
Altered Cytokine Response of Human Brain Endothelial Cells after Stimulation with Malaria Patient Plasma

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Blood Plasma of Malaria Patients and Healthy Control Individuals
2.2. HBEC-5i Brain Endothelial Cell Line
2.3. Stimulation Assay of ECs with Plasma of Malaria Patients and Healthy Control Individuals
2.4. LEGENDplexTM Assay
2.5. ANGPTL4 and TNF-α ELISA
2.6. RNA Isolation
3. Results
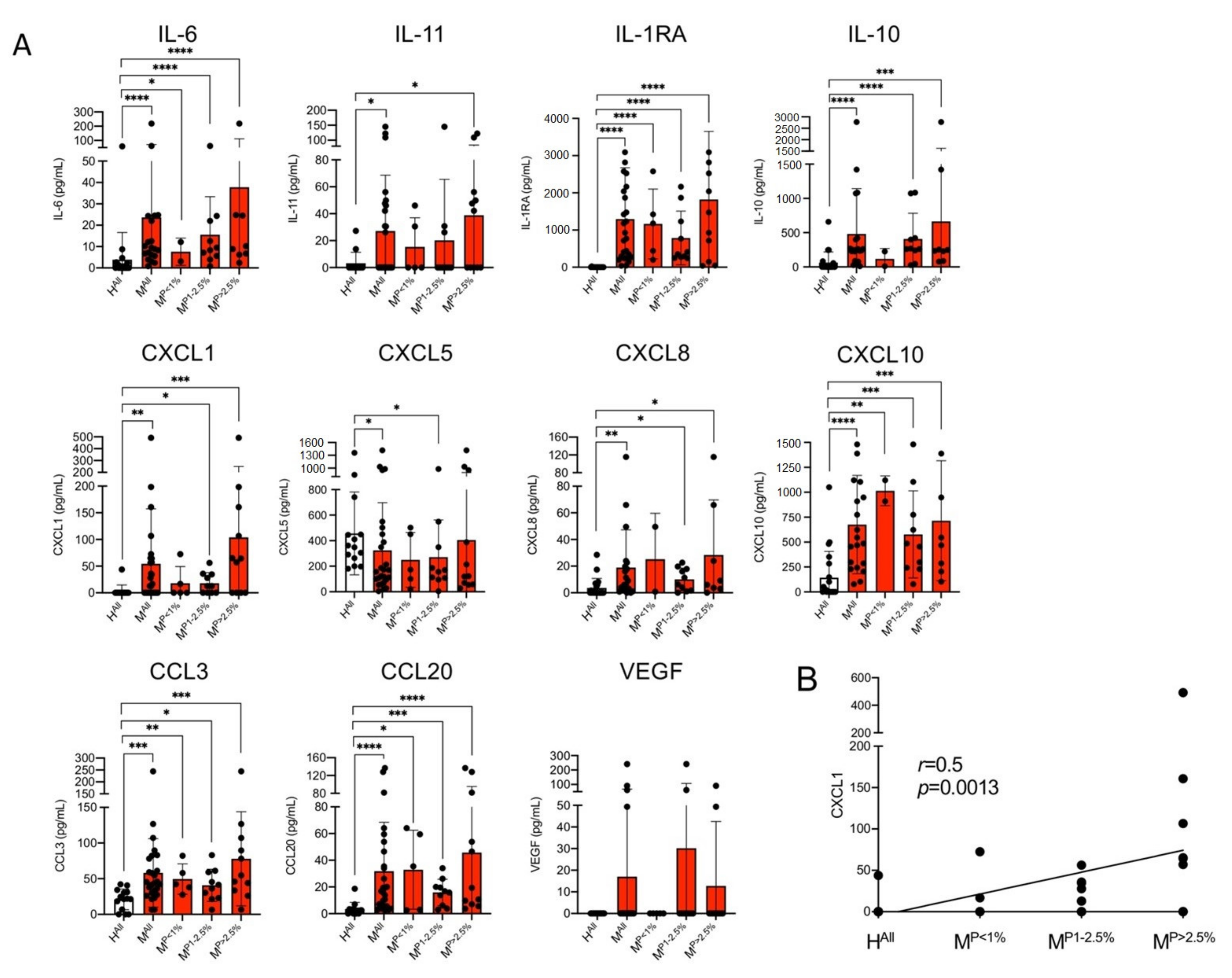
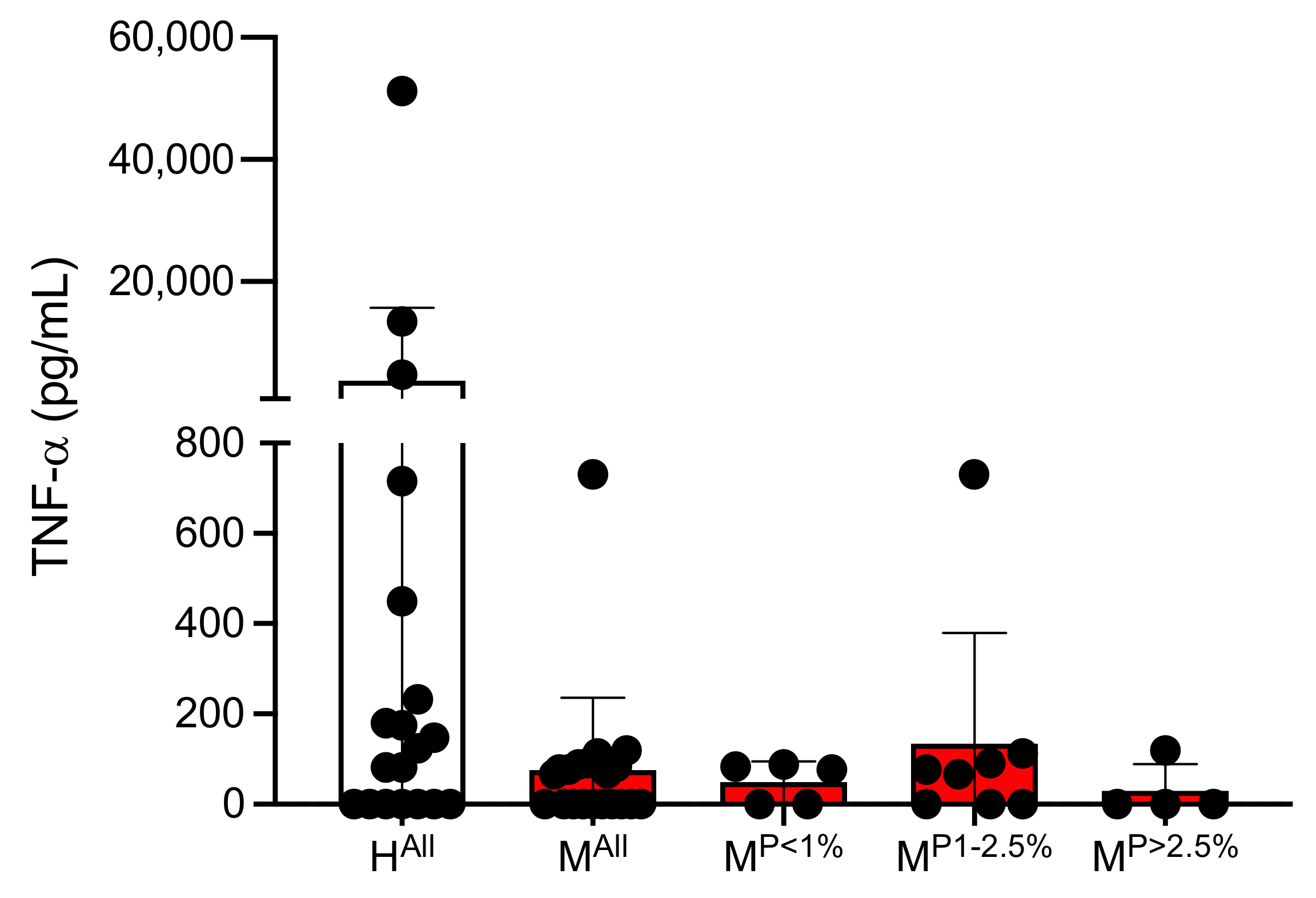
3.1. Determination of Concentrations of Different Cytokines in Plasmas of Malaria Patients and Healthy Controls
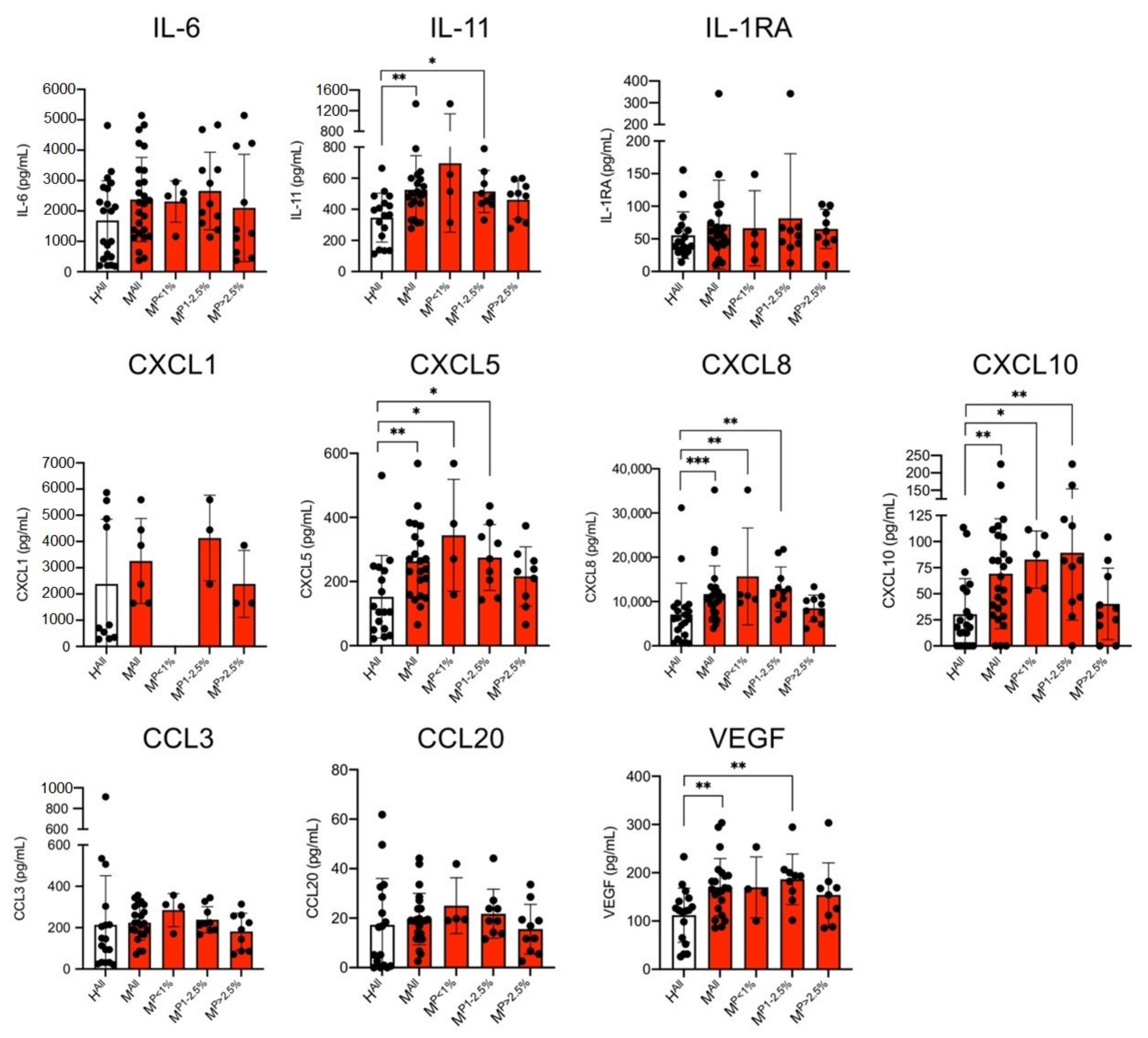
3.2. Determination of Concentrations of Various Cytokines in the Culture Supernatant of ECs Stimulated with Plasma from Malaria Patients and Healthy Controls
3.3. Amount of Secreted Angiopoietin-like Protein 4 (ANGPTL4) in Culture Supernatant of ECs Stimulated with Plasma Derived from Malaria Patients and Healthy Individuals
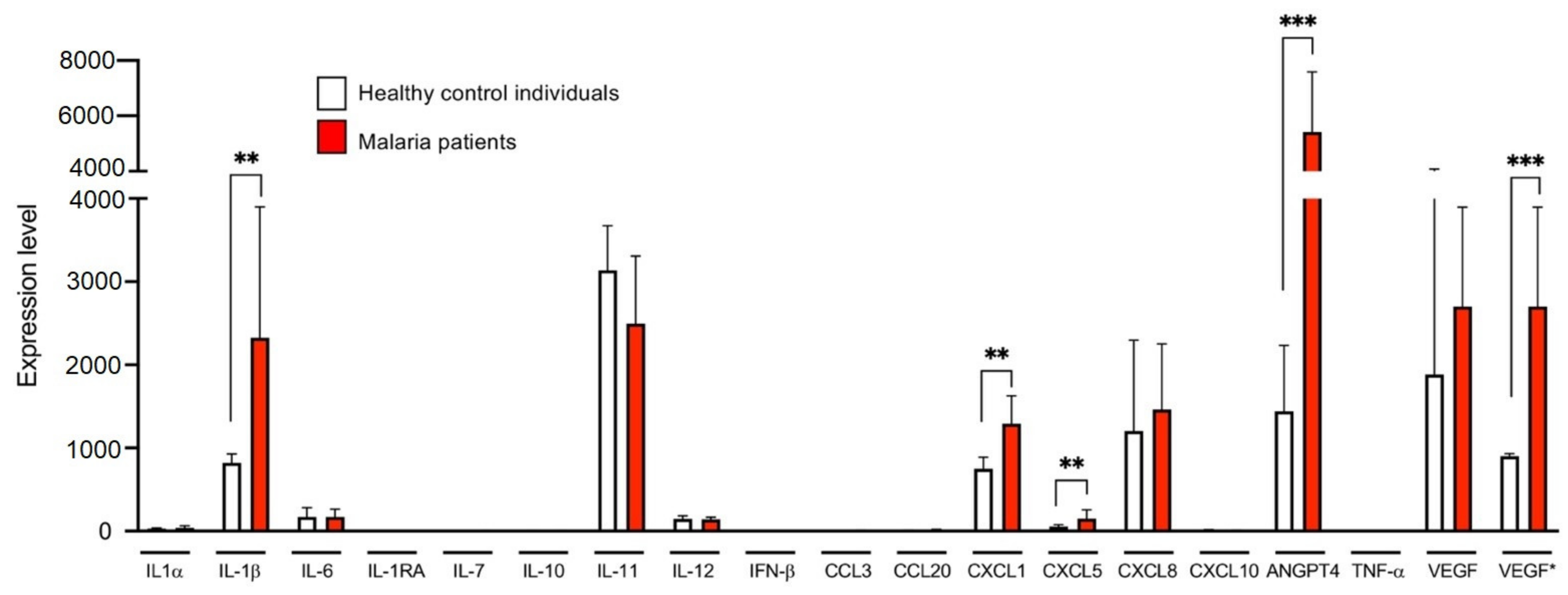
3.4. Comparative Transcriptome Analyses of ECs Stimulated with Plasma of Malaria Patients and Healthy Individuals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2020; WHO Team, Global Malaria Programme: Geneva, Switzerland, 2020; ISBN 978-92-4-001579-1. [Google Scholar]
- Milner, D.A., Jr.; Lee, J.J.; Frantzreb, C.; Whitten, R.O.; Kamiza, S.; Carr, R.A.; Pradham, A.; Factor, R.E.; Playforth, K.; Liomba, G.; et al. Quantitative assessment of multiorgan sequestration of parasites in fatal pediatric cerebral malaria. J. Infect. Dis. 2015, 212, 1317–1321. [Google Scholar] [CrossRef] [PubMed]
- Milner, D., Jr.; Factor, R.; Whitten, R.; Carr, R.A.; Kamiza, S.; Pinkus, G.; Molyneux, M.; Taylor, T. Pulmonary pathology in pediatric cerebral malaria. Hum. Pathol. 2013, 44, 2719–2726. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.E.; Fu, W.J.; Carr, R.A.; Whitten, R.O.; Mueller, J.S.; Fosiko, N.G.; Lewallen, S.; Liomba, N.G.; Molyneux, M.E. Differentiating the pathologies of cerebral malaria by postmortem parasite counts. Nat. Med. 2004, 10, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Lyke, K.E.; Burges, R.; Cissoko, Y.; Sangare, L.; Dao, M.; Diarra, I.; Kone, A.; Harley, R.; Plowe, C.V.; Doumbo, O.K.; et al. Serum levels of the proinflammatory cytokines interleukin-1 beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis factor alpha, and IL-12(p70) in Malian children with severe Plasmodium falciparum malaria and matched uncomplicated malaria or healthy controls. Infect. Immun. 2004, 72, 5630–5637. [Google Scholar] [CrossRef]
- Nishanth, G.; Schluter, D. Blood-brain barrier in cerebral malaria: Pathogenesis and therapeutic intervention. Trends Parasitol. 2019, 35, 516–528. [Google Scholar] [CrossRef]
- Dunst, J.; Kamena, F.; Matuschewski, K. Cytokines and chemokines in cerebral malaria pathogenesis. Front. Cell. Infect. Microbiol. 2017, 7, 324. [Google Scholar] [CrossRef]
- Hasday, J.D.; Bannerman, D.; Sakarya, S.; Cross, A.S.; Singh, I.S.; Howard, D.; Drysdale, B.E.; Goldblum, S.E. Exposure to febrile temperature modifies endothelial cell response to tumor necrosis factor-alpha. J. Appl. Physiol. 2001, 90, 90–98. [Google Scholar] [CrossRef]
- Oakley, M.S.; Gerald, N.; McCutchan, T.F.; Aravind, L.; Kumar, S. Clinical and molecular aspects of malaria fever. Trends Parasitol. 2011, 27, 442–449. [Google Scholar] [CrossRef]
- Oakley, M.S.; Kumar, S.; Anantharaman, V.; Zheng, H.; Mahajan, B.; Haynes, J.D.; Moch, J.K.; Fairhurst, R.; McCutchan, T.F.; Aravind, L. Molecular factors and biochemical pathways induced by febrile temperature in intraerythrocytic Plasmodium falciparum parasites. Infect. Immun. 2007, 75, 2012–2025. [Google Scholar] [CrossRef]
- Phillips, M.A.; Burrows, J.N.; Manyando, C.; van Huijsduijnen, R.H.; Van Voorhis, W.C.; Wells, T.N.C. Malaria. Nat. Rev. Dis. Primers 2017, 3, 17050. [Google Scholar] [CrossRef]
- Cunnington, A.J.; Riley, E.M.; Walther, M. Microvascular dysfunction in severe Plasmodium falciparum malaria. J. Infect. Dis. 2013, 207, 369–370. [Google Scholar] [CrossRef]
- Gazzinelli, R.T.; Kalantari, P.; Fitzgerald, K.A.; Golenbock, D.T. Innate sensing of malaria parasites. Nat. Rev. Immunol. 2014, 14, 744–757. [Google Scholar] [CrossRef]
- Boivin, M.J.; Bangirana, P.; Byarugaba, J.; Opoka, R.O.; Idro, R.; Jurek, A.M.; John, C.C. Cognitive impairment after cerebral malaria in children: A prospective study. Pediatrics 2007, 119, e360–e366. [Google Scholar] [CrossRef]
- Idro, R.; Marsh, K.; John, C.C.; Newton, C.R. Cerebral malaria: Mechanisms of brain injury and strategies for improved neurocognitive outcome. Pediatric Res. 2010, 68, 267–274. [Google Scholar] [CrossRef]
- Roze, E.; Thiebaut, M.M.; Mazevet, D.; Bricaire, F.; Danis, M.; Deseilligny, C.P.; Caumes, E. Neurologic sequelae after severe falciparum malaria in adult travelers. Eur. Neurol. 2001, 46, 192–197. [Google Scholar] [CrossRef]
- Chakravorty, S.J.; Carret, C.; Nash, G.B.; Ivens, A.; Szestak, T.; Craig, A.G. Altered phenotype and gene transcription in endothelial cells, induced by Plasmodium falciparum-infected red blood cells: Pathogenic or protective? Int. J. Parasitol. 2007, 37, 975–987. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tripathi, A.K.; Sha, W.; Shulaev, V.; Stins, M.F.; Sullivan, D.J., Jr. Plasmodium falciparum-infected erythrocytes induce NF-kappaB regulated inflammatory pathways in human cerebral endothelium. Blood 2009, 114, 4243–4252. [Google Scholar] [CrossRef] [PubMed]
- Viebig, N.K.; Wulbrand, U.; Forster, R.; Andrews, K.T.; Lanzer, M.; Knolle, P.A. Direct activation of human endothelial cells by Plasmodium falciparum-infected erythrocytes. Infect. Immun. 2005, 73, 3271–3277. [Google Scholar] [CrossRef]
- Schofield, L.; Hackett, F. Signal transduction in host cells by a glycosylphosphatidylinositol toxin of malaria parasites. J. Exp. Med. 1993, 177, 145–153. [Google Scholar] [CrossRef]
- Zhu, J.; Krishnegowda, G.; Gowda, D.C. Induction of proinflammatory responses in macrophages by the glycosylphosphatidylinositols of Plasmodium falciparum: The requirement of extracellular signal-regulated kinase, p38, c-Jun N-terminal kinase and NF-kappaB pathways for the expression of proinflammatory cytokines and nitric oxide. J. Biol. Chem. 2005, 280, 8617–8627. [Google Scholar] [CrossRef] [PubMed]
- Giribaldi, G.; Prato, M.; Ulliers, D.; Gallo, V.; Schwarzer, E.; Akide-Ndunge, O.B.; Valente, E.; Saviozzi, S.; Calogero, R.A.; Arese, P. Involvement of inflammatory chemokines in survival of human monocytes fed with malarial pigment. Infect. Immun. 2010, 78, 4912–4921. [Google Scholar] [CrossRef]
- Jaramillo, M.; Godbout, M.; Olivier, M. Hemozoin induces macrophage chemokine expression through oxidative stress-dependent and -independent mechanisms. J. Immunol. 2005, 174, 475–484. [Google Scholar] [CrossRef]
- Schwarzer, E.; Bellomo, G.; Giribaldi, G.; Ulliers, D.; Arese, P. Phagocytosis of malarial pigment haemozoin by human monocytes: A confocal microscopy study. Parasitology 2001, 123, 125–131. [Google Scholar] [CrossRef]
- Armah, H.B.; Wilson, N.O.; Sarfo, B.Y.; Powell, M.D.; Bond, V.C.; Anderson, W.; Adjei, A.A.; Gyasi, R.K.; Tettey, Y.; Wiredu, E.K.; et al. Cerebrospinal fluid and serum biomarkers of cerebral malaria mortality in Ghanaian children. Malar. J. 2007, 6, 147. [Google Scholar] [CrossRef] [PubMed]
- Dieye, Y.; Mbengue, B.; Dagamajalu, S.; Fall, M.M.; Loke, M.F.; Nguer, C.M.; Thiam, A.; Vadivelu, J.; Dieye, A. Cytokine response during non-cerebral and cerebral malaria: Evidence of a failure to control inflammation as a cause of death in African adults. PeerJ 2016, 4, e1965. [Google Scholar] [CrossRef]
- Jain, V.; Armah, H.B.; Tongren, J.E.; Ned, R.M.; Wilson, N.O.; Crawford, S.; Joel, P.K.; Singh, M.P.; Nagpal, A.C.; Dash, A.P.; et al. Plasma IP-10, apoptotic and angiogenic factors associated with fatal cerebral malaria in India. Malar. J. 2008, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Herbert, F.; Tchitchek, N.; Bansal, D.; Jacques, J.; Pathak, S.; Becavin, C.; Fesel, C.; Dalko, E.; Cazenave, P.A.; Preda, C.; et al. Evidence of IL-17, IP-10, and IL-10 involvement in multiple-organ dysfunction and IL-17 pathway in acute renal failure associated to Plasmodium falciparum malaria. J. Transl. Med. 2015, 13, 369. [Google Scholar] [CrossRef]
- Wilson, N.O.; Jain, V.; Roberts, C.E.; Lucchi, N.; Joel, P.K.; Singh, M.P.; Nagpal, A.C.; Dash, A.P.; Udhayakumar, V.; Singh, N.; et al. CXCL4 and CXCL10 predict risk of fatal cerebral malaria. Dis. Markers 2011, 30, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Berg, A.; Patel, S.; Gonca, M.; David, C.; Otterdal, K.; Ueland, T.; Dalen, I.; Kvaloy, J.T.; Mollnes, T.E.; Aukrust, P.; et al. Cytokine network in adults with falciparum malaria and HIV-1: Increased IL-8 and IP-10 levels are associated with disease severity. PLoS ONE 2014, 9, e114480. [Google Scholar] [CrossRef] [PubMed]
- Prakash, D.; Fesel, C.; Jain, R.; Cazenave, P.A.; Mishra, G.C.; Pied, S. Clusters of cytokines determine malaria severity in Plasmodium falciparum-infected patients from endemic areas of Central India. J. Infect. Dis. 2006, 194, 198–207. [Google Scholar] [CrossRef]
- Colborn, J.M.; Ylostalo, J.H.; Koita, O.A.; Cisse, O.H.; Krogstad, D.J. Human gene expression in uncomplicated Plasmodium falciparum malaria. J. Immunol. Res. 2015, 2015, 162639. [Google Scholar] [CrossRef]
- Bwanika, R.; Kato, C.D.; Welishe, J.; Mwandah, D.C. Cytokine profiles among patients co-infected with Plasmodium falciparum malaria and soil borne helminths attending Kampala International University Teaching Hospital, in Uganda. Allergy Asthma Clin. Immunol. 2018, 14, 10. [Google Scholar] [CrossRef]
- Thuma, P.E.; van Dijk, J.; Bucala, R.; Debebe, Z.; Nekhai, S.; Kuddo, T.; Nouraie, M.; Weiss, G.; Gordeuk, V.R. Distinct clinical and immunologic profiles in severe malarial anemia and cerebral malaria in Zambia. J. Infect. Dis. 2011, 203, 211–219. [Google Scholar] [CrossRef]
- Che, J.N.; Nmorsi, O.P.; Nkot, B.P.; Isaac, C.; Okonkwo, B.C. Chemokines responses to Plasmodium falciparum malaria and co-infections among rural Cameroonians. Parasitol. Int. 2015, 64, 139–144. [Google Scholar] [CrossRef]
- Ayimba, E.; Hegewald, J.; Segbena, A.Y.; Gantin, R.G.; Lechner, C.J.; Agosssou, A.; Banla, M.; Soboslay, P.T. Proinflammatory and regulatory cytokines and chemokines in infants with uncomplicated and severe Plasmodium falciparum malaria. Clin. Exp. Immunol. 2011, 166, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Burgmann, H.; Hollenstein, U.; Wenisch, C.; Thalhammer, F.; Looareesuwan, S.; Graninger, W. Serum concentrations of MIP-1 alpha and interleukin-8 in patients suffering from acute Plasmodium falciparum malaria. Clin. Immunol. Immunopathol. 1995, 76, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Otterdal, K.; Berg, A.; Michelsen, A.E.; Patel, S.; Gregersen, I.; Sagen, E.L.; Halvorsen, B.; Yndestad, A.; Ueland, T.; Langeland, N.; et al. Plasma levels of interleukin 27 in falciparum malaria is increased independently of co-infection with HIV: Potential immune-regulatory role during malaria. BMC Infect. Dis. 2020, 20, 65. [Google Scholar] [CrossRef]
- John, C.C.; Panoskaltsis-Mortari, A.; Opoka, R.O.; Park, G.S.; Orchard, P.J.; Jurek, A.M.; Idro, R.; Byarugaba, J.; Boivin, M.J. Cerebrospinal fluid cytokine levels and cognitive impairment in cerebral malaria. Am. J. Trop. Med. Hyg. 2008, 78, 198–205. [Google Scholar] [CrossRef] [PubMed]
- John, C.C.; Park, G.S.; Sam-Agudu, N.; Opoka, R.O.; Boivin, M.J. Elevated serum levels of IL-1ra in children with Plasmodium falciparum malaria are associated with increased severity of disease. Cytokine 2008, 41, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate—A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, A.; Ma, T.; Menon, D.; Deshpande, M.; Jee, K.; Dinabandhu, A.; Vancel, J.; Lu, D.; Montaner, S. Angiopoietin-like 4 binds neuropilins and cooperates with VEGF to induce diabetic macular edema. J. Clin. Investig. 2019, 129, 4593–4608. [Google Scholar] [CrossRef]
- Gealekman, O.; Burkart, A.; Chouinard, M.; Nicoloro, S.M.; Straubhaar, J.; Corvera, S. Enhanced angiogenesis in obesity and in response to PPARgamma activators through adipocyte VEGF and ANGPTL4 production. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1056–E1064. [Google Scholar] [CrossRef] [PubMed]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Snel, B.; Lehmann, G.; Bork, P.; Huynen, M.A. STRING: A web-server to retrieve and display the repeatedly occurring neighbourhood of a gene. Nucleic Acids Res. 2000, 28, 3442–3444. [Google Scholar] [CrossRef]
- Ioannidis, L.J.; Nie, C.Q.; Hansen, D.S. The role of chemokines in severe malaria: More than meets the eye. Parasitology 2014, 141, 602–613. [Google Scholar] [CrossRef]
- Wu, X.; Gowda, N.M.; Kumar, S.; Gowda, D.C. Protein-DNA complex is the exclusive malaria parasite component that activates dendritic cells and triggers innate immune responses. J. Immunol. 2010, 184, 4338–4348. [Google Scholar] [CrossRef]
- Opitz, B.; Hippenstiel, S.; Eitel, J.; Suttorp, N. Extra- and intracellular innate immune recognition in endothelial cells. Thromb. Haemost. 2007, 98, 319–326. [Google Scholar] [CrossRef]
- Nourshargh, S.; Alon, R. Leukocyte migration into inflamed tissues. Immunity 2014, 41, 694–707. [Google Scholar] [CrossRef]
- Middleton, J.; Patterson, A.M.; Gardner, L.; Schmutz, C.; Ashton, B.A. Leukocyte extravasation: Chemokine transport and presentation by the endothelium. Blood 2002, 100, 3853–3860. [Google Scholar] [CrossRef]
- Mordelet, E.; Davies, H.A.; Hillyer, P.; Romero, I.A.; Male, D. Chemokine transport across human vascular endothelial cells. Endothelium 2007, 14, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Adams, Y.; Olsen, R.W.; Bengtsson, A.; Dalgaard, N.; Zdioruk, M.; Satpathi, S.; Behera, P.K.; Sahu, P.K.; Lawler, S.E.; Qvortrup, K.; et al. Plasmodium falciparum erythrocyte membrane protein 1 variants induce cell swelling and disrupt the blood-brain barrier in cerebral malaria. J. Exp. Med. 2021, 218. [Google Scholar] [CrossRef]
- Nilsen, E.M.; Johansen, F.E.; Jahnsen, F.L.; Lundin, K.E.; Scholz, T.; Brandtzaeg, P.; Haraldsen, G. Cytokine profiles of cultured microvascular endothelial cells from the human intestine. Gut 1998, 42, 635–642. [Google Scholar] [CrossRef]
- Briones, M.A.; Phillips, D.J.; Renshaw, M.A.; Hooper, W.C. Expression of chemokine by human coronary-artery and umbilical-vein endothelial cells and its regulation by inflammatory cytokines. Coron. Artery Dis. 2001, 12, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Basilico, N.; Corbett, Y.; D’Alessandro, S.; Parapini, S.; Prato, M.; Girelli, D.; Misiano, P.; Olliaro, P.; Taramelli, D. Malaria pigment stimulates chemokine production by human microvascular endothelium. Acta Trop. 2017, 172, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Gillrie, M.R.; Lee, K.; Gowda, D.C.; Davis, S.P.; Monestier, M.; Cui, L.; Hien, T.T.; Day, N.P.J.; Ho, M. Plasmodium falciparum histones induce endothelial proinflammatory response and barrier dysfunction. Am. J. Pathol. 2012, 180, 1028–1039. [Google Scholar] [CrossRef]
- Chakravorty, S.J.; Hughes, K.R.; Craig, A.G. Host response to cytoadherence in Plasmodium falciparum. Biochem. Soc. Trans. 2008, 36, 221–228. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hillyer, P.; Mordelet, E.; Flynn, G.; Male, D. Chemokines, chemokine receptors and adhesion molecules on different human endothelia: Discriminating the tissue-specific functions that affect leucocyte migration. Clin. Exp. Immunol. 2003, 134, 431–441. [Google Scholar] [CrossRef]
- Wichers, J.S.; Tonkin-Hill, G.; Thye, T.; Krumkamp, R.; Kreuels, B.; Strauss, J.; von Thien, H.; Scholz, J.A.; Smedegaard Hansson, H.; Weisel Jensen, R.; et al. Common virulence gene expression in adult first-time infected malaria patients and severe cases. Elife 2021, 10. [Google Scholar] [CrossRef]
- Oggungwan, K.; Glaharn, S.; Ampawong, S.; Krudsood, S.; Viriyavejakul, P. FTY720 restores endothelial cell permeability induced by malaria sera. Sci. Rep. 2018, 8, 10959. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Sullivan, D.J.; Stins, M.F. Plasmodium falciparum-infected erythrocytes increase intercellular adhesion molecule 1 expression on brain endothelium through NF-kappaB. Infect. Immun. 2006, 74, 3262–3270. [Google Scholar] [CrossRef]
- Mohan, A.; Sharma, S.K.; Bollineni, S. Acute lung injury and acute respiratory distress syndrome in malaria. J. Vector Borne Dis. 2008, 45, 179–193. [Google Scholar] [PubMed]
- Pal, P.; Daniels, B.P.; Oskman, A.; Diamond, M.S.; Klein, R.S.; Goldberg, D.E. Plasmodium falciparum histidine-rich protein II compromises brain endothelial barriers and may promote cerebral malaria pathogenesis. mBio 2016, 7. [Google Scholar] [CrossRef]
- Gallego-Delgado, J.; Basu-Roy, U.; Ty, M.; Alique, M.; Fernandez-Arias, C.; Movila, A.; Gomes, P.; Weinstock, A.; Xu, W.; Edagha, I.; et al. Angiotensin receptors and beta-catenin regulate brain endothelial integrity in malaria. J. Clin. Investig. 2016, 126, 4016–4029. [Google Scholar] [CrossRef] [PubMed]
- Luster, A.D.; Unkeless, J.C.; Ravetch, J.V. Gamma-interferon transcriptionally regulates an early-response gene containing homology to platelet proteins. Nature 1985, 315, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Mai, J.; Virtue, A.; Shen, J.; Wang, H.; Yang, X.F. An evolving new paradigm: Endothelial cells—Conditional innate immune cells. J. Hematol. Oncol. 2013, 6, 61. [Google Scholar] [CrossRef]
- Wassmer, S.C.; Combes, V.; Candal, F.J.; Juhan-Vague, I.; Grau, G.E. Platelets potentiate brain endothelial alterations induced by Plasmodium falciparum. Infect. Immun. 2006, 74, 645–653. [Google Scholar] [CrossRef]
- Dorpinghaus, M.; Furstenwerth, F.; Roth, L.K.; Bouws, P.; Rakotonirinalalao, M.; Jordan, V.; Sauer, M.; Rehn, T.; Pansegrau, E.; Hohn, K.; et al. Stringent selection of knobby Plasmodium falciparum-infected erythrocytes during cytoadhesion at febrile temperature. Microorganisms 2020, 8, 174. [Google Scholar] [CrossRef] [PubMed]
- Semple, B.D.; Kossmann, T.; Morganti-Kossmann, M.C. Role of chemokines in CNS health and pathology: A focus on the CCL2/CCR2 and CXCL8/CXCR2 networks. J. Cereb. Blood Flow Metab. 2010, 30, 459–473. [Google Scholar] [CrossRef]
- Li, A.; Dubey, S.; Varney, M.L.; Dave, B.J.; Singh, R.K. IL-8 directly enhanced endothelial cell survival, proliferation, and matrix metalloproteinases production and regulated angiogenesis. J. Immunol. 2003, 170, 3369–3376. [Google Scholar] [CrossRef]
- Heidemann, J.; Ogawa, H.; Dwinell, M.B.; Rafiee, P.; Maaser, C.; Gockel, H.R.; Otterson, M.F.; Ota, D.M.; Lugering, N.; Domschke, W.; et al. Angiogenic effects of interleukin 8 (CXCL8) in human intestinal microvascular endothelial cells are mediated by CXCR2. J. Biol. Chem. 2003, 278, 8508–8515. [Google Scholar] [CrossRef]
- Yu, M.; Ma, X.; Jiang, D.; Wang, L.; Zhan, Q.; Zhao, J. CXC chemokine ligand 5 (CXCL5) disrupted the permeability of human brain microvascular endothelial cells via regulating p38 signal. Microbiol. Immunol. 2021, 65, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Haarmann, A.; Schuhmann, M.K.; Silwedel, C.; Monoranu, C.M.; Stoll, G.; Buttmann, M. Human brain endothelial CXCR2 is inflammation-inducible and mediates CXCL5- and CXCL8-triggered paraendothelial barrier breakdown. Int. J. Mol. Sci. 2019, 20, 602. [Google Scholar] [CrossRef]
- Chandrasekar, B.; Melby, P.C.; Sarau, H.M.; Raveendran, M.; Perla, R.P.; Marelli-Berg, F.M.; Dulin, N.O.; Singh, I.S. Chemokine-cytokine cross-talk. The ELR+ CXC chemokine LIX (CXCL5) amplifies a proinflammatory cytokine response via a phosphatidylinositol 3-kinase-NF-kappa B pathway. J. Biol. Chem. 2003, 278, 4675–4686. [Google Scholar] [CrossRef]
- Kossodo, S.; Monso, C.; Juillard, P.; Velu, T.; Goldman, M.; Grau, G.E. Interleukin-10 modulates susceptibility in experimental cerebral malaria. Immunology 1997, 91, 536–540. [Google Scholar] [CrossRef]
- Sanni, L.A.; Jarra, W.; Li, C.; Langhorne, J. Cerebral edema and cerebral hemorrhages in interleukin-10-deficient mice infected with Plasmodium chabaudi. Infect. Immun. 2004, 72, 3054–3058. [Google Scholar] [CrossRef] [PubMed]
- Mandala, W.L.; Msefula, C.L.; Gondwe, E.N.; Drayson, M.T.; Molyneux, M.E.; MacLennan, C.A. Cytokine profiles in Malawian children presenting with uncomplicated malaria, severe malarial anemia, and cerebral malaria. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef]
- Seydel, K.B.; Kampondeni, S.D.; Valim, C.; Potchen, M.J.; Milner, D.A.; Muwalo, F.W.; Birbeck, G.L.; Bradley, W.G.; Fox, L.L.; Glover, S.J.; et al. Brain swelling and death in children with cerebral malaria. N. Engl. J. Med. 2015, 372, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Harawa, V.; Njie, M.; Kessler, A.; Choko, A.; Kumwenda, B.; Kampondeni, S.; Potchen, M.; Kim, K.; Jaworowski, A.; Taylor, T.; et al. Brain swelling is independent of peripheral plasma cytokine levels in Malawian children with cerebral malaria. Malar. J. 2018, 17, 435. [Google Scholar] [CrossRef]
- Jason, J.; Archibald, L.K.; Nwanyanwu, O.C.; Byrd, M.G.; Kazembe, P.N.; Dobbie, H.; Jarvis, W.R. Comparison of serum and cell-specific cytokines in humans. Clin. Diagn. Lab. Immunol. 2001, 8, 1097–1103. [Google Scholar] [CrossRef]
- Worzfeld, T.; Schwaninger, M. Apicobasal polarity of brain endothelial cells. J. Cereb. Blood Flow Metab. 2016, 36, 340–362. [Google Scholar] [CrossRef]
- Ferrara, N.; Chen, H.; Davis-Smyth, T.; Gerber, H.P.; Nguyen, T.N.; Peers, D.; Chisholm, V.; Hillan, K.J.; Schwall, R.H. Vascular endothelial growth factor is essential for corpus luteum angiogenesis. Nat. Med. 1998, 4, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Gerber, H.P.; Dixit, V.; Ferrara, N. Vascular endothelial growth factor induces expression of the antiapoptotic proteins Bcl-2 and A1 in vascular endothelial cells. J. Biol. Chem. 1998, 273, 13313–13316. [Google Scholar] [CrossRef]
- Yuan, F.; Chen, Y.; Dellian, M.; Safabakhsh, N.; Ferrara, N.; Jain, R.K. Time-dependent vascular regression and permeability changes in established human tumor xenografts induced by an anti-vascular endothelial growth factor/vascular permeability factor antibody. Proc. Natl. Acad. Sci. USA 1996, 93, 14765–14770. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Kimura, M.; Watanabe, N. Elevated levels of vascular endothelial growth factor (VEGF) and soluble vascular endothelial growth factor receptor (VEGFR)-2 in human malaria. Am. J. Trop. Med. Hyg. 2010, 82, 136–139. [Google Scholar] [CrossRef]
- Leung, D.W.; Cachianes, G.; Kuang, W.J.; Goeddel, D.V.; Ferrara, N. Vascular endothelial growth factor is a secreted angiogenic mitogen. Science 1989, 246, 1306–1309. [Google Scholar] [CrossRef] [PubMed]
- Azzouz, M.; Ralph, G.S.; Storkebaum, E.; Walmsley, L.E.; Mitrophanous, K.A.; Kingsman, S.M.; Carmeliet, P.; Mazarakis, N.D. VEGF delivery with retrogradely transported lentivector prolongs survival in a mouse ALS model. Nature 2004, 429, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Krum, J.M.; Khaibullina, A. Inhibition of endogenous VEGF impedes revascularization and astroglial proliferation: Roles for VEGF in brain repair. Exp. Neurol. 2003, 181, 241–257. [Google Scholar] [CrossRef]
- Canavese, M.; Spaccapelo, R. Protective or pathogenic effects of vascular endothelial growth factor (VEGF) as potential biomarker in cerebral malaria. Pathog. Glob. Health 2014, 108, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Deininger, M.H.; Winkler, S.; Kremsner, P.G.; Meyermann, R.; Schluesener, H.J. Angiogenic proteins in brains of patients who died with cerebral malaria. J. Neuroimmunol. 2003, 142, 101–111. [Google Scholar] [CrossRef]
- Kwiatkowski, D.P. How malaria has affected the human genome and what human genetics can teach us about malaria. Am. J. Hum. Genet. 2005, 77, 171–192. [Google Scholar] [CrossRef]
- La Paglia, L.; Listi, A.; Caruso, S.; Amodeo, V.; Passiglia, F.; Bazan, V.; Fanale, D. Potential role of ANGPTL4 in the cross talk between metabolism and cancer through PPAR signaling pathway. PPAR Res. 2017, 2017, 8187235. [Google Scholar] [CrossRef] [PubMed]
- Le Jan, S.; Amy, C.; Cazes, A.; Monnot, C.; Lamande, N.; Favier, J.; Philippe, J.; Sibony, M.; Gasc, J.M.; Corvol, P.; et al. Angiopoietin-like 4 is a proangiogenic factor produced during ischemia and in conventional renal cell carcinoma. Am. J. Pathol. 2003, 162, 1521–1528. [Google Scholar] [CrossRef]
- Liu, Y.; Cox, S.R.; Morita, T.; Kourembanas, S. Hypoxia regulates vascular endothelial growth factor gene expression in endothelial cells. Identification of a 5’ enhancer. Circ. Res. 1995, 77, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Park, M.K.; Ko, E.J.; Jeon, K.Y.; Kim, H.; Jo, J.O.; Baek, K.W.; Kang, Y.J.; Choi, Y.H.; Hong, Y.; Ock, M.S.; et al. Induction of angiogenesis by malarial infection through hypoxia dependent manner. Korean J. Parasitol. 2019, 57, 117–125. [Google Scholar] [CrossRef]
- Angiolillo, A.L.; Sgadari, C.; Taub, D.D.; Liao, F.; Farber, J.M.; Maheshwari, S.; Kleinman, H.K.; Reaman, G.H.; Tosato, G. Human interferon-inducible protein 10 is a potent inhibitor of angiogenesis in vivo. J. Exp. Med. 1995, 182, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Strieter, R.M.; Kunkel, S.L.; Arenberg, D.A.; Burdick, M.D.; Polverini, P.J. Interferon gamma-inducible protein 10 (IP-10), a member of the C-X-C chemokine family, is an inhibitor of angiogenesis. Biochem. Biophys. Res. Commun. 1995, 210, 51–57. [Google Scholar] [CrossRef]
- Campanella, G.S.; Colvin, R.A.; Luster, A.D. CXCL10 can inhibit endothelial cell proliferation independently of CXCR3. PLoS ONE 2010, 5, e12700. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, E.W.; Lian, J.; Ozga, A.J.; Miyabe, Y.; Ji, S.W.; Bromley, S.K.; Mempel, T.R.; Luster, A.D. CXCL10 stabilizes T cell-brain endothelial cell adhesion leading to the induction of cerebral malaria. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Bodnar, R.J.; Yates, C.C.; Wells, A. IP-10 blocks vascular endothelial growth factor-induced endothelial cell motility and tube formation via inhibition of calpain. Circ. Res. 2006, 98, 617–625. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
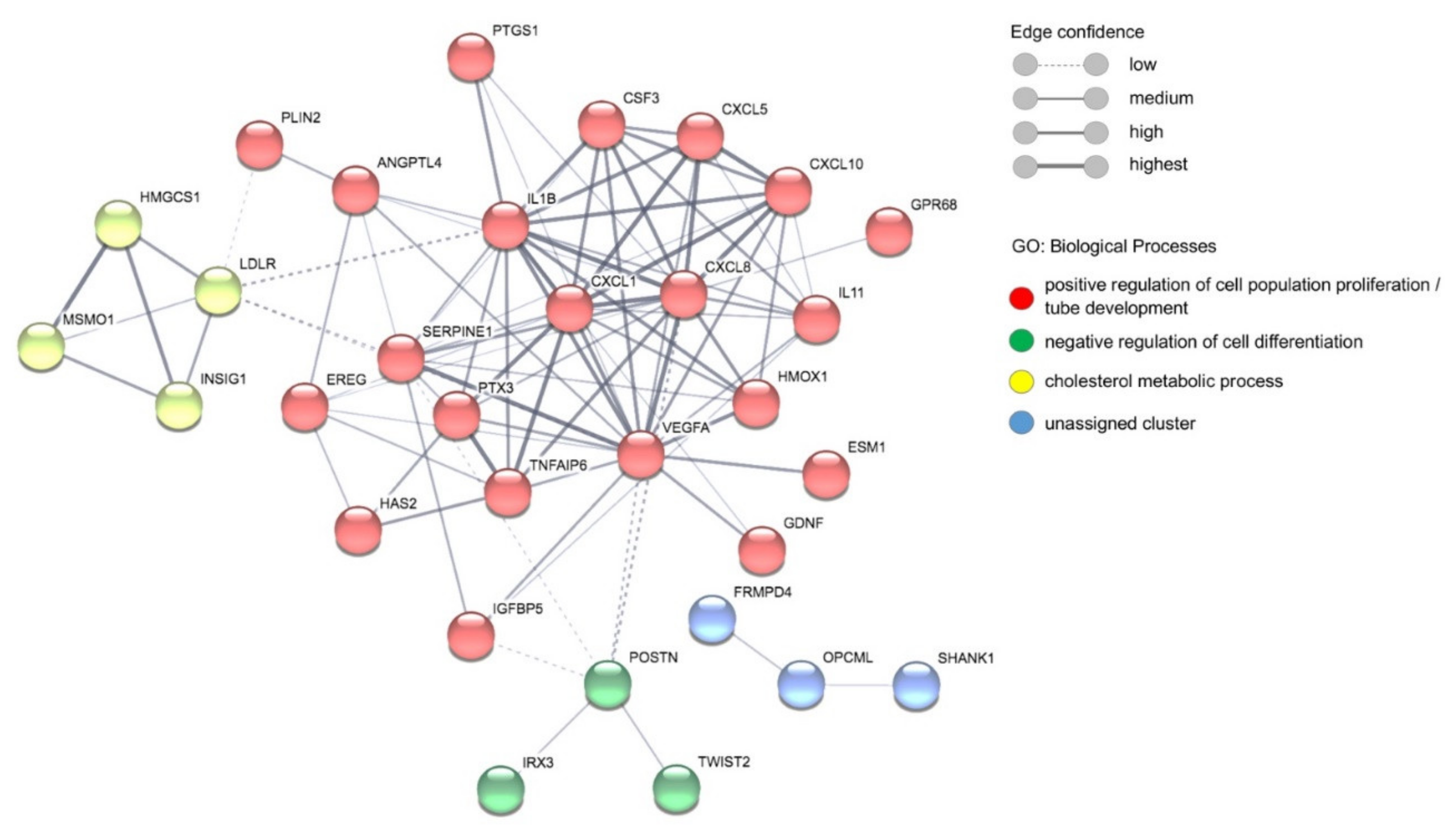{kind=link}
| Gene | Expression Level | Fold Change | padj | GO:BP | KEGG | MCL | Name | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H5, 10, 8 | M6, 9, 10, 11 | 1 | 2 | 3 | 4 | 5 | ||||||
| 1 | CSF3 | 16 | 73 | 4.5 | 0.016 | Colony Stimulating Factor 3 | ||||||
| 2 | ANGPTL4 | 1442 | 5419 | 3.8 | 4.17 × 10−6 | Angiopoietin Like 4 | ||||||
| 3 | IL1B | 821 | 2324 | 2.8 | 0.019 | Interleukin 1b | ||||||
| 4 | CXCL5 | 55 | 149 | 2.7 | 0.049 | Chemokine cxcl5 | ||||||
| 5 | FOXS1 | 18 | 47 | 2.7 | 0.002 | Forkhead Box S1 | ||||||
| 6 | HMOX1 | 410 | 1083 | 2.6 | 0.033 | Heme Oxygenase 1 | ||||||
| 7 | GDNF | 39 | 98 | 2.5 | 0.044 | Glial Cell Derived Neurotrophic Factor | ||||||
| 8 | OPCML | 29 | 71 | 2.4 | 3.1 × 10−5 | Opioid Binding Protein/Cell Adhesion Mol. Like | ||||||
| 9 | GPR68 | 23 | 57 | 2.4 | 0.015 | G Protein-Coupled Receptor 68 | ||||||
| 10 | INSIG1 | 1065 | 2543 | 2.4 | 0.002 | Insulin Induced Gene 1 | ||||||
| 11 | TNFAIP6 | 21 | 49 | 2.3 | 0.026 | TNF Alpha Induced Protein 6 | ||||||
| 12 | IGFBP5 | 3873 | 8901 | 2.3 | 3.15 × 10−7 | Insulin Like Growth Factor Binding Protein 5 | ||||||
| 13 | ESM1 | 809 | 1783 | 2.2 | 0.022 | Endothelial Cell Specific Molecule 1 | ||||||
| 14 | POSTN | 920 | 1922 | 2.1 | 5.43 × 10−5 | Periostin | ||||||
| 15 | SERPINA9 | 57 | 116 | 2 | 3.75 × 10−5 | Serpin Family A Member 9 | ||||||
| 16 | ADTRP | 49 | 100 | 2 | 0.023 | Androgen Dependent TFPI Regulating Protein | ||||||
| 17 | MSMO1 | 1097 | 2235 | 2 | 0.0002 | Methylsterol Monooxygenase 1 | ||||||
| 18 | HAS2 | 1299 | 2642 | 2 | 0.041 | Hyaluron Synthase 2 | ||||||
| 19 | LDLR | 1491 | 3030 | 2 | 0.0012 | Low Density Lipoprotein Receptor | ||||||
| 20 | SHANK1 | 25 | 51 | 2 | 0.0036 | SH3 And Multiple Ankyrin Repeat Domains 1_2 | ||||||
| 21 | RFX8 | 38 | 76 | 2 | 0.0031 | RFX Family Member 8, Lacking RFX DNA bd. | ||||||
| 22 | CHST2 | 305 | 606 | 2 | 0.0012 | Carbohydrate Sulfotransferase 2 | ||||||
| 23 | EREG | 128 | 253 | 2 | 0.0126 | Epiregulin | ||||||
| 24 | PTX3 | 6486 | 12598 | 1.9 | 0.00012 | Pentraxin 3 | ||||||
| 25 | PTGS1 | 66 | 128 | 1.9 | 0.0123 | Prostaglandin-Endoperoxide Synthase 1 | ||||||
| 26 | RPSAP52 | 69 | 133 | 1.9 | 1.2 × 10−7 | Ribosomal Protein SA Pseudogene 52 | ||||||
| 27 | FAM84A | 241 | 455 | 1.9 | 0.0012 | LRAT Domain Containing 1 | ||||||
| 28 | LAMC2 | 1258 | 2329 | 1.9 | 0.0341 | Laminin Subunit Gamma 2 | ||||||
| 29 | SERPINE1 | 57674 | 106566 | 1.8 | 0.0002 | Serpin Family E Member 1 | ||||||
| 30 | TMEM158 | 752 | 1377 | 1.8 | 3.63 × 10−5 | Transmembrane Protein 158 | ||||||
| 31 | FRMPD4 | 54 | 97 | 1.8 | 0.023 | FERM And PDZ Domain Containing | ||||||
| 32 | B3GNT5 | 150 | 267 | 1.8 | 0.0099 | UDP-GlcNAc:BetaGal Beta-1,3-N-Acetylglucosaminyltransferase 5 | ||||||
| 33 | CAMK1G | 111 | 197 | 1.8 | 4.31 × 10−5 | Calcium/Calmodulin Dependent Protein Kinase IG | ||||||
| 34 | HMGCS1 | 1014 | 1798 | 1.8 | 0.016 | 3-Hydroxy-3-Methylglutaryl-CoA Synthase 1 | ||||||
| 35 | PQLC2L | 37 | 65 | 1.7 | 0.0176 | Solute Carrier Family 66 Member 1 Like | ||||||
| 36 | CD93 | 114 | 197 | 1.7 | 0.00764 | CD93 Molecule | ||||||
| 37 | TWIST2 | 122 | 212 | 1.7 | 0.0095 | Twist Family BHLH Transcription Factor 2 | ||||||
| 38 | IRX3 | 82 | 143 | 1.7 | 0.0035 | Iroquois Homeobox 3 | ||||||
| 39 | CXCL1 | 749 | 1293 | 1.7 | 0.0017 | C-X-C Motif Chemokine Ligand 1 | ||||||
| 40 | RRAD | 146 | 251 | 1.7 | 0.0301 | Ras Related Glycolysis Inhibitor/Calcium Channel Reg. | ||||||
| 41 | POU2F2 | 104 | 180 | 1.7 | 0.0065 | POU Class 2 Homeobox 2 | ||||||
| 42 | HSPA1B | 3762 | 6447 | 1.7 | 0.0055 | Heat Shock Protein Family A (Hsp70) Member 1B | ||||||
| 43 | PLIN2 | 2740 | 4688 | 1.7 | 0.0022 | Perilipin 2 | ||||||
| 44 * | VEGF | 1884 | 2700 | 1.4 | ns | Vascular Epidermal Growth Factor | ||||||
| 45 * | IL11 | 3135 | 2494 | 0.8 | ns | Interleukin 11 | ||||||
| 46 * | CXCL10 | 7.3 | 5.2 | 0.67 | ns | C-X-C Motif Chemokine Ligand 10 | ||||||
| 47 * | CXCL8 | 1203 | 1462 | 1.2 | ns | C-X-C Motif Chemokine Ligand 8 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raacke, M.; Kerr, A.; Dörpinghaus, M.; Brehmer, J.; Wu, Y.; Lorenzen, S.; Fink, C.; Jacobs, T.; Roeder, T.; Sellau, J.; et al. Altered Cytokine Response of Human Brain Endothelial Cells after Stimulation with Malaria Patient Plasma. Cells 2021, 10, 1656. https://doi.org/10.3390/cells10071656
Raacke M, Kerr A, Dörpinghaus M, Brehmer J, Wu Y, Lorenzen S, Fink C, Jacobs T, Roeder T, Sellau J, et al. Altered Cytokine Response of Human Brain Endothelial Cells after Stimulation with Malaria Patient Plasma. Cells. 2021; 10(7):1656. https://doi.org/10.3390/cells10071656
Chicago/Turabian StyleRaacke, Michaela, Amy Kerr, Michael Dörpinghaus, Jana Brehmer, Yifan Wu, Stephan Lorenzen, Christine Fink, Thomas Jacobs, Thomas Roeder, Julie Sellau, and et al. 2021. "Altered Cytokine Response of Human Brain Endothelial Cells after Stimulation with Malaria Patient Plasma" Cells 10, no. 7: 1656. https://doi.org/10.3390/cells10071656
APA StyleRaacke, M., Kerr, A., Dörpinghaus, M., Brehmer, J., Wu, Y., Lorenzen, S., Fink, C., Jacobs, T., Roeder, T., Sellau, J., Bachmann, A., Metwally, N. G., & Bruchhaus, I. (2021). Altered Cytokine Response of Human Brain Endothelial Cells after Stimulation with Malaria Patient Plasma. Cells, 10(7), 1656. https://doi.org/10.3390/cells10071656