
Identification of Putative Markers That Predict the In Vitro Senescence of Mesenchymal Progenitor Cells


Abstract
1. Introduction
2. Materials and Methods
2.1. Culture of Human PSCs
2.2. Differentiation of Human PSCs into MPCs
2.3. Flow Cytometry
2.4. Mesodermal Lineage Differentiation
2.5. Cell Proliferation Assay
2.6. Microarray Analysis
2.7. Reverse Transcription Polymerase Chain Reaction (RT-PCR) and Quantitative Real-Time PCR (qRT-PCR)
3. Results
3.1. Differentiation of MPCs from PSCs
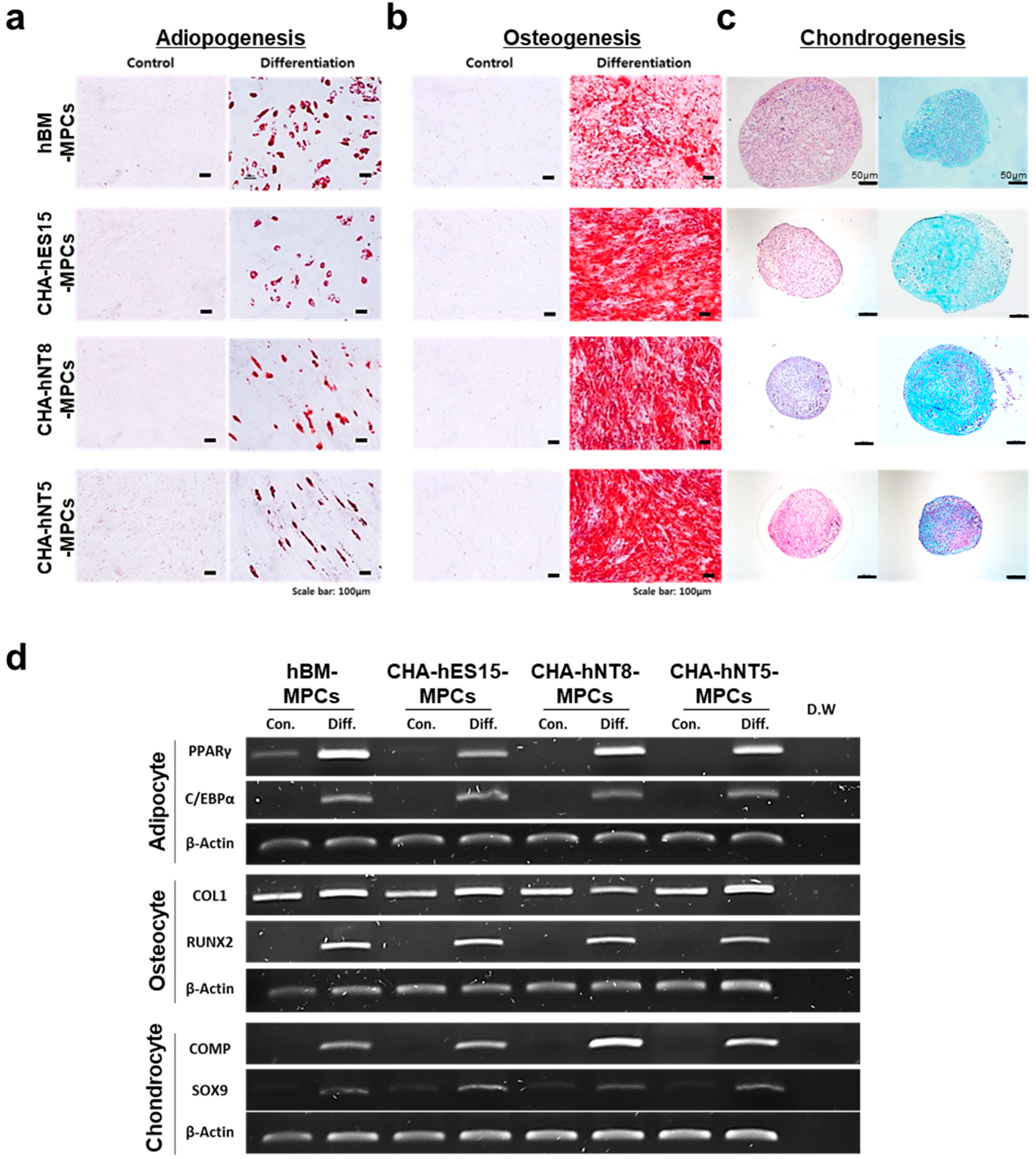
3.2. PSC-Derived MPCs Show Typical MSC Markers and Have Differentiation Capacity
3.3. PSC-MPCs Showed a Robust Growth Potential
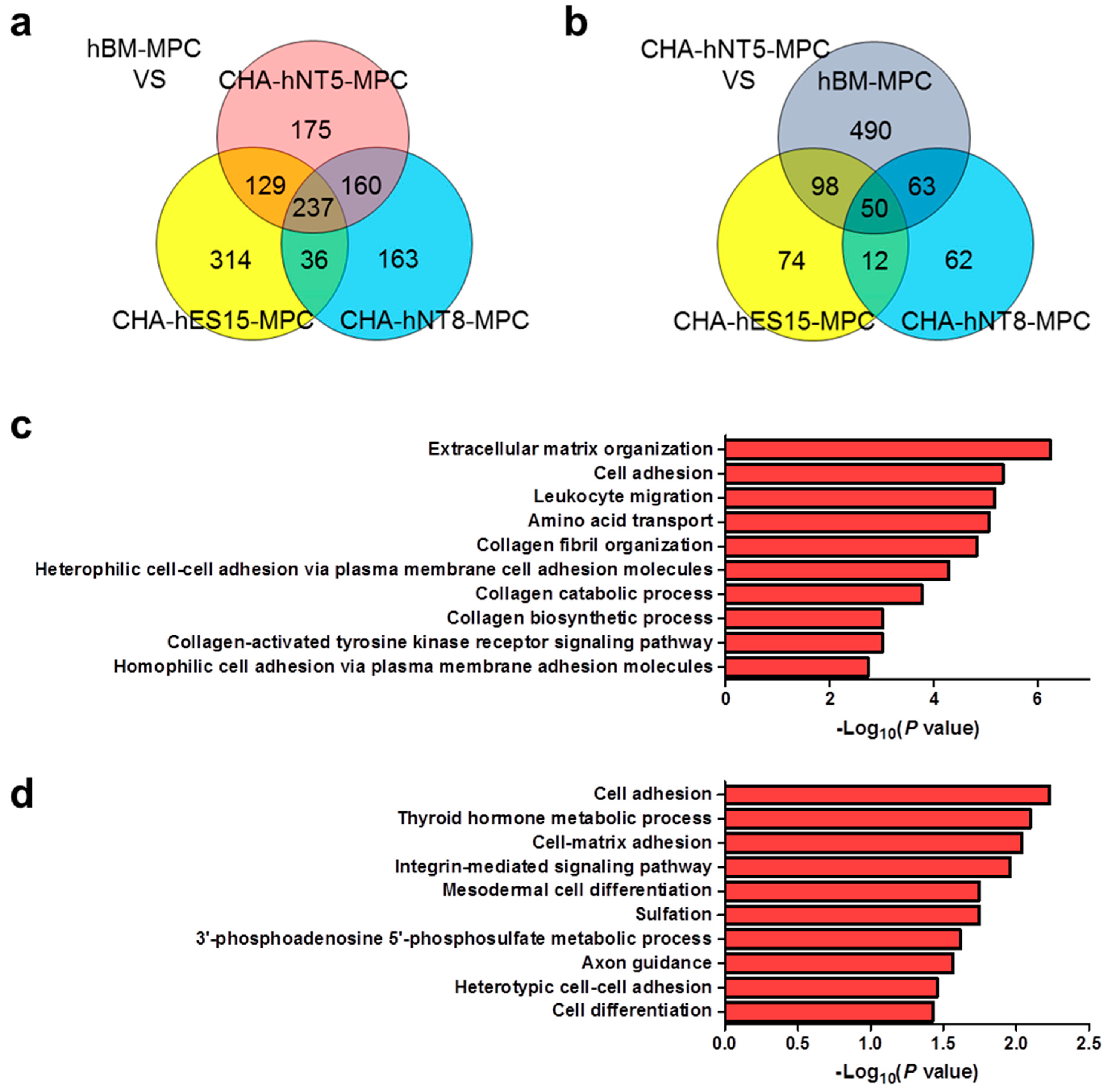
3.4. Identification of Differentially Expressed Genes in High Proliferative MPCs
3.5. Evaluation of Selected Genes on Different Passages of MPCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jimenez-Puerta, G.J.; Marchal, J.A.; López-Ruiz, E.; Gálvez-Martín, P. Role of mesenchymal stromal cells as therapeutic agents: Potential mechanisms of action and implications in their clinical use. J. Clin. Med. 2020, 9, 445. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Liu, S.; De Castro, L.F.; Jin, P.; Civini, S.; Ren, J.; Reems, J.-A.; Cancelas, J.; Nayak, R.; Shaw, G.; O’Brien, T. Manufacturing differences affect human bone marrow stromal cell characteristics and function: Comparison of production methods and products from multiple centers. Sci. Rep. 2017, 7, 46731. [Google Scholar] [CrossRef] [PubMed]
- Boregowda, S.V.; Krishnappa, V.; Haga, C.L.; Ortiz, L.A.; Phinney, D.G. A clinical indications prediction scale based on TWIST1 for human mesenchymal stem cells. EBioMedicine 2016, 4, 62–73. [Google Scholar] [CrossRef]
- Menard, C.; Pacelli, L.; Bassi, G.; Dulong, J.; Bifari, F.; Bezier, I.; Zanoncello, J.; Ricciardi, M.; Latour, M.; Bourin, P. Clinical-grade mesenchymal stromal cells produced under various good manufacturing practice processes differ in their immunomodulatory properties: Standardization of immune quality controls. Stem Cells Dev. 2013, 22, 1789–1801. [Google Scholar] [CrossRef] [PubMed]
- Siegel, G.; Kluba, T.; Hermanutz-Klein, U.; Bieback, K.; Northoff, H.; Schäfer, R. Phenotype, donor age and gender affect function of human bone marrow-derived mesenchymal stromal cells. BMC Med. 2013, 11, 146. [Google Scholar] [CrossRef]
- Phinney, D.G.; Galipeau, J. Manufacturing mesenchymal stromal cells for clinical applications: A survey of Good Manufacturing Practices at US academic centers. Cytotherapy 2019, 21, 782–792. [Google Scholar] [CrossRef]
- Jiang, B.; Yan, L.; Wang, X.; Li, E.; Murphy, K.; Vaccaro, K.; Li, Y.; Xu, R.H. Concise Review: Mesenchymal Stem Cells Derived from Human Pluripotent Cells, an Unlimited and Quality-Controllable Source for Therapeutic Applications. Stem Cells 2019, 37, 572–581. [Google Scholar] [CrossRef]
- Kimbrel, E.A.; Kouris, N.A.; Yavanian, G.J.; Chu, J.; Qin, Y.; Chan, A.; Singh, R.P.; McCurdy, D.; Gordon, L.; Levinson, R.D. Mesenchymal stem cell population derived from human pluripotent stem cells displays potent immunomodulatory and therapeutic properties. Stem Cells Dev. 2014, 23, 1611–1624. [Google Scholar] [CrossRef]
- Li, E.; Zhang, Z.; Jiang, B.; Yan, L.; Park, J.W.; Xu, R.-H. Generation of mesenchymal stem cells from human embryonic stem cells in a complete serum-free condition. Int. J. Biol. Sci. 2018, 14, 1901. [Google Scholar] [CrossRef]
- Zhang, J.; Chan, Y.-C.; Ho, J.C.-Y.; Siu, C.-W.; Lian, Q.; Tse, H.-F. Regulation of cell proliferation of human induced pluripotent stem cell-derived mesenchymal stem cells via ether-a-go-go 1 (hEAG1) potassium channel. Am. J. Physiol. Cell Physiol. 2012, 303, C115–C125. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.M.; Park, M.; Lee, J.Y.; Jung, S.; Lee, J.E.; Shim, S.H.; Song, H.; Lee, D.R. Single cell-derived clonally expanded mesenchymal progenitor cells from somatic cell nuclear transfer-derived pluripotent stem cells ameliorate the endometrial function in the uterus of a murine model with Asherman’s syndrome. Cell Prolif. 2019, 52, e12597. [Google Scholar] [CrossRef] [PubMed]
- Billing, A.M.; Dib, S.S.; Bhagwat, A.M.; da Silva, I.T.; Drummond, R.D.; Hayat, S.; Al-Mismar, R.; Ben-Hamidane, H.; Goswami, N.; Engholm-Keller, K. A systems-level characterization of the differentiation of human embryonic stem cells into mesenchymal stem cells. Mol. Cell Proteom. 2019, 18, 1950–1966. [Google Scholar] [CrossRef]
- Frobel, J.; Hemeda, H.; Lenz, M.; Abagnale, G.; Joussen, S.; Denecke, B.; Šarić, T.; Zenke, M.; Wagner, W. Epigenetic rejuvenation of mesenchymal stromal cells derived from induced pluripotent stem cells. Stem Cell Rep. 2014, 3, 414–422. [Google Scholar] [CrossRef]
- Spitzhorn, L.-S.; Megges, M.; Wruck, W.; Rahman, M.S.; Otte, J.; Degistirici, Ö.; Meisel, R.; Sorg, R.V.; Oreffo, R.O.; Adjaye, J. Human iPSC-derived MSCs (iMSCs) from aged individuals acquire a rejuvenation signature. Stem Cell Res. Ther. 2019, 10, 100. [Google Scholar] [CrossRef]
- Vodyanik, M.A.; Yu, J.; Zhang, X.; Tian, S.; Stewart, R.; Thomson, J.A.; Slukvin, I.I. A mesoderm-derived precursor for mesenchymal stem and endothelial cells. Cell Stem Cell 2010, 7, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Gadue, P.; Huber, T.L.; Paddison, P.J.; Keller, G.M. Wnt and TGF-β signaling are required for the induction of an in vitro model of primitive streak formation using embryonic stem cells. Proc. Natl. Acad. Sci. USA 2006, 103, 16806–16811. [Google Scholar] [CrossRef]
- Lipsitz, Y.Y.; Timmins, N.E.; Zandstra, P.W. Quality cell therapy manufacturing by design. Nat. Biotechnol. 2016, 34, 393–400. [Google Scholar] [CrossRef]
- Zaim, M.; Karaman, S.; Cetin, G.; Isik, S. Donor age and long-term culture affect differentiation and proliferation of human bone marrow mesenchymal stem cells. Ann. Hematol. 2012, 91, 1175–1186. [Google Scholar] [CrossRef]
- Barekzai, J.; Petry, F.; Zitzmann, J.; Czermak, P.; Salzig, D. Bioprocess Development for Human Mesenchymal Stem Cell Therapy Products. In New Advances on Fermentation Processes; IntechOpen: London, UK, 2019. [Google Scholar]
- Shi, Z.; Rockey, D.C. Upregulation of the actin cytoskeleton via myocardin leads to increased expression of type 1 collagen. Lab. Investig. 2017, 97, 1412–1426. [Google Scholar] [CrossRef]
- Creemers, E.E.; Sutherland, L.B.; McAnally, J.; Richardson, J.A.; Olson, E.N. Myocardin is a direct transcriptional target of Mef2, Tead and Foxo proteins during cardiovascular development. Development 2006, 133, 4245–4256. [Google Scholar] [CrossRef] [PubMed]
- Lacolley, P.; Li, Z.; Challande, P.; Regnault, V. SRF/Myocardin: A Novel Molecular Axis Regulating Vascular Smooth Muscle Cell Stiffening in Hypertension; Oxford University Press: Oxford, UK, 2017. [Google Scholar]
- Tucker, R.P.; Chiquet-Ehrismann, R. Teneurins: A conserved family of transmembrane proteins involved in intercellular signaling during development. Dev. Biol. 2006, 290, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Leamey, C.A.; Sawatari, A. The teneurins: New players in the generation of visual topography. Semin. Cell Dev. Biol. 2014, 35, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, J.; Schubert, R.; Chiquet-Ehrismann, R.; Müller, D.J. Deciphering teneurin domains that facilitate cellular recognition, cell-cell adhesion, and neurite outgrowth using atomic force microscopy-based single-cell force spectroscopy. Nano Lett. 2013, 13, 2937–2946. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.K.; Chen, C.S. Cell adhesion and mechanical stimulation in the regulation of mesenchymal stem cell differentiation. J. Cell. Mol. Med. 2013, 17, 823–832. [Google Scholar] [CrossRef]
- Razinia, Z.; Castagnino, P.; Xu, T.; Vázquez-Salgado, A.; Puré, E.; Assoian, R.K. Stiffness-dependent motility and proliferation uncoupled by deletion of CD44. Sci. Rep. 2017, 7, 16499. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ -> 3′) | |
|---|---|---|
| β-ACTIN | Forward | TGAAGTGTGACGTGGACATC |
| Reverse | GGAGGAGCAATGATCTTGAT | |
| PPARγ | Forward | TGTCTCATAATGCCATCAGGTTTG |
| Reverse | GATAACGATGGTGATTTGTCTGTT | |
| C/EBPα | Forward | GCAAACTCACCGCTCCAATG |
| Reverse | TTAGGTTCCAAGCCCCAAGTC | |
| COL-1 | Forward | AGAACATCACCTACCACTGC |
| Reverse | ATGTCCAAAGGTGCAATATC | |
| RUNX2 | Forward | CCGCACGACAACCGCACCAT |
| Reverse | CGCTCCGGCCCACAAATCTC | |
| COMP | Forward | AACGCTGAAGTCACGCTCAC |
| Reverse | GGTAGCCAAAGATGAAGCCC | |
| SOX9 | Forward | TTCATGAAGATGACCGACGA |
| Reverse | CACACCATGAAGGCGTTCAT | |
| GAPDH | Forward | AGAAGGCTGGGGCTCATTTG |
| Reverse | AGGGGCCATCCACAGTCTTC | |
| GPR87 | Forward | CTACCTTGTCTGGTAGGGGAGATG |
| Reverse | TCAGCATAGGTTATTCCTGGTTTG | |
| MYCT1 | Forward | GCCAGAAAACTTTTGGGAGGA |
| Reverse | ATCCAGTTCTGTTGAGGCCG | |
| SULT1E1 | Forward | AAAGAGGGTGATGTGGAA |
| Reverse | AAATGAGGCAGGAAGAAG | |
| KAL1 | Forward | AGCGGAGAAAGACTACGGATGG |
| Reverse | GGACACCTTTGCACTCTTCAGC | |
| ODZ2 | Forward | CCTCTCGAAATGTGACCAGCATC |
| Reverse | GCGGTAGATTCTCCTGCTGTTG | |
| MYOCD | Forward | CCACCTATGGACTCAGCCTAC |
| Reverse | CTCAGTGGCGTTGAAGAAGAG | |
| mRNA Accession | Gene Symbol | Gene Description | Gene Accession | Log2 Fold Change (Relative to hBM-MPCs) | ||
|---|---|---|---|---|---|---|
| CHA-hES15-MPCs | CHA-hNT8-MPCs | CHA-hNT5-MPCs | ||||
| NM_001302777 | - | - | - | 0.973 | 0.253 | 4.896 |
| NM_001143981 | KAL1 | Kallmann syndrome 1 sequence | NM_000216 | 1.299 | 2.210 | 4.853 |
| NR_026776 | MYCT1 | myc target 1 | NM_025107 | 1.222 | 1.887 | 4.588 |
| XR_937373 | LINC00312 | long intergenic non-protein coding RNA 312 | NR_024065 | 1.954 | 1.690 | 4.227 |
| NONHSAT052653 | MYOCD | myocardin | NM_001146312 | 2.098 | 3.046 | 4.112 |
| NM_001011655 | GPR87 | G protein-coupled receptor 87 | NM_023915 | −0.140 | 1.986 | 3.969 |
| - | - | - | - | 2.151 | 1.539 | 3.842 |
| NM_001102562 | ODZ2 | odz, odd Oz/ten-m homolog 2 (Drosophila) | NM_001122679 | 1.534 | 1.310 | 3.599 |
| NM_001271156 | - | - | - | 1.990 | 0.109 | 3.461 |
| NM_001128843 | SULT1E1 | sulfotransferase family 1E, estrogen-preferring, member 1 | NM_005420 | 0.684 | 0.989 | 3.406 |
| NM_001308394 | LMCD1 | LIM and cysteine-rich domains 1 | ENST00000157600 | 0.972 | 1.376 | 3.189 |
| NM_002196 | - | - | - | 1.290 | 0.161 | 3.097 |
| BC037342 | MYH10 | myosin, heavy chain 10, non-muscle | NM_001256012 | 1.303 | 1.462 | 2.796 |
| NM_017709 | PTGFRN | prostaglandin F2 receptor negative regulator | NM_020440 | 1.779 | 1.433 | 2.793 |
| ENST00000391302 | - | - | - | 1.157 | 1.432 | 2.733 |
| NR_121212 | EPPK1 | epiplakin 1 | ENST00000525985 | 0.160 | 1.127 | 2.583 |
| NM_001127266 | DCLK2 | doublecortin-like kinase 2 | NM_001040261 | 0.644 | 1.540 | 2.542 |
| ENST00000549438 | KRT81 | keratin 81 | NM_002281 | 0.987 | 0.356 | 2.427 |
| NM_001256155 | LOC100506610 | uncharacterized LOC100506610 | ENST00000446964 | −0.176 | 0.449 | 2.400 |
| NR_104625 | - | - | - | −0.263 | 0.693 | 2.363 |
| NM_005558 | B3GALT2 | UDP-Gal:betaGlcNAc beta 1,3-galactosyltransferase, polypeptide 2 | ENST00000367434 | −0.224 | −0.865 | 2.338 |
| NM_001084 | INHBA | inhibin, beta A | ENST00000242208 | 0.554 | 1.050 | 2.272 |
| XR_241687 | TLL1 | tolloid-like 1 | NM_012464 | −0.448 | 0.162 | 2.234 |
| NONHSAT098134 | SULT1B1 | sulfotransferase family, cytosolic, 1B, member 1 | ENST00000310613 | 0.545 | 0.829 | 2.102 |
| NONHSAT002938 | SZT2-AS1 | SZT2 antisense RNA 1 (non-protein coding) | ENST00000396885 | 0.782 | 0.995 | 2.076 |
| ENST00000432942 | - | - | - | 0.214 | −0.136 | 2.008 |
| NM_004295 | FLJ34690 | uncharacterized protein FLJ34690 | NR_034145 | 0.525 | 0.906 | 1.991 |
| NM_001017991 | HEPH | hephaestin | ENST00000519389 | 0.922 | −0.077 | 1.948 |
| NM_001143974 | - | - | - | −0.001 | 0.336 | 1.944 |
| NM_020726 | ADAMTS16 | ADAM metallopeptidase with thrombospondin type 1 motif, 16 | NM_139056 | −0.633 | −0.084 | 1.856 |
| ENST00000375094 | CTGF | connective tissue growth factor | ENST00000367976 | 0.346 | 0.607 | 1.806 |
| GENSCAN00000011063 | PPME1 | protein phosphatase methylesterase 1 | ENST00000328257 | −0.014 | 0.263 | 1.738 |
| NM_000829 | FAT3 | FAT tumor suppressor homolog 3 (Drosophila) | NM_001008781 | 0.369 | 0.634 | 1.726 |
| NM_021915 | GADD45B | growth arrest and DNA-damage-inducible, beta | NM_015675 | 0.485 | 0.596 | 1.703 |
| NM_001101421 | ANO4 | anoctamin 4 | NM_178826 | 0.273 | 0.362 | 1.659 |
| NM_003804 | C5orf46 | chromosome 5 open reading frame 46 | NM_206966 | 0.496 | −0.157 | 1.591 |
| NM_001009185 | GCNT4 | glucosaminyl (N-acetyl) transferase 4, core 2 | NM_016591 | 0.024 | −0.001 | 1.581 |
| NM_203425 | ITGB3 | integrin, beta 3 (platelet glycoprotein IIIa, antigen CD61) | NM_000212 | −0.963 | 0.126 | 1.470 |
| NM_006168 | SEL1L3 | sel-1 suppressor of lin-12-like 3 (C. elegans) | NM_015187 | −0.007 | 0.069 | 1.426 |
| NM_004061 | KCNIP1 | Kv channel interacting protein 1 | NM_001034837 | 0.050 | −0.247 | 1.388 |
| NM_001412 | DIAPH2 | diaphanous homolog 2 (Drosophila) | NM_007309 | 0.150 | 0.166 | 1.329 |
| NR_001553 | ACSL4 | acyl-CoA synthetase long-chain family member 4 | NM_022977 | 0.313 | 0.283 | 1.328 |
| NONHSAT138126 | LOC389906 | zinc finger protein 839 pseudogene | NR_034031 | 0.220 | 0.073 | 1.263 |
| ENST00000365229 | - | - | - | −0.130 | −0.083 | 1.220 |
| NM_030661 | MGAM | maltase-glucoamylase (alpha-glucosidase) | ENST00000549489 | −0.150 | 0.068 | 1.219 |
| XM_011519004 | MAMDC2 | MAM domain containing 2 | NM_153267 | 0.118 | −0.130 | 1.174 |
| NM_001300974 | GDF15 | growth differentiation factor 15 | NM_004864 | −0.074 | −0.060 | 1.132 |
| ENST00000362934 | TRAJ59 | T cell receptor alpha joining 59 (non-functional) | ENST00000390480 | −0.452 | 0.088 | 1.124 |
| XR_427224 | ITGA4 | integrin, alpha 4 (antigen CD49D, alpha 4 subunit of VLA-4 receptor) | NM_000885 | −0.292 | −0.566 | 1.029 |
| NM_031273 | - | - | - | −0.206 | −0.849 | 1.019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, E.-Y.; Yoon, Y.-J.; Lee, J.E.; Shim, S.H.; Park, G.H.; Lee, D.R. Identification of Putative Markers That Predict the In Vitro Senescence of Mesenchymal Progenitor Cells. Cells 2021, 10, 1301. https://doi.org/10.3390/cells10061301
Shin E-Y, Yoon Y-J, Lee JE, Shim SH, Park GH, Lee DR. Identification of Putative Markers That Predict the In Vitro Senescence of Mesenchymal Progenitor Cells. Cells. 2021; 10(6):1301. https://doi.org/10.3390/cells10061301
Chicago/Turabian StyleShin, Eun-Young, Yeo-Joon Yoon, Jeoung Eun Lee, Sung Han Shim, Gene Hong Park, and Dong Ryul Lee. 2021. "Identification of Putative Markers That Predict the In Vitro Senescence of Mesenchymal Progenitor Cells" Cells 10, no. 6: 1301. https://doi.org/10.3390/cells10061301
APA StyleShin, E.-Y., Yoon, Y.-J., Lee, J. E., Shim, S. H., Park, G. H., & Lee, D. R. (2021). Identification of Putative Markers That Predict the In Vitro Senescence of Mesenchymal Progenitor Cells. Cells, 10(6), 1301. https://doi.org/10.3390/cells10061301