
Bone Marrow-Derived VSELs Engraft as Lung Epithelial Progenitor Cells after Bleomycin-Induced Lung Injury

 , , ,
, , ,
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Mice
2.2. Isolation of Murine VSELs and Non-VSELs/Nonhematopoietic Stem Cells by FACS
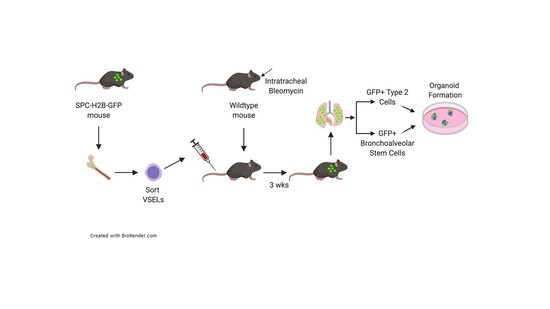
2.3. Intravenous Transplantation of BM Cells Following Intratracheal Bleomycin Administration
2.4. Lung Harvest and Lung Single Cell Suspension
2.5. Isolation of Murine Alveolar Type II Cells and Bronchioalveolar Stem Cells by FACS and Flow Cytometry Analysis
2.6. Lung Tissue Paraffin-Embedded Sections
2.7. Lung Organoid Model System
2.8. Immunofluorescence Staining on Lung Tissue Sections and Organoids
2.9. qRT-PCR for GFP Expression
2.10. Statistical Analysis
3. Results
3.1. Transplantation of VSELs Versus Non-VSEL/Non-HSCs Cells into Bleomycin-Induced Lung Injured Mice
3.2. Detection of GFP in Lungs of Mice Receiving VSELs Versus HSPCs
3.3. Detection of GFP+ Alveolar type II Cells Derived from VSELs by Confocal Microscopy on Tissue Sections
3.4. Detection of GFP+AT2 Cells and BASC in the Lung Using Flow Cytometry
3.5. Detection of GFP+ Epithelial Contribution of Transplanted VSELs Versus Non-VSEL/Non-HSCs in Organoids
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fritz, H.; Flower, G.; Weeks, L.; Cooley, K.; Callachan, M.; McGowan, J.; Skidmore, B.; Kirchner, L.; Seely, D. Intravenous Vitamin C and Cancer: A Systematic Review. Integr. Cancer Ther. 2014, 13, 280–300. [Google Scholar] [CrossRef] [PubMed]
- Basil, M.C.; E Morrisey, E. BASC-ing in the glow: Bronchioalveolar stem cells get their place in the lung. EMBO J. 2019, 38, 38. [Google Scholar] [CrossRef] [PubMed]
- Giangreco, A.; Reynolds, S.D.; Stripp, B.R. Terminal Bronchioles Harbor a Unique Airway Stem Cell Population That Localizes to the Bronchoalveolar Duct Junction. Am. J. Pathol. 2002, 161, 173–182. [Google Scholar] [CrossRef]
- Kim, C.F.; Jackson, E.L.; Woolfenden, A.E.; Lawrence, S.; Babar, I.; Vogel, S.; Crowley, D.; Bronson, R.T.; Jacks, T. Identification of Bronchioalveolar Stem Cells in Normal Lung and Lung Cancer. Cell 2005, 121, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Tropea, K.A.; Leder, E.; Aslam, M.; Lau, A.; Raiser, D.M.; Lee, J.-H.; Balasubramaniam, V.; Fredenburgh, L.E.; Mitsialis, S.A.; Kourembanas, S.; et al. Bronchioalveolar stem cells increase after mesenchymal stromal cell treatment in a mouse model of bronchopulmonary dysplasia. Am. J. Physiol. Cell. Mol. Physiol. 2012, 302, L829–L837. [Google Scholar] [CrossRef] [PubMed]
- Hogan, B.L.; Barkauskas, C.E.; Chapman, H.A.; Epstein, J.A.; Jain, R.; Hsia, C.C.; Niklason, L.; Calle, E.; Le, A.; Randell, S.H.; et al. Repair and Regeneration of the Respiratory System: Complexity, Plasticity, and Mechanisms of Lung Stem Cell Function. Cell Stem Cell 2014, 15, 123–138. [Google Scholar] [CrossRef]
- Liu, Q.; Liu, K.; Cui, G.; Huang, X.; Yao, S.; Guo, W.; Qin, Z.; Li, Y.; Yang, R.; Pu, W.; et al. Lung regeneration by multipotent stem cells residing at the bronchioalveolar-duct junction. Nat. Genet. 2019, 51, 728–738. [Google Scholar] [CrossRef]
- Barkauskas, C.E.; Cronce, M.J.; Rackley, C.R.; Bowie, E.; Keene, D.R.; Stripp, B.R.; Randell, S.H.; Noble, P.W.; Hogan, B.L. Type 2 alveolar cells are stem cells in adult lung. J. Clin. Investig. 2013, 123, 3025–3036. [Google Scholar] [CrossRef]
- Salwig, I.; Spitznagel, B.; Vazquez-Armendariz, A.I.; Khalooghi, K.; Guenther, S.; Herold, S.; Szibor, M.; Braun, T. Bronchioalveolar stem cells are a main source for regeneration of distal lung epithelia in vivo. EMBO J. 2019, 38, 38. [Google Scholar] [CrossRef]
- Kassmer, S.H.; Jin, H.; Zhang, P.-X.; Bruscia, E.M.; Heydari, K.; Lee, J.-H.; Kim, C.F.; Krause, D.S.; Krouse, D. Very Small Embryonic-Like Stem Cells from the Murine Bone Marrow Differentiate into Epithelial Cells of the Lung. Stem Cells 2013, 31, 2759–2766. [Google Scholar] [CrossRef]
- Szade, K.; Bukowska-Strakova, K.; Nowak, W.N.; Szade, A.; Kachamakova-Trojanowska, N.; Zukowska, M.; Jozkowicz, A.; Dulak, J. Murine Bone Marrow Lin−Sca-1+CD45− Very Small Embryonic-Like (VSEL) Cells Are Heterogeneous Population Lacking Oct-4A Expression. PLoS ONE 2013, 8, e63329. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Zuba-Surma, E.; Kucia, M.; Poniewierska, A.; Suszynska, M.; Ratajczak, J. Pluripotent and multipotent stem cells in adult tissues. Adv. Med Sci. 2012, 57, 1–17. [Google Scholar] [CrossRef]
- Kucia, M.; Reca, R.; Campbell, F.R.; Zuba-Surma, E.; Majka, M.; Ratajczak, J.; Ratajczak, M.Z. A population of very small embryonic-like (VSEL) CXCR4+SSEA-1+Oct-4+ stem cells identified in adult bone marrow. Leukemia 2006, 20, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Kucia, M.; Zuba-Surma, E.; Wysoczynski, M.; Dobrowolska, H.; Reca, R.; Ratajczak, J.; Ratajczak, M.Z. Physiological and pathological consequences of identification of very small embryonic like (VSEL) stem cells in adult bone marrow. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2006, 57, 5–18. [Google Scholar]
- Ratajczak, M.Z.; Zuba-Surma, E.; Shin, D.-M.; Ratajczak, J.; Kucia, M. Very small embryonic-like (VSEL) stem cells in adult organs and their potential role in rejuvenation of tissues and longevity. Exp. Gerontol. 2008, 43, 1009–1017. [Google Scholar] [CrossRef]
- Hałasa, M.; Baśkiewicz-Masiuk, M.; Dąbkowska, E.; Machalinski, B. An efficient two-step method to purify very small embryonic-like (VSEL) stem cells from umbilical cord blood (UCB). Folia Histochem. Cytobiol. 2008, 46, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.; Imberti, B.; Bianchi, N.; Pezzotta, A.; Morigi, M.; Del Fante, C.; Redi, C.A.; Perotti, C. A Novel Method for Isolation of Pluripotent Stem Cells from Human Umbilical Cord Blood. Stem Cells Dev. 2017, 26, 1258–1269. [Google Scholar] [CrossRef] [PubMed]
- Sovalat, H.; Scrofani, M.; Eidenschenk, A.; Pasquet, S.; Rimelen, V.; Hénon, P. Identification and isolation from either adult human bone marrow or G-CSF-mobilized peripheral blood of CD34+/CD133+/CXCR4+/Lin−CD45− cells, featuring morphological, molecular, and phenotypic characteristics of very small embryonic-like (VSEL) stem cells. Exp. Hematol. 2011, 39, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Gounari, E.L.E.N.I.; Daniilidis, A.; Tsagias, N.; Michopoulou, A.; Kouzi, K.; Koliakos, G. Isolation of a novel embryonic stem cell cord blood-derived population with in vitro hematopoietic capacity in the presence of Wharton’s jelly-derived mesenchymal stromal cells. Cytotherapy 2019, 21, 246–259. [Google Scholar] [CrossRef]
- Kassmer, S.H.; Bruscia, E.M.; Zhang, P.-X.; Krause, D.S. Nonhematopoietic Cells are the Primary Source of Bone Marrow-Derived Lung Epithelial Cells. Stem Cells 2011, 30, 491–499. [Google Scholar] [CrossRef]
- Barkauskas, C.E.; Chung, M.-I.; Fioret, B.; Gao, X.; Katsura, H.; Hogan, B.L.M. Lung organoids: Current uses and future promise. Development 2017, 144, 986–997. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kim, J.; Gludish, D.; Roach, R.R.; Saunders, A.H.; Barrios, J.; Woo, A.; Chen, H.; Conner, D.A.; Fujiwara, Y.; et al. Surfactant Protein–C Chromatin-Bound Green Fluorescence Protein Reporter Mice Reveal Heterogeneity of Surfactant Protein C–Expressing Lung Cells. Am. J. Respir. Cell Mol. Biol. 2013, 48, 288–298. [Google Scholar] [CrossRef]
- Suszyńska, M.; Zuba-Surma, E.K.; Maj, M.; Mierzejewska, K.; Ratajczak, J.; Kucia, M.; Ratajczak, M.Z. The Proper Criteria for Identification and Sorting of Very Small Embryonic-Like Stem Cells, and Some Nomenclature Issues. Stem Cells Dev. 2014, 23, 702–713. [Google Scholar] [CrossRef]
- Zuba-Surma, E.K.; Klich, I.; Greco, N.; Laughlin, M.J.; Ratajczak, J.; Ratajczak, M.Z. Optimization of isolation and further characterization of umbilical cord blood-derived very small embryonic/ epiblast-like stem cells (VSELs). Eur. J. Haematol. 2010, 84, 34–46. [Google Scholar] [CrossRef]
- Zuba-Surma, E.K.; Ratajczak, M.Z. Overview of Very Small Embryonic-Like Stem Cells (VSELs) and Methodology of Their Identification and Isolation by Flow Cytometric Methods. Curr. Protoc. Cytom. 2010, 51, 9.29.1–9.29.15. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, L.; Zuba-Surma, E.K.; Peng, X.; Kucia, M.; Ratajczak, M.Z.; Wang, W.; Enzman, V.; Kaplan, H.J.; Dean, D.C. Identification of small Sca-1+, Lin−, CD45− multipotential cells in the neonatal murine retina. Exp. Hematol. 2009, 37, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Herzog, E.L.; Van Arnam, J.; Hu, B.; Krause, D.S. Threshold of Lung Injury Required for the Appearance of Marrow-Derived Lung Epithelia. Stem Cells 2006, 24, 1986–1992. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Ciechanowicz, A.; Kaplan, A.R.; Wang, L.; Zhang, P.-X.; Lu, Y.-C.; Tobin, R.E.; Tobin, B.A.; Cohn, L.; Zeiss, C.J.; et al. Surfactant protein C dampens inflammation by decreasing JAK/STAT activation during lung repair. Am. J. Physiol. Cell. Mol. Physiol. 2018, 314, L882–L892. [Google Scholar] [CrossRef]
- McQualter, J.L.; Yuen, K.; Williams, B.; Bertoncello, I. Evidence of an epithelial stem/progenitor cell hierarchy in the adult mouse lung. Proc. Natl. Acad. Sci. USA 2010, 107, 1414–1419. [Google Scholar] [CrossRef]
- Teisanu, R.M.; Chen, H.; Matsumoto, K.; McQualter, J.L.; Potts, E.; Foster, W.M.; Bertoncello, I.; Stripp, B.R. Functional Analysis of Two Distinct Bronchiolar Progenitors during Lung Injury and Repair. Am. J. Respir. Cell Mol. Biol. 2011, 44, 794–803. [Google Scholar] [CrossRef]
- Izbicki, G.; Segel, M.J.; Christensen, T.G.; Conner, M.W.; Breuer, R. Time course of bleomycin-induced lung fibrosis. Int. J. Exp. Pathol. 2002, 83, 111–119. [Google Scholar] [CrossRef]
- Viksman, M.Y.; Liu, M.C.; Schleimer, R.P.; Bochner, B.S. Application of a flow cytometric method using autofluorescence and a tandem fluorescent dye to analyze human alveolar macrophage surface markers. J. Immunol. Methods 1994, 172, 17–24. [Google Scholar] [CrossRef]
- Pataki, G.; Czopf, L.; A Holm, B.; Matalon, S. Quantitation of the alveolar distribution of surfactant mixtures in normal and injured lungs. Am. J. Respir. Cell Mol. Biol. 1996, 15, 451–459. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Zuba-Surma, E.; Wojakowski, W.; Suszynska, M.; Mierzejewska, K.; Liu, R.; Ratajczak, J.; Shin, D.M.; Kucia, M. Very small embryonic-like stem cells (VSELs) represent a real challenge in stem cell biology: Recent pros and cons in the midst of a lively debate. Leukemia 2013, 28, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Desai, T.J.; Brownfield, D.G.; Krasnow, M.A. Alveolar progenitor and stem cells in lung development, renewal and cancer. Nat. Cell Biol. 2014, 507, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Nadkarni, R.R.; Abed, S.; Draper, J.S. Organoids as a model system for studying human lung development and disease. Biochem. Biophys. Res. Commun. 2016, 473, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Choi, K.M.; Sicard, D.; Tschumperlin, D.J. Human airway organoid engineering as a step toward lung regeneration and disease modeling. Biomaterials 2017, 113, 118–132. [Google Scholar] [CrossRef]
- Miyanishi, M.; Mori, Y.; Seita, J.; Chen, J.Y.; Karten, S.; Chan, C.K.; Nakauchi, H.; Weissman, I.L. Do Pluripotent Stem Cells Exist in Adult Mice as Very Small Embryonic Stem Cells? Stem Cell Rep. 2013, 1, 198–208. [Google Scholar] [CrossRef]
- Chen, Z.-H.; Lv, X.; Dai, H.; Liu, C.; Lou, D.; Chen, R.; Zou, G.-M. Hepatic Regenerative Potential of Mouse Bone Marrow Very Small Embryonic-Like Stem Cells. J. Cell. Physiol. 2015, 230, 1852–1861. [Google Scholar] [CrossRef]
- Golipoor, Z.; Mehraein, F.; Zafari, F.; Alizadeh, A.; Ababzadeh, S.; Baazm, M. Migration of Bone Marrow-Derived Very Small Embryonic-Like Stem Cells toward An Injured Spinal Cord. Cell J. 2016, 17, 639–647. [Google Scholar]
- Guerin, C.L.; Loyer, X.; Vilar, J.; Cras, A.; Mirault, T.; Gaussem, P.; Silvestre, J.-S.; Smadja, D.M. Bone-marrow-derived very small embryonic-like stem cells in patients with critical leg ischaemia: Evidence of vasculogenic potential. Thromb. Haemost. 2015, 113, 1084–1094. [Google Scholar] [CrossRef] [PubMed]
- Lahlil, R.; Scrofani, M.; Barbet, R.; Tancrédi, C.; Aries, A.; Hénon, P. VSELs Maintain their Pluripotency and Competence to Differentiate after Enhanced Ex Vivo Expansion. Stem Cell Rev. Rep. 2018, 14, 510–524. [Google Scholar] [CrossRef]
- Shaikh, A.; Nagvenkar, P.; Pethe, P.; Hinduja, I.; Bhartiya, D. Molecular and phenotypic characterization of CD133 and SSEA4 enriched very small embryonic-like stem cells in human cord blood. Leukemia 2015, 29, 1909–1917. [Google Scholar] [CrossRef]
- Wang, J.; Guo, X.; Lui, M.; Chu, P.-J.; Yoo, J.; Chang, M.; Yen, Y. Identification of a Distinct Small Cell Population from Human Bone Marrow Reveals Its Multipotency In Vivo and In Vitro. PLoS ONE 2014, 9, e85112. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, L.; Wang, J.; Chen, N.; Yan, J.; Pan, X. HIF-2α and Oct4 have synergistic effects on survival and myocardial repair of very small embryonic-like mesenchymal stem cells in infarcted hearts. Cell Death Dis. 2018, 8, e2548. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciechanowicz, A.K.; Sielatycka, K.; Cymer, M.; Skoda, M.; Suszyńska, M.; Bujko, K.; Ratajczak, M.Z.; Krause, D.S.; Kucia, M. Bone Marrow-Derived VSELs Engraft as Lung Epithelial Progenitor Cells after Bleomycin-Induced Lung Injury. Cells 2021, 10, 1570. https://doi.org/10.3390/cells10071570
Ciechanowicz AK, Sielatycka K, Cymer M, Skoda M, Suszyńska M, Bujko K, Ratajczak MZ, Krause DS, Kucia M. Bone Marrow-Derived VSELs Engraft as Lung Epithelial Progenitor Cells after Bleomycin-Induced Lung Injury. Cells. 2021; 10(7):1570. https://doi.org/10.3390/cells10071570
Chicago/Turabian StyleCiechanowicz, Andrzej K., Katarzyna Sielatycka, Monika Cymer, Marta Skoda, Malwina Suszyńska, Kamila Bujko, Mariusz Z. Ratajczak, Diane S. Krause, and Magdalena Kucia. 2021. "Bone Marrow-Derived VSELs Engraft as Lung Epithelial Progenitor Cells after Bleomycin-Induced Lung Injury" Cells 10, no. 7: 1570. https://doi.org/10.3390/cells10071570
APA StyleCiechanowicz, A. K., Sielatycka, K., Cymer, M., Skoda, M., Suszyńska, M., Bujko, K., Ratajczak, M. Z., Krause, D. S., & Kucia, M. (2021). Bone Marrow-Derived VSELs Engraft as Lung Epithelial Progenitor Cells after Bleomycin-Induced Lung Injury. Cells, 10(7), 1570. https://doi.org/10.3390/cells10071570