
Spatiotemporal Patterns of Menin Localization in Developing Murine Brain: Co-Expression with the Elements of Cholinergic Synaptic Machinery

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals: Brain Slices and Neuronal Cell Culture
2.2. Immunocytochemistry and Immunohistochemistry
2.3. Quantitative PCR (qPCR)
2.4. GFP-Cre_AAV Stereotaxic Injections in Homozygous Floxed MEN1 Mice
2.5. Microscopy
2.6. Experimental Design and Statistical Analysis
3. Results
3.1. Specificity of Menin Expression Pattern
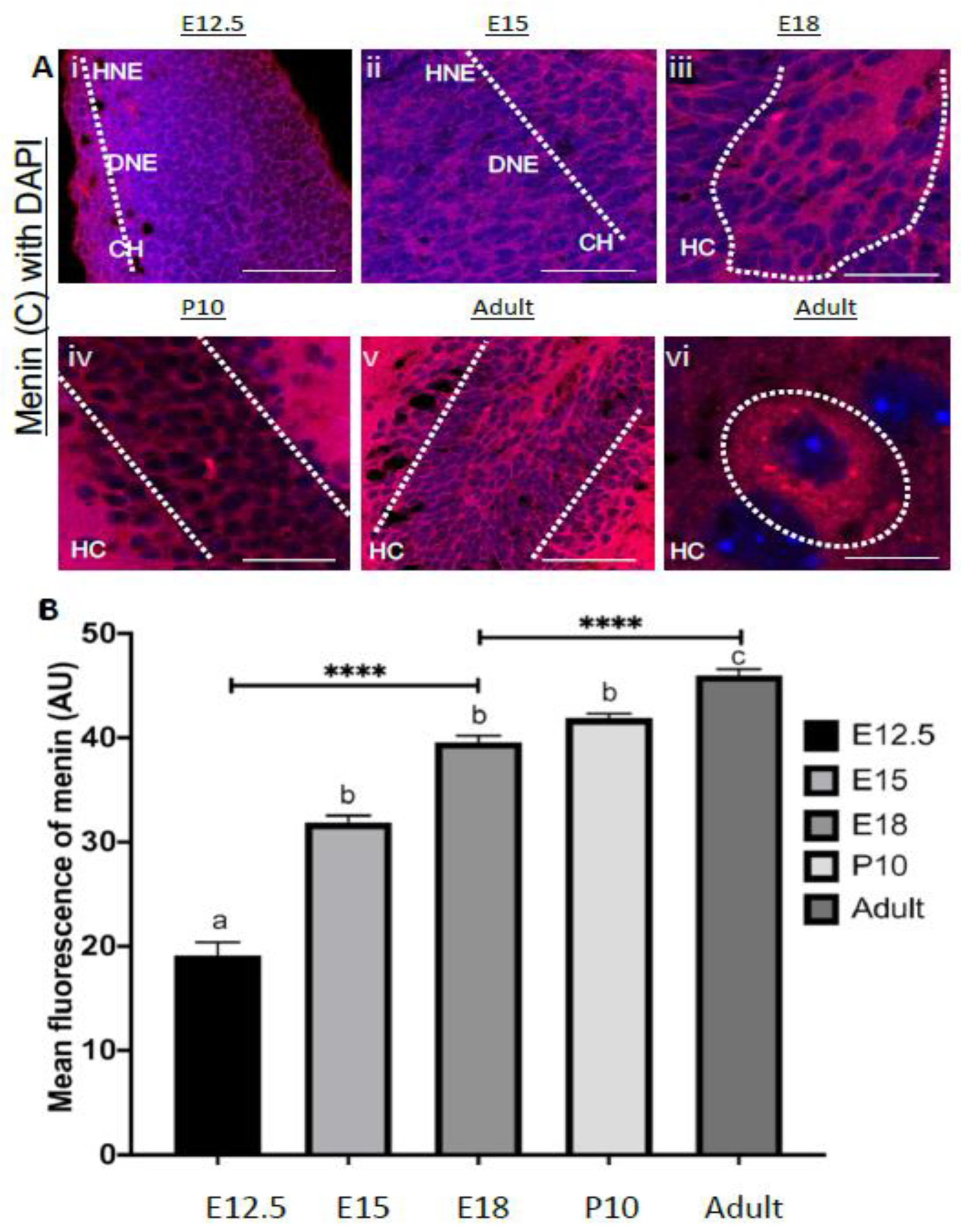
3.2. Spatiotemporal Specific Localization of MEN1 Gene and Menin Protein Fragments in Neurons
3.3. Neuron Specific Expression of α-C-terminal Menin in Hippocampal Neurons (In-Situ)
3.4. Astrocytes—Specific Expression of α-C-terminal Menin in Mouse Brain (In Situ) and (In Vitro)
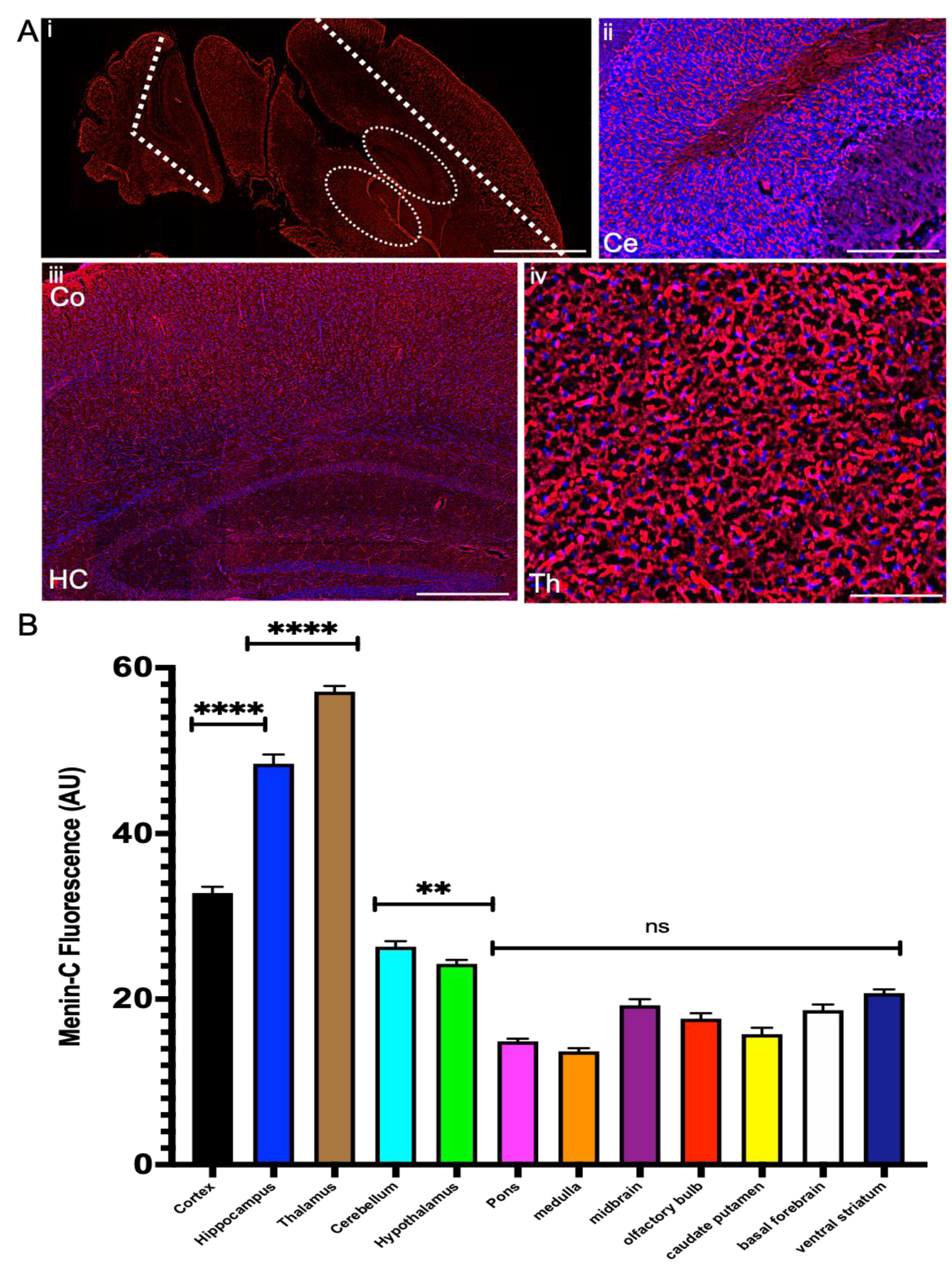
3.5. Menin Expression Was Highly Localized to Telencephalon and Diencephalon during Embryonic Developmental Period in Mouse Brain
3.6. Menin Exhibited Highest Expression in Thalamus, Hippocampus, Cortex and Cerebellum in Both P10 and Adult Mouse Brains
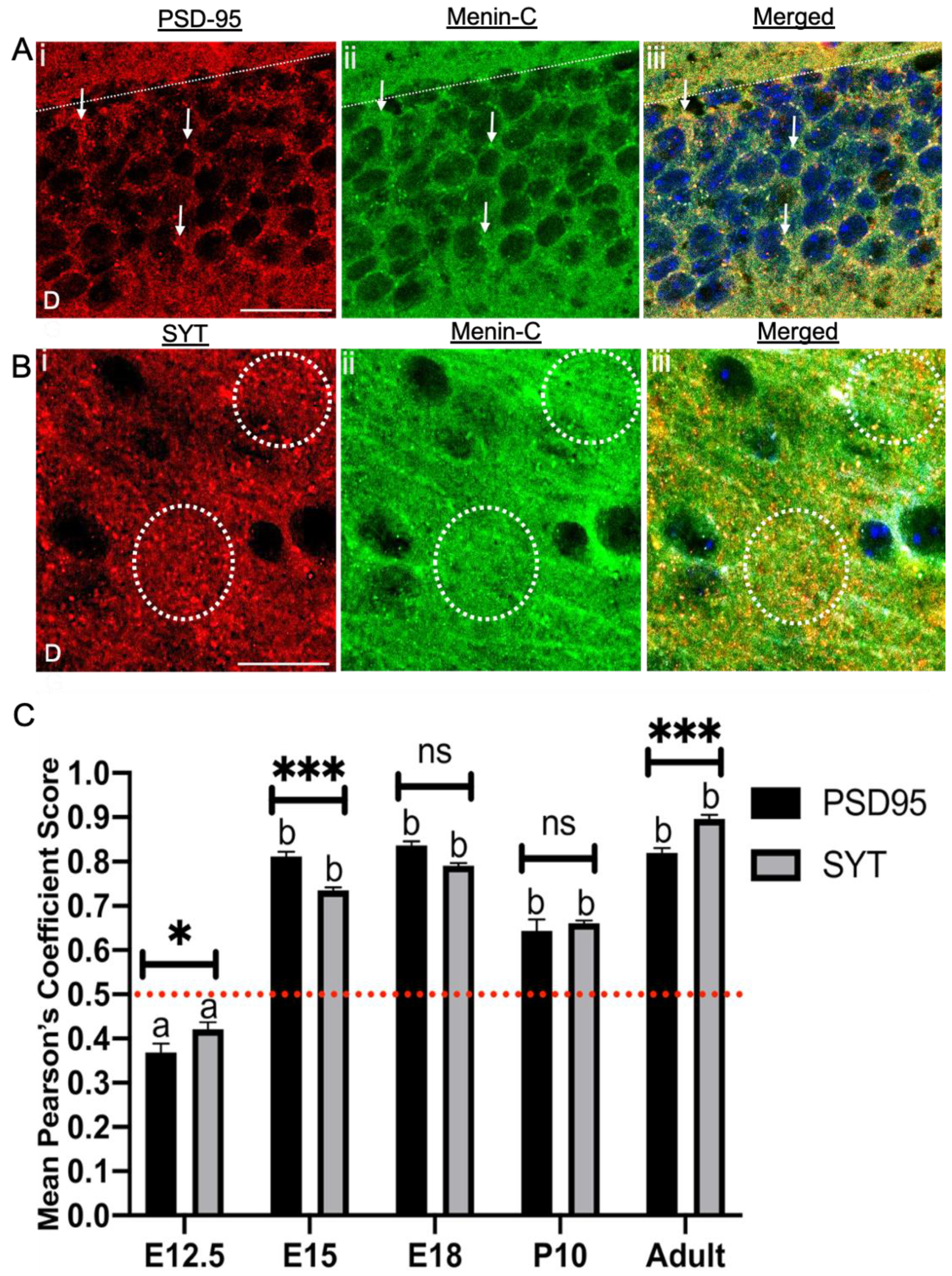
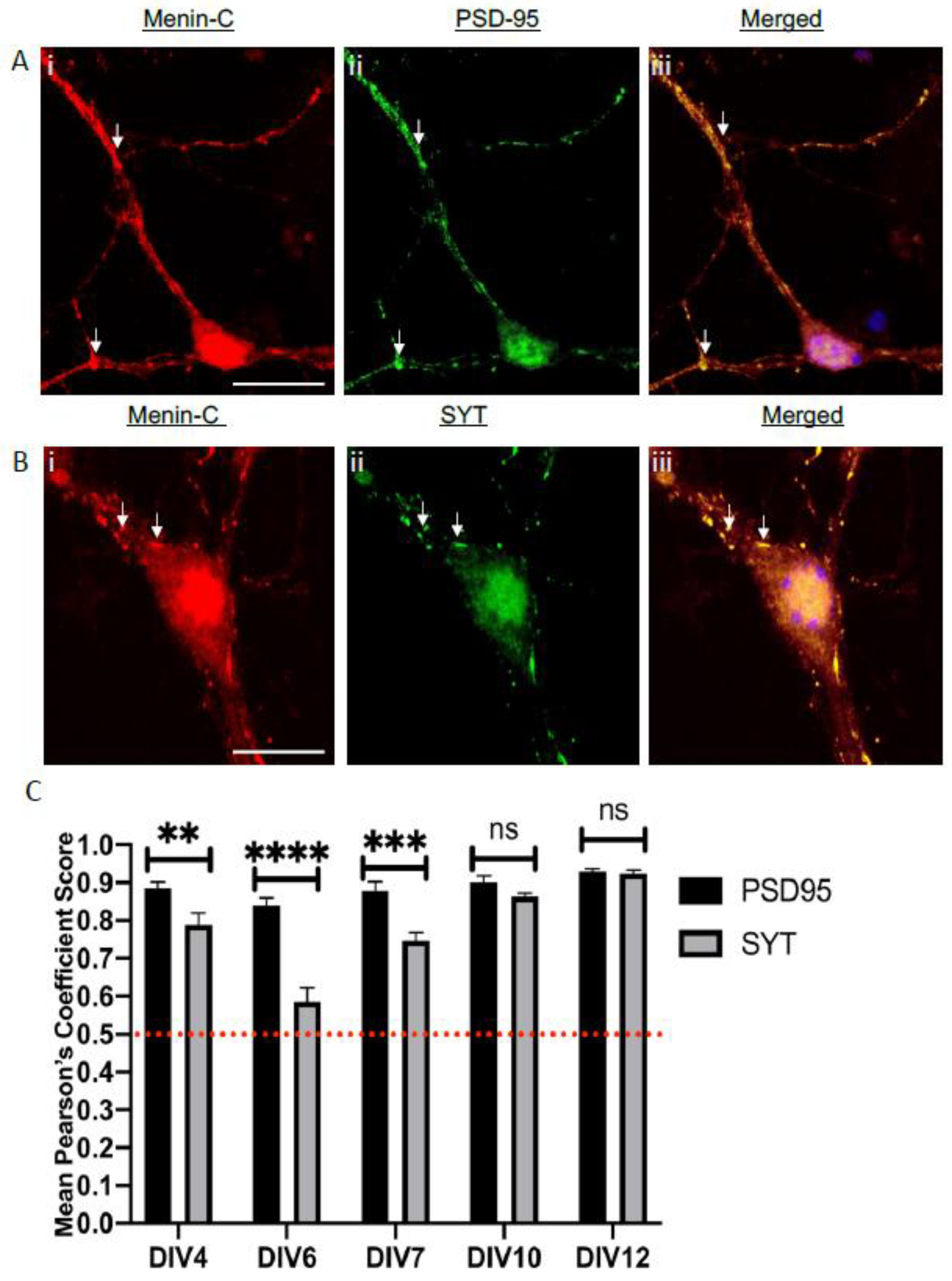
3.7. Menin Was Colocalized with Both Pre- and Postsynaptic Protein in a Whole Mouse Brain
3.8. Menin Expression Was Significantly Aligned Along Excitatory versus Inhibitory Receptors in an Adult Mouse Brain Topography
3.9. Menin Colocalizes with Nicotinic, Cholinergic Receptors (nAChR) Specific Subunits α5 and α7 in a Spatiotemporal Manner
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATN | anterior thalamic nuclei |
| BL | Basolateral amygdaloid nucleus |
| BM | Basomedial amygdaloid nucleus |
| CA1 | Cornu Ammonis-1 |
| CA2 | Cornu Ammonis-2 |
| CA3 | Cornu Ammonis-3 |
| Che | choroid plexus |
| CH | cortical hem |
| Cp | cortical plate |
| Cpu | Caudate putamen |
| DG | Dentate gyrus |
| Dgn | Dentate gyrus neuroepithelium |
| DP | dorsal plate |
| DNE | dentate neuroepithelium |
| Ec | External capsule |
| Fpc | preculminate fissure |
| Fpl | posterolateral fissure |
| Fppd | prepyramidal fissure |
| Fpr | primary fissure |
| Fsec | secondary fissure |
| FStr | Fundus Striati |
| HNE | hippocampal neuroepithelium |
| IZ | Intermediate zone |
| La | Lateral nucleus of amygdala |
| LGE | lateral ganglionic eminence |
| LV | lateral ventricle |
| Met | mesencephalic tegmentum |
| MGE | medial ganglionic eminence |
| MP | medioposterior nucleus |
| MV | medioventral |
| ncx | neocortex |
| Par | Parietal cortex |
| PIR | piriform cortex |
| PP | preplate |
| PSE | paraseptal subpallial sector |
| PT | pretectum |
| PV | paraventricular |
| Rs | retrosplenial cortex |
| SP | Subplate |
| SPT | septum |
| Sta | Striatum |
| SVZ | subventricular zone |
| Th | thalamus |
| v3 | third ventricle |
| vThal | ventral thalamus |
| VZ | ventral zone |
References
- Thakker, R.V.; Newey, P.J.; Walls, G.V.; Bilezikian, J.; Dralle, H.; Ebeling, P.R.; Melmed, S.; Sakurai, A.; Tonelli, F.; Brandi, M.L.; et al. Clinical practice guidelines for multiple endocrine neoplasia type 1 (MEN1). J. Clin. Endocrinol. Metab. 2012, 97, 2990–3011. [Google Scholar] [PubMed]
- Stewart, C.; Parente, F.; Piehl, F.; Farnebo, F.; Quincey, D.; Silins, G.; Bergman, L.; Carle, G.F.; Lemmens, I.; Grimmond, S.; et al. Characterization of the mouse Men1 gene and its expression during development. Oncogene 1998, 17, 2485. [Google Scholar] [PubMed]
- Zhuang, K.; Huang, C.; Leng, L.; Zheng, H.; Gao, Y.; Chen, G.; Ji, Z.; Sun, H.; Hu, Y.; Wu, D.; et al. Neuron-specific menin deletion leads to synaptic dysfunction and cognitive impairment by modulating p35 expression. Cell Rep. 2018, 24, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Guru, S.C.; Prasad, N.B.; Shin, E.J.; Hemavathy, K.; Lu, J.; Ip, Y.T.; Agarwal, S.K.; Marx, S.J.; Spiegel, A.M.; Collins, F.S.; et al. Characterization of a MEN1 ortholog from Drosophila melanogaster. Gene 2001, 263, 31. [Google Scholar] [CrossRef]
- Getz, A.M.; Visser, F.; Bell, E.M.; Xu, F.; Flynn, N.M.; Zaidi, W.; Syed, N.I. Two proteolytic fragments of menin coordinate the nuclear transcription and postsynaptic clustering of neurotransmitter receptors during synaptogenesis between Lymnaea neurons. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Getz, A.M.; Xu, F.; Visser, F.; Persson, R.; Syed, N.I. Tumor suppressor menin is required for subunit-specific nAChR α5 transcription and nAChR-dependent presynaptic facilitation in cultured mouse hippocampal neurons. Sci. Rep. 2017, 7, 1. [Google Scholar]
- Murai, M.J.; Chruszcz, M.; Reddy, G.; Grembecka, J.; Cierpicki, T. Crystal structure of menin reveals binding site for mixed lineage leukemia (MLL) protein. J. Biol. Chem. 2011, 286, 31742–31748. [Google Scholar] [CrossRef] [PubMed]
- Matkar, S.; Thiel, A.; Hua, X. Menin: A scaffold protein that controls gene expression and cell signaling. Trends Biochem. Sci. 2013, 38, 394–402. [Google Scholar] [CrossRef]
- Xu, S.; Wu, H.; Wang, X.; Shen, X.; Guo, X.; Shen, R.; Wang, F. Tumor suppressor menin mediates peripheral nerve injury-induced neuropathic pain through potentiating synaptic plasticity. Neuroscience 2012, 223, 473–485. [Google Scholar] [CrossRef]
- Van Kesteren, R.E.; Syed, N.I.; Munno, D.W.; Bouwman, J.; Feng, Z.P.; Geraerts, W.P.; Smit, A.B. Synapse formation between central neurons requires postsynaptic expression of the MEN1 tumor suppressor gene. J. Neurosci. 2001, 21, 1–5. [Google Scholar] [CrossRef]
- Flynn, N.; Getz, A.; Visser, F.; Janes, T.A.; Syed, N.I. Menin: A tumor suppressor that mediates postsynaptic receptor expression and synaptogenesis between central neurons of Lymnaea stagnalis. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Park, J.J.; Cunningham, M.G. Thin sectioning of slice preparations for immunohistochemistry. J. Vis. Exp. 2007. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Tseng, Y.C.; Chu, S.W. High spatio-temporal-resolution detection of chlorophyll fluorescence dynamics from a single chloroplast with confocal imaging fluorometer. Plant Methods 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Lein, E.S.; Hawrylycz, M.J.; Ao, N.; Ayres, M.; Bensinger, A.; Bernard, A.; Boe, A.F.; Boguski, M.S.; Brockway, K.S.; Byrnes, E.J.; et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature 2007, 445, 168. [Google Scholar] [CrossRef] [PubMed]
- Bolte, S.; Cordelieres, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Yap, B.W.; Sim, C.H. Comparisons of various types of normality tests. J. Stat. Comput. Simul. 2011, 81, 2141–2155. [Google Scholar] [CrossRef]
- Dang, C.; Sodt, A.; Lau, C.; Youngstrom, B..; Ng, L.; Kuan, L.; Pathak, S.; Jones, A.; Hawrylycz, M. The Allen Brain Atlas: Delivering Neuroscience to the Web on a Genome Wide Scale. In Data Integration in the Life Sciences. DILS 2007. Lecture Notes in Computer Science; Springer: Berlin/Heidelberg, Germany; Volume 4544, pp. 17–26.
- Bacaj, T.; Wu, D.; Burre, J.; Malenka, R.C.; Liu, X.; Sudhof, T.C. Synaptotagmin-1 and -7 are redundantly essential for maintaining the capacity of the readily-releasable pool of synaptic vesicles (SYT-11 and SYT-17 maintain the RRP size). PLoS Biol. 2015, 13, e1002267. [Google Scholar] [CrossRef] [PubMed]
- Crupi, R.; Impellizzeri, D.; Cuzzocrea, S. Role of metabotropic glutamate receptors in neurological disorders. Front. Mol. Neurosci. 2019, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-K.; Takamiya, K.; He, K.; Song, L.; Huganir, R.L. Specific roles of AMPA receptor subunit GluR1 (GluA1) phosphorylation sites in regulating synaptic plasticity in the CA1 region of hippocampus. J. Neurophysiol. 2010, 103, 479–489. [Google Scholar] [CrossRef]
- Pinard, A.; Seddik, R.; Bettler, B. GABAB receptors: Physiological functions and mechanisms of diversity. Adv. Pharmacol. 2010, 58, 231–255. [Google Scholar] [PubMed]
- Yakel, J. Cholinergic receptors: Functional role of nicotinic ACh receptors in brain circuits and disease. Pflügers Arch. Eur. J. Physiol. 2013, 465, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Couturier, S.; Bertrand, D.; Matter, J.M.; Hernandez, M.C.; Bertrand, S.; Millar, N.; Valera, S.; Barkas, T.; Ballivet, M. A neuronal nicotinic acetylcholine receptor subunit (α7) is developmentally regulated and forms a homo-oligomeric channel blocked by α-BTX. Neuron 1990, 5, 847–856. [Google Scholar] [CrossRef]
- Parri, H.R.; Hernandez, C.M.; Dineley, K.T. Research update: Alpha7 nicotinic acetylcholine receptor mechanisms in Alzheimer’s disease. Biochem. Pharmacol. 2011, 82, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Foucault-Fruchard, L.; Antier, D. Therapeutic potential of alpha 7 nicotinic receptor agonists to regulate neuroinflammation in neurodegenerative diseases. Neural Regen. Res. 2017, 12, 1418–1421. [Google Scholar] [CrossRef]
- Fierro, J.; Haynes, D.R.; Washbourne, P. 4.1 Ba is necessary for glutamatergic synapse formation in the sensorimotor circuit of developing zebrafish. PLoS ONE 2018, 13, e0205255. [Google Scholar] [CrossRef]
- Brzezinka, K.; Nevedomskaya, E.; Lesche, R.; Haegebarth, A.; Ter Laak, A.; Fernandez-Montalvan, A.E.; Eberspaecher, Y.; Werbeck, N.D.; Moenning, U.; Siegel, S.; et al. Characterization of the Menin-MLL interaction as therapeutic cancer target. Cancers 2020, 12, 201. [Google Scholar] [CrossRef]
- Leng, L.; Zhuang, K.; Liu, Z.; Huang, C.; Gao, Y.; Chen, G.; Lin, H.; Hu, Y.; Wu, D.; Shi, M.; et al. Menin deficiency leads to depressive-like behaviors in mice by modulating astrocyte-mediated neuroinflammation. Neuron 2018, 100, 551–563. [Google Scholar] [CrossRef]
- Gerrow, K.; Romorini, S.; Nabi, S.M.; Colicos, M.A.; Sala, C.; El-Husseini, A. A preformed complex of postsynaptic proteins is involved in excitatory synapse development. Neuron 2006, 49, 547–562. [Google Scholar] [CrossRef]
- Urban, N.; Guillemot, F. Neurogenesis in the embryonic and adult brain: Same regulators, different roles. Front. Cell. Neurosci. 2014, 8, 396. [Google Scholar] [CrossRef]
- Goetz, M.; Huttner, W.B. The cell biology of neurogenesis. Nat. Rev. Mol. Cell Biol. 2005, 6, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Paridaen, J.; Huttner, W.B. Neurogenesis during development of the vertebrate central nervous system. EMBO Rep. 2014, 15, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Fuentealba, L.C.; Obernier, K.; Alvarez-Buylla, A. Adult neural stem cells bridge their niche. Cell Stem Cell 2012, 10, 698–708. [Google Scholar] [CrossRef]
- Holzman, D.C. MRI-based atlas of the developing mouse brain debuts. Environ. Health Perspect. 2011, 119, A165. [Google Scholar] [CrossRef]
- Pierfelice, T.; Alberi, L.; Gaiano, N. Notch in the vertebrate nervous system: An old dog with new tricks. Neuron 2011, 69, 840–855. [Google Scholar] [CrossRef] [PubMed]
- Harrison-Uy, S.J.; Pleasure, S.J. Wnt signaling and forebrain development. Cold Spring Harb. Perspect. Biol. 2012, 4, a008094. [Google Scholar] [CrossRef]
- Saade, M.; Gutierrez-Vallejo, I.; Le Dreau, G.; Rabadan, M.A.; Miguez, D.G.; Buceta, J.; Marti, E. Sonic hedgehog signaling switches the mode of division in the developing nervous system. Cell Rep. 2013, 4, 492–503. [Google Scholar] [CrossRef]
- Kaji, H. Menin and bone metabolism. J. Bone Miner. Metab. 2012, 30, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Jingbo, A.; Wang, M.; Farley, S.; Lee, L.Y.; Lee, L.C.; Sawicki, M.P. Menin promotes the Wnt signaling pathway in pancreatic endocrine cells. Mol. Cancer Res. 2008, 6, 1894–1907. [Google Scholar] [CrossRef]
- Gurung, B.; Feng, Z.; Iwamoto, D.V.; Thiel, A.; Jin, G.; Fan, C.-M.; Ng, J.M.Y.; Curran, T.; Hua, X. Menin epigenetically represses Hedgehog signaling in MEN1 tumor syndrome. Cancer Res. 2013, 73, 2650–2658. [Google Scholar] [CrossRef]
- Matas-Rico, E.; van Veen, M.; Moolenaar, W.H. Neuronal differentiation through GPI-anchor cleavage. Cell Cycle 2017, 16, 388–389. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Stoeckli, E.T. Understanding axon guidance: Are we nearly there yet? Development 2018, 145. [Google Scholar] [CrossRef] [PubMed]
- Südhof, T.C. Towards an understanding of synapse formation. Neuron 2018, 100, 276–293. [Google Scholar] [CrossRef] [PubMed]
- Sabo, S.L.; Gomes, R.A.; McAllister, A.K. Formation of presynaptic terminals at predefined sites along axons. J. Neurosci. 2006, 26, 10813–10825. [Google Scholar] [CrossRef] [PubMed]
- Batool, S.; Raza, H.; Zaidi, J.; Riaz, S.; Hasan, S.; Syed, N.I. Synapse formation: From cellular and molecular mechanisms to neurodevelopmental and neurodegenerative disorders. J. Neurophysiol. 2019, 121, 1381–1397. [Google Scholar] [CrossRef]
- Sytnyk, V.; Leschnynska, I.; Dityatev, A.; Schachner, M. Trans-Golgi network delivery of synaptic proteins in synaptogenesis. J. Cell Sci. 2004, 117, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Lippa, C.F. Review: Disruption of the postsynaptic density in Alzheimer’s disease and other neurodegenerative dementias. Am. J. Alzheimer Dis. Other Dement. 2010, 25, 547–555. [Google Scholar] [CrossRef]
- Wang, C.; Kang, X.; Zhou, L.; Chai, Z.; Wu, Q.; Huang, R.; Xu, H.; Hu, M.; Sun, X.; Sun, S.; et al. Synaptotagmin-11 is a critical mediator of parkin-linked neurotoxicity and Parkinson’s disease-like pathology. Nat. Commun. 2018, 9, 81. [Google Scholar] [CrossRef]
- Ohrfelt, A.; Brinkmalm, A.; Dumurgier, J.; Brinkmalm, G.; Hansson, O.; Zetterberg, H.; Bouaziz-Amar, E.; Hugon, J.; Paquet, C.; Blennow, K. The pre-synaptic vesicle protein synaptotagmin is a novel biomarker for Alzheimer’s disease. Alzheimer Res. Ther. 2016, 8. [Google Scholar] [CrossRef]
- Epp, J.R.; Chow, C.; HGalea, L.A.M. Hippocampus-dependent learning influences hippocampal neurogenesis. Front. Neurosci. 2013, 7, 57. [Google Scholar] [CrossRef]
- Berdugo-Vega, G.; Arias-Gil, G.; Lopez-Fernandez, A.; Artegiani, B.; Wasielewska, J.M.; Lee, C.C.; Lippert, M.T.; Kempermann, G.; Takagaki, K.; Calegari, F. Increasing neurogenesis refines hippocampal activity rejuvenating navigational learning strategies and contextual memory throughout life. Nat. Commun. 2020, 11, 135. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Sun, D. GABA receptors in brain development, function, and injury. Metab. Brain Dis. 2015, 30, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Luo, D.; Yang, J.; Xu, X.Y.; Zhu, B.L.; Wang, X.F.; Yan, Z.; Chen, G.J. Modulation of AMPA receptor mediated current by nicotinic acetylcholine receptor in layer I neurons of rat prefrontal cortex. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef]
- Broide, R.S.; Winzer-Serhan, U.H.; Chen, Y.; Leslie, F.M. Distribution of α7 nicotinic acetylcholine receptor subunit mRNA in the developing mouse. Front. Neuroanat. 2019, 13, 76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, C.; Miao, H.; Gong, Z.H.; Nordberg, A. Postnatal changes of nicotinic acetylcholine receptor alpha 2, alpha 3, alpha 4, alpha 7 and beta 2 subunits genes expression in rat brain. Int. J. Dev. Neurosci. 1998, 16, 507–518. [Google Scholar] [CrossRef]
- Cheng, Q.; Yakel, J.L. The effect of α7 nicotinic receptor activation on glutamatergic transmission in the hippocampus. Biochem. Pharmacol. 2015, 97, 439–444. [Google Scholar] [CrossRef]
- Campbell, N.R.; Fernandes, C.C.; Halff, A.W.; Berg, D.K. Endogenous signaling through α7-containing nicotinic receptors promotes maturation and integration of adult-born neurons in the hippocampus. J. Neurosci. 2010, 30, 8734–8744. [Google Scholar] [CrossRef]
- Lozada, A.F.; Wang, X.; Guonko, N.V.; Massey, K.A.; Duan, J.; Liu, Z.; Berg, D.K. Glutamatergic synapse formation is promoted by alpha 7-containing nicotinic acetylcholine receptors. J. Neurosci. 2012, 32, 7651–7661. [Google Scholar] [CrossRef]
- Terry, A.V.; Buccafusco, J.J. The cholinergic hypothesis of age and Alzheimer’s disease-related cognitive deficits: Recent challenges and their implications for novel drug development. J. Pharmacol. Exp. Ther. 2003, 306, 821–827. [Google Scholar] [CrossRef]
- Dani, J.A.; Bertrand, D. Nicotinic acetylcholine receptors and nicotinic cholinergic mechanisms of the central nervous system. Ann. Rev. Pharmacol. Toxicol. 2007, 47, 699–729. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batool, S.; Zaidi, J.; Akhter, B.; Ulfat, A.K.; Visser, F.; Syed, N.I. Spatiotemporal Patterns of Menin Localization in Developing Murine Brain: Co-Expression with the Elements of Cholinergic Synaptic Machinery. Cells 2021, 10, 1215. https://doi.org/10.3390/cells10051215
Batool S, Zaidi J, Akhter B, Ulfat AK, Visser F, Syed NI. Spatiotemporal Patterns of Menin Localization in Developing Murine Brain: Co-Expression with the Elements of Cholinergic Synaptic Machinery. Cells. 2021; 10(5):1215. https://doi.org/10.3390/cells10051215
Chicago/Turabian StyleBatool, Shadab, Jawwad Zaidi, Basma Akhter, Anosha Kiran Ulfat, Frank Visser, and Naweed I. Syed. 2021. "Spatiotemporal Patterns of Menin Localization in Developing Murine Brain: Co-Expression with the Elements of Cholinergic Synaptic Machinery" Cells 10, no. 5: 1215. https://doi.org/10.3390/cells10051215
APA StyleBatool, S., Zaidi, J., Akhter, B., Ulfat, A. K., Visser, F., & Syed, N. I. (2021). Spatiotemporal Patterns of Menin Localization in Developing Murine Brain: Co-Expression with the Elements of Cholinergic Synaptic Machinery. Cells, 10(5), 1215. https://doi.org/10.3390/cells10051215