
NR4A1 Regulates Tamoxifen Resistance by Suppressing ERK Signaling in ER-Positive Breast Cancer


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatment
2.2. Plasmid and siRNA Transfection
2.3. Total RNA Isolation and Quantitative Real-Time PCR
2.4. Cell Proliferation Assay
2.5. Spheroid Formation Assay
2.6. Invasion and Migration Assays
2.7. Apoptosis Assay by Annexin V-FITC/Propidium Iodide (PI) Staining
2.8. Nuclear and Cytoplasmic Protein Fractionation
2.9. Immunocytochemisty
2.10. Western Blot Analysis
2.11. In Silico Analysis
2.11.1. Gene Expression Omnibus (GEO) Dataset
2.11.2. Kaplan-Meier Survival Analysis
2.11.3. cBioportal
2.11.4. Gene Ontology Functional Enrichment Analysis
2.11.5. Gene Set Enrichment Analysis (GSEA)
2.12. Statistical Analysis
3. Results
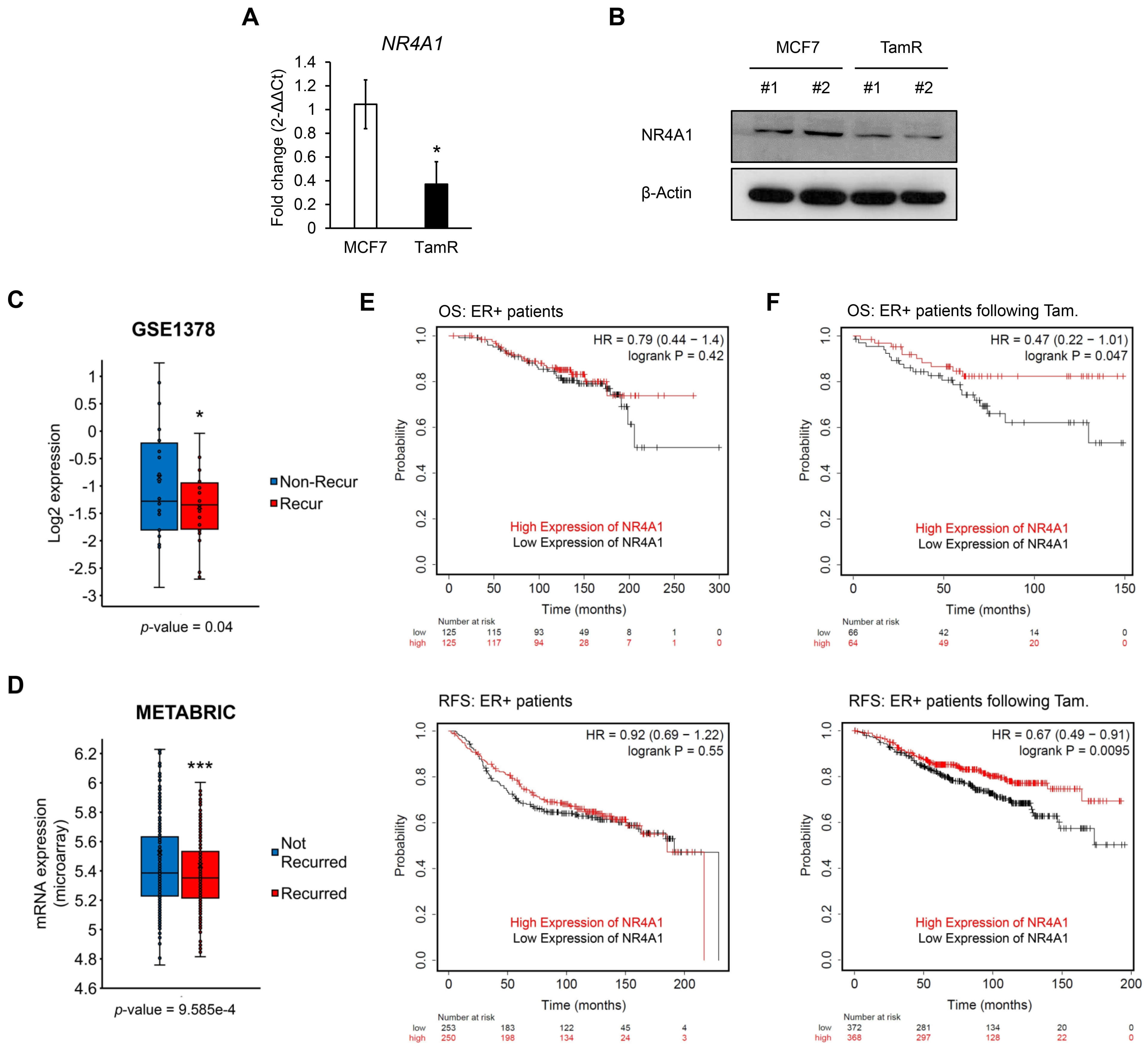
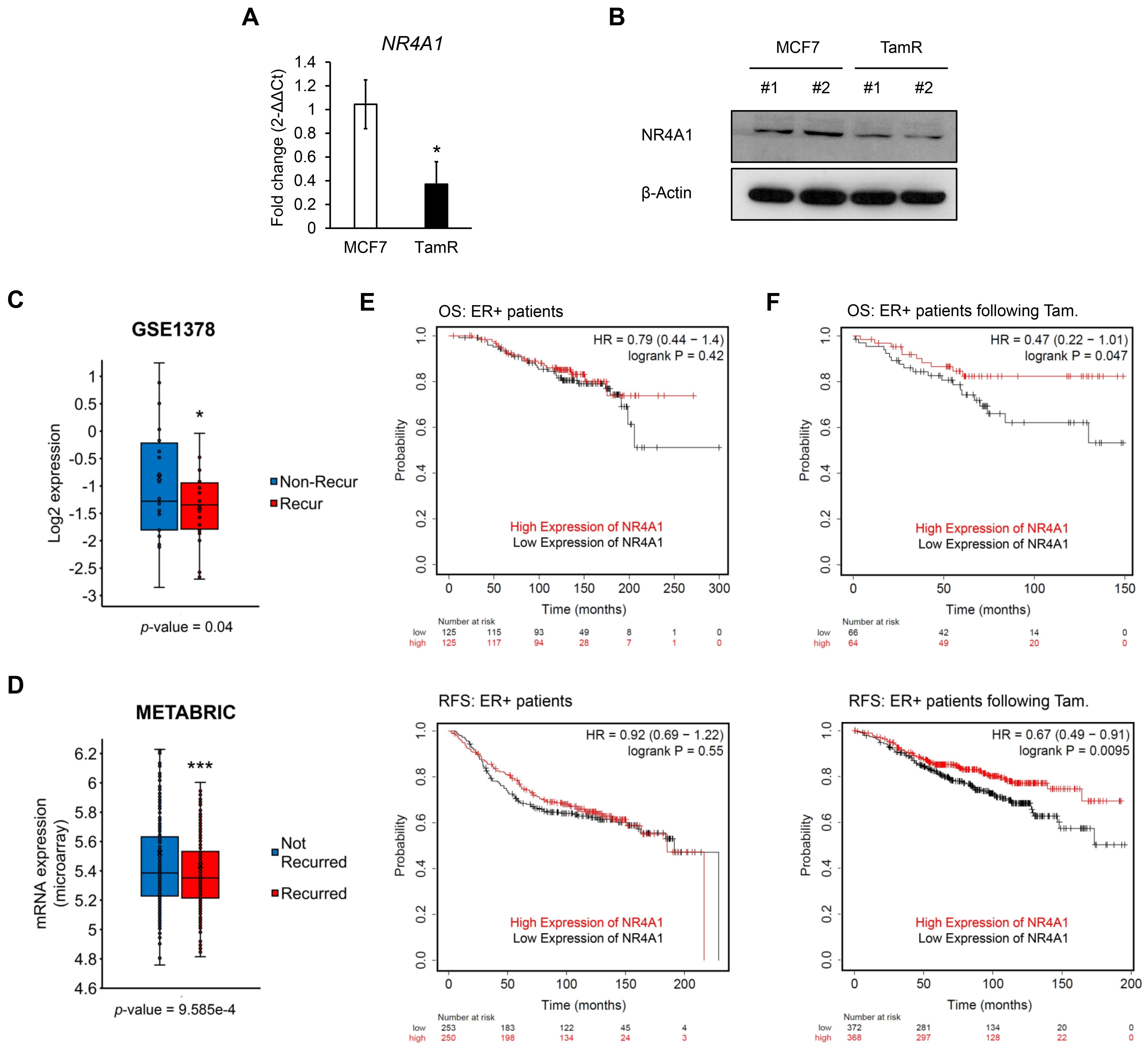
3.1. NR4A1 Is Associated with Tamoxifen Sensitivity in ER-Positive Breast Cancer
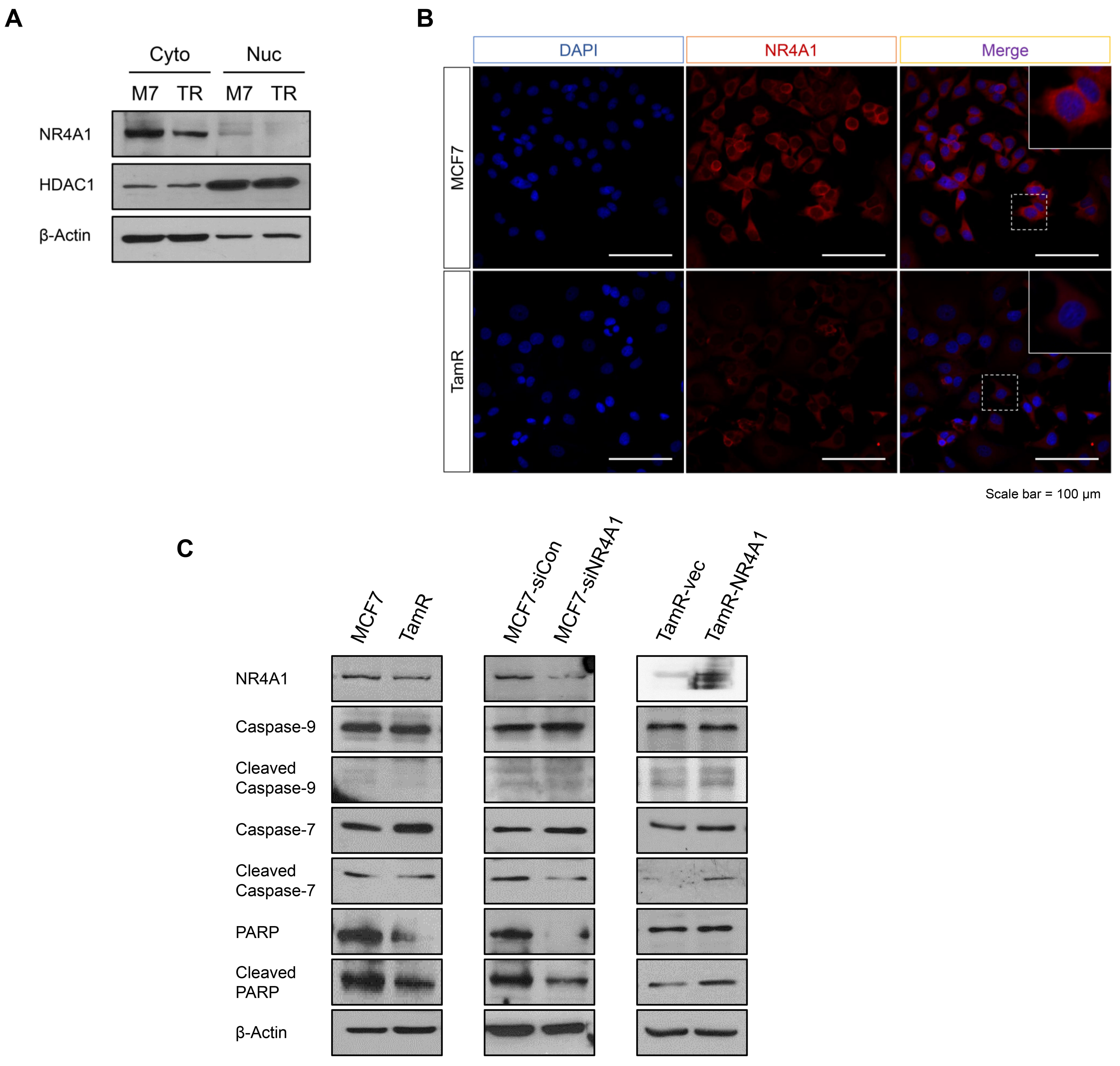
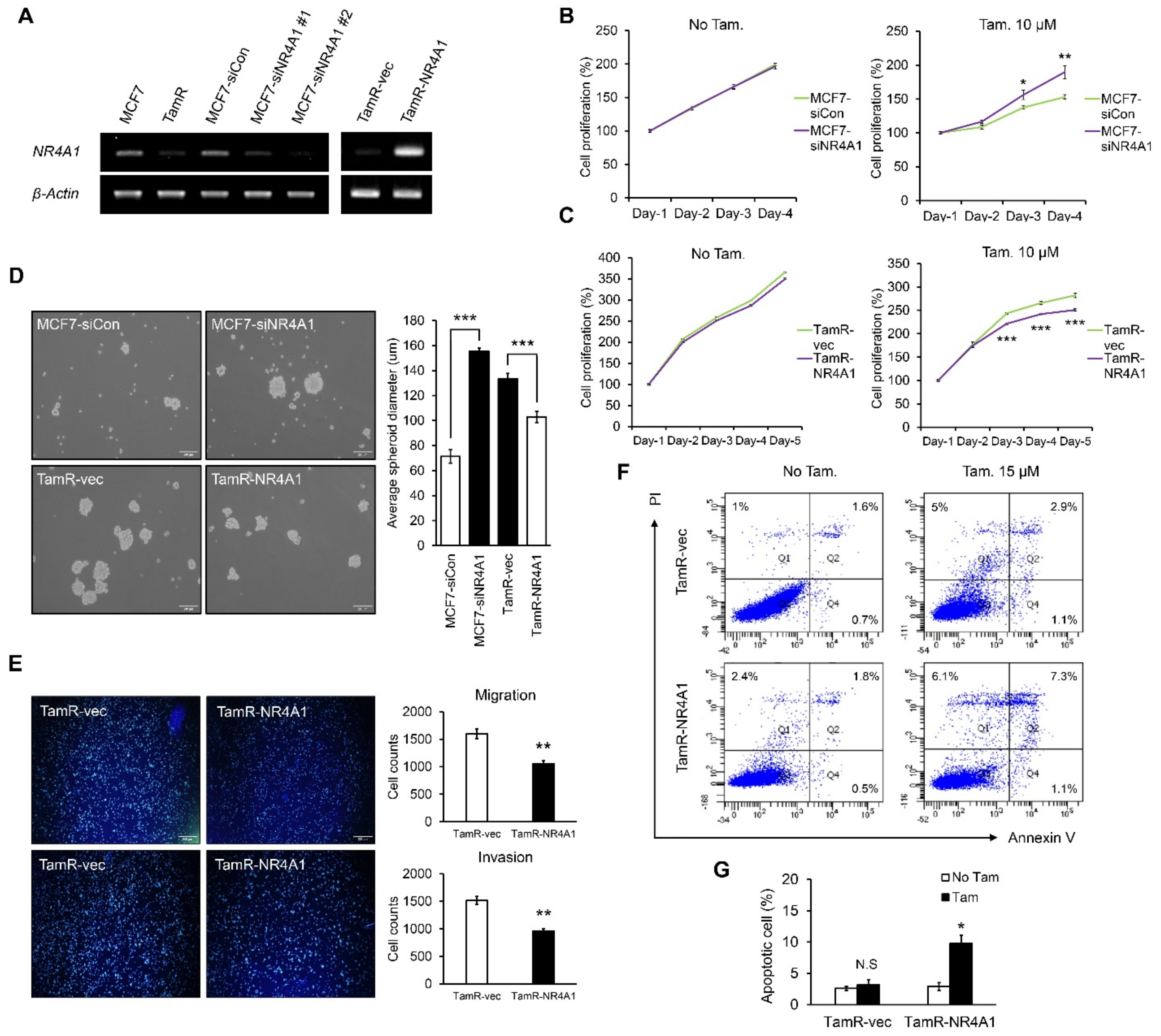
3.2. NR4A1 Suppresses Proliferation, Migration, and Invasion Ability, While Increasing Ttamoxifen-Induced Apoptosis in TamR Cells
3.3. NR4A1 Localized in the Cytoplasm Affects Apoptosis
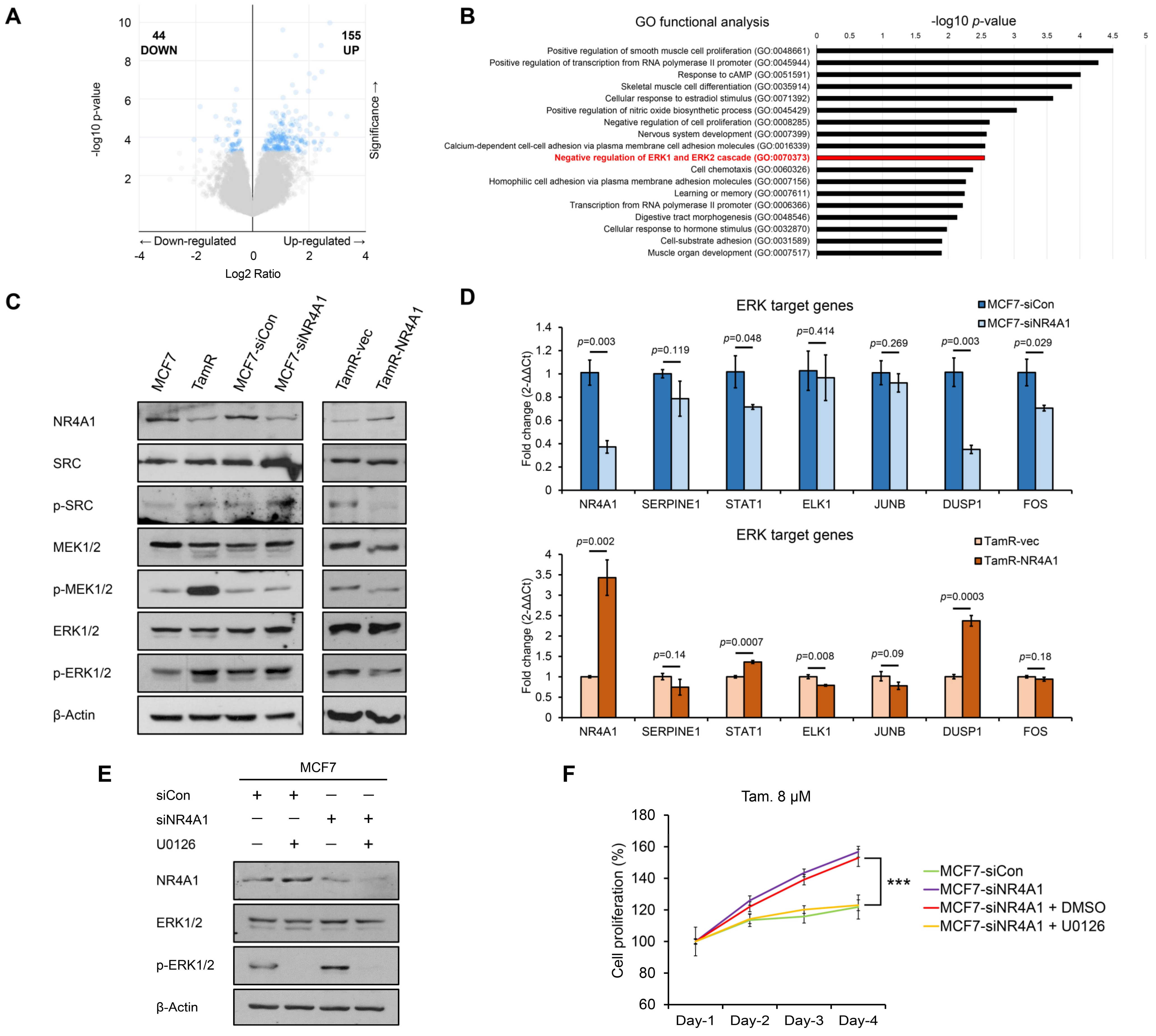
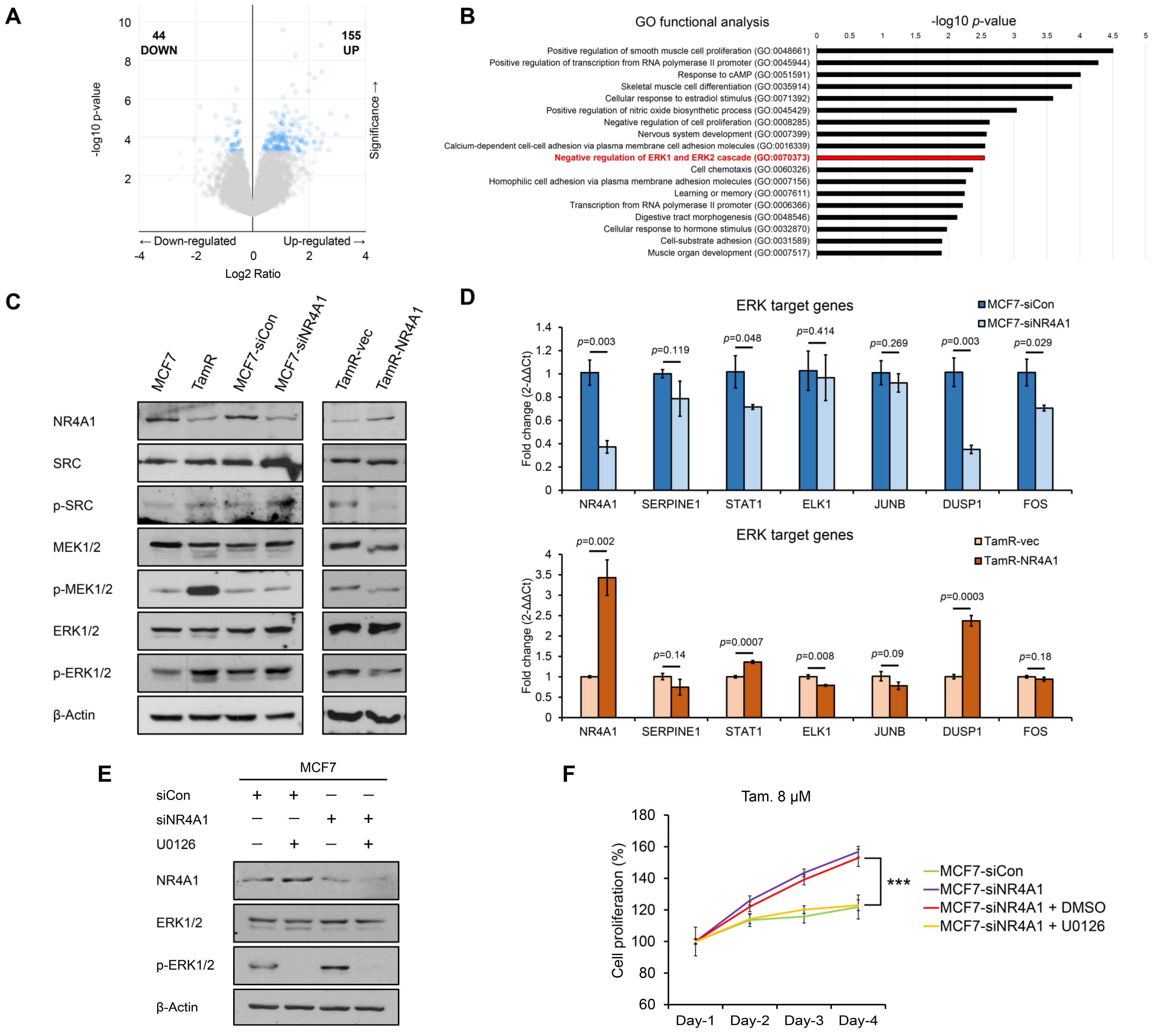
3.4. NR4A1 Alters Tamoxifen Sensitivity in ER-Positive Breast Cancer Cells by Suppressing ERK Signaling Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeSantis, C.E.; Ma, J.; Gaudet, M.M.; Newman, L.A.; Mph, K.D.M.; Sauer, A.G.; Jemal, A.; Siegel, R.L. Breast cancer statistics, 2019. CA A Cancer J. Clin. 2019, 69, 438–451. [Google Scholar] [CrossRef]
- Fragomeni, S.M.; Sciallis, A.; Jeruss, J.S. Molecular Subtypes and Local-Regional Control of Breast Cancer. Surg. Oncol. Clin. N. Am. 2018, 27, 95–120. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Coombes, R.C. Endocrine-responsive breast cancer and strategies for combating resistance. Nat. Rev. Cancer 2002, 2, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Lumachi, F.; Luisetto, G.; Basso, S.M.; Basso, U.; Brunello, A.; Camozzi, V. Endocrine Therapy of Breast Cancer. Curr. Med. Chem. 2011, 18, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Early Breast Cancer Trialists’ Collaborative Group (EBCTCG). Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15-year survival: An overview of the randomised trials. Lancet 2005, 365, 1687–1717. [Google Scholar] [CrossRef]
- Normanno, N.; Di Maio, M.; De Maio, E.; De Luca, A.; De Matteis, A.; Giordano, A.; Perrone, F. Mechanisms of endocrine resistance and novel therapeutic strategies in breast cancer. Endocrine-Related Cancer 2005, 12, 721–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herring, J.A.; Elison, W.S.; Tessem, J.S. Function of Nr4a Orphan Nuclear Receptors in Proliferation, Apoptosis and Fuel Utilization Across Tissues. Cells 2019, 8, 1373. [Google Scholar] [CrossRef] [Green Version]
- Mohan, H.M.; Aherne, C.M.; Rogers, A.; Baird, A.W.; Winter, D.C.; Murphy, E.P. Molecular Pathways: The Role of NR4A Orphan Nuclear Receptors in Cancer. Clin. Cancer Res. 2012, 18, 3223–3228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Wang, Q.; Liu, W.; Liu, F.; Ji, A.; Li, Y. The Orphan Nuclear Receptor 4A1: A Potential New Therapeutic Target for Metabolic Diseases. J. Diabetes Res. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [Green Version]
- McMorrow, J.; Murphy, E.P. Inflammation: A role for NR4A orphan nuclear receptors? Biochem. Soc. Trans. 2011, 39, 688–693. [Google Scholar] [CrossRef]
- Cho, S.D.; Yoon, K.; Chintharlapalli, S.; Abdelrahim, M.; Lei, P.; Hamilton, S.; Khan, S.; Ramaiah, S.K.; Safe, S. Nur77 Agonists Induce Proapoptotic Genes and Responses in Colon Cancer Cells through Nuclear Receptor–Dependent and Nuclear Receptor–Independent Pathways. Cancer Res. 2007, 67, 674–683. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.-X.; Chen, H.-Z.; Du, X.-D.; Luo, J.; He, J.-P.; Wang, R.-H.; Wang, Y.; Wu, R.; Hou, R.-R.; Hong, M.; et al. Orphan Receptor TR3 Enhances p53 Transactivation and Represses DNA Double-Strand Break Repair in Hepatoma Cells under Ionizing Radiation. Mol. Endocrinol. 2011, 25, 1337–1350. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, E.; Mohankumar, K.; Safe, S. TGFβ-Induced Lung Cancer Cell Migration Is NR4A1-Dependent. Mol. Cancer Res. 2018, 16, 1991–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-X.; Wang, Z.-J.; Zheng, Y.; Guan, Y.-F.; Yang, P.-B.; Chen, X.; Peng, C.; He, J.-P.; Ai, Y.-L.; Wu, S.-F.; et al. Nuclear Receptor Nur77 Facilitates Melanoma Cell Survival under Metabolic Stress by Protecting Fatty Acid Oxidation. Mol. Cell 2018, 69, 480–492.e7. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Herrick, A.M.; Mullican, S.E.; Sheehan, A.M.; Conneely, O.M. Reduced NR4A gene dosage leads to mixed myelodysplastic/myeloproliferative neoplasms in mice. Blood 2011, 117, 2681–2690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullican, S.E.; Zhang, S.; Konopleva, M.; Ruvolo, V.; Andreeff, M.; Milbrandt, J.; Conneely, O.M. Abrogation of nuclear receptors Nr4a3 andNr4a1 leads to development of acute myeloid leukemia. Nat. Med. 2007, 13, 730–735. [Google Scholar] [CrossRef]
- Crawford, P.A.; Sadovsky, Y.; Woodson, K.; Lee, S.L.; Milbrandt, J. Adrenocortical function and regulation of the steroid 21-hydroxylase gene in NGFI-B-deficient mice. Mol. Cell. Biol. 1995, 15, 4331–4336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Drabsch, Y.; Dekker, T.J.A.; De Vinuesa, A.G.; Li, Y.; Hawinkels, L.J.A.C.; Sheppard, K.-A.; Goumans, M.-J.; Luwor, R.B.; De Vries, C.J.; et al. Nuclear receptor NR4A1 promotes breast cancer invasion and metastasis by activating TGF-β signalling. Nat. Commun. 2014, 5, 3388. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.-F.; Wu, Q.; Liu, S.; Lin, X.-F.; Zhang, B.; Wu, J.-F.; Cai, J.-H.; Zhang, M.-Q.; Su, W.-J. Distinct role and functional mode of TR3 and RARα in mediating ATRA-induced signalling pathway in breast and gastric cancer cells. Int. J. Biochem. Cell Biol. 2004, 36, 98–113. [Google Scholar] [CrossRef]
- Wu, H.; Bi, J.; Peng, Y.; Huo, L.; Yu, X.; Yang, Z.; Zhou, Y.; Qin, L.; Xu, Y.; Liao, L.; et al. Nuclear receptor NR4A1 is a tumor suppressor down-regulated in triple-negative breast cancer. Oncotarget 2017, 8, 54364–54377. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]
- Yuan, J.; Dong, X.; Yap, J.; Hu, J. The MAPK and AMPK signalings: Interplay and implication in targeted cancer therapy. J. Hematol. Oncol. 2020, 13, 113. [Google Scholar] [CrossRef]
- Santen, R.J.; Song, R.X.; McPherson, R.; Kumar, R.; Adam, L.; Jeng, M.-H.; Yue, W. The role of mitogen-activated protein (MAP) kinase in breast cancer. J. Steroid Biochem. Mol. Biol. 2002, 80, 239–256. [Google Scholar] [CrossRef]
- Salaroglio, I.C.; Mungo, E.; Gazzano, E.; Kopecka, J.; Riganti, C. ERK is a Pivotal Player of Chemo-Immune-Resistance in Cancer. Int. J. Mol. Sci. 2019, 20, 2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeyinka, A.; Nui, Y.; Cherlet, T.; Snell, L.; Watson, P.; Murphy, L.C. Activated mitogen-activated protein kinase expression during human breast tumorigenesis and breast cancer progression. Clin. Cancer Res. 2002, 8, 1747–1753. [Google Scholar] [PubMed]
- Xie, F.; Li, Z.; Wang, N.; Fang, J.; Huang, J.; Tian, F.; Li, C. Role of PKC-ERK signaling in tamoxifen-induced apoptosis and tamoxifen resistance in human breast cancer cells. Oncol. Rep. 2012, 27, 1879–1886. [Google Scholar] [CrossRef] [PubMed]
- Mueller, H.; Flury, N.; Eppenberger-Castori, S.; Kueng, W.; Eppenberger, U. Potential prognostic value of mitogen-activated protein kinase activity for disease-free survival of primary breast cancer patients. Int. J. Cancer 2000, 89, 384–388. [Google Scholar] [CrossRef]
- Gee, J.M.; Robertson, J.F.; Ellis, I.O.; Nicholson, R.I. Phosphorylation of ERK1/2 mitogen-activated protein kinase is associated with poor response to anti-hormonal therapy and decreased patient survival in clinical breast cancer. Int. J. Cancer 2001, 95, 247–254. [Google Scholar] [CrossRef]
- Yang, S.; Lee, J.-Y.; Hur, H.; Oh, J.H.; Kim, M.H. Up-regulation of HOXB cluster genes are epigenetically regulated in tamoxifen-resistant MCF7 breast cancer cells. BMB Rep. 2018, 51, 450–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iglesias, J.M.; Beloqui, I.; Garcia-Garcia, F.; Leis, O.; Vázquez-Martín, A.; Eguiara, A.; Cufí, S.; Pavon, A.; Menendez, J.A.; Dopazo, J.; et al. Mammosphere Formation in Breast Carcinoma Cell Lines Depends upon Expression of E-cadherin. PLoS ONE 2013, 8, e77281. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.-Y.; Hur, H.; Yun, H.J.; Kim, Y.; Yang, S.; Kim, S.I.; Kim, M.H. HOXB5 Promotes the Proliferation and Invasion of Breast Cancer Cells. Int. J. Biol. Sci. 2015, 11, 701–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooding, A.J.; Schiemann, W.P. Epithelial–Mesenchymal Transition Programs and Cancer Stem Cell Phenotypes: Mediators of Breast Cancer Therapy Resistance. Mol. Cancer Res. 2020, 18, 1257–1270. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, A.J.A.; Rinner, B.; Wenzl, K.; Pichler, M.; Troppan, K.; Steinbauer, E.; Schwarzenbacher, D.; Reitter, S.; Feichtinger, J.; Tierling, S.; et al. NR4A1-mediated apoptosis suppresses lymphomagenesis and is associated with a favorable cancer-specific survival in patients with aggressive B-cell lymphomas. Blood 2014, 123, 2367–2377. [Google Scholar] [CrossRef]
- Kolluri, S.K.; Bruey-Sedano, N.; Cao, X.; Lin, B.; Lin, F.; Han, Y.-H.; Dawson, M.I.; Zhang, X.-K. Mitogenic Effect of Orphan Receptor TR3 and Its Regulation by MEKK1 in Lung Cancer Cells. Mol. Cell. Biol. 2003, 23, 8651–8667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, B.; Kolluri, S.K.; Lin, F.; Liu, W.; Han, Y.-H.; Cao, X.; Dawson, M.I.; Reed, J.C.; Zhang, X.-K. Conversion of Bcl-2 from Protector to Killer by Interaction with Nuclear Orphan Receptor Nur77/TR3. Cell 2004, 116, 527–540. [Google Scholar] [CrossRef] [Green Version]
- Fechter, K.; Feichtinger, J.; Prochazka, K.; Unterluggauer, J.J.; Pansy, K.; Steinbauer, E.; Pichler, M.; Haybaeck, J.; Prokesch, A.; Greinix, H.T.; et al. Cytoplasmic location of NR4A1 in aggressive lymphomas is associated with a favourable cancer specific survival. Sci. Rep. 2018, 8, 14528. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Yang, J.-R.; Jia, Y.; Zhang, P.; Shen, L.; Li, X.-L.; Li, J.; Wang, B. Overexpression of NR4A1 is associated with tumor recurrence and poor survival in non-small-cell lung carcinoma. Oncotarget 2017, 8, 113977–113986. [Google Scholar] [CrossRef]
- Yan, H.; Xiao, F.; Zou, J.; Qiu, C.; Sun, W.; Gu, M.; Zhang, L. NR4A1-induced increase in the sensitivity of a human gastric cancer line to TNFα-mediated apoptosis is associated with the inhibition of JNK/Parkin-dependent mitophagy. Int. J. Oncol. 2017, 52, 367–378. [Google Scholar] [CrossRef]
- Wilson, A.J.; Liu, A.Y.; Roland, J.; Adebayo, O.B.; Fletcher, S.A.; Slaughter, J.C.; Saskowski, J.; Crispens, M.A.; Jones, H.W.; James, S.; et al. TR3 Modulates Platinum Resistance in Ovarian Cancer. Cancer Res. 2013, 73, 4758–4769. [Google Scholar] [CrossRef] [Green Version]
- Alexopoulou, A.N.; Leao, M.; Caballero, O.L.; Da Silva, L.; Reid, L.; Lakhani, S.R.; Simpson, A.J.; Marshall, J.F.; Neville, A.M.; Jat, P.S. Dissecting the transcriptional networks underlying breast cancer: NR4A1 reduces the migration of normal and breast cancer cell lines. Breast Cancer Res. 2010, 12, R51. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, S. Protein mislocalization: Mechanisms, functions and clinical applications in cancer. Biochim. Biophys. Acta Bioenergy 2014, 1846, 13–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Yuan, L.; Yu, W.; Sun, Y.; Jiang, D.; Wang, X.; Feng, X.; Wang, Z.; Xu, J.; Yang, R.; et al. A Regulation Loop between YAP and NR4A1 Balances Cell Proliferation and Apoptosis. Cell Rep. 2020, 33, 108284. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Du, X.; Chen, H.; Liu, J.; Zhao, B.; Huang, D.; Li, G.; Xu, Q.; Zhang, M.; Weimer, B.C.; et al. Cytosporone B is an agonist for nuclear orphan receptor Nur77. Nat. Chem. Biol. 2008, 4, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Abdelrahim, M.; Cho, S.D.; Liu, X.; Liu, X.; Safe, S. Structure-dependent activation of endoplasmic reticulum stress-mediated apoptosis in pancreatic cancer by 1,1-bis(3′-indoly)-1-(p-substituted phenyl)methanes. Mol. Cancer Ther. 2008, 7, 3363–3372. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.C.; Kim, C.Y.; Oh, J.H.; Kim, M.H. NR4A1 Regulates Tamoxifen Resistance by Suppressing ERK Signaling in ER-Positive Breast Cancer. Cells 2021, 10, 1633. https://doi.org/10.3390/cells10071633
Kim YC, Kim CY, Oh JH, Kim MH. NR4A1 Regulates Tamoxifen Resistance by Suppressing ERK Signaling in ER-Positive Breast Cancer. Cells. 2021; 10(7):1633. https://doi.org/10.3390/cells10071633
Chicago/Turabian StyleKim, Yu Cheon, Clara Yuri Kim, Ji Hoon Oh, and Myoung Hee Kim. 2021. "NR4A1 Regulates Tamoxifen Resistance by Suppressing ERK Signaling in ER-Positive Breast Cancer" Cells 10, no. 7: 1633. https://doi.org/10.3390/cells10071633
APA StyleKim, Y. C., Kim, C. Y., Oh, J. H., & Kim, M. H. (2021). NR4A1 Regulates Tamoxifen Resistance by Suppressing ERK Signaling in ER-Positive Breast Cancer. Cells, 10(7), 1633. https://doi.org/10.3390/cells10071633