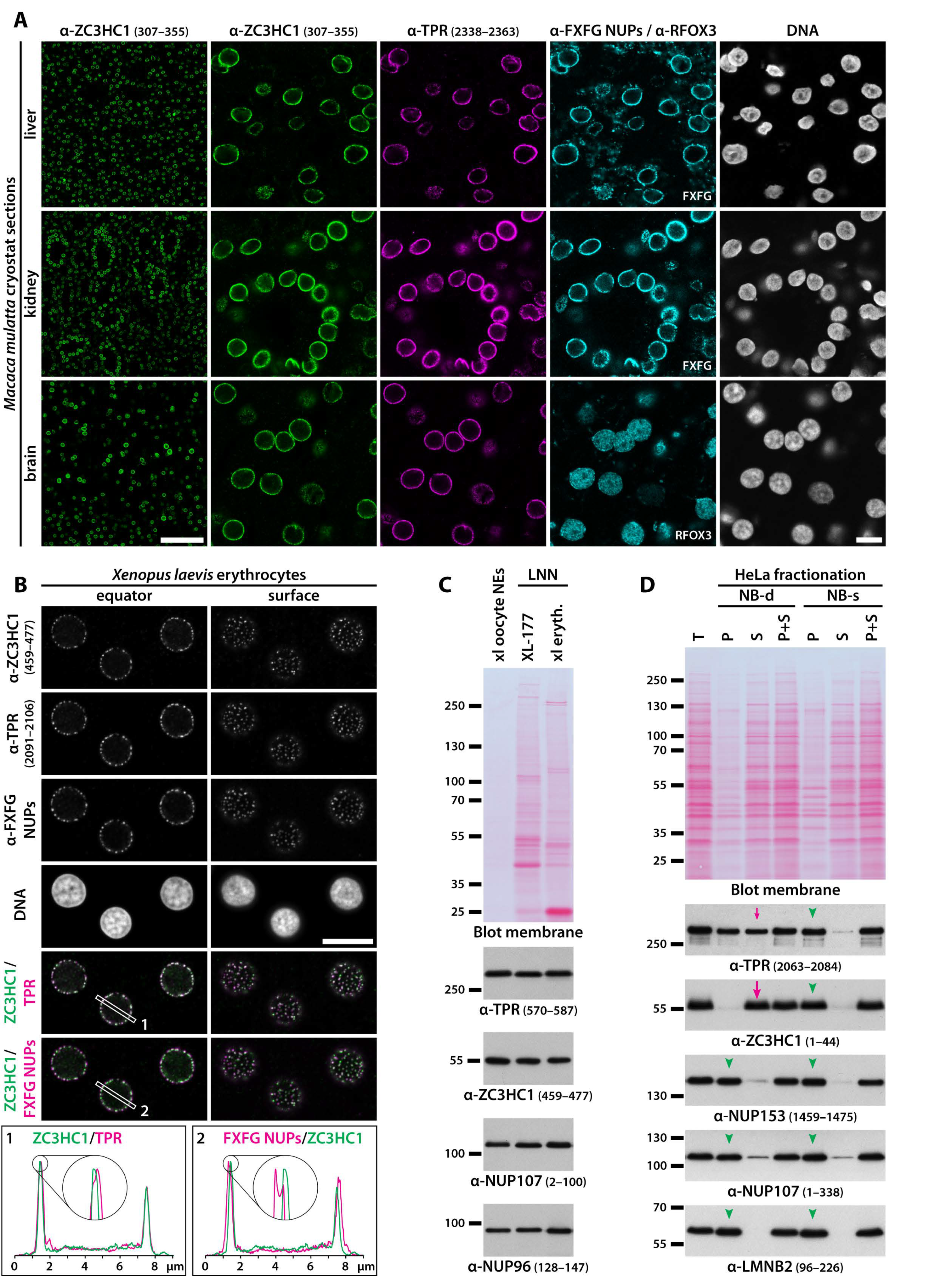
3.2. Residency of ZC3HC1 at the NE and Colocalisation with NB Scaffold Protein TPR Is Common in Proliferating Cells and Post-Mitotic Tissue Cells of Different Germ Layer Origin
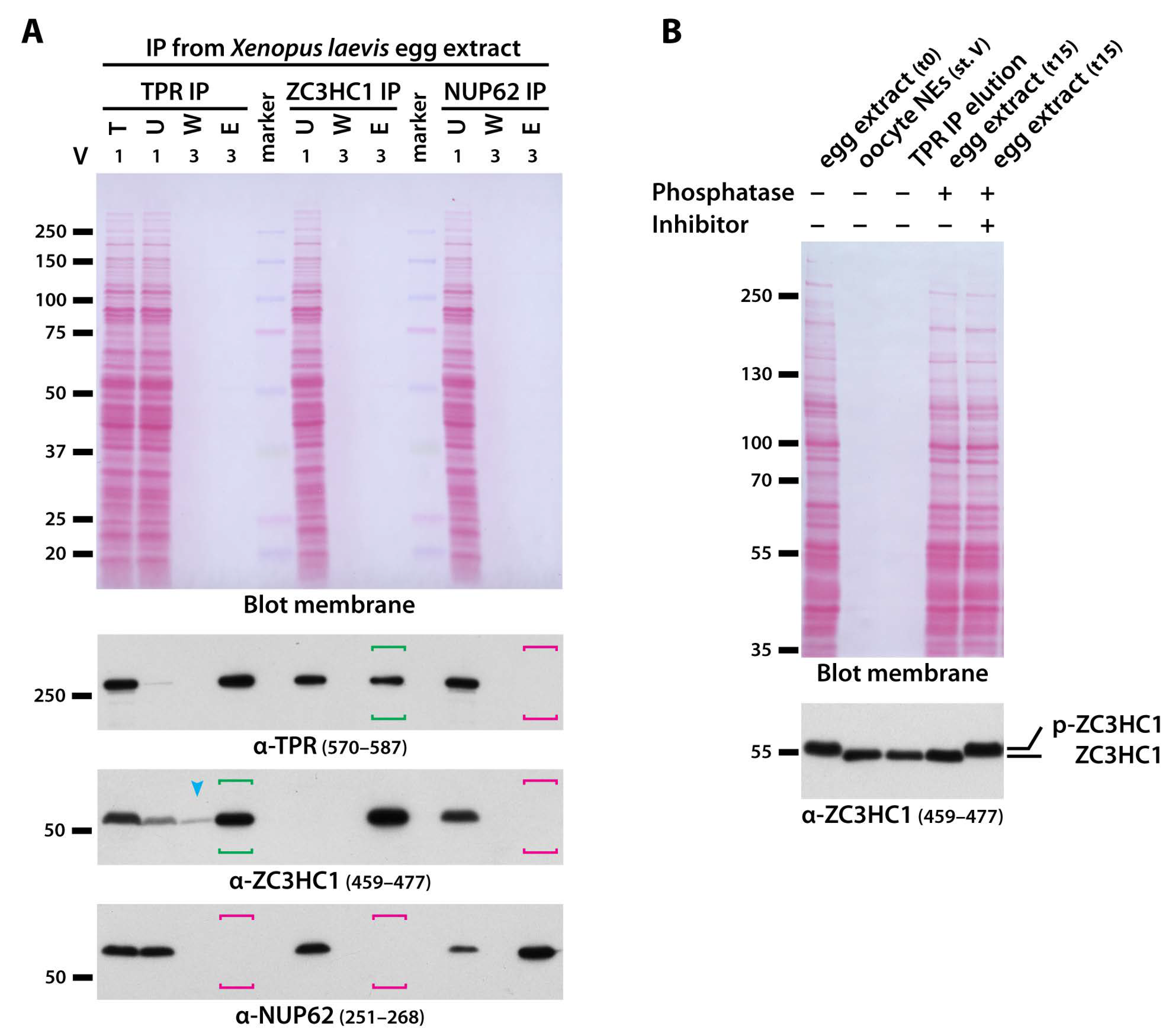
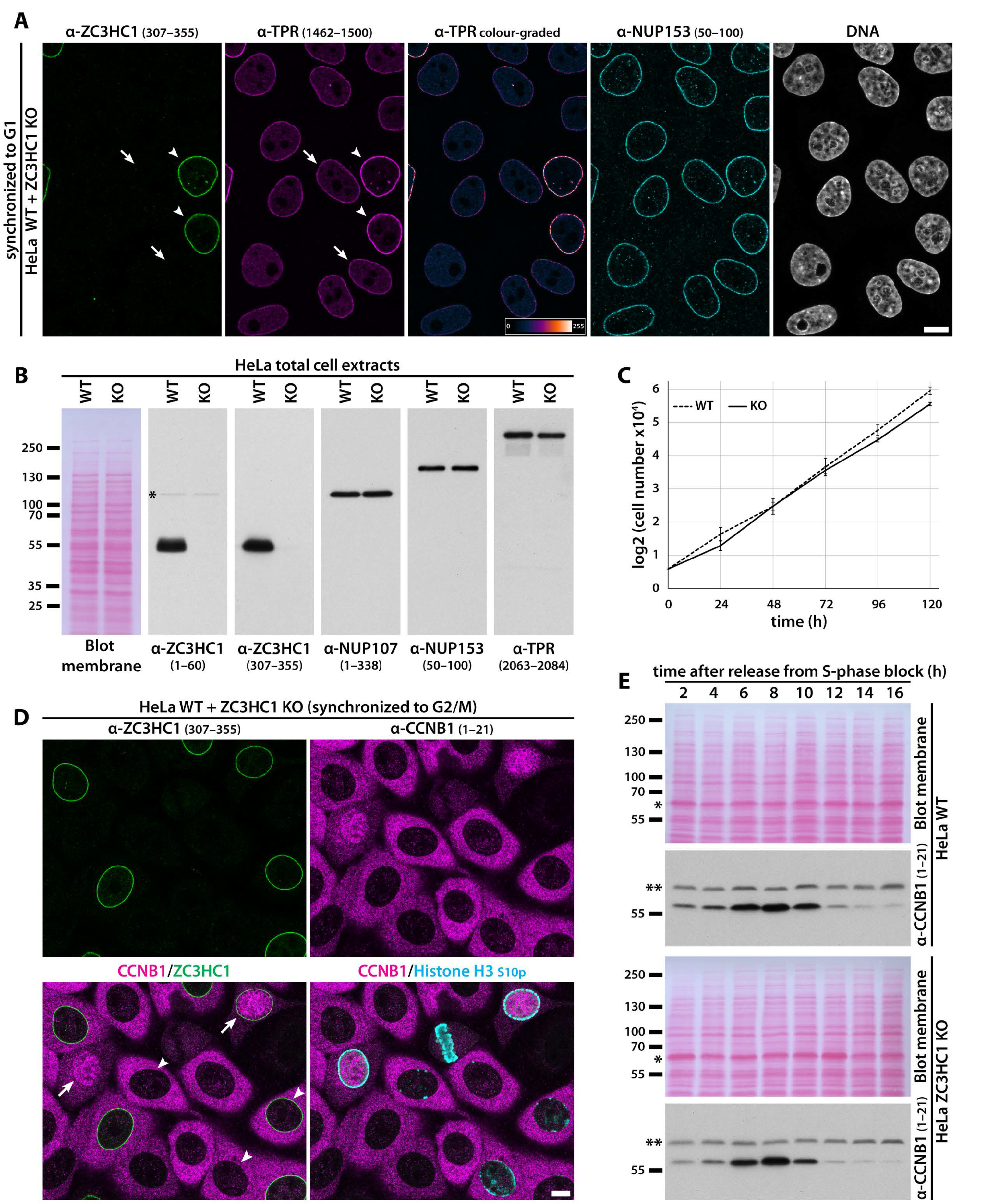
3.3. ZC3HC1 and TPR Engage in Physical Interactions and Depend on Each Other for Undiminished NE-Association of Both Proteins
3.4. ZC3HC1 Is Not Required for Cellular Housekeeping Activities in Human Tumour and Non-Tumour Cell Lines of Ectodermal, Mesodermal, and Endodermal Origin
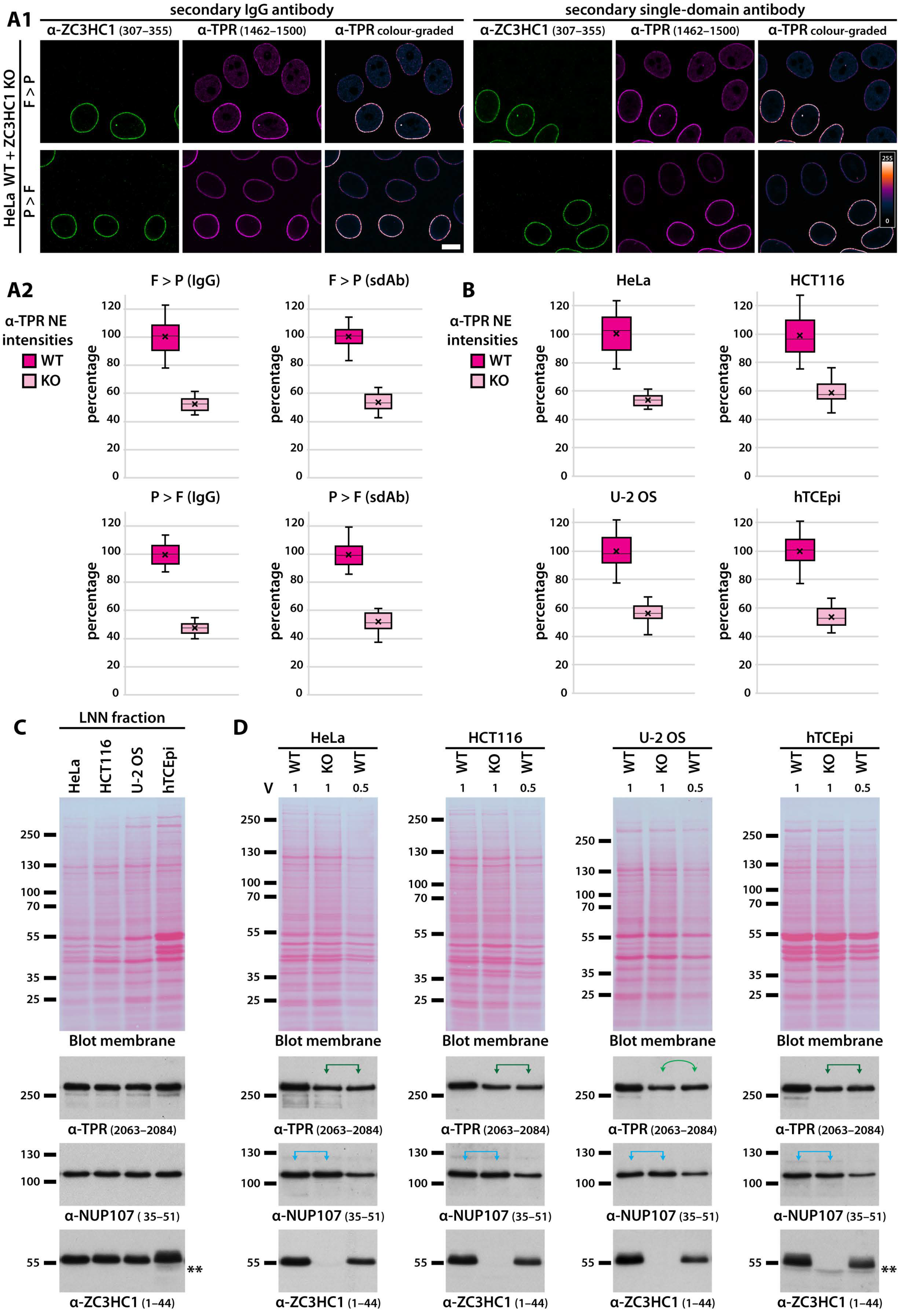
3.5. Presence of ZC3HC1 Is Required for NE-Localisation of about Half the There Positioned Amount of TPR in Several Human Cell Lines
4. Discussion
4.1. ZC3HC1 Qualifying as an NB Protein
4.2. ZC3HC1 as a Non-Essential Protein at the NB
4.3. ZC3HC1 and TPR as a Functional Ensemble at the NB?
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Franke, W.W.; Scheer, U. The Ultrastructure of the Nuclear Envelope of Amphibian Oocytes: A Reinvestigation. I. The Mature Oocyte. J. Ultrastruct. Res. 1970, 30, 288–316. [Google Scholar] [CrossRef]
- Maul, G.G. On the Octagonality of the Nuclear Pore Complex. J. Cell Biol. 1971, 51, 558–563. [Google Scholar] [CrossRef] [PubMed]
- Scheer, U. The Ultrastructure of the Nuclear Envelope of Amphibian Oocytes. IV. On the Chemical Nature of the Nuclear Pore Complex Material. Z. Zellforsch. Mikrosk. Anat. 1972, 127, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Maul, G.G. Fibrils Attached to the Nuclear Pore Prevent Egress of SV40 Particles from the Infected Nucleus. J. Cell Biol. 1976, 70, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Ris, H. Three-Dimensional Imaging of Cell Ultrastructure with High Resolution Low-Voltage SEM. Int. Phys. Conf. Ser. 1989, 98, 657–662. [Google Scholar]
- Ris, H. The Three-Dimensional Structure of the Nuclear Pore Complex as Seen by High Voltage Electron Microscopy and High Resolution Low Voltage Scanning Electron Microscopy. EMSA Bull. 1991, 21, 54–56. [Google Scholar]
- Jarnik, M.; Aebi, U. Toward a More Complete 3-D Structure of the Nuclear Pore Complex. J. Struct. Biol. 1991, 107, 291–308. [Google Scholar] [CrossRef]
- Kiseleva, E.; Goldberg, M.W.; Daneholt, B.; Allen, T.D. RNP Export Is Mediated by Structural Reorganization of the Nuclear Pore Basket. J. Mol. Biol. 1996, 260, 304–311. [Google Scholar] [CrossRef]
- Kiseleva, E.; Allen, T.D.; Rutherford, S.; Bucci, M.; Wente, S.R.; Goldberg, M.W. Yeast Nuclear Pore Complexes Have a Cytoplasmic Ring and Internal Filaments. J. Struct. Biol. 2004, 145, 272–288. [Google Scholar] [CrossRef]
- Beck, M.; Förster, F.; Ecke, M.; Plitzko, J.M.; Melchior, F.; Gerisch, G.; Baumeister, W.; Medalia, O. Nuclear Pore Complex Structure and Dynamics Revealed by Cryoelectron Tomography. Science 2004, 306, 1387–1390. [Google Scholar] [CrossRef]
- Krull, S.; Dörries, J.; Boysen, B.; Reidenbach, S.; Magnius, L.; Norder, H.; Thyberg, J.; Cordes, V.C. Protein TPR Is Required for Establishing Nuclear Pore-Associated Zones of Heterochromatin Exclusion. EMBO J. 2010, 29, 1659–1673. [Google Scholar] [CrossRef]
- Niepel, M.; Molloy, K.R.; Williams, R.; Farr, J.C.; Meinema, A.C.; Vecchietti, N.; Cristea, I.M.; Chait, B.T.; Rout, M.P.; Strambio-De-Castillia, C. The Nuclear Basket Proteins Mlp1p and Mlp2p Are Part of a Dynamic Interactome Including Esc1p and the Proteasome. Mol. Biol. Cell 2013, 24, 3920–3938. [Google Scholar] [CrossRef] [PubMed]
- Snow, C.J.; Paschal, B.M. Roles of the Nucleoporin TPR in Cancer and Aging. Adv. Exp. Med. Biol. 2014, 773, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Krull, S.; Thyberg, J.; Björkroth, B.; Rackwitz, H.-R.; Cordes, V.C. Nucleoporins as Components of the Nuclear Pore Complex Core Structure and TPR as the Architectural Element of the Nuclear Basket. Mol. Biol. Cell 2004, 15, 4261–4277. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.R.; Silver, P.A. Transcriptional Regulation at the Nuclear Pore Complex. Curr. Opin. Genet. Dev. 2007, 17, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Palancade, B.; Doye, V. Sumoylating and Desumoylating Enzymes at Nuclear Pores: Underpinning Their Unexpected Duties? Trends Cell Biol. 2008, 18, 174–183. [Google Scholar] [CrossRef]
- Dieppois, G.; Stutz, F. Connecting the Transcription Site to the Nuclear Pore: A Multi-Tether Process That Regulates Gene Expression. J. Cell Sci. 2010, 123, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Köhler, A.; Hurt, E. Gene Regulation by Nucleoporins and Links to Cancer. Mol. Cell 2010, 38, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Strambio-De-Castillia, C.; Niepel, M.; Rout, M.P. The Nuclear Pore Complex: Bridging Nuclear Transport and Gene Regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 490–501. [Google Scholar] [CrossRef] [PubMed]
- García-Oliver, E.; García-Molinero, V.; Rodríguez-Navarro, S. MRNA Export and Gene Expression: The SAGA-TREX-2 Connection. Biochim. Biophys. Acta 2012, 1819, 555–565. [Google Scholar] [CrossRef]
- Delaleau, M.; Borden, K.L.B. Multiple Export Mechanisms for MRNAs. Cells 2015, 4, 452–473. [Google Scholar] [CrossRef]
- Wilson, K.L. Nuclear Import Pathway Key to Rescuing Dominant Progerin Phenotypes. Sci. Signal. 2018, 11, eaat9448. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.J.; Cooper, C.S. The Human Tpr Gene Encodes a Protein of 2094 Amino Acids That Has Extensive Coiled-Coil Regions and an Acidic C-Terminal Domain. Oncogene 1992, 7, 2329–2333. [Google Scholar] [CrossRef]
- Byrd, D.A.; Sweet, D.J.; Panté, N.; Konstantinov, K.N.; Guan, T.; Saphire, A.C.; Mitchell, P.J.; Cooper, C.S.; Aebi, U.; Gerace, L. TPR, a Large Coiled Coil Protein Whose Amino Terminus Is Involved in Activation of Oncogenic Kinases, Is Localized to the Cytoplasmic Surface of the Nuclear Pore Complex. J. Cell Biol. 1994, 127, 1515–1526. [Google Scholar] [CrossRef] [PubMed]
- Cordes, V.C.; Reidenbach, S.; Rackwitz, H.R.; Franke, W.W. Identification of Protein P270/TPR as a Constitutive Component of the Nuclear Pore Complex-Attached Intranuclear Filaments. J. Cell Biol. 1997, 136, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Frosst, P.; Guan, T.; Subauste, C.; Hahn, K.; Gerace, L. TPR Is Localized within the Nuclear Basket of the Pore Complex and Has a Role in Nuclear Protein Export. J. Cell Biol. 2002, 156, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Strambio-de-Castillia, C.; Blobel, G.; Rout, M.P. Proteins Connecting the Nuclear Pore Complex with the Nuclear Interior. J. Cell Biol. 1999, 144, 839–855. [Google Scholar] [CrossRef]
- Kosova, B.; Panté, N.; Rollenhagen, C.; Podtelejnikov, A.; Mann, M.; Aebi, U.; Hurt, E. Mlp2p, a Component of Nuclear Pore Attached Intranuclear Filaments, Associates with Nic96p. J. Biol. Chem. 2000, 275, 343–350. [Google Scholar] [CrossRef]
- Funasaka, T.; Tsuka, E.; Wong, R.W. Regulation of Autophagy by Nucleoporin TPR. Sci. Rep. 2012, 2, 878. [Google Scholar] [CrossRef] [PubMed]
- Duheron, V.; Chatel, G.; Sauder, U.; Oliveri, V.; Fahrenkrog, B. Structural Characterization of Altered Nucleoporin NUP153 Expression in Human Cells by Thin-Section Electron Microscopy. Nucleus 2014, 5, 601–612. [Google Scholar] [CrossRef]
- Scott, R.J.; Lusk, C.P.; Dilworth, D.J.; Aitchison, J.D.; Wozniak, R.W. Interactions between Mad1p and the Nuclear Transport Machinery in the Yeast Saccharomyces Cerevisiae. Mol. Biol. Cell 2005, 16, 4362–4374. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Sterling, H.; Burlingame, A.; McCormick, F. TPR Directly Binds to MAD1 and MAD2 and Is Important for the MAD1-MAD2-Mediated Mitotic Spindle Checkpoint. Genes Dev. 2008, 22, 2926–2931. [Google Scholar] [CrossRef]
- Ding, D.; Muthuswamy, S.; Meier, I. Functional Interaction between the Arabidopsis Orthologs of Spindle Assembly Checkpoint Proteins MAD1 and MAD2 and the Nucleoporin NUA. Plant Mol. Biol. 2012, 79, 203–216. [Google Scholar] [CrossRef]
- Palancade, B.; Zuccolo, M.; Loeillet, S.; Nicolas, A.; Doye, V. Pml39, a Novel Protein of the Nuclear Periphery Required for Nuclear Retention of Improper Messenger Ribonucleoparticles. Mol. Biol. Cell 2005, 16, 5258–5268. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wu, C.-Y.; Blobel, G. Mlp-Dependent Anchorage and Stabilization of a Desumoylating Enzyme Is Required to Prevent Clonal Lethality. J. Cell Biol. 2004, 167, 605–611. [Google Scholar] [CrossRef]
- Schweizer, N.; Ferrás, C.; Kern, D.M.; Logarinho, E.; Cheeseman, I.M.; Maiato, H. Spindle Assembly Checkpoint Robustness Requires TPR-Mediated Regulation of MAD1/MAD2 Proteostasis. J. Cell Biol. 2013, 203, 883–893. [Google Scholar] [CrossRef]
- Umlauf, D.; Bonnet, J.; Waharte, F.; Fournier, M.; Stierle, M.; Fischer, B.; Brino, L.; Devys, D.; Tora, L. The Human TREX-2 Complex Is Stably Associated with the Nuclear Pore Basket. J. Cell Sci. 2013, 126, 2656–2667. [Google Scholar] [CrossRef] [PubMed]
- Aksenova, V.; Smith, A.; Lee, H.; Bhat, P.; Esnault, C.; Chen, S.; Iben, J.; Kaufhold, R.; Yau, K.C.; Echeverria, C.; et al. Nucleoporin TPR Is an Integral Component of the TREX-2 MRNA Export Pathway. Nat. Commun. 2020, 11, 4577. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Guo, P.; Takeda, K.; Fu, Q.; Fang, H.; Frucht, D.M. ERK1/2 Inactivation Promotes a Rapid Redistribution of COP1 and Degradation of COP1 Substrates. Proc. Natl. Acad. Sci. USA 2020, 117, 4078–4087. [Google Scholar] [CrossRef]
- HUGO Gene Nomenclature Committee. Available online: https://www.genenames.org/data/gene-symbol-report/#!/hgnc_id/HGNC:29913 (accessed on 25 July 2021).
- Zhang, Q.H.; Ye, M.; Wu, X.Y.; Ren, S.X.; Zhao, M.; Zhao, C.J.; Fu, G.; Shen, Y.; Fan, H.Y.; Lu, G.; et al. Cloning and Functional Analysis of CDNAs with Open Reading Frames for 300 Previously Undefined Genes Expressed in CD34+ Hematopoietic Stem/Progenitor Cells. Genome Res. 2000, 10, 1546–1560. [Google Scholar] [CrossRef]
- Higashi, K.; Takasawa, R.; Yoshimori, A.; Goh, T.; Tanuma, S.; Kuchitsu, K. Identification of a Novel Gene Family, Paralogs of Inhibitor of Apoptosis Proteins Present in Plants, Fungi, and Animals. Apoptosis 2005, 10, 471–480. [Google Scholar] [CrossRef]
- Ouyang, T.; Bai, R.-Y.; Bassermann, F.; von Klitzing, C.; Klumpen, S.; Miething, C.; Morris, S.W.; Peschel, C.; Duyster, J. Identification and Characterization of a Nuclear Interacting Partner of Anaplastic Lymphoma Kinase (NIPA). J. Biol. Chem. 2003, 278, 30028–30036. [Google Scholar] [CrossRef]
- Bassermann, F.; von Klitzing, C.; Münch, S.; Bai, R.-Y.; Kawaguchi, H.; Morris, S.W.; Peschel, C.; Duyster, J. NIPA Defines an SCF-Type Mammalian E3 Ligase That Regulates Mitotic Entry. Cell 2005, 122, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Bassermann, F.; Peschel, C.; Duyster, J. Mitotic Entry: A Matter of Oscillating Destruction. Cell Cycle 2005, 4, 1515–1517. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bassermann, F.; von Klitzing, C.; Illert, A.L.; Münch, S.; Morris, S.W.; Pagano, M.; Peschel, C.; Duyster, J. Multisite Phosphorylation of Nuclear Interaction Partner of ALK (NIPA) at G2/M Involves Cyclin B1/Cdk1. J. Biol. Chem. 2007, 282, 15965–15972. [Google Scholar] [CrossRef] [PubMed]
- von Klitzing, C.; Huss, R.; Illert, A.L.; Fröschl, A.; Wötzel, S.; Peschel, C.; Bassermann, F.; Duyster, J. APC/C(Cdh1)-Mediated Degradation of the F-Box Protein NIPA Is Regulated by Its Association with SKP1. PLoS ONE 2011, 6, e28998. [Google Scholar] [CrossRef]
- Illert, A.L.; Zech, M.; Moll, C.; Albers, C.; Kreutmair, S.; Peschel, C.; Bassermann, F.; Duyster, J. Extracellular Signal-Regulated Kinase 2 (ERK2) Mediates Phosphorylation and Inactivation of Nuclear Interaction Partner of Anaplastic Lymphoma Kinase (NIPA) at G2/M. J. Biol. Chem. 2012, 287, 37997–38005. [Google Scholar] [CrossRef]
- Lu, Z.H.; Xu, H.; Leno, G.H. DNA Replication in Quiescent Cell Nuclei: Regulation by the Nuclear Envelope and Chromatin Structure. Mol. Biol. Cell 1999, 10, 4091–4106. [Google Scholar] [CrossRef]
- Hase, M.E.; Cordes, V.C. Direct Interaction with NUP153 Mediates Binding of TPR to the Periphery of the Nuclear Pore Complex. Mol. Biol. Cell 2003, 14, 1923–1940. [Google Scholar] [CrossRef]
- Ma, H.T.; Poon, R.Y.C. Synchronization of HeLa Cells. Methods Mol. Biol. 2011, 761, 151–161. [Google Scholar] [CrossRef]
- Cordes, V.C.; Reidenbach, S.; Franke, W.W. High Content of a Nuclear Pore Complex Protein in Cytoplasmic Annulate Lamellae of Xenopus Oocytes. Eur. J. Cell Biol. 1995, 68, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Krohne, G.; Franke, W.W. Proteins of Pore Complex—Lamina Structures from Nuclei and Nuclear Membranes. Methods Enzymol. 1983, 96, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, N.V.; Sandblad, L.; Hase, M.E.; Hunziker, A.; Hergt, M.; Cordes, V.C. The Evolutionarily Conserved Single-Copy Gene for Murine TPR Encodes One Prevalent Isoform in Somatic Cells and Lacks Paralogs in Higher Eukaryotes. Chromosoma 2002, 111, 236–255. [Google Scholar] [CrossRef]
- Cronshaw, J.M.; Krutchinsky, A.N.; Zhang, W.; Chait, B.T.; Matunis, M.J. Proteomic Analysis of the Mammalian Nuclear Pore Complex. J. Cell Biol. 2002, 158, 915–927. [Google Scholar] [CrossRef]
- Blow, J.J.; Laskey, R.A. Initiation of DNA Replication in Nuclei and Purified DNA by a Cell-Free Extract of Xenopus Eggs. Cell 1986, 47, 577–587. [Google Scholar] [CrossRef]
- Thomas, J.O.; Kornberg, R.D. An Octamer of Histones in Chromatin and Free in Solution. Proc. Natl. Acad. Sci. USA 1975, 72, 2626–2630. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Lin, C.-Y.; Gootenberg, J.S.; Konermann, S.; Trevino, A.E.; Scott, D.A.; Inoue, A.; Matoba, S.; Zhang, Y.; et al. Double Nicking by RNA-Guided CRISPR Cas9 for Enhanced Genome Editing Specificity. Cell 2013, 154, 1380–1389. [Google Scholar] [CrossRef]
- Optimized CRISPR Design. Available online: http://crispr.mit.edu/ (accessed on 20 July 2016).
- Hsu, P.D.; Scott, D.A.; Weinstein, J.A.; Ran, F.A.; Konermann, S.; Agarwala, V.; Li, Y.; Fine, E.J.; Wu, X.; Shalem, O.; et al. DNA Targeting Specificity of RNA-Guided Cas9 Nucleases. Nat. Biotechnol. 2013, 31, 827–832. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Wright, J.; Agarwala, V.; Scott, D.A.; Zhang, F. Genome Engineering Using the CRISPR-Cas9 System. Nat. Protoc. 2013, 8, 2281–2308. [Google Scholar] [CrossRef]
- Addgene. Available online: http://n2t.net/addgene:62987 (accessed on 25 July 2021).
- Dumont, J.N. Oogenesis in Xenopus Laevis (Daudin). I. Stages of Oocyte Development in Laboratory Maintained Animals. J. Morphol. 1972, 136, 153–179. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.W.; Allen, T.D. High Resolution Scanning Electron Microscopy of the Nuclear Envelope: Demonstration of a New, Regular, Fibrous Lattice Attached to the Baskets of the Nucleoplasmic Face of the Nuclear Pores. J. Cell Biol. 1992, 119, 1429–1440. [Google Scholar] [CrossRef] [PubMed]
- Cordes, V.C.; Reidenbach, S.; Köhler, A.; Stuurman, N.; van Driel, R.; Franke, W.W. Intranuclear Filaments Containing a Nuclear Pore Complex Protein. J. Cell Biol. 1993, 123, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Ris, H.; Malecki, M. High-Resolution Field Emission Scanning Electron Microscope Imaging of Internal Cell Structures after Epon Extraction from Sections: A New Approach to Correlative Ultrastructural and Immunocytochemical Studies. J. Struct. Biol. 1993, 111, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Arlucea, J.; Andrade, R.; Alonso, R.; Aréchaga, J. The Nuclear Basket of the Nuclear Pore Complex Is Part of a Higher-Order Filamentous Network That Is Related to Chromatin. J. Struct. Biol. 1998, 124, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Kiseleva, E.; Drummond, S.P.; Goldberg, M.W.; Rutherford, S.A.; Allen, T.D.; Wilson, K.L. Actin- and Protein-4.1-Containing Filaments Link Nuclear Pore Complexes to Subnuclear Organelles in Xenopus Oocyte Nuclei. J. Cell Sci. 2004, 117, 2481–2490. [Google Scholar] [CrossRef]
- Walther, P. High-Resolution Cryo-SEM Allows Direct Identification of F-Actin at the Inner Nuclear Membrane of Xenopus Oocytes by Virtue of Its Structural Features. J. Microsc. 2008, 232, 379–385. [Google Scholar] [CrossRef]
- Starr, C.M.; D’Onofrio, M.; Park, M.K.; Hanover, J.A. Primary Sequence and Heterologous Expression of Nuclear Pore Glycoprotein P62. J. Cell Biol. 1990, 110, 1861–1871. [Google Scholar] [CrossRef]
- Cordes, V.C.; Waizenegger, I.; Krohne, G. Nuclear Pore Complex Glycoprotein P62 of Xenopus Laevis and Mouse: CDNA Cloning and Identification of Its Glycosylated Region. Eur. J. Cell Biol. 1991, 55, 31–47. [Google Scholar]
- Carmo-Fonseca, M.; Kern, H.; Hurt, E.C. Human Nucleoporin P62 and the Essential Yeast Nuclear Pore Protein NSP1 Show Sequence Homology and a Similar Domain Organization. Eur. J. Cell Biol. 1991, 55, 17–30. [Google Scholar]
- Grote, M.; Kubitscheck, U.; Reichelt, R.; Peters, R. Mapping of Nucleoporins to the Center of the Nuclear Pore Complex by Post-Embedding Immunogold Electron Microscopy. J. Cell Sci. 1995, 108, 2963–2972. [Google Scholar] [CrossRef]
- Sukegawa, J.; Blobel, G. A Nuclear Pore Complex Protein That Contains Zinc Finger Motifs, Binds DNA, and Faces the Nucleoplasm. Cell 1993, 72, 29–38. [Google Scholar] [CrossRef]
- Kessel, R.G. The Annulate Lamellae—From Obscurity to Spotlight. Electron Microsc. Rev. 1989, 2, 257–348. [Google Scholar] [CrossRef]
- Kessel, R.G. Annulate Lamellae: A Last Frontier in Cellular Organelles. Int. Rev. Cytol. 1992, 133, 43–120. [Google Scholar] [CrossRef]
- Imoh, H.; Okamoto, M.; Eguchi, G. Accumulation of Annulate Lamellae in the Subcortical Layer during Progesterone-Induced Oocyte Maturation in Xenopus Laevis. Dev. Growth Differ. 1983, 25, 1–10. [Google Scholar] [CrossRef]
- Cordes, V.C.; Gajewski, A.; Stumpp, S.; Krohne, G. Immunocytochemistry of Annulate Lamellae: Potential Cell Biological Markers for Studies of Cell Differentiation and Pathology. Differentiation 1995, 58, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Cordes, V.C.; Rackwitz, H.R.; Reidenbach, S. Mediators of Nuclear Protein Import Target Karyophilic Proteins to Pore Complexes of Cytoplasmic Annulate Lamellae. Exp. Cell Res. 1997, 237, 419–433. [Google Scholar] [CrossRef] [PubMed]
- Dabauvalle, M.C.; Loos, K.; Merkert, H.; Scheer, U. Spontaneous Assembly of Pore Complex-Containing Membranes (“annulate Lamellae”) in Xenopus Egg Extract in the Absence of Chromatin. J. Cell Biol. 1991, 112, 1073–1082. [Google Scholar] [CrossRef]
- Kleinschmidt, J.A.; Scheer, U.; Dabauvalle, M.C.; Bustin, M.; Franke, W.W. High Mobility Group Proteins of Amphibian Oocytes: A Large Storage Pool of a Soluble High Mobility Group-1-like Protein and Involvement in Transcriptional Events. J. Cell Biol. 1983, 97, 838–848. [Google Scholar] [CrossRef]
- Miller, L.; Daniel, J.C. Comparison of in Vivo and in Vitro Ribosomal RNA Synthesis in Nucleolar Mutants of Xenopus Laevis. In Vitro 1977, 13, 557–563. [Google Scholar] [CrossRef]
- Ellison, T.R.; Mathisen, P.M.; Miller, L. Developmental Changes in Keratin Patterns during Epidermal Maturation. Dev. Biol. 1985, 112, 329–337. [Google Scholar] [CrossRef]
- Bastos, R.; Lin, A.; Enarson, M.; Burke, B. Targeting and Function in MRNA Export of Nuclear Pore Complex Protein NUP153. J. Cell Biol. 1996, 134, 1141–1156. [Google Scholar] [CrossRef]
- Enarson, P.; Enarson, M.; Bastos, R.; Burke, B. Amino-Terminal Sequences That Direct Nucleoporin NUP153 to the Inner Surface of the Nuclear Envelope. Chromosoma 1998, 107, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Vasu, S.; Shah, S.; Orjalo, A.; Park, M.; Fischer, W.H.; Forbes, D.J. Novel Vertebrate Nucleoporins NUP133 and NUP160 Play a Role in MRNA Export. J. Cell Biol. 2001, 155, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, B.; Lorenz, M.; Moreno-Andrés, D.; Bodenhöfer, M.; De Magistris, P.; Astrinidis, S.A.; Schooley, A.; Flötenmeyer, M.; Leptihn, S.; Antonin, W. NUP153 Recruits the NUP107-160 Complex to the Inner Nuclear Membrane for Interphasic Nuclear Pore Complex Assembly. Dev. Cell 2015, 33, 717–728. [Google Scholar] [CrossRef] [PubMed]
- Walther, T.C.; Fornerod, M.; Pickersgill, H.; Goldberg, M.; Allen, T.D.; Mattaj, I.W. The Nucleoporin NUP153 Is Required for Nuclear Pore Basket Formation, Nuclear Pore Complex Anchoring and Import of a Subset of Nuclear Proteins. EMBO J. 2001, 20, 5703–5714. [Google Scholar] [CrossRef]
- Scheer, U.; Dabauvalle, M.C.; Merkert, H.; Benevente, R. The Nuclear Envelope and the Organization of the Pore Complexes. Cell Biol. Int. Rep. 1988, 12, 669–689. [Google Scholar] [CrossRef][Green Version]
- Maclean, N.; Hilder, V.A.; Baynes, Y.A. RNA Synthesis in Xenopus Erythrocytes. Cell Differ. 1973, 2, 261–269. [Google Scholar] [CrossRef]
- Krohne, G.; Dabauvalle, M.C.; Franke, W.W. Cell Type-Specific Differences in Protein Composition of Nuclear Pore Complex-Lamina Structures in Oocytes and Erythrocytes of Xenopus Laevis. J. Mol. Biol. 1981, 151, 121–141. [Google Scholar] [CrossRef]
- Gambino, J.; Weatherbee, J.A.; Gavin, R.H.; Eckhardt, R.A. Studies on the Cytoskeletal and Nuclear Architecture of Xenopus Erythrocytes. J. Cell Sci. 1984, 72, 275–294. [Google Scholar] [CrossRef]
- Maul, G.G.; Deaven, L.L.; Freed, J.J.; Campbell, G.L.; Beçak, W. Investigation of the Determinants of Nuclear Pore Number. Cytogenet. Cell Genet. 1980, 26, 175–190. [Google Scholar] [CrossRef]
- Savas, J.N.; Toyama, B.H.; Xu, T.; Yates, J.R.; Hetzer, M.W. Extremely Long-Lived Nuclear Pore Proteins in the Rat Brain. Science 2012, 335, 942. [Google Scholar] [CrossRef] [PubMed]
- Ori, A.; Banterle, N.; Iskar, M.; Andrés-Pons, A.; Escher, C.; Khanh Bui, H.; Sparks, L.; Solis-Mezarino, V.; Rinner, O.; Bork, P.; et al. Cell Type-Specific Nuclear Pores: A Case in Point for Context-Dependent Stoichiometry of Molecular Machines. Mol. Syst. Biol. 2013, 9, 648. [Google Scholar] [CrossRef]
- Toyama, B.H.; Savas, J.N.; Park, S.K.; Harris, M.S.; Ingolia, N.T.; Yates, J.R.; Hetzer, M.W. Identification of Long-Lived Proteins Reveals Exceptional Stability of Essential Cellular Structures. Cell 2013, 154, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Mathieson, T.; Franken, H.; Kosinski, J.; Kurzawa, N.; Zinn, N.; Sweetman, G.; Poeckel, D.; Ratnu, V.S.; Schramm, M.; Becher, I.; et al. Systematic Analysis of Protein Turnover in Primary Cells. Nat. Commun. 2018, 9, 689. [Google Scholar] [CrossRef] [PubMed]
- Cordes, V.C.; Reidenbach, S.; Franke, W.W. Cytoplasmic Annulate Lamellae in Cultured Cells: Composition, Distribution, and Mitotic Behavior. Cell Tissue Res. 1996, 284, 177–191. [Google Scholar] [CrossRef]
- Finlay, D.R.; Forbes, D.J. Reconstitution of Biochemically Altered Nuclear Pores: Transport Can Be Eliminated and Restored. Cell 1990, 60, 17–29. [Google Scholar] [CrossRef]
- Bodoor, K.; Shaikh, S.; Salina, D.; Raharjo, W.H.; Bastos, R.; Lohka, M.; Burke, B. Sequential Recruitment of NPC Proteins to the Nuclear Periphery at the End of Mitosis. J. Cell Sci. 1999, 112, 2253–2264. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, T.; Koujin, T.; Hayakawa, T.; Kaneda, T.; Tsutsumi, C.; Imamoto, N.; Akazawa, C.; Sukegawa, J.; Yoneda, Y.; Hiraoka, Y. Live Fluorescence Imaging Reveals Early Recruitment of Emerin, LBR, RanBP2, and NUP153 to Reforming Functional Nuclear Envelopes. J. Cell Sci. 2000, 113, 779–794. [Google Scholar] [CrossRef]
- Gengenbacher, A.; Müller-Rudorf, A.; Poggio, T.; Gräßel, L.; Dumit, V.I.; Kreutmair, S.; Lippert, L.J.; Duyster, J.; Illert, A.L. Proteomic Phosphosite Analysis Identified Crucial NPM-ALK-Mediated NIPA Serine and Threonine Residues. Int. J. Mol. Sci. 2019, 20, 4060. [Google Scholar] [CrossRef]
- Illert, A.L.; Kawaguchi, H.; Antinozzi, C.; Bassermann, F.; Quintanilla-Martinez, L.; von Klitzing, C.; Hiwatari, M.; Peschel, C.; de Rooij, D.G.; Morris, S.W.; et al. Targeted Inactivation of Nuclear Interaction Partner of ALK Disrupts Meiotic Prophase. Development 2012, 139, 2523–2534. [Google Scholar] [CrossRef]
- International Mouse Phenotyping Consortium. Available online: https://www.mousephenotype.org/data/genes/MGI:1916023 (accessed on 25 July 2021).
- Liu, Y.-Q.; Wang, X.-L.; Cheng, X.; Lu, Y.; Wang, G.; Li, X.-C.; Zhang, J.; Wen, Z.-S.; Huang, Z.-L.; Gao, Q.-L.; et al. SKP1 in Lung Cancer: Clinical Significance and Therapeutic Efficacy of Its Small Molecule Inhibitors. Oncotarget 2015, 6, 34953–34967. [Google Scholar] [CrossRef]
- Jones, P.D.; Kaiser, M.A.; Ghaderi Najafabadi, M.; McVey, D.G.; Beveridge, A.J.; Schofield, C.L.; Samani, N.J.; Webb, T.R. The Coronary Artery Disease-Associated Coding Variant in Zinc Finger C3HC-Type Containing 1 (ZC3HC1) Affects Cell Cycle Regulation. J. Biol. Chem. 2016, 291, 16318–16327. [Google Scholar] [CrossRef] [PubMed]
- Linseman, T.; Soubeyrand, S.; Martinuk, A.; Nikpay, M.; Lau, P.; McPherson, R. Functional Validation of a Common Nonsynonymous Coding Variant in ZC3HC1 Associated with Protection from Coronary Artery Disease. Circ. Cardiovasc. Genet. 2017, 10, e001498. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef]
- Mali, P.; Aach, J.; Stranges, P.B.; Esvelt, K.M.; Moosburner, M.; Kosuri, S.; Yang, L.; Church, G.M. CAS9 Transcriptional Activators for Target Specificity Screening and Paired Nickases for Cooperative Genome Engineering. Nat. Biotechnol. 2013, 31, 833–838. [Google Scholar] [CrossRef]
- Gey, G.O.; Coffman, W.D.; Kubicek, M. Tissue Culture Studies of the Proliferative Capacity of Cervical Carcinoma and Normal Epithelium. Cancer Res. 1952, 12, 264–265. [Google Scholar]
- Jones, H.W.; McKusick, V.A.; Harper, P.S.; Wuu, K.D. George Otto Gey (1899–1970). The HeLa Cell and a Reappraisal of Its Origin. Obstet. Gynecol. 1971, 38, 945–949. [Google Scholar]
- Berman, J.J. Tumor Classification: Molecular Analysis Meets Aristotle. BMC Cancer 2004, 4, 10. [Google Scholar] [CrossRef]
- Pontén, J.; Saksela, E. Two Established in Vitro Cell Lines from Human Mesenchymal Tumours. Int. J. Cancer 1967, 2, 434–447. [Google Scholar] [CrossRef] [PubMed]
- American Type Culture Collection (ATCC). Available online: https://www.atcc.org/products/htb-96 (accessed on 25 July 2021).
- Brattain, M.G.; Fine, W.D.; Khaled, F.M.; Thompson, J.; Brattain, D.E. Heterogeneity of Malignant Cells from a Human Colonic Carcinoma. Cancer Res. 1981, 41, 1751–1756. [Google Scholar] [PubMed]
- Robertson, D.M.; Li, L.; Fisher, S.; Pearce, V.P.; Shay, J.W.; Wright, W.E.; Cavanagh, H.D.; Jester, J.V. Characterization of Growth and Differentiation in a Telomerase-Immortalized Human Corneal Epithelial Cell Line. Investig. Ophthalmol. Vis. Sci. 2005, 46, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Landry, J.J.M.; Pyl, P.T.; Rausch, T.; Zichner, T.; Tekkedil, M.M.; Stütz, A.M.; Jauch, A.; Aiyar, R.S.; Pau, G.; Delhomme, N.; et al. The Genomic and Transcriptomic Landscape of a HeLa Cell Line. G3 2013, 3, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Adey, A.; Burton, J.N.; Kitzman, J.O.; Hiatt, J.B.; Lewis, A.P.; Martin, B.K.; Qiu, R.; Lee, C.; Shendure, J. The Haplotype-Resolved Genome and Epigenome of the Aneuploid HeLa Cancer Cell Line. Nature 2013, 500, 207–211. [Google Scholar] [CrossRef]
- Rout, M.P.; Aitchison, J.D.; Suprapto, A.; Hjertaas, K.; Zhao, Y.; Chait, B.T. The Yeast Nuclear Pore Complex: Composition, Architecture, and Transport Mechanism. J. Cell Biol. 2000, 148, 635–651. [Google Scholar] [CrossRef]
- Yoon, J.H. Schizosaccharomyces Pombe Rsm1 Genetically Interacts with Spmex67, Which Is Involved in MRNA Export. J. Microbiol. 2004, 42, 32–36. [Google Scholar]
- Hart, T.; Chandrashekhar, M.; Aregger, M.; Steinhart, Z.; Brown, K.R.; MacLeod, G.; Mis, M.; Zimmermann, M.; Fradet-Turcotte, A.; Sun, S.; et al. High-Resolution CRISPR Screens Reveal Fitness Genes and Genotype-Specific Cancer Liabilities. Cell 2015, 163, 1515–1526. [Google Scholar] [CrossRef]
- Rual, J.-F.; Ceron, J.; Koreth, J.; Hao, T.; Nicot, A.; Hirozane-Kishikawa, T.; Vandenhaute, J.; Orkin, S.H.; Hill, D.E.; van den Heuvel, S.; et al. Toward Improving Caenorhabditis Elegans Phenome Mapping with an ORFeome-Based RNAi Library. Genome Res. 2004, 14, 2162–2168. [Google Scholar] [CrossRef]
- Sönnichsen, B.; Koski, L.B.; Walsh, A.; Marschall, P.; Neumann, B.; Brehm, M.; Alleaume, A.-M.; Artelt, J.; Bettencourt, P.; Cassin, E.; et al. Full-Genome RNAi Profiling of Early Embryogenesis in Caenorhabditis Elegans. Nature 2005, 434, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Kreutmair, S.; Erlacher, M.; Andrieux, G.; Istvanffy, R.; Mueller-Rudorf, A.; Zwick, M.; Rückert, T.; Pantic, M.; Poggio, T.; Shoumariyeh, K.; et al. Loss of the Fanconi Anemia–Associated Protein NIPA Causes Bone Marrow Failure. J. Clin. Investig. 2020, 130, 2827–2844. [Google Scholar] [CrossRef] [PubMed]
- Mouse Genome Database (MGD) at the Mouse Genome Informatics Website, The Jackson Laboratory. Available online: http://www.informatics.jax.org/marker/MGI:1916023 (accessed on 25 July 2021).
- Boumendil, C.; Hari, P.; Olsen, K.C.F.; Acosta, J.C.; Bickmore, W.A. Nuclear Pore Density Controls Heterochromatin Reorganization during Senescence. Genes Dev. 2019, 33, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Hase, M.E.; Kuznetsov, N.V.; Cordes, V.C. Amino Acid Substitutions of Coiled-Coil Protein TPR Abrogate Anchorage to the Nuclear Pore Complex but Not Parallel, in-Register Homodimerization. Mol. Biol. Cell 2001, 12, 2433–2452. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.; Görlich, D. A Saturated FG-Repeat Hydrogel Can Reproduce the Permeability Properties of Nuclear Pore Complexes. Cell 2007, 130, 512–523. [Google Scholar] [CrossRef]
- Schmidt, H.B.; Görlich, D. Transport Selectivity of Nuclear Pores, Phase Separation, and Membraneless Organelles. Trends Biochem. Sci. 2016, 41, 46–61. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gunkel, P.; Iino, H.; Krull, S.; Cordes, V.C. ZC3HC1 Is a Novel Inherent Component of the Nuclear Basket, Resident in a State of Reciprocal Dependence with TPR. Cells 2021, 10, 1937. https://doi.org/10.3390/cells10081937
Gunkel P, Iino H, Krull S, Cordes VC. ZC3HC1 Is a Novel Inherent Component of the Nuclear Basket, Resident in a State of Reciprocal Dependence with TPR. Cells. 2021; 10(8):1937. https://doi.org/10.3390/cells10081937
Chicago/Turabian StyleGunkel, Philip, Haruki Iino, Sandra Krull, and Volker C. Cordes. 2021. "ZC3HC1 Is a Novel Inherent Component of the Nuclear Basket, Resident in a State of Reciprocal Dependence with TPR" Cells 10, no. 8: 1937. https://doi.org/10.3390/cells10081937
APA StyleGunkel, P., Iino, H., Krull, S., & Cordes, V. C. (2021). ZC3HC1 Is a Novel Inherent Component of the Nuclear Basket, Resident in a State of Reciprocal Dependence with TPR. Cells, 10(8), 1937. https://doi.org/10.3390/cells10081937