
Increased HERV-K(HML-2) Transcript Levels Correlate with Clinical Parameters of Liver Damage in Hepatitis C Patients

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Peripheral Blood Mononuclear Cells (PBMC)
2.2. Cryoconservation and Thawing of PBMC
2.3. Isolation of RNA Using a Trizol Gradient
2.4. DNAse Treatment
2.5. First Strand cDNA Synthesis Using Random Hexamer Primers
2.6. Quantitative Real-Time PCR
2.7. One-Step Quantitative Real-Time PCR
2.8. Analysis of Quantitative Real-Time PCR
2.9. Statistical Analysis
3. Results
3.1. Cohort Information
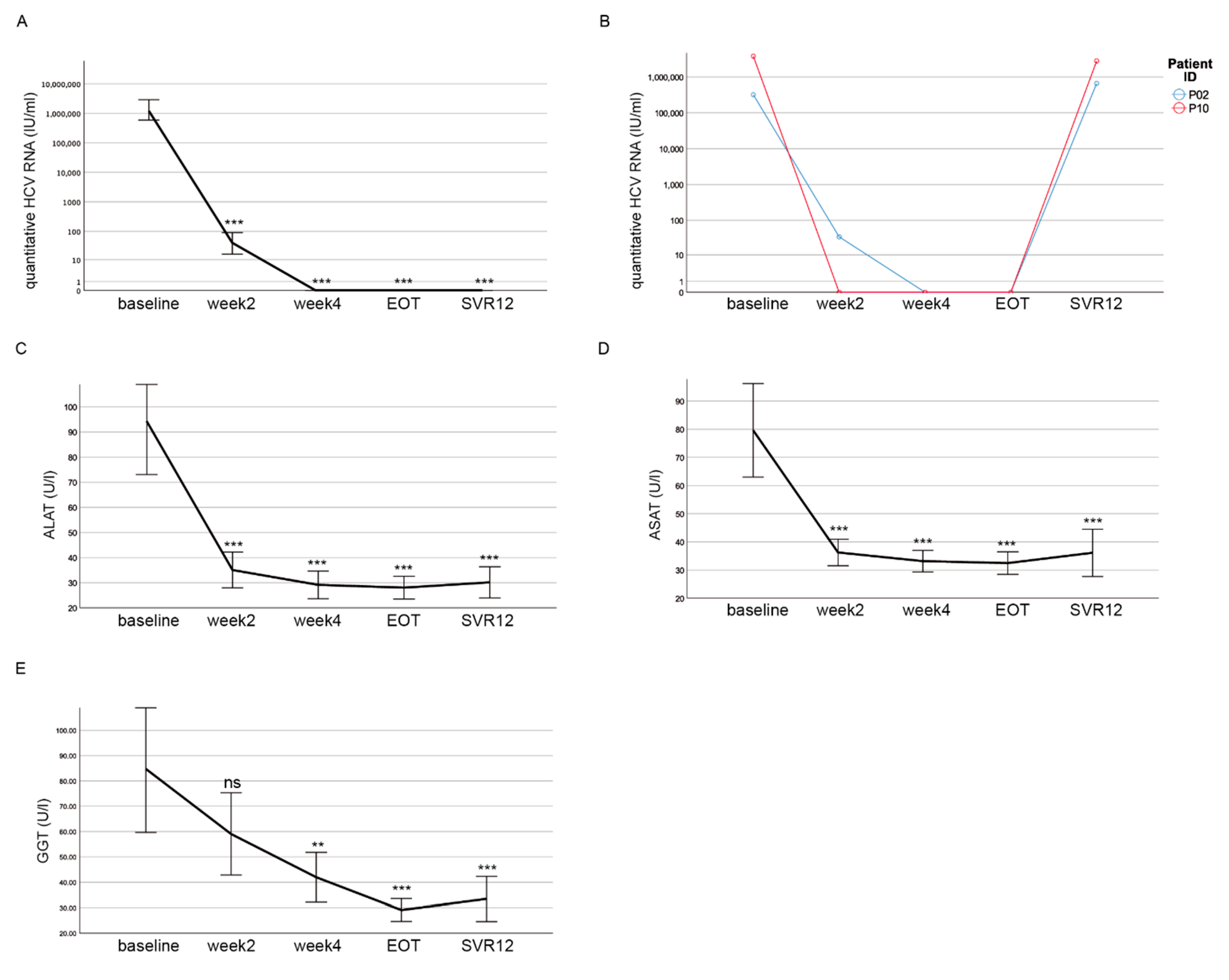
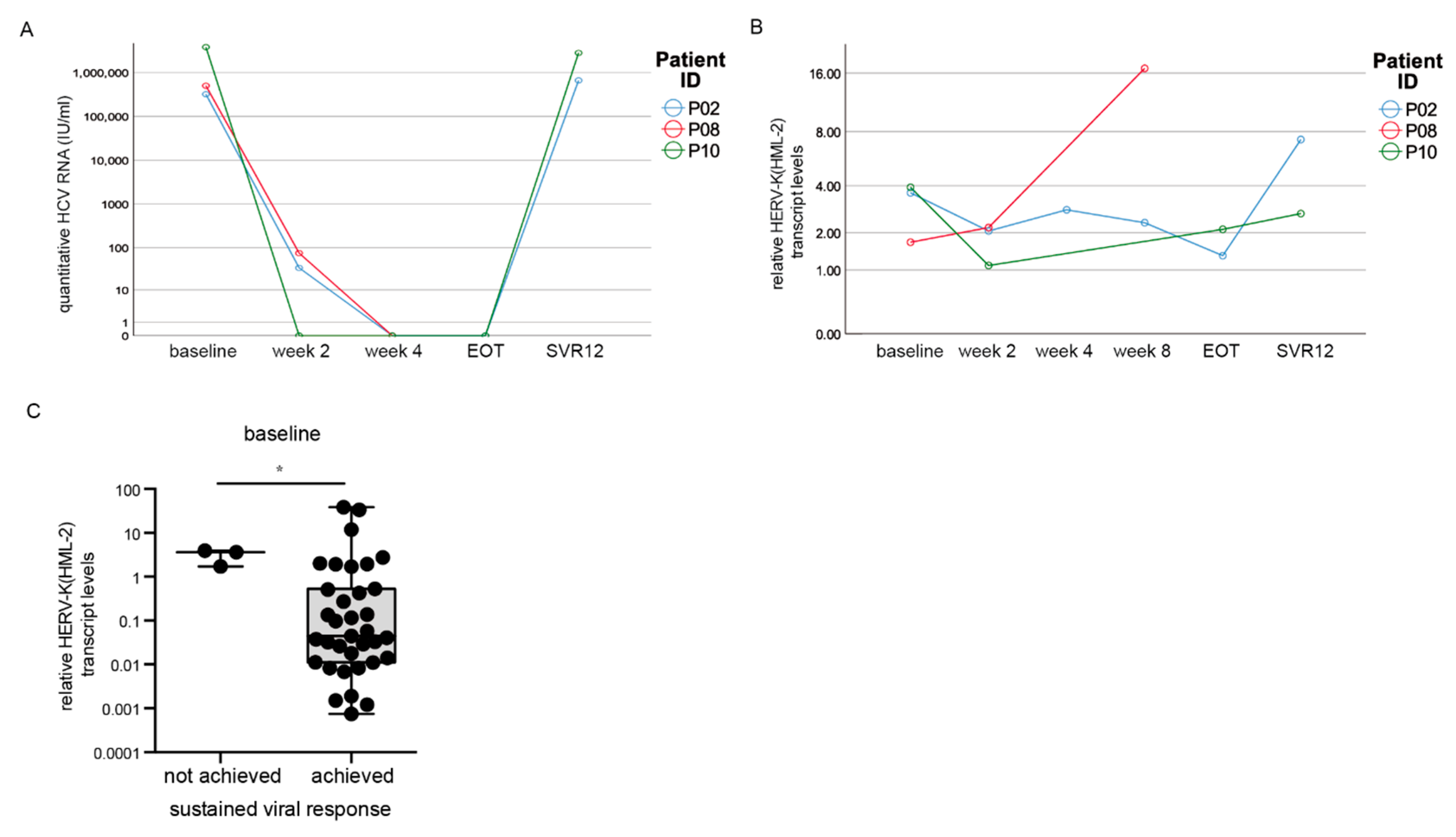
3.2. DAA Treatment Results in Decreased HCV RNA Levels
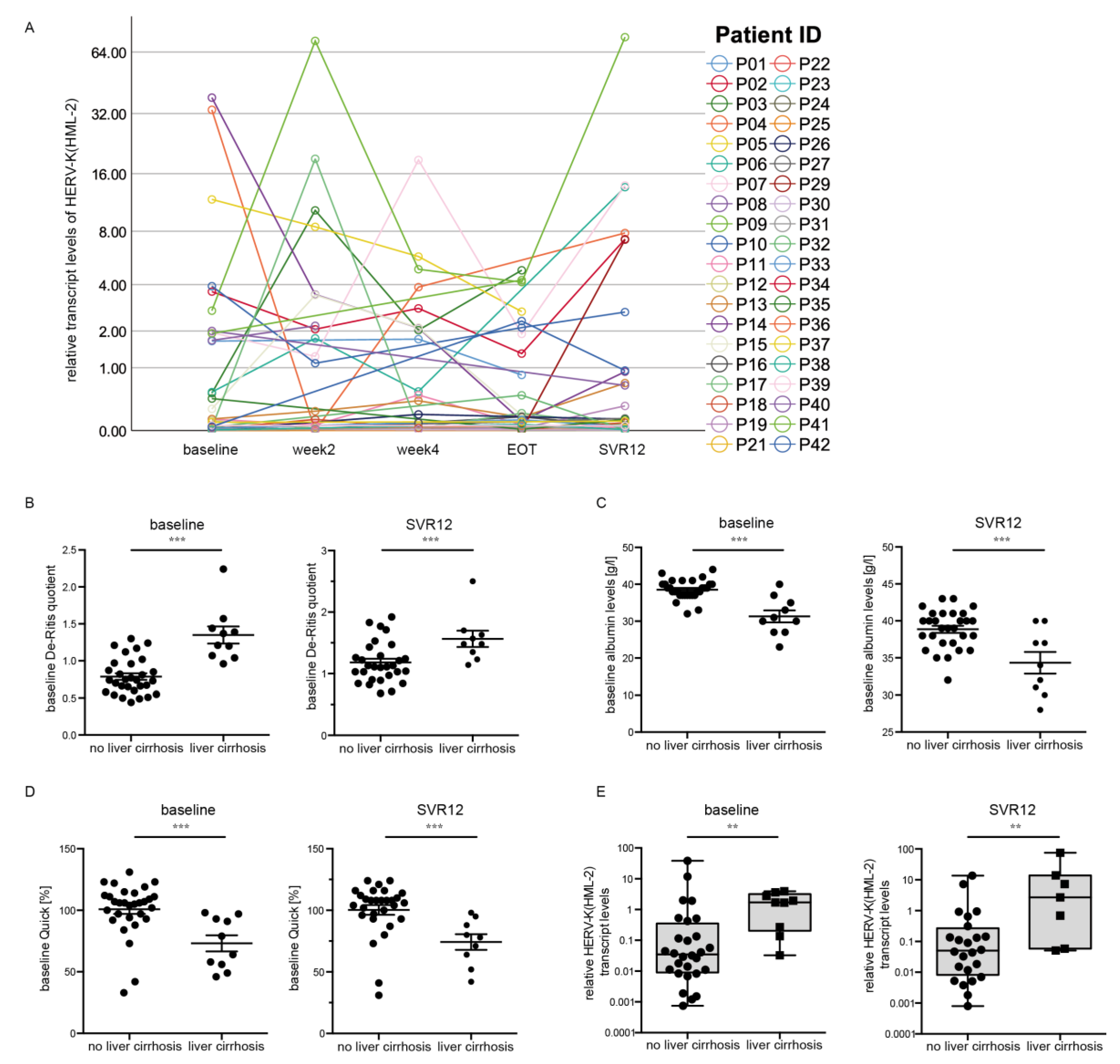
3.3. HERV-K(HML-2) Transcript Levels Are Increased in Patients with Liver Cirrhosis
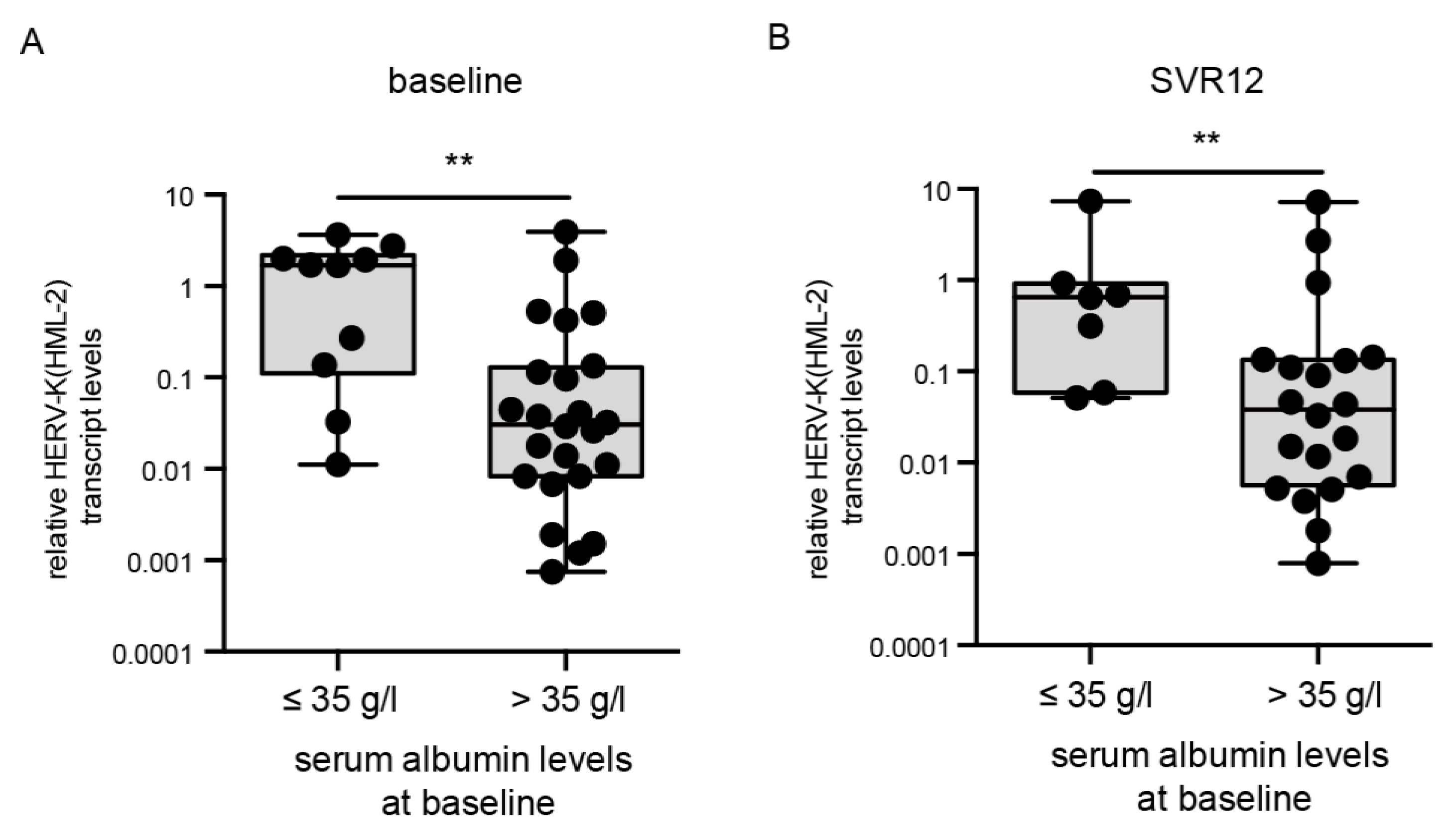
3.4. HERV-K(HML-2) Transcript Levels Are Elevated in Patients with Low Albumin Levels
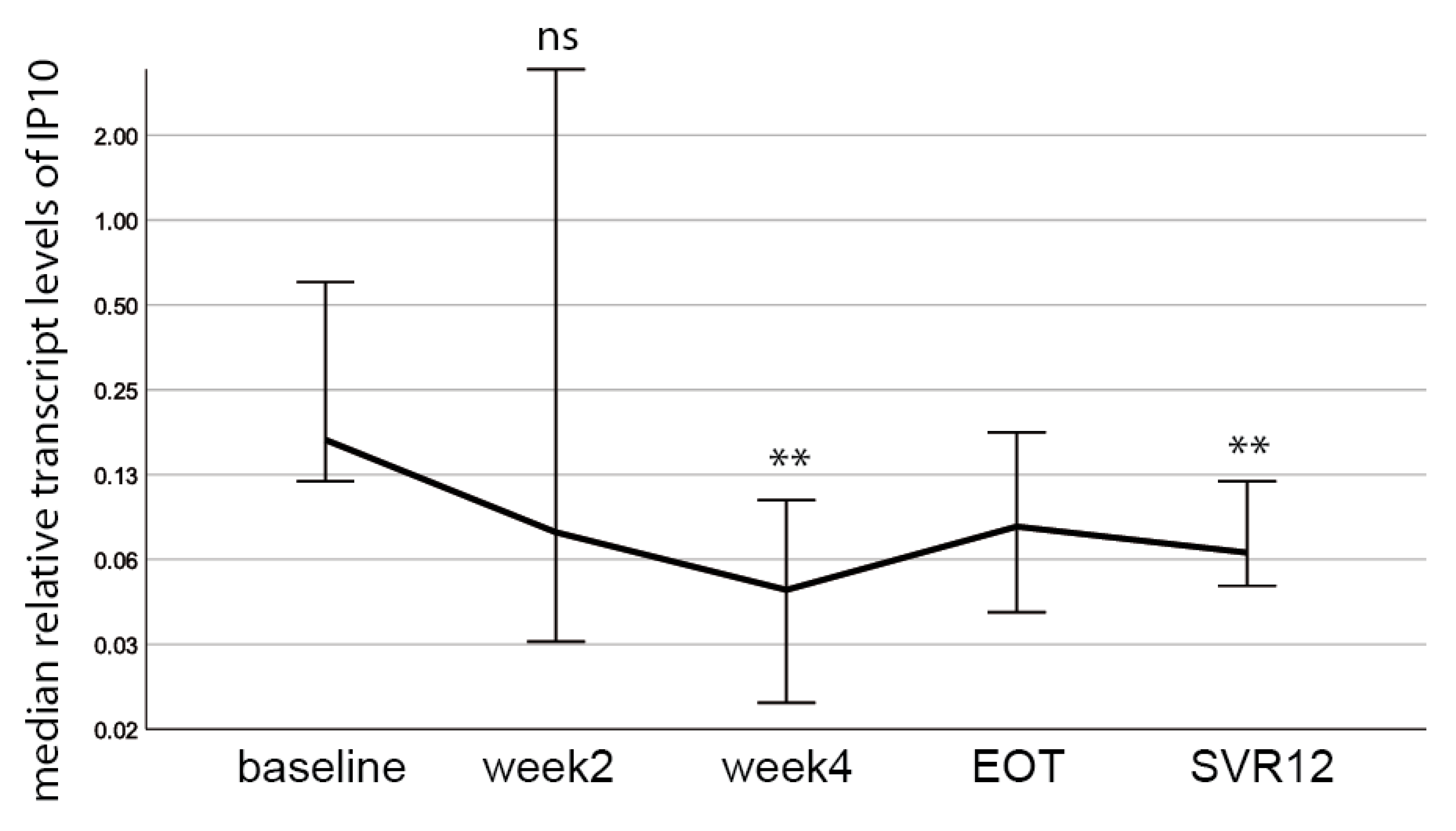
3.5. The Inflammatory Marker Interferon γ-Inducible Protein 10 (IP 10) Decreases during DAA Treatment and Does Not Correlate with HERV-K(HML-2) Transcript Levels
3.6. HERV-K(HML-2) Transcript Levels Are Increased in Patients without Sustained Viral Response
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Suwanthawornkul, T.; Anothaisintawee, T.; Sobhonslidsuk, A.; Thakkinstian, A.; Teerawattananon, Y. Efficacy of Second Generation Direct-Acting Antiviral Agents for Treatment Naïve Hepatitis C Genotype 1: A Systematic Review and Network Meta-Analysis. PLoS ONE 2015, 10, e0145953. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Ji, F.; Yeo, Y.H.; Ogawa, E.; Stave, C.D.; Dang, S.; Li, Z.; Furusyo, N.; Cheung, R.C.; Nguyen, M.H. Systematic review and meta-analysis: Real-world effectiveness of direct-acting antiviral therapies in chronic hepatitis C genotype 3 in Asia. BMJ Open Gastroenterol. 2018, 5, e000209. [Google Scholar] [CrossRef]
- Ramos-Casals, M.; Stone, J.H.; Cid, M.C.; Bosch, X. The cryoglobulinaemias. Lancet 2012, 379, 348–360. [Google Scholar] [CrossRef]
- Mager, D.L.; Medstrand, P. Retroviral repeat sequences. In Nature Encyclopedia of the Human Genome; Cooper, D., Ed.; Nature Publishing Group: London, UK, 2003. [Google Scholar]
- Grange, J.M.; Krone, B.; Kölmel, K. Human endogenous retroviruses in health and disease. J. R. Soc. Med. 2005, 98, 134. [Google Scholar] [CrossRef] [PubMed]
- Belshaw, R.; Katzourakis, A.; Paces, J.; Burt, A.; Tristem, M. High copy number in human endogenous retrovirus families is associated with copying mechanisms in addition to reinfection. Mol. Biol. Evol. 2005, 22, 814–817. [Google Scholar] [CrossRef] [PubMed]
- Belshaw, R.; Pereira, V.; Katzourakis, A.; Talbot, G.; Paces, J.; Burt, A.; Tristem, M. Long-term reinfection of the human genome by endogenous retroviruses. Proc. Natl. Acad. Sci. USA 2004, 101, 4894–4899. [Google Scholar] [CrossRef]
- Friedli, M.; Trono, D. The developmental control of transposable elements and the evolution of higher species. Annu. Rev. Cell Dev. Biol. 2015, 31, 429–451. [Google Scholar] [CrossRef] [PubMed]
- Vincendeau, M.; Gottesdorfer, I.; Schreml, J.M.; Wetie, A.G.; Mayer, J.; Greenwood, A.D.; Helfer, M.; Kramer, S.; Seifarth, W.; Hadian, K.; et al. Modulation of human endogenous retrovirus (HERV) transcription during persistent and de novo HIV-1 infection. Retrovirology 2015, 12, 27. [Google Scholar] [CrossRef]
- Carnell, A.N.; Goodman, J.I. The long (LINEs) and the short (SINEs) of it: Altered methylation as a precursor to toxicity. Toxicol. Sci. 2003, 75, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.; Meese, E. Human endogenous retroviruses in the primate lineage and their influence on host genomes. Cytogenet. Genome Res. 2005, 110, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Buzdin, A.; Kovalskaya-Alexandrova, E.; Gogvadze, E.; Sverdlov, E. At least 50% of human-specific HERV-K (HML-2) long terminal repeats serve in vivo as active promoters for host nonrepetitive DNA transcription. J. Virol. 2006, 80, 10752–10762. [Google Scholar] [CrossRef] [PubMed]
- Bannert, N.; Kurth, R. Retroelements and the human genome: New perspectives on an old relation. Proc. Natl. Acad. Sci. USA 2004, 101 (Suppl. S2), 14572–14579. [Google Scholar] [CrossRef]
- Mangeney, M.; de Parseval, N.; Thomas, G.; Heidmann, T. The full-length envelope of an HERV-H human endogenous retrovirus has immunosuppressive properties. J. General Virol. 2001, 82 Pt 10, 2515–2518. [Google Scholar] [CrossRef]
- Grandi, N.; Tramontano, E. HERV Envelope Proteins: Physiological Role and Pathogenic Potential in Cancer and Autoimmunity. Front. Microbiol. 2018, 9, 462. [Google Scholar] [CrossRef] [PubMed]
- Denne, M.; Sauter, M.; Armbruester, V.; Licht, J.D.; Roemer, K.; Mueller-Lantzsch, N. Physical and functional interactions of human endogenous retrovirus proteins Np9 and rec with the promyelocytic leukemia zinc finger protein. J. Virol. 2007, 81, 5607–5616. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Montojo, M.; Doucet-O’Hare, T.; Henderson, L.; Nath, A. Human endogenous retrovirus-K (HML-2): A comprehensive review. Crit. Rev. Microbiol. 2018, 44, 715–738. [Google Scholar] [CrossRef]
- Dembny, P.; Newman, A.G.; Singh, M.; Hinz, M.; Szczepek, M.; Kruger, C.; Adalbert, R.; Dzaye, O.; Trimbuch, T.; Wallach, T.; et al. Human endogenous retrovirus HERV-K(HML-2) RNA causes neurodegeneration through Toll-like receptors. JCI Insight 2020, 5, e131093. [Google Scholar] [CrossRef]
- Li, W.; Lee, M.H.; Henderson, L.; Tyagi, R.; Bachani, M.; Steiner, J.; Campanac, E.; Hoffman, D.A.; von Geldern, G.; Johnson, K.; et al. Human endogenous retrovirus-K contributes to motor neuron disease. Sci. Transl. Med. 2015, 7, 307ra153. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Sechi, L.A.; Kelvin, D.J. Human Endogenous Retrovirus K (HML-2) in Health and Disease. Front. Microbiol. 2020, 11, 1690. [Google Scholar] [CrossRef]
- Steele, A.J.; Al-Chalabi, A.; Ferrante, K.; Cudkowicz, M.E.; Brown, R.H., Jr.; Garson, J.A. Detection of serum reverse transcriptase activity in patients with ALS and unaffected blood relatives. Neurology 2005, 64, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Hong, Z.; Liu, H.; Chen, X.; Ding, L.; Liu, Z.; Zhou, F.; Yuan, Y. Human Endogenous Retroviruses-K (HML-2) Expression Is Correlated with Prognosis and Progress of Hepatocellular Carcinoma. BioMed Res. Int. 2016, 2016, 8201642. [Google Scholar] [CrossRef] [PubMed]
- Tovo, P.A.; Garazzino, S.; Daprà, V.; Alliaudi, C.; Silvestro, E.; Calvi, C.; Montanari, P.; Galliano, I.; Bergallo, M. Chronic HCV Infection Is Associated with Overexpression of Human Endogenous Retroviruses that Persists after Drug-Induced Viral Clearance. Int. J. Mol. Sci. 2020, 21, 3980. [Google Scholar] [CrossRef]
- Maliniemi, P.; Vincendeau, M.; Mayer, J.; Frank, O.; Hahtola, S.; Karenko, L.; Carlsson, E.; Mallet, F.; Seifarth, W.; Leib-Mosch, C.; et al. Expression of human endogenous retrovirus-w including syncytin-1 in cutaneous T-cell lymphoma. PLoS ONE 2013, 8, e76281. [Google Scholar] [CrossRef] [PubMed]
- Hösel, M.; Quasdorff, M.; Ringelhan, M.; Kashkar, H.; Debey-Pascher, S.; Sprinzl, M.F.; Bockmann, J.H.; Arzberger, S.; Webb, D.; von Olshausen, G.; et al. Hepatitis B Virus Activates Signal Transducer and Activator of Transcription 3 Supporting Hepatocyte Survival and Virus Replication. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 339–363. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Fontana, S.; Zeerleder, S.; Lämmle, B. Derailed oral anticoagulation with very high INR values and poor response to oral vitamin K--cholestasis as a possible cause. Ther. Umschau. Revue Ther. 1999, 56, 477–480. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, G.; Garcia-Tsao, G.; Pagliaro, L. Natural history and prognostic indicators of survival in cirrhosis: A systematic review of 118 studies. J. Hepatol. 2006, 44, 217–231. [Google Scholar] [CrossRef] [PubMed]
- Heller, A.R.; Fischer, S.; Rössel, T.; Geiger, S.; Siegert, G.; Ragaller, M.; Zimmermann, T.; Koch, T. Impact of n-3 fatty acid supplemented parenteral nutrition on haemostasis patterns after major abdominal surgery. Br. J. Nutr. 2002, 87 (Suppl. S1), S95–S101. [Google Scholar] [CrossRef]
- Burri, S.; Demarmels Biasiutti, F.; Lämmle, B.; Wuillemin, W.A. Comparison of Quick/INR values of whole capillary blood (CoaguChek Plus) and venous citrate plasma in patients with and without oral anticoagulation. Schweiz. Med. Wochenschr. 1998, 128, 1723–1729. [Google Scholar] [PubMed]
- Carvalho, J.R.; Verdelho Machado, M. New Insights About Albumin and Liver Disease. Ann. Hepatol. 2018, 17, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Neesgaard, B.; Ruhwald, M.; Weis, N. Inducible protein-10 as a predictive marker of antiviral hepatitis C treatment: A systematic review. World J. Hepatol. 2017, 9, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Buzdin, A.; Kovalskaya-Alexandrova, E.; Gogvadze, E.; Sverdlov, E. GREM, a technique for genome-wide isolation and quantitative analysis of promoter active repeats. Nucleic Acids Res. 2006, 34, e67. [Google Scholar] [CrossRef] [PubMed]
- Vergara Bermejo, A.; Ragonnaud, E.; Daradoumis, J.; Holst, P. Cancer Associated Endogenous Retroviruses: Ideal Immune Targets for Adenovirus-Based Immunotherapy. Int. J. Mol. Sci. 2020, 21, 4843. [Google Scholar] [CrossRef]
- Küry, P.; Nath, A.; Créange, A.; Dolei, A.; Marche, P.; Gold, J.; Giovannoni, G.; Hartung, H.P.; Perron, H. Human Endogenous Retroviruses in Neurological Diseases. Trends Mol. Med. 2018, 24, 379–394. [Google Scholar] [CrossRef]
- Armbruester, V.; Sauter, M.; Roemer, K.; Best, B.; Hahn, S.; Nty, A.; Schmid, A.; Philipp, S.; Mueller, A.; Mueller-Lantzsch, N. Np9 protein of human endogenous retrovirus K interacts with ligand of numb protein X. J. Virol. 2004, 78, 10310–10319. [Google Scholar] [CrossRef][Green Version]
- Medstrand, P.; Mager, D.L. Human-specific integrations of the HERV-K endogenous retrovirus family. J. Virol. 1998, 72, 9782–9787. [Google Scholar] [CrossRef] [PubMed]
- Brattas, P.L.; Jonsson, M.E.; Fasching, L.; Nelander Wahlestedt, J.; Shahsavani, M.; Falk, R.; Falk, A.; Jern, P.; Parmar, M.; Jakobsson, J. TRIM28 Controls a Gene Regulatory Network Based on Endogenous Retroviruses in Human Neural Progenitor Cells. Cell Rep. 2017, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Salavatiha, Z.; Soleimani-Jelodar, R.; Jalilvand, S. The role of endogenous retroviruses-K in human cancer. Rev. Med. Virol. 2020, 30, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bonacci, M.; Lens, S.; Mariño, Z.; Londoño, M.C.; Rodriguez-Tajes, S.; Sánchez-Tapias, J.M.; Ramos-Casals, M.; Hernández-Rodríguez, J.; Forns, X. Long-Term Outcomes of Patients With HCV-Associated Cryoglobulinemic Vasculitis After Virologic Cure. Gastroenterology 2018, 155, 311–315.e6. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N.; Beste, L.A.; Green, P.K.; Singal, A.G.; Tapper, E.B.; Waljee, A.K.; Sterling, R.K.; Feld, J.J.; Kaplan, D.E.; Taddei, T.H.; et al. Increased Risk for Hepatocellular Carcinoma Persists Up to 10 Years After HCV Eradication in Patients With Baseline Cirrhosis or High FIB-4 Scores. Gastroenterology 2019, 157, 1264–1278.e4. [Google Scholar] [CrossRef] [PubMed]
- Terziroli Beretta-Piccoli, B.; Di Bartolomeo, C.; Deleonardi, G.; Grondona, A.G.; Silvestri, T.; Tesei, C.; Melidona, L.; Cerny, A.; Mertens, J.; Semmo, N.; et al. Autoimmune liver serology before and after successful treatment of chronic hepatitis C by direct acting antiviral agents. J. Autoimmun. 2019, 102, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Scarsi, K.K.; Havens, J.P.; Podany, A.T.; Avedissian, S.N.; Fletcher, C.V. HIV-1 Integrase Inhibitors: A Comparative Review of Efficacy and Safety. Drugs 2020, 80, 1649–1676. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, R.; Li, W.; Parades, D.; Bianchet, M.A.; Nath, A. Inhibition of human endogenous retrovirus-K by antiretroviral drugs. Retrovirology 2017, 14, 21. [Google Scholar] [CrossRef] [PubMed]
- Nath, A. HERV-K Suppression Using Antiretroviral Therapy in Volunteers with Amyotrophic Lateral Sclerosis (ALS). National Institute of Neurological Disorders and Stroke, ClinicalTrials.gov identifier: NCT02437110. 2015. Available online: https://clinicaltrials.gov/ct2/show/NCT02437110 (accessed on 27 January 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug Combinations | Number of Patients |
|---|---|
| Simeprevir/Sofosbuvir/Ribavirin | 5 |
| Simeprevir/Sofosbuvir | 6 |
| Daclatasvir/Sofosbuvir | 16 |
| Daclatasvir/Sofosbuvir/Ribavirin | 3 |
| Ledipasvir/Sofosbuvir | 5 |
| Ledipasvir/Sofosbuvir/Ribavirin | 2 |
| Viekirax/Exviera | 4 |
| Viekirax/Ribavirin | 1 |
| Mean Albumin Levels | |||
|---|---|---|---|
| Time Point | HERV-K(HML-2) < 1 | HERV-K(HML-2) > 1 | p-Value |
| baseline | 38.04 g/L | 32.10 g/L | p = 0.005 |
| week 2 | 37.52 g/L | 31.89 g/L | p = 0.003 |
| week 4 | 38.45 g/L | 32.56 g/L | p = 0.001 |
| end of treatment (EOT) | 38.35 g/L | 34.10 g/L | p = 0.004 |
| SVR 12 | 38.68 g/L | 35.30 g/L | p = 0.048 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weber, M.; Padmanabhan Nair, V.; Bauer, T.; Sprinzl, M.F.; Protzer, U.; Vincendeau, M. Increased HERV-K(HML-2) Transcript Levels Correlate with Clinical Parameters of Liver Damage in Hepatitis C Patients. Cells 2021, 10, 774. https://doi.org/10.3390/cells10040774
Weber M, Padmanabhan Nair V, Bauer T, Sprinzl MF, Protzer U, Vincendeau M. Increased HERV-K(HML-2) Transcript Levels Correlate with Clinical Parameters of Liver Damage in Hepatitis C Patients. Cells. 2021; 10(4):774. https://doi.org/10.3390/cells10040774
Chicago/Turabian StyleWeber, Melanie, Vidya Padmanabhan Nair, Tanja Bauer, Martin F. Sprinzl, Ulrike Protzer, and Michelle Vincendeau. 2021. "Increased HERV-K(HML-2) Transcript Levels Correlate with Clinical Parameters of Liver Damage in Hepatitis C Patients" Cells 10, no. 4: 774. https://doi.org/10.3390/cells10040774
APA StyleWeber, M., Padmanabhan Nair, V., Bauer, T., Sprinzl, M. F., Protzer, U., & Vincendeau, M. (2021). Increased HERV-K(HML-2) Transcript Levels Correlate with Clinical Parameters of Liver Damage in Hepatitis C Patients. Cells, 10(4), 774. https://doi.org/10.3390/cells10040774