
Neurogenic and Neuroprotective Potential of Stem/Stromal Cells Derived from Adipose Tissue

 , , and
, , and
Abstract
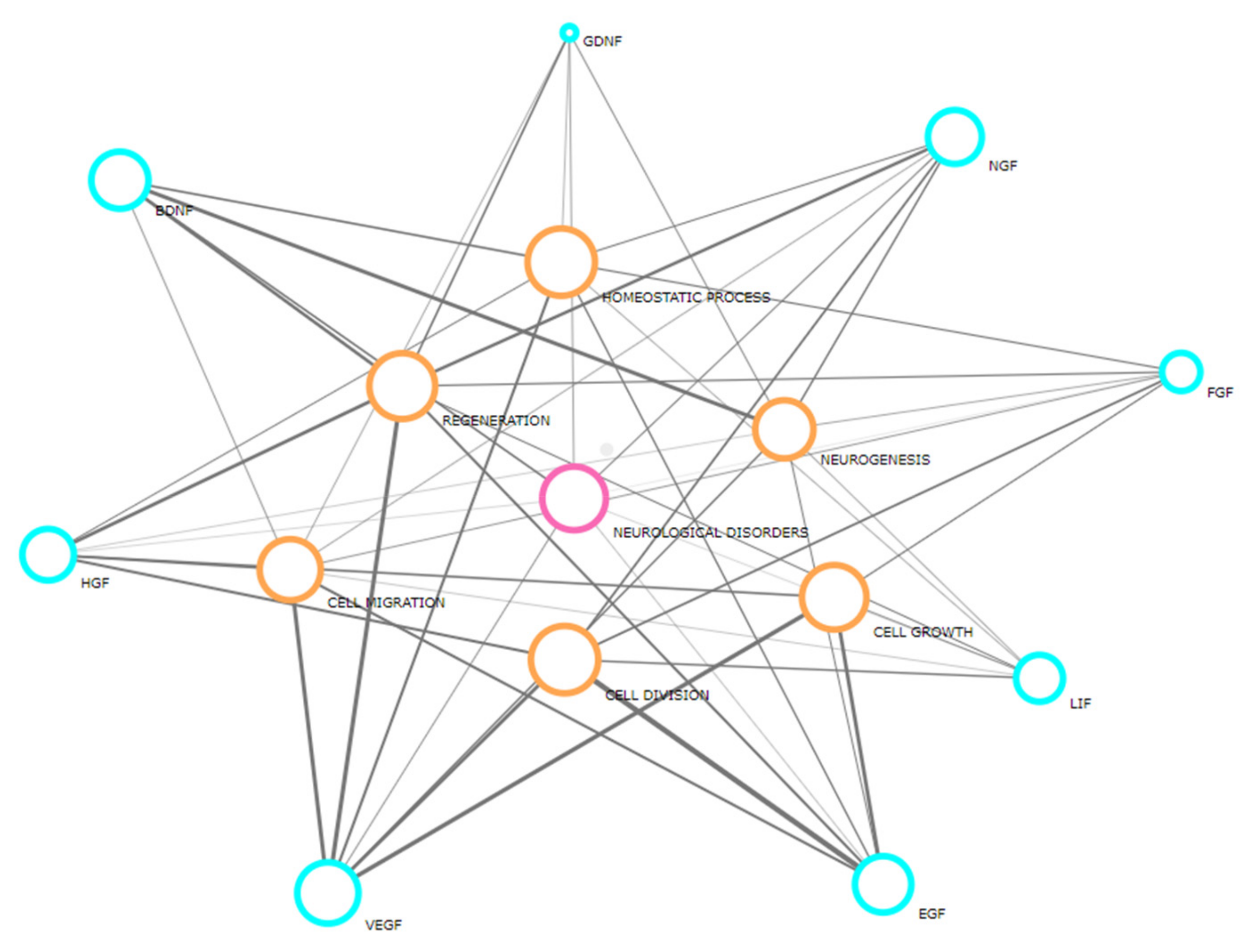
1. Introduction
2. Materials and Methods
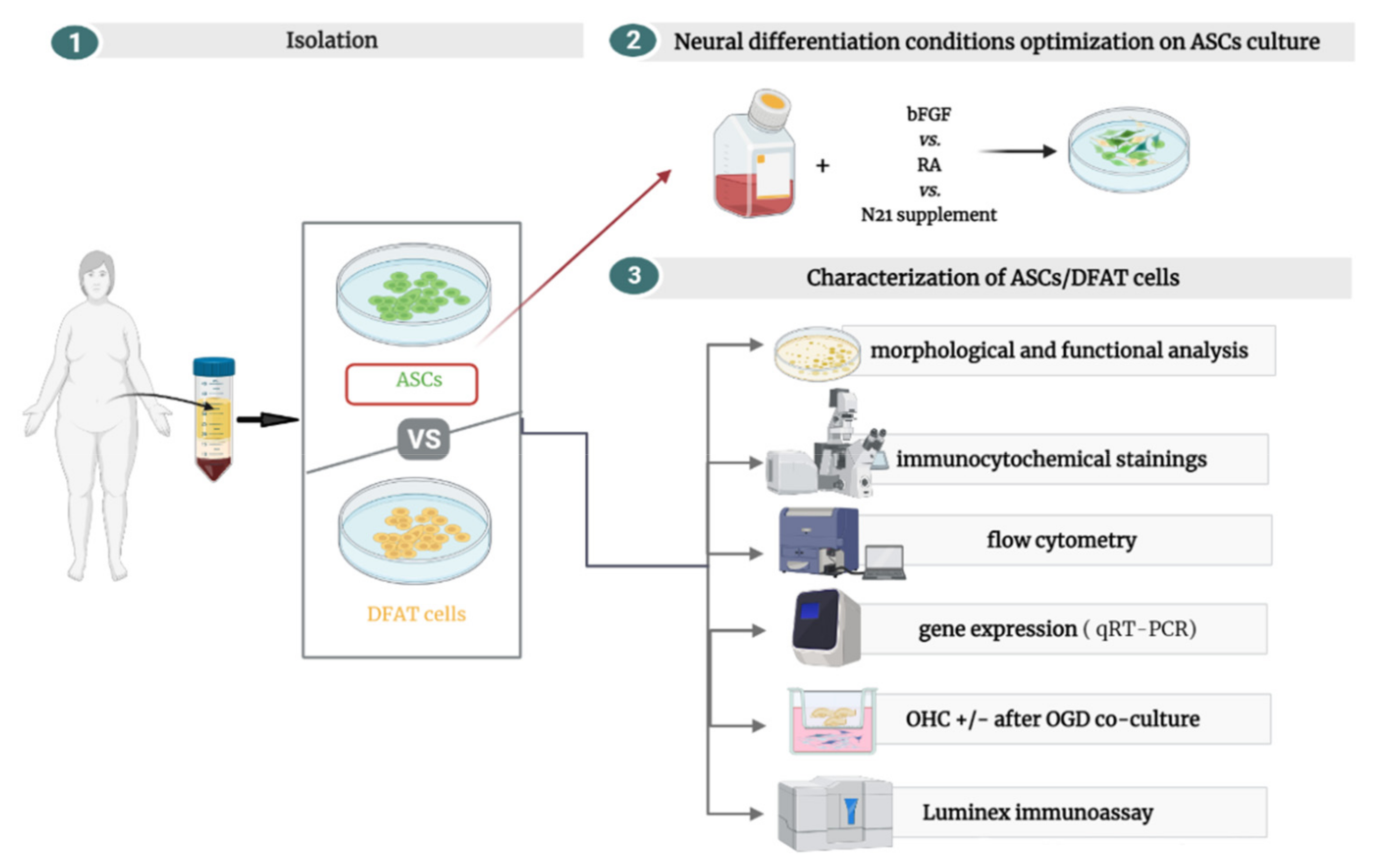
2.1. Cell Isolation
2.2. Cell Culture
2.3. Flow Cytometry Analysis
2.4. Mesodermal Lineage Differentiation
2.5. Cell Proliferation Analysis
2.6. Senescence Assay
2.7. CFU-F Assay
2.8. Neural Lineage Differentiation
- cell growth medium with Human Platelet Lysate concentration decreased to 5% and with the addition of bFGF (0.1%; Gibco) for 21 days,
- cell growth medium with Human Platelet Lysate concentration decreased to 5% and with the addition of bFGF (0.1%; Gibco) for 10 days, then with the addition of retinoic acid (RA; Sigma-Aldrich) for the next 15 days,
- cell growth medium with the addition of N21 supplement (1:49; Sigma-Aldrich) for 21 days.
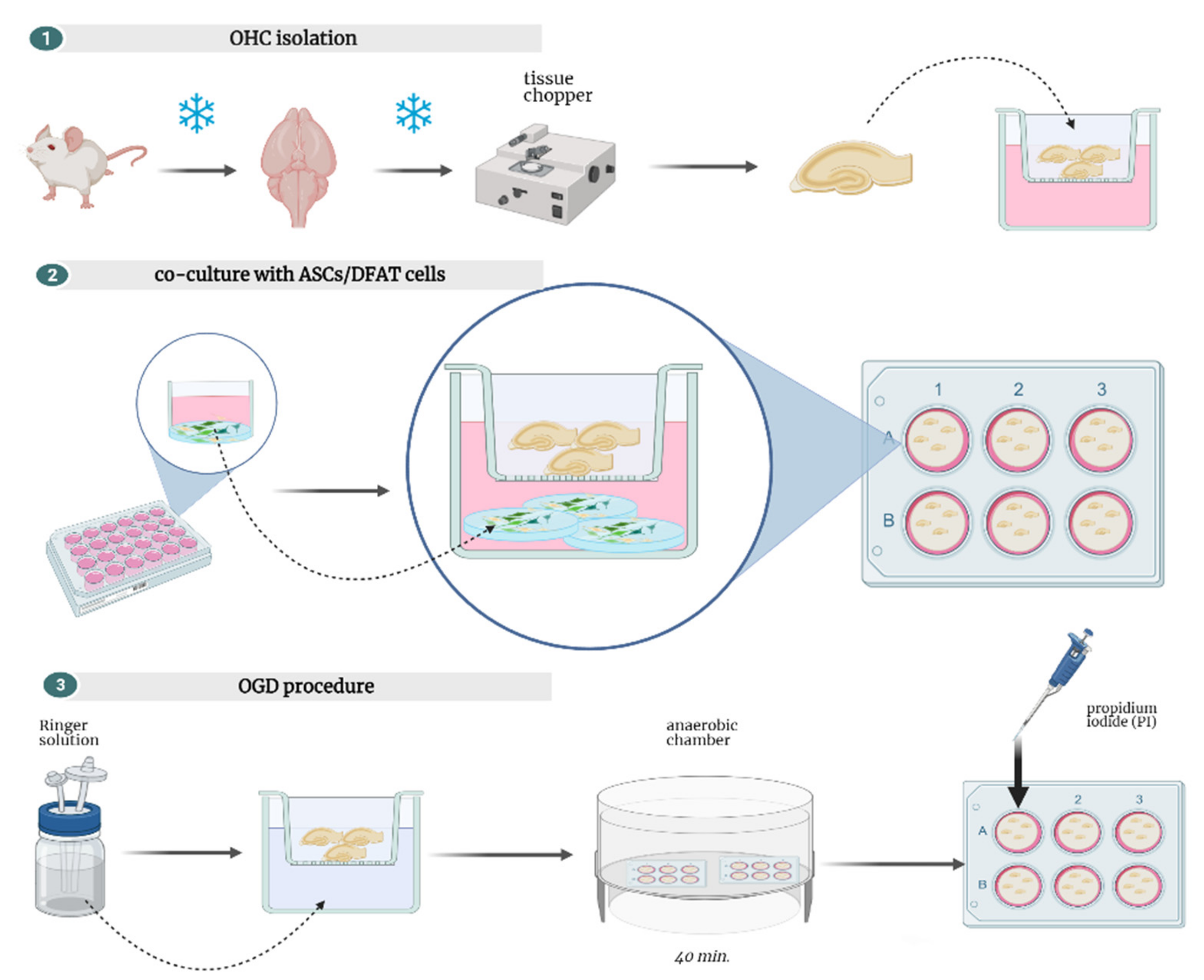
2.9. Organotypic Hippocampal Slices Culture (OHC)
2.10. Oxygen-Glucose-Deprivation (OGD)
2.11. Immunocytochemistry
2.12. Three Germ Layer Differentiation Potential
2.13. Quantitative RT-PCR Analysis
2.14. Cytokine and Chemokine Assays with Luminex Kit
2.15. Statistical Analysis
3. Results
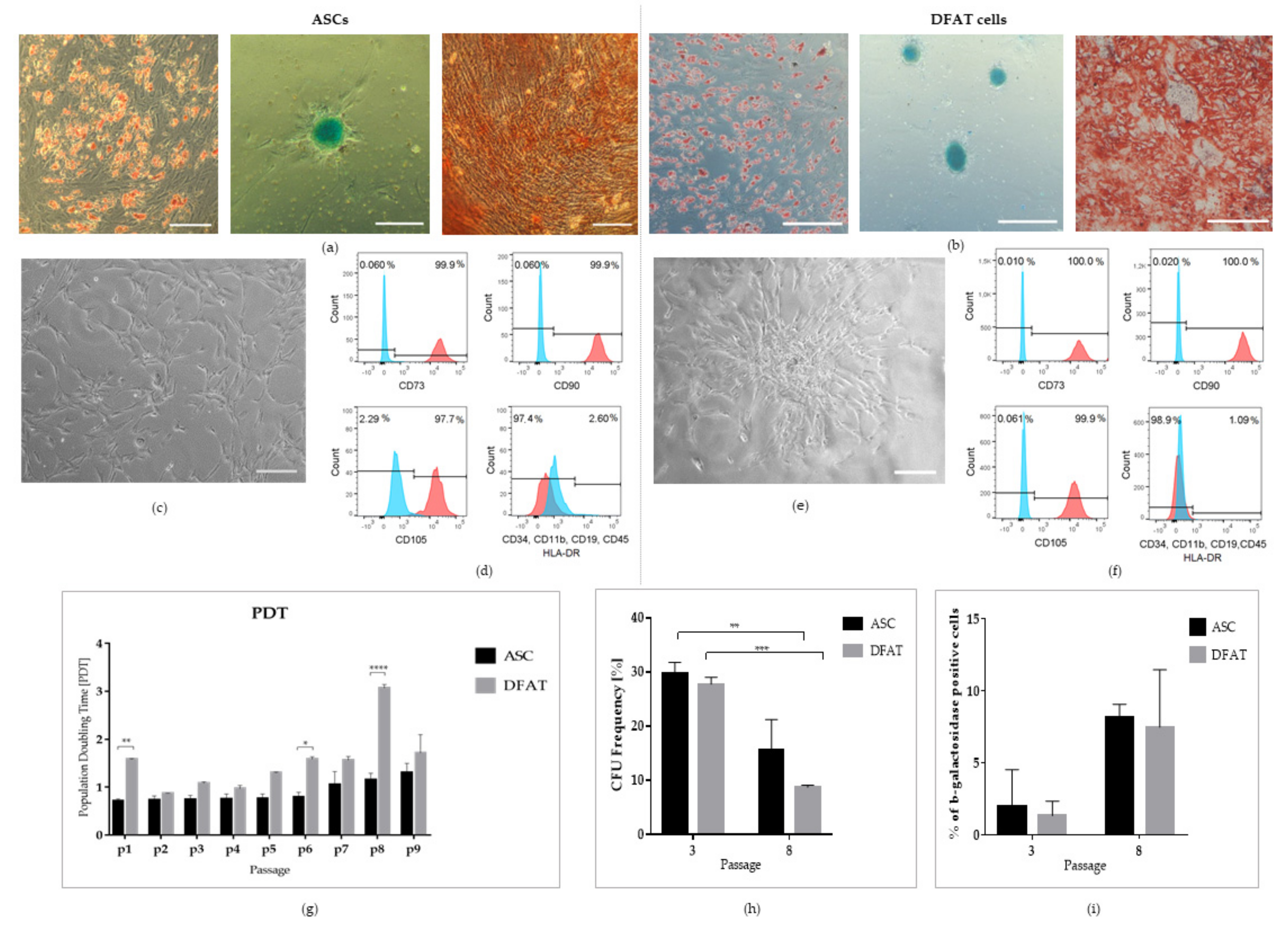
3.1. Multipotent Properties of Mesenchymal Stem Cells
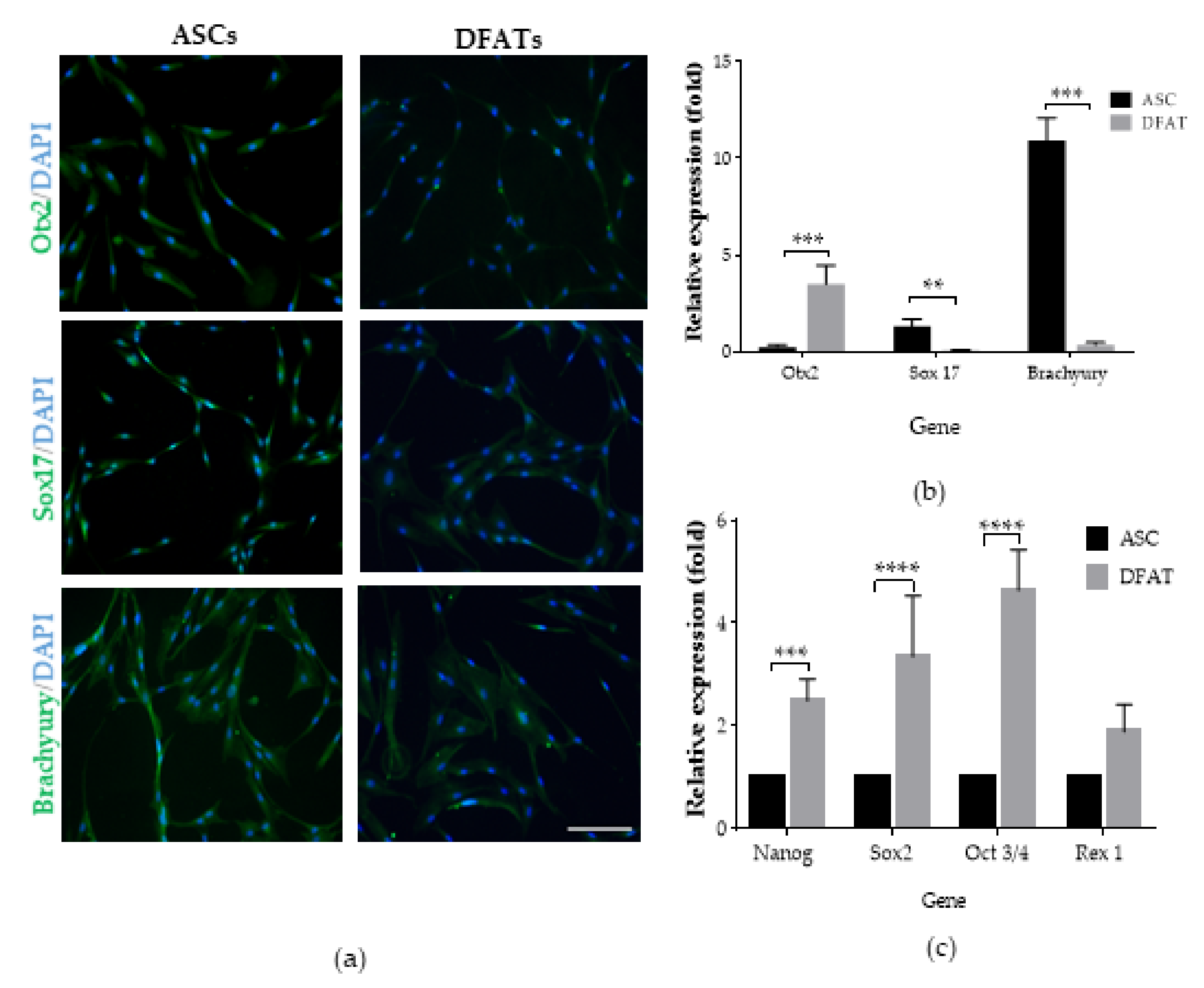
3.2. Pluripotent Properties of ASCs and DFATs
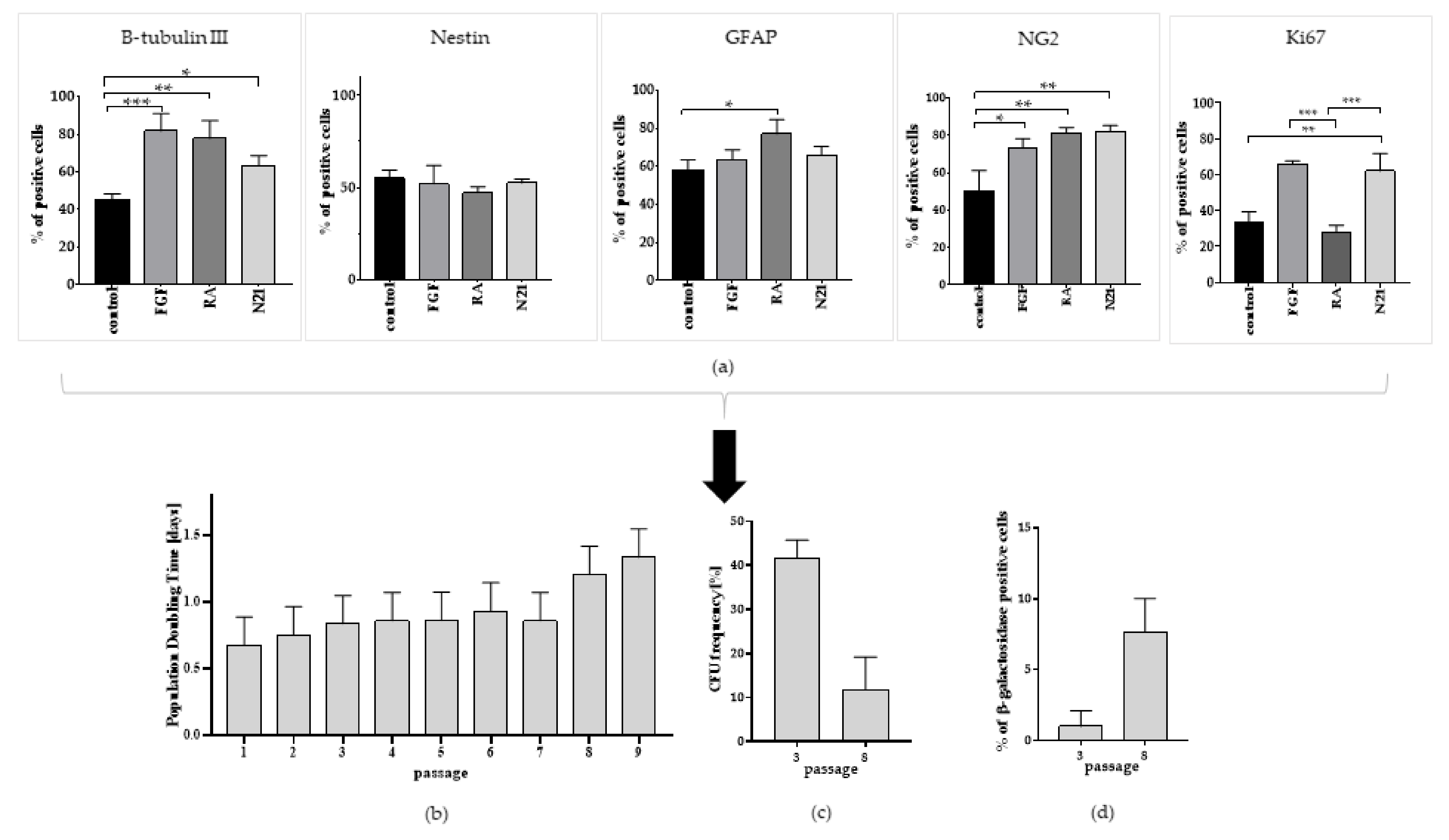
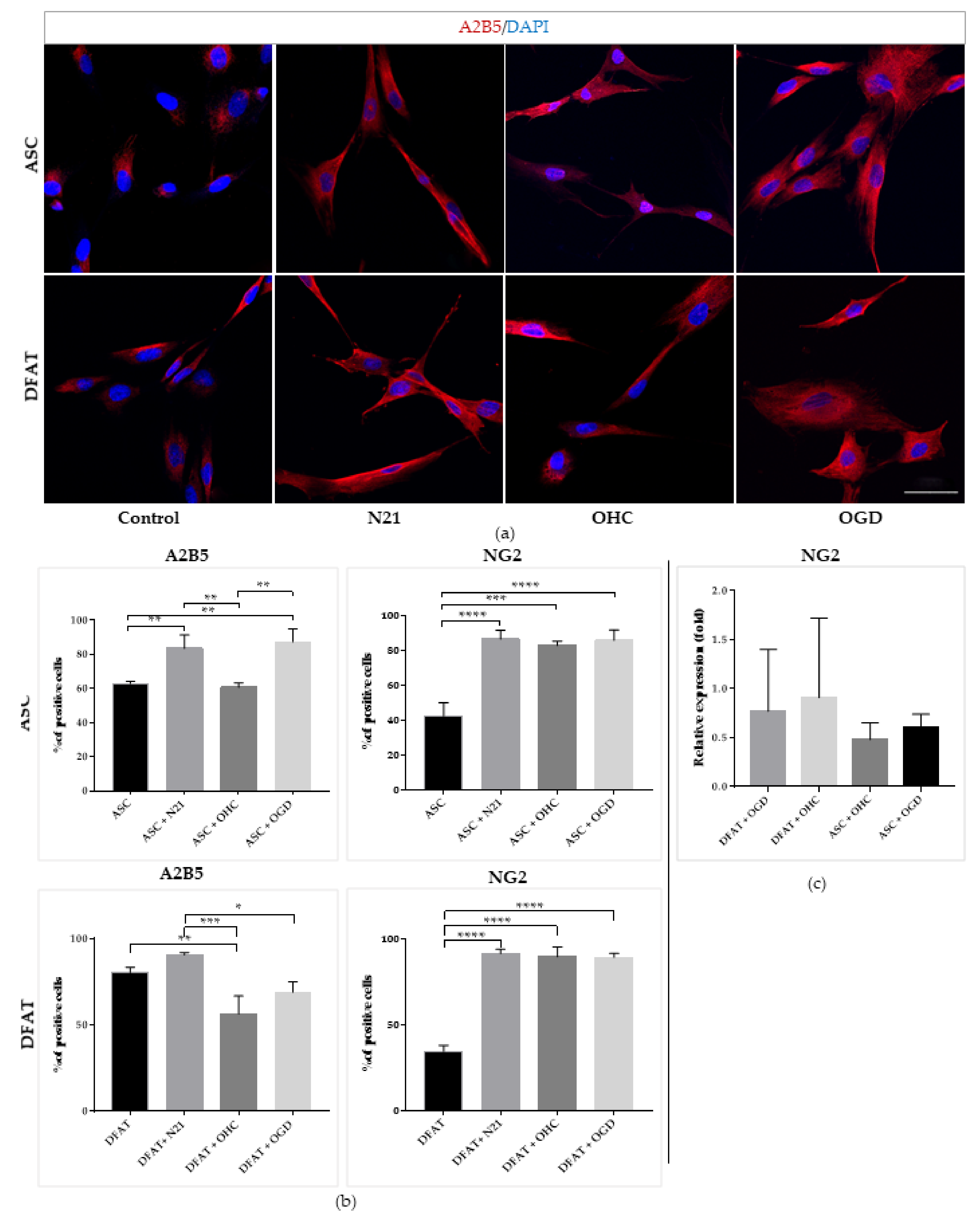
3.3. Optimization of ASCs Capacity for Neural Lineage Differentiation
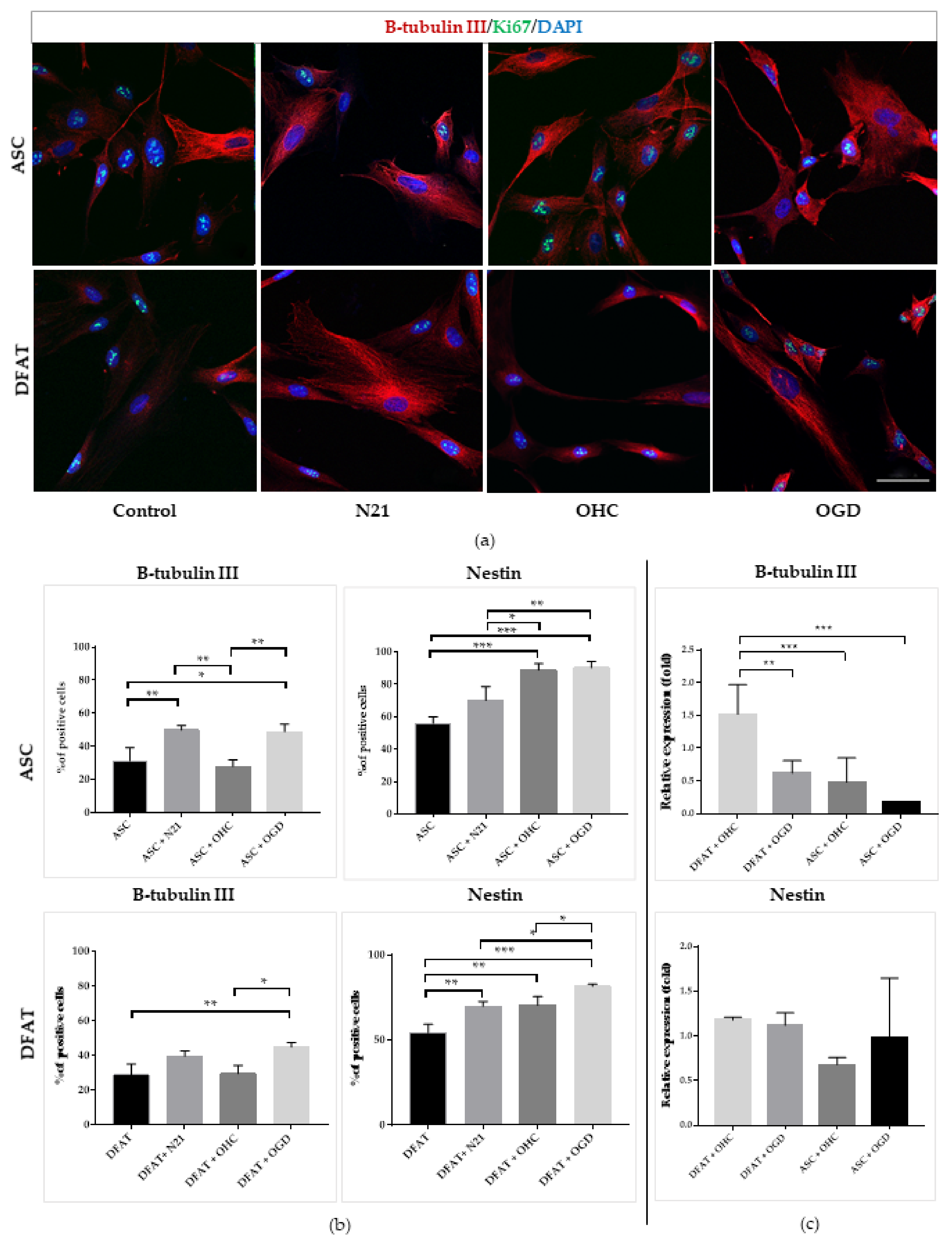
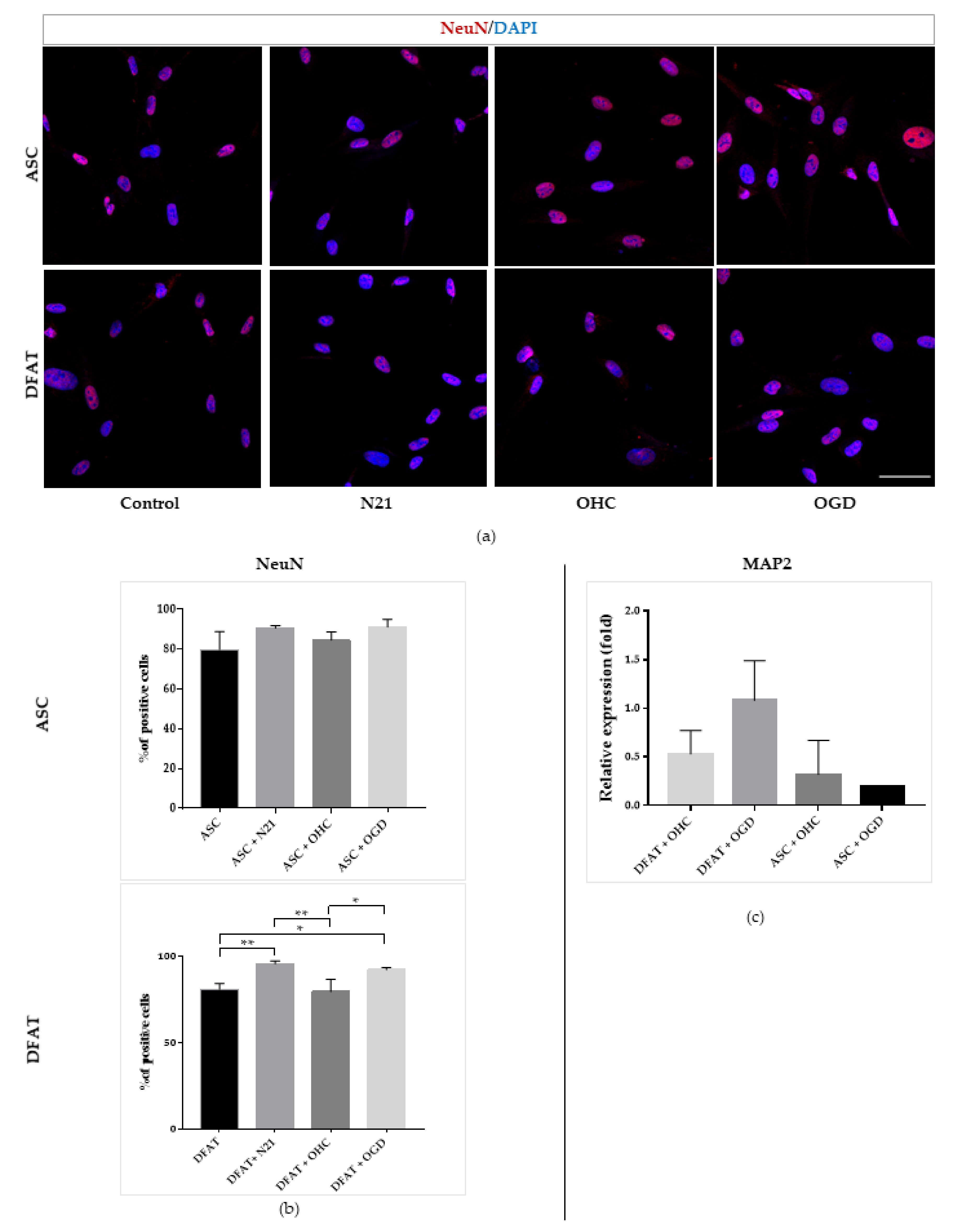
3.4. Comparison of ASCs and DFAT Cells Neural Differentiation Capacity
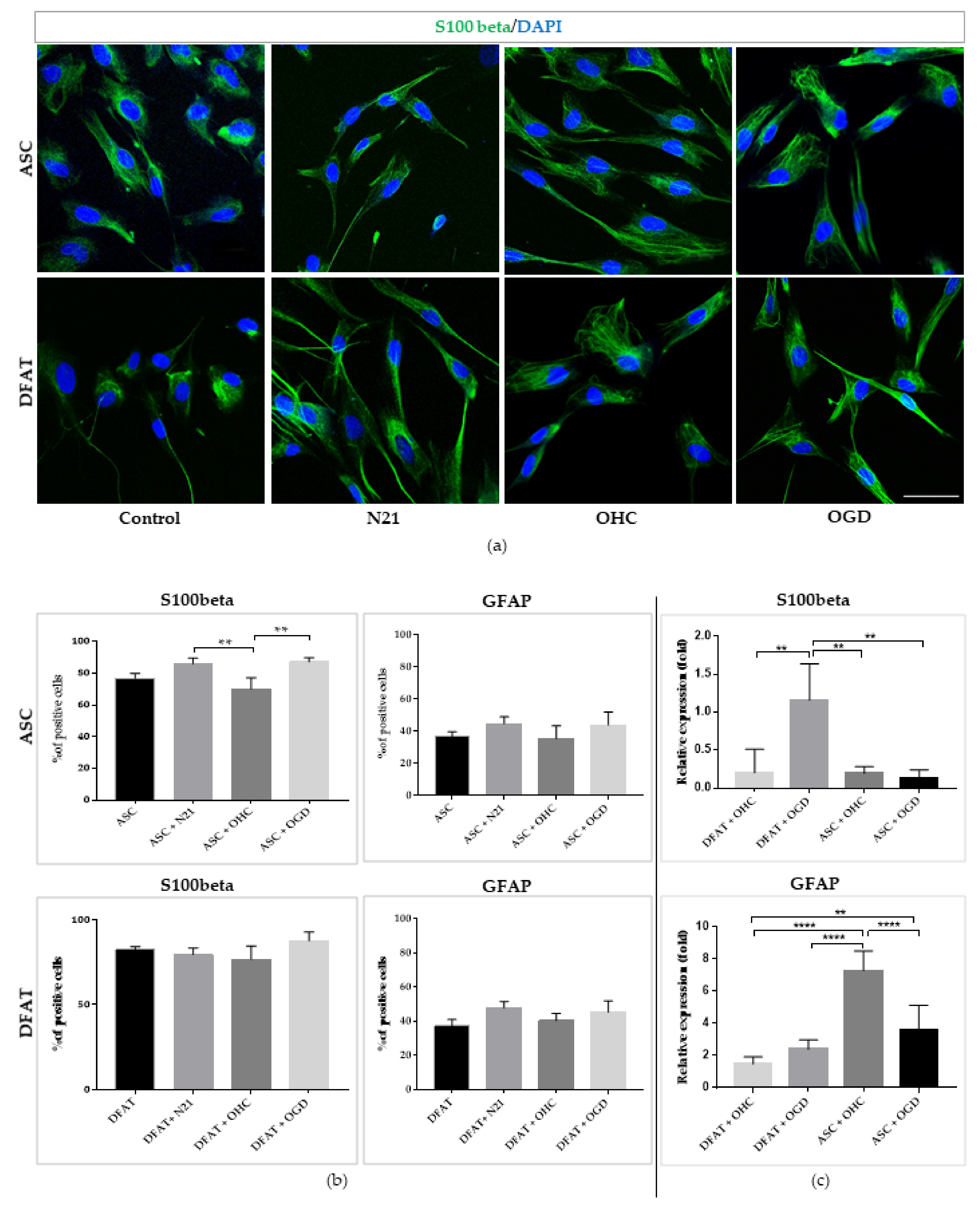
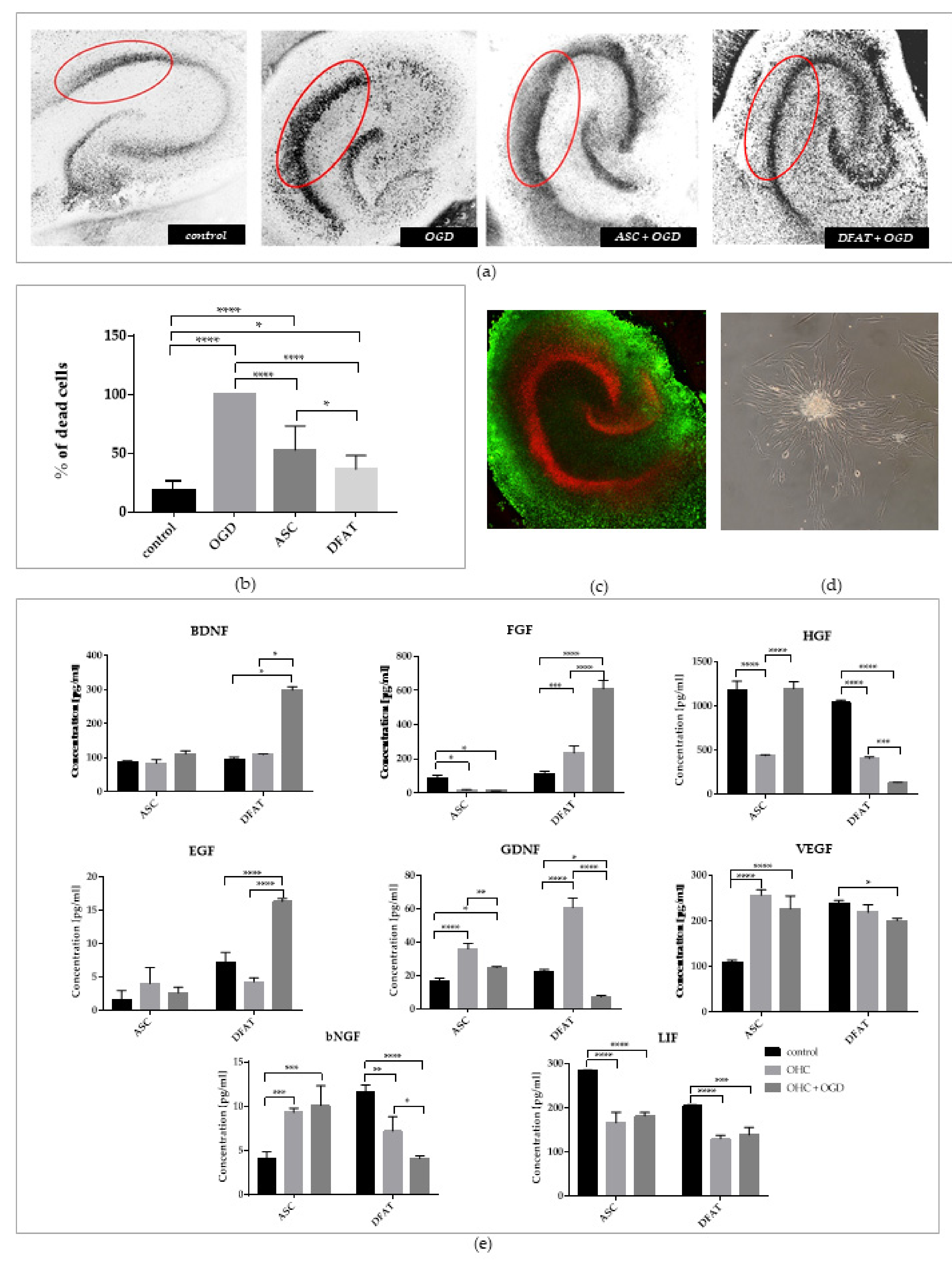
3.5. Neuroprotective Abilities of ASCs and DFAT Cells in the Ex Vivo Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Onorati, M.; Camnasio, S.; Binetti, M.; Jung, C.B.; Moretti, A.; Cattaneo, E. Neuropotent self-renewing neural stem (NS) cells derived from mouse induced pluripotent stem (iPS) cells. Mol. Cell. Neurosci. 2010, 43, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Spiliotopoulos, D.; Goffredo, D.; Conti, L.; Di Febo, F.; Biella, G.; Toselli, M.; Cattaneo, E. An optimized experimental strategy for efficient conversion of embryonic stem (ES)-derived mouse neural stem (NS) cells into a nearly homogeneous mature neuronal population. Neurobiol. Dis. 2009, 34, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, S. Pluripotent Stem Cell-Based Cell Therapy—Promise and Challenges. Cell Stem Cell 2020, 27, 523–531. [Google Scholar] [CrossRef]
- Hepler, C.; Shan, B.; Zhang, Q.; Henry, G.H.; Shao, M.; Vishvanath, L.; Gupta, R.K.; Ghaben, A.L.; Mobley, A.B.; Strand, D. Identification of functionally distinct fibro-inflammatory and adipogenic stromal subpopulations in visceral adipose tissue of adult mice. Elife 2018, 7, e39636. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Hoogduijn, M.J.; Baan, C.C.; Korevaar, S.S.; De Kuiper, R.; Yan, L.; Wang, L.; Van Besouw, N.M. Adipose Tissue-Derived Mesenchymal Stem Cells Have a Heterogenic Cytokine Secretion Profile. Stem Cells Int. 2017, 2017, 1–7. [Google Scholar] [CrossRef]
- Zanata, F.; Shaik, S.; Devireddy, R.V.; Wu, X.; Ferreira, L.M.; Gimble, J.M. Cryopreserved Adipose Tissue-Derived Stromal/Stem Cells: Potential for Applications in Clinic and Therapy. In Advances in Experimental Medicine and Biology; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2016; Volume 951, pp. 137–146. [Google Scholar]
- Liu, M.; Lei, H.; Dong, P.; Fu, X.; Yang, Z.; Yang, Y.; Ma, J.; Liu, X.; Cao, Y.; Xiao, R. Adipose-Derived Mesenchymal Stem Cells from the Elderly Exhibit Decreased Migration and Differentiation Abilities with Senescent Properties. Cell Transplant. 2017, 26, 1505–1519. [Google Scholar] [CrossRef]
- Assoni, A.; Coatti, G.; Valadares, M.C.; Beccari, M.; Gomes, J.; Pelatti, M.; Mitne-Neto, M.; Carvalho, V.M.; Zatz, M. Different Donors Mesenchymal Stromal Cells Secretomes Reveal Heterogeneous Profile of Relevance for Therapeutic Use. Stem Cells Dev. 2017, 26, 206–214. [Google Scholar] [CrossRef]
- Sakai, D.; Wakamatsu, Y. Regulatory Mechanisms for Neural Crest Formation. Cells Tissues Organs 2005, 179, 24–35. [Google Scholar] [CrossRef]
- Sowa, Y.; Imura, T.; Numajiri, T.; Takeda, K.; Mabuchi, Y.; Matsuzaki, Y.; Nishino, K. Adipose Stromal Cells Contain Phenotypically Distinct Adipogenic Progenitors Derived from Neural Crest. PLoS ONE 2013, 8, e84206. [Google Scholar] [CrossRef]
- Wrage, P.C.; Tran, T.; To, K.; Keefer, E.W.; Ruhn, K.A.; Hong, J.; Hattangadi, S.; Treviño, I.; Tansey, M.G. The Neuro-Glial Properties of Adipose-Derived Adult Stromal (ADAS) Cells Are Not Regulated by Notch 1 and Are Not Derived from Neural Crest Lineage. PLoS ONE 2008, 3, e1453. [Google Scholar] [CrossRef]
- Yagi, K.; Kondo, D.; Okazaki, Y.; Kano, K. A novel preadipocyte cell line established from mouse adult mature adipocytes. Biochem. Biophys. Res. Commun. 2004, 321, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Kano, K.; Kondo, D.; Fukuda, N.; Iribe, Y.; Tanaka, N.; Matsubara, Y.; Sakuma, T.; Satomi, A.; Otaki, M.; et al. Mature adipocyte-derived dedifferentiated fat cells exhibit multilineage potential. J. Cell. Physiol. 2008, 215, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Goto, S.; Kato, R.; Komiyama, S.; Nagaoka, Y.; Kazama, T.; Yamamoto, C.; Li, Y.; Konuma, N.; Hagikura, K.; et al. The neovascularization effect of dedifferentiated fat cells. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Ohta, Y.; Takenaga, M.; Tokura, Y.; Hamaguchi, A.; Matsumoto, T.; Kano, K.; Mugishima, H.; Okano, H.; Igarashi, R. Mature adipocyte-derived cells, dedifferentiated fat cells (DFAT), promoted functional recovery from spinal cord injury-induced motor dysfunction in rats. Cell Transplant. 2008, 17, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Fujimaki, H.; Matsumine, H.; Osaki, H.; Ueta, Y.; Kamei, W.; Shimizu, M.; Hashimoto, K.; Fujii, K.; Kazama, T.; Matsumoto, T.; et al. Dedifferentiated fat cells in polyglycolic acid-collagen nerve conduits promote rat facial nerve regeneration. Regen. Ther. 2019, 11, 240–248. [Google Scholar] [CrossRef]
- Mikrogeorgiou, A.; Kondo, T.; Ito, M.; Nakanishi, K.; Hayakawa, M.; Sato, Y.; Hattori, T.; Sugiyama, Y.; Saito, A.; Tsuji, M.; et al. Dedifferentiated Fat Cells as a Novel Source for Cell Therapy to Target Neonatal Hypoxic-Ischemic Encephalopathy. Dev. Neurosci. 2017, 39, 273–286. [Google Scholar] [CrossRef]
- Lin, L.; Du, L. The role of secreted factors in stem cells-mediated immune regulation. Cell. Immunol. 2018, 326, 24–32. [Google Scholar] [CrossRef]
- Cao, W.; Cao, K.; Cao, J.; Wang, Y.; Shi, Y. Mesenchymal stem cells and adaptive immune responses. Immunol. Lett. 2015, 168, 147–153. [Google Scholar] [CrossRef]
- Le Blanc, K.; Mougiakakos, D. Multipotent mesenchymal stromal cells and the innate immune system. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef]
- Shi, Y.; Du, L.; Lin, L.; Wang, Y. Tumour-associated mesenchymal stem/stromal cells: Emerging therapeutic targets. Nat. Rev. Drug Discov. 2017, 16, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, E.S.; Brock, J.H.; Lu, P.; Kumamaru, H.; A Salegio, E.; Kadoya, K.; Weber, J.L.; Liang, J.J.; Moseanko, R.; Hawbecker, S.; et al. Restorative effects of human neural stem cell grafts on the primate spinal cord. Nat. Med. 2018, 24, 484–490. [Google Scholar] [CrossRef]
- Liang, X.; Ding, Y.; Zhang, Y.; Tse, H.F.; Lian, Q. Paracrine Mechanisms of Mesenchymal Stem Cell-Based Therapy: Current Status and Perspectives. Cell Transplant. 2014, 23, 1045–1059. [Google Scholar] [CrossRef] [PubMed]
- Balu, D.T.; Lucki, I. Adult hippocampal neurogenesis: Regulation, functional implications, and contribution to disease pathology. Neurosci. Biobehav. Rev. 2009, 33, 232–252. [Google Scholar] [CrossRef]
- Qian, L.; Saltzman, W.M. Improving the expansion and neuronal differentiation od mesenchymal stem cells through culture surface modification. Biomaterials 2003, 25, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- Urdzíková, L.M.; Růžička, J.; Labagnara, M.; Kárová, K.; Kubinová, Š.; Jiráková, K.; Murali, R.; Syková, E.; Jhanwar-Uniyal, M.; Jendelová, P. Human Mesenchymal Stem Cells Modulate Inflammatory Cytokines after Spinal Cord Injury in Rat. Int. J. Mol. Sci. 2014, 15, 11275–11293. [Google Scholar] [CrossRef]
- Kingham, P.J.; Kolar, M.K.; Novikova, L.N.; Novikov, L.N.; Wiberg, M. Stimulating the Neurotrophic and Angiogenic Properties of Human Adipose-Derived Stem Cells Enhances Nerve Repair. Stem Cells Dev. 2014, 23, 741–754. [Google Scholar] [CrossRef] [PubMed]
- Marconi, S.; Castiglione, G.; Turano, E.; Bissolotti, G.; Angiari, S.; Farinazzo, A.; Constantin, G.; Bedogni, G.; Bedogni, A.; Bonetti, B. Human Adipose-Derived Mesenchymal Stem Cells Systemically Injected Promote Peripheral Nerve Regeneration in the Mouse Model of Sciatic Crush. Tissue Eng. Part A 2012, 18, 1264–1272. [Google Scholar] [CrossRef]
- Kuhn, H.G.; Winkler, J.; Kempermann, G.; Thal, L.J.; Gage, F.H. Epidermal Growth Factor and Fibroblast Growth Factor-2 Have Different Effects on Neural Progenitors in the Adult Rat Brain. J. Neurosci. 1997, 17, 5820–5829. [Google Scholar] [CrossRef]
- Mashayekhi, F. Neural cell death is induced by neutralizing antibody to nerve growth factor: An in vivo study. Brain Dev. 2008, 30, 112–117. [Google Scholar] [CrossRef]
- Prautsch, K.M.; Schmidt, A.; Paradiso, V.; Schaefer, D.J.; Guzman, R.; Kalbermatten, D.F.; Madduri, S. Modulation of Human Adipose Stem Cells’ Neurotrophic Capacity Using a Variety of Growth Factors for Neural Tissue Engineering Applications: Axonal Growth, Transcriptional, and Phosphoproteomic Analyses In Vitro. Cells 2020, 9, 1939. [Google Scholar] [CrossRef]
- Rehman, J.; Traktuev, D.; Li, J.; Merfeld-Clauss, S.; Temm-Grove, C.J.; Bovenkerk, J.E.; Pell, C.L.; Johnstone, B.H.; Considine, R.V.; March, K.L. Secretion of Angiogenic and Antiapoptotic Factors by Human Adipose Stromal Cells. Circulation 2004, 109, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Chan, T.-M.; Chen, J.Y.-R.; Ho, L.-I.; Lin, H.-P.; Hsueh, K.-W.; Liu, D.D.; Chen, Y.-H.; Hsieh, A.-C.; Tsai, N.-M.; Hueng, D.-Y.; et al. ADSC Therapy in Neurodegenerative Disorders. Cell Transplant. 2014, 23, 549–557. [Google Scholar] [CrossRef]
- Skalska, U.; Kontny, E. Regenerative and immunomodulatory properties of adipose-derived mesenchymal stem cells. Postępy Biol. Komórki 2011, 38, 363–378. [Google Scholar]
- Anghileri, E.; Marconi, S.; Pignatelli, A.; Cifelli, P.; Galie’, M.; Sbarbati, A.; Krampera, M.; Belluzzi, O.; Bonetti, B. Neuronal Differentiation Potential of Human Adipose-Derived Mesenchymal Stem Cells. Stem Cells Dev. 2008, 17, 909–916. [Google Scholar] [CrossRef]
- Hu, F.; Wang, X.; Liang, G.; Lv, L.; Zhu, Y.; Sun, B.; Xiao, Z. Effects of Epidermal Growth Factor and Basic Fibroblast Growth Factor on the Proliferation and Osteogenic and Neural Differentiation of Adipose-Derived Stem Cells. Cell. Reprogramm. 2013, 15, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Zhou, J.; Jing’e Wang, B.X.; Li, L.; Zhu, F.; Han, J.; Luo, X. Neuronal differentiation of adipose-derived stem cells and their transplantation for cerebral ischemia. Neural Regen. Res. 2012, 7, 1992. [Google Scholar]
- Hu, F.; Sun, B.; Xu, P.; Zhu, Y.; Meng, X.-H.; Teng, G.-J.; Xiao, Z.-D. MiR-218 Induces Neuronal Differentiation of ASCs in a Temporally Sequential Manner with Fibroblast Growth Factor by Regulation of the Wnt Signaling Pathway. Sci. Rep. 2017, 7, 39427. [Google Scholar] [CrossRef]
- Moore, T.J.; Abrahamse, H. Neuronal Differentiation of Adipose Derived Stem Cells: Progress So Far. Int. J. Photoenergy 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Jang, S.; Cho, H.-H.; Cho, Y.-B.; Park, J.-S.; Jeong, H.-S. Functional neural differentiation of human adipose tissue-derived stem cells using bFGF and forskolin. BMC Cell Biol. 2010, 11, 25. [Google Scholar] [CrossRef]
- Takahashi, J.; Palmer, T.D.; Gage, F.H. Retinoic acid and neurotrophins collaborate to regulate neurogenesis in adult-derived neural stem cell cultures. J. Neurobiol. 1999, 38, 65–81. [Google Scholar] [CrossRef]
- George, S.; Hamblin, M.R.; Abrahamse, H. Current and Future Trends in Adipose Stem Cell Differentiation into Neuroglia. Photomed. Laser Surg. 2018, 36, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Janesick, A.; Wu, S.C.; Blumberg, B. Retinoic acid signaling and neuronal differentiation. Cell. Mol. Life Sci. 2015, 72, 1559–1576. [Google Scholar] [CrossRef] [PubMed]
- Koutmani, Y.; Gampierakis, I.A.; Polissidis, A.; Ximerakis, M.; Koutsoudaki, P.N.; Polyzos, A.; Agrogiannis, G.; Karaliota, S.; Thomaidou, D.; Rubin, L.; et al. CRH Promotes the Neurogenic Activity of Neural Stem Cells in the Adult Hippocampus. Cell Rep. 2019, 29, 932–945.e7. [Google Scholar] [CrossRef]
- Jumabay, M.; Abdmaulen, R.; Ly, A.; Cubberly, M.R.; Shahmirian, L.J.; Heydarkhan-Hagvall, S.; Dumesic, D.A.; Yao, Y.; Boström, K.I. Pluripotent Stem Cells Derived from Mouse and Human White Mature Adipocytes. Stem Cells Transl. Med. 2014, 3, 161–171. [Google Scholar] [CrossRef]
- Wei, S.; Zan, L.; Hausman, G.J.; Rasmussen, T.P.; Bergen, W.G.; Dodson, M.V. Dedifferentiated adipocyte-derived progeny cells (DFAT cells). Adipocyte 2013, 2, 122–127. [Google Scholar] [CrossRef]
- Gao, Q.; Zhao, L.; Song, Z.; Yang, G. Expression pattern of embryonic stem cell markers in DFAT cells and ADSCs. Mol. Biol. Rep. 2012, 39, 5791–5804. [Google Scholar] [CrossRef]
- Dezawa, M. Muse Cells Provide the Pluripotency of Mesenchymal Stem Cells: Direct Contribution of Muse Cells to Tissue Regeneration. Cell Transplant. 2016, 25, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Jouhilahti, E.-M.; Peltonen, S.; Peltonen, J. Class III β-Tubulin Is a Component of the Mitotic Spindle in Multiple Cell Types. J. Histochem. Cytochem. 2008, 56, 1113–1119. [Google Scholar] [CrossRef]
- Dráberová, E.; Del Valle, L.; Gordon, J.; Marková, V.; Šmejkalová, B.; Bertrand, L.; De Chadarévian, J.-P.; Agamanolis, D.P.; Legido, A.; Khalili, K.; et al. Class III β-Tubulin Is Constitutively Coexpressed with Glial Fibrillary Acidic Protein and Nestin in Midgestational Human Fetal Astrocytes: Implications for Phenotypic Identity. J. Neuropathol. Exp. Neurol. 2008, 67, 341–354. [Google Scholar] [CrossRef]
- Zheng, Y.; Huang, C.; Liu, F.; Lin, H.; Yang, X.; Zhang, Z. Comparison of the neuronal differentiation abilities of bone marrow-derived and adipose tissue-derived mesenchymal stem cells. Mol. Med. Rep. 2017, 16, 3877–3886. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yang, H.; Xia, Y.; Lu, S.Q.; Soong, T.W.; Feng, Z.W. Basic Fibroblast Growth Factor-induced Neuronal Differentiation of Mouse Bone Marrow Stromal Cells Requires FGFR-1, MAPK/ERK, and Transcription Factor AP-1. J. Biol. Chem. 2008, 283, 5287–5295. [Google Scholar] [CrossRef]
- Cardozo, A.J.; Gómez, D.E.; Argibay, P.F. Transcriptional Characterization of Wnt and Notch Signaling Pathways in Neuronal Differentiation of Human Adipose Tissue-Derived Stem Cells. J. Mol. Neurosci. 2011, 44, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Stevens, B.; Chang, J.; Milbrandt, J.; Barres, B.A.; Hell, J.W. NS21: Re-defined and modified supplement B27 for neuronal cultures. J. Neurosci. Methods 2008, 171, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Drela, K.; Siedlecka, P.; Sarnowska, A.; Domanska-Janik, K. Human mesenchymal stem cells in the treatment of neurological diseases. Acta Neurobiol. Exp. 2013, 73, 38–56. [Google Scholar]
- Basciano, L.; Nemos, C.; Foliguet, B.; De Isla, N.; De Carvalho, M.; Tran, N.; Dalloul, A. Long term culture of mesenchymal stem cells in hypoxia promotes a genetic program maintaining their undifferentiated and multipotent status. BMC Cell Biol. 2011, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Namekata, K.; Guo, X.; Harada, C.; Harada, T. Neuroprotection, Growth Factors and BDNF-TrkB Signalling in Retinal Degeneration. Int. J. Mol. Sci. 2016, 17, 1584. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, D.; Feng, G.; Ma, Y.; Hu, H. Recombinant AAV-mediated expression of human BDNF protects neurons against cell apoptosis in Aβ-induced neuronal damage model. Acta Acad. Med. 2007, 27, 233–236. [Google Scholar] [CrossRef]
- Chen, A.; Xiong, L.-J.; Tong, Y.; Mao, M. The neuroprotective roles of BDNF in hypoxic ischemic brain injury. Biomed. Rep. 2012, 1, 167–176. [Google Scholar] [CrossRef]
- Schäbitz, W.-R.; Berger, C.; Kollmar, R.; Seitz, M.; Tanay, E.; Kiessling, M.; Schwab, S.; Sommer, C. Effect of Brain-Derived Neurotrophic Factor Treatment and Forced Arm Use on Functional Motor Recovery After Small Cortical Ischemia. Stroke 2004, 35, 992–997. [Google Scholar] [CrossRef]
- Kume, T.; Kouchiyama, H.; Kaneko, S.; Maeda, T.; Kaneko, S.; Akaike, A.; Shimohama, S.; Kihara, T.; Kimura, J.; Wada, K.; et al. BDNF prevents NO mediated glutamate cytotoxicity in cultured cortical neurons. Brain Res. 1997, 756, 200–204. [Google Scholar] [CrossRef]
- Sakai, R.; Ukai, W.; Sohma, H.; Hashimoto, E.; Yamamoto, M.; Ikeda, H.; Saito, T. Attenuation of brain derived neurotrophic factor (BDNF) by ethanol and cytoprotective effect of exogenous BDNF against ethanol damage in neuronal cells. J. Neural Transm. 2004, 112, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, T.; Kalinina, N.; Karagyaur, M.; Stambolsky, D.; Rubina, K.; Revischin, A.; Pavlova, G.; Parfyonova, Y.; Tkachuk, V. Adipose-Derived Stem Cells Stimulate Regeneration of Peripheral Nerves: BDNF Secreted by These Cells Promotes Nerve Healing and Axon Growth De Novo. PLoS ONE 2011, 6, e17899. [Google Scholar] [CrossRef]
- Salgado, A.J.B.O.G.; Reis, R.L.; Sousa, N.J.C.; Gimble, J.M. Adipose Tissue Derived Stem Cells Secretome: Soluble Factors and Their Roles in Regenerative Medicine. Curr. Stem Cell Res. Ther. 2010, 5, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Sarnowska, A.; Braun, H.; Sauerzweig, S.; Reymann, K.G.; Minhas, G. The neuroprotective effect of bone marrow stem cells is not dependent on direct cell contact with hypoxic injured tissue. Ann. Neurosci. 2011, 18, 21. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tan, B.; Luan, Z.; Wei, X.; He, Y.; Wei, G.; Johnstone, B.; Farlow, M.; Du, Y. AMP-activated kinase mediates adipose stem cell-stimulated neuritogenesis of PC12 cells. Neuroscience 2011, 181, 40–47. [Google Scholar] [CrossRef]
- Lu, S.; Lu, C.; Han, Q.; Li, J.; Du, Z.; Liao, L.; Zhao, R.C. Adipose-derived mesenchymal stem cells protect PC12 cells from glutamate excitotoxicity-induced apoptosis by upregulation of XIAP through PI3-K/Akt activation. Toxicology 2011, 279, 189–195. [Google Scholar] [CrossRef]
- Crigler, L.; Robey, R.C.; Asawachaicharn, A.; Gaupp, D.; Phinney, D.G. Human mesenchymal stem cell subpopulations express a variety of neuro-regulatory molecules and promote neuronal cell survival and neuritogenesis. Exp. Neurol. 2006, 198, 54–64. [Google Scholar] [CrossRef]
- Obtulowicz, P.; Lech, W.; Strojek, L.; Sarnowska, A.; Domanska-Janik, K. Induction of Endothelial Phenotype from Wharton’s Jelly-Derived MSCs and Comparison of Their Vasoprotective and Neuroprotective Potential with Primary WJ-MSCs in CA1 Hippocampal Region Ex Vivo. Cell Transplant. 2016, 25, 715–727. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Differentiation Lineage | Used Stain | Visualized Compound | Time of Incubation (Minutes) | Concentration |
|---|---|---|---|---|
| Adipogenesis | Oil Red O | Oil Red | 5 | 0.5% |
| Osteogenesis | Alizarin Red S | Calcium deposits | 3 | 2% |
| Chondrogenesis | Alcian blue | Mucopolysaccharides | 30 | 1% |
| Antibody | Catalogue Number | Source | Isotype | Dilution | Manufacturer |
|---|---|---|---|---|---|
| anti-β-Tubulin III | T8660 | Mouse monoclonal | IgG2b | 1:1000 | Sigma-Aldrich |
| anti-GFAP | Z0334 | Rabbit polyclonal | IgG | 1:400 | Dako |
| anti-Ki67 | AB15580 | Rabbit polyclonal | IgG | 1:200 | Abcam |
| anti-Nestin | MAB5326 | Mouse monoclonal | IgG1 | 1:500 | Millipore |
| anti-Fibronectin | F3648 | Mouse monoclonal | IgG | 1:400 | Sigma-Aldrich |
| anti-Vimentin | AB1620 | Mouse monoclonal | IgG1 | 1:200 | Abcam |
| anti-NeuN | MAB377 | Mouse monoclonal | IgG1 | 1:50 | Millipore |
| anti-A2B5 | MAB312R | Mouse monoclonal | IgM | 1:200 | Millipore |
| anti-NG2 | AB5320 | Rabbit polyclonal | IgG | 1:150 | Millipore |
| anti-S100 beta | AB52642 | Rabbit polyclonal | IgG | 1:100 | Abcam |
| Antibody | Fluorochrome | Catalogue Number | Isotype | Dilution | Manufacturer |
|---|---|---|---|---|---|
| Alexa Fluor Goat (anti-rabbit) | Alexa 546 | A11035 | IgG | 1:1000 | Life Technologies |
| Alexa FluorGoat (anti-mouse) | Alexa 546 | A21123 | IgG1 | 1:1000 | Thermo Fisher Scientific |
| Alexa Fluor Goat (anti-mouse) | Alexa 488 | A21121 | IgG1 | 1:1000 | Life Technologies |
| Alexa Fluor Goat (anti-mouse) | Alexa 488 | A21141 | IgG2b | 1:1000 | Life Technologies |
| Alexa Fluor Goat (anti-mouse) | Alexa 546 | A21045 | IgM | 1:1000 | Life Technologies |
| Gene | NCBI Reference Sequence | Product Size | Primer Sequence (5e -> 3-) |
|---|---|---|---|
| β-Actin | NM_001101.5 | 250 bp | F: CATGTACGTTGCTATCCAGGC R: CTCCTTAATGTCACGCACGAT |
| Nanog | NM_024865.4 | 103 bp | F: GAACCTCAGCTACAAACAGG R: CGTCACACCATTGCTATTCT |
| Sox2 | NM_003106.4 | 93 bp | F: GTGGAAACTTTTGTCGGAGA R: TTATAATCCGGGTGCTCCTT |
| Oct3/4 | NM_001285986.2 | 331 bp | F: CCTGAAGCAGAAGAGGATCACC R: AAAGCGGCAGATGGTCGTTTGG |
| Rex1 | NM_001304358.2 | 107 bp | F: GCTCCCTTGAATGTTCTTTG R: GCCTGTCATGTACTCAGAAT |
| Oxt2 | NM_001270523.2 | 98 bp | F: TTCATGCGAGAGGAGGTGGCA R: TGCTGTTGTTGGCGGCACTT |
| Sox17 | NM_022454.4 | 110 bp | F:AACTATCCTGACGTGTGACA R:CAAAAACCCAGGAGTCTGAG |
| Brachyury | NM_001379200.1 | 104 bp | F:ACGGCCACATTATTCTGAAT R:GAAGTTCTCCTCGGCATATT |
| Nestin | NM_006617.2 | 64 bp | F: GGGAAGAGGTGATGGAACCA R: AAGCCCTGAACCCTCTTTGC |
| GFAP | NM_001363846.2 | 100 bp | F: CCGACAGCAGGTCCATGT R: GTTGCTGGACGCCATTG |
| MAP-2 | NM_001375545.1 | 99 bp | F: TTGGTGCCGAGTGAGAAGA R: GTCTGGCAGTGGTTGGTTAA |
| β-Tubulin III | NM_001197181.2 | 126 bp | F: GGAAGAGGGCGAGATGTACG R: GGGTTTAGACACTGCTGGCT |
| S100beta | NM_006272.3 | 91 bp | F: AGCGCTCCTGGAAAAAGCAA R: TTGAATCGCATGGGTCACGG |
| NG2 | NM_001897.5 | 118 bp | F: GTCTACACCATCGAGCAGCC R: TGTGTGAGAACAGCACGAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figiel-Dabrowska, A.; Radoszkiewicz, K.; Rybkowska, P.; Krzesniak, N.E.; Sulejczak, D.; Sarnowska, A. Neurogenic and Neuroprotective Potential of Stem/Stromal Cells Derived from Adipose Tissue. Cells 2021, 10, 1475. https://doi.org/10.3390/cells10061475
Figiel-Dabrowska A, Radoszkiewicz K, Rybkowska P, Krzesniak NE, Sulejczak D, Sarnowska A. Neurogenic and Neuroprotective Potential of Stem/Stromal Cells Derived from Adipose Tissue. Cells. 2021; 10(6):1475. https://doi.org/10.3390/cells10061475
Chicago/Turabian StyleFigiel-Dabrowska, Anna, Klaudia Radoszkiewicz, Paulina Rybkowska, Natalia Ewa Krzesniak, Dorota Sulejczak, and Anna Sarnowska. 2021. "Neurogenic and Neuroprotective Potential of Stem/Stromal Cells Derived from Adipose Tissue" Cells 10, no. 6: 1475. https://doi.org/10.3390/cells10061475
APA StyleFigiel-Dabrowska, A., Radoszkiewicz, K., Rybkowska, P., Krzesniak, N. E., Sulejczak, D., & Sarnowska, A. (2021). Neurogenic and Neuroprotective Potential of Stem/Stromal Cells Derived from Adipose Tissue. Cells, 10(6), 1475. https://doi.org/10.3390/cells10061475