
ACPA Status Correlates with Differential Immune Profile in Patients with Rheumatoid Arthritis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Synovial Tissue Single Cell Suspensions
2.2. Flow Cytometric Analysis
2.3. RNAseq
2.4. Pathway Enrichment Analysis
2.5. Hierarchical Clustering and PCA
2.6. Statistical Analysis
2.7. Study Approval
3. Results
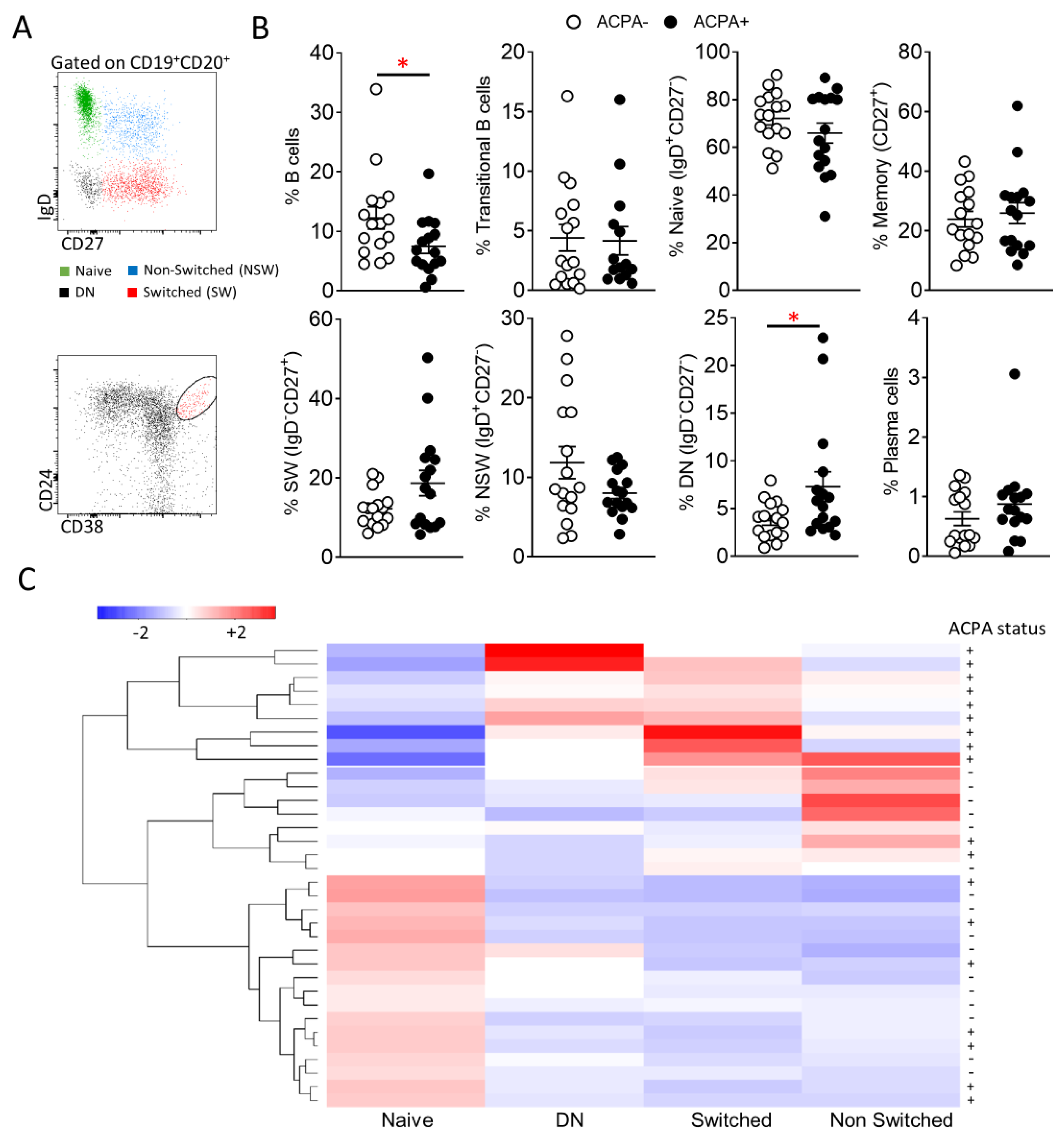
3.1. Peripheral Blood B Cell Subpopulation Profile of ACPA− and ACPA+RA Patients
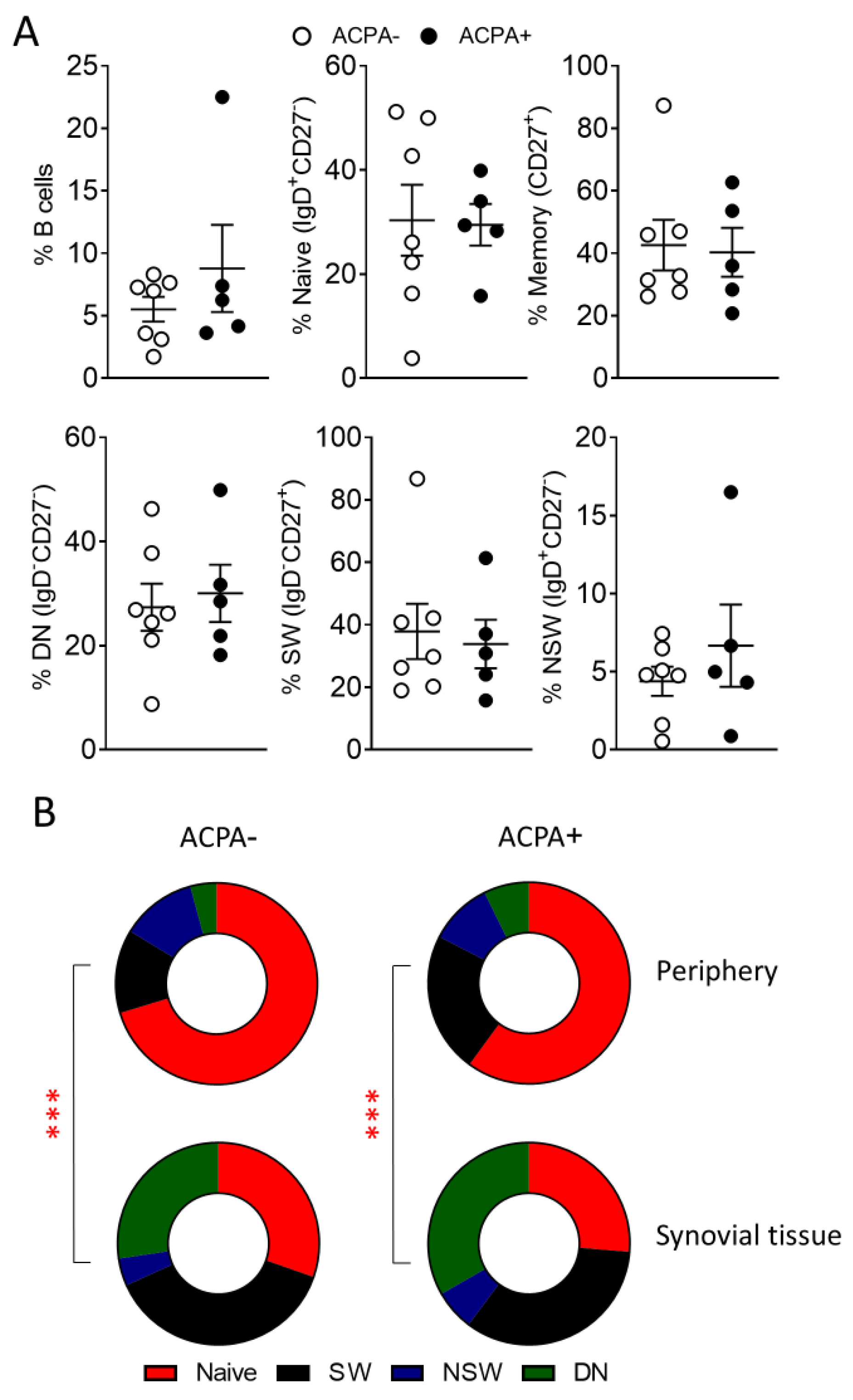
3.2. Predominance of Switched and DN Memory B Cells in the Synovial Tissue of ACPA− and ACPA+ RA Patients
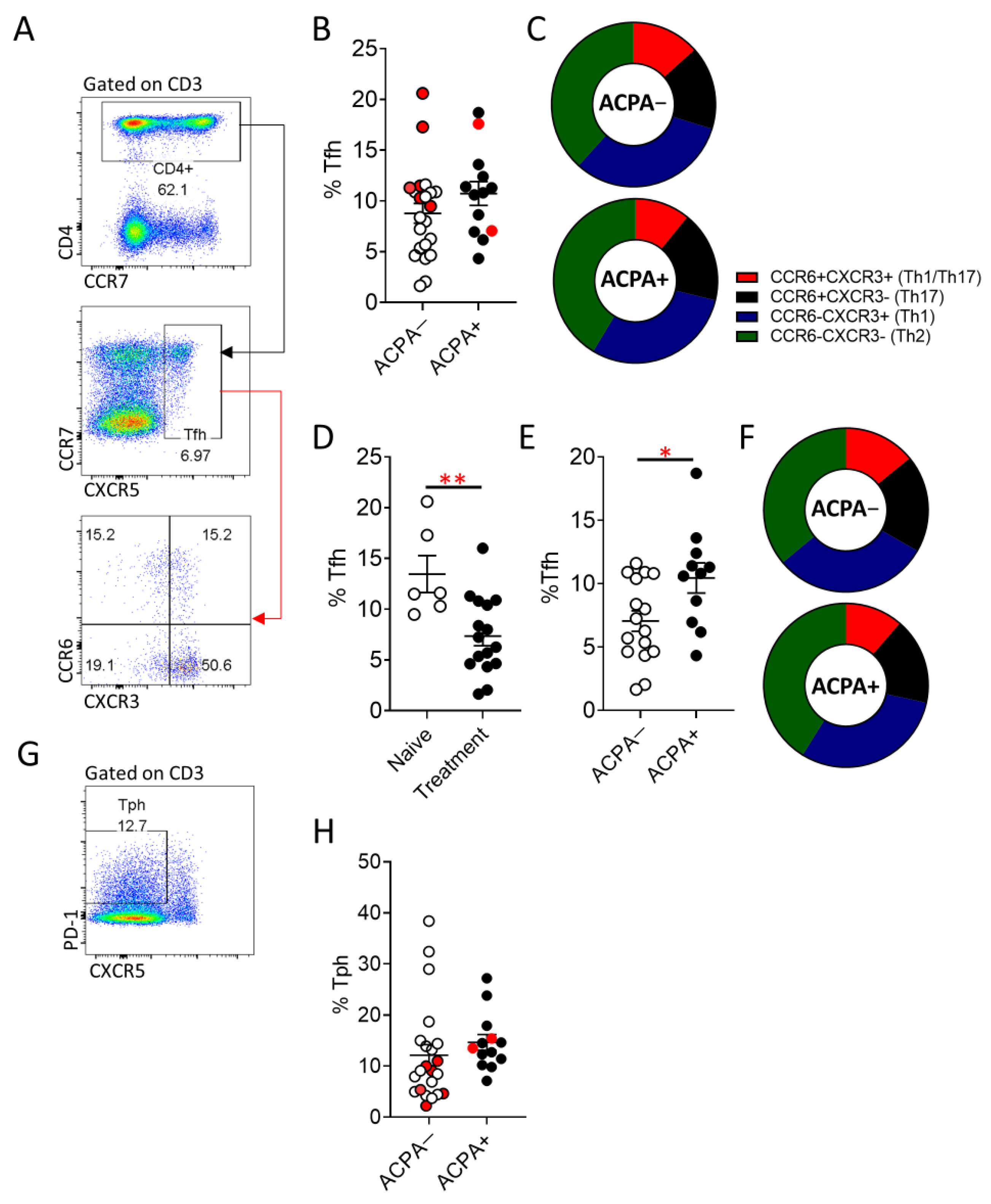
3.3. Increased Peripheral Blood T Follicular Helper (Tfh) Cell Frequency in ACPA+ Compared to ACPA− RA Patients
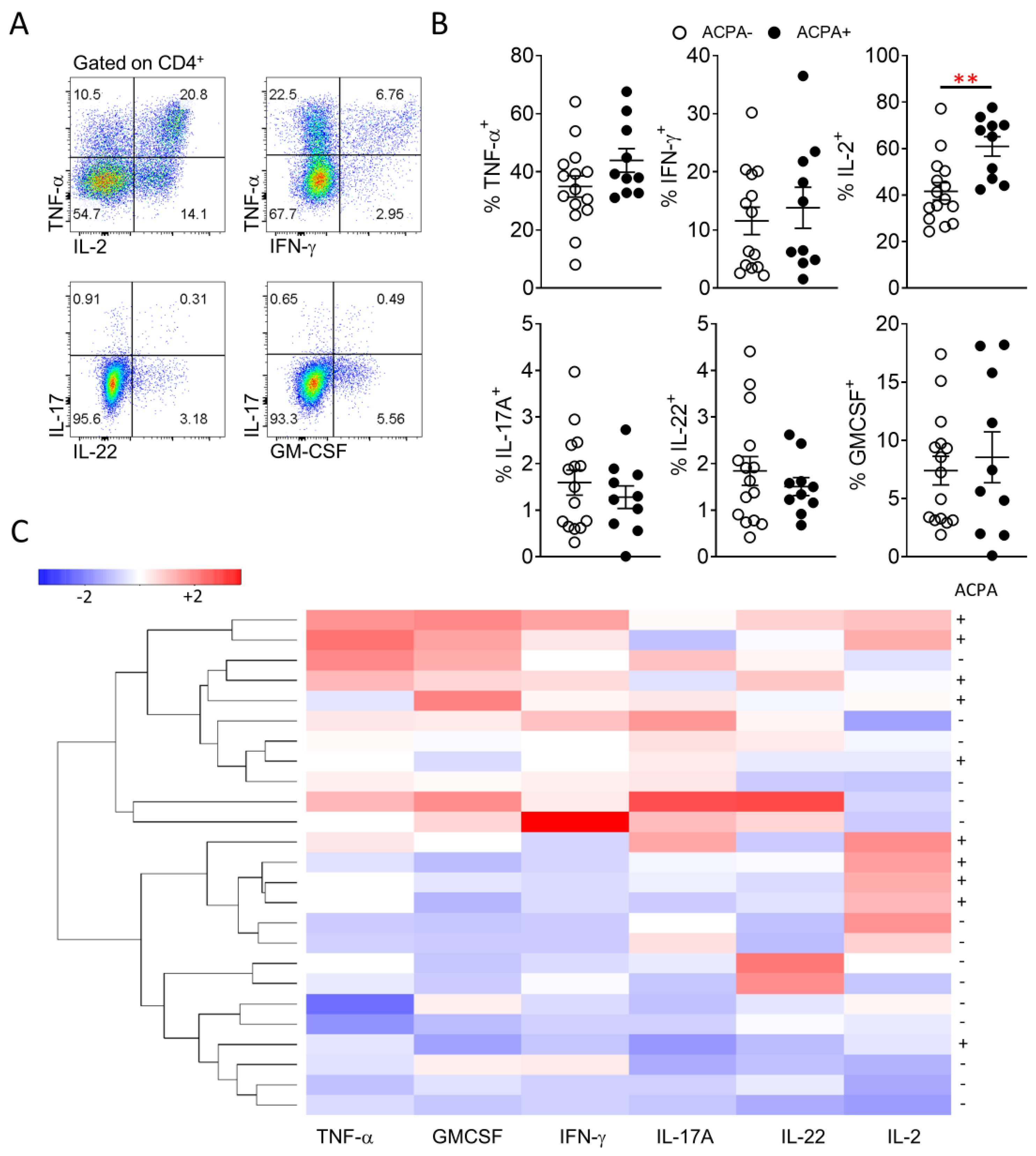
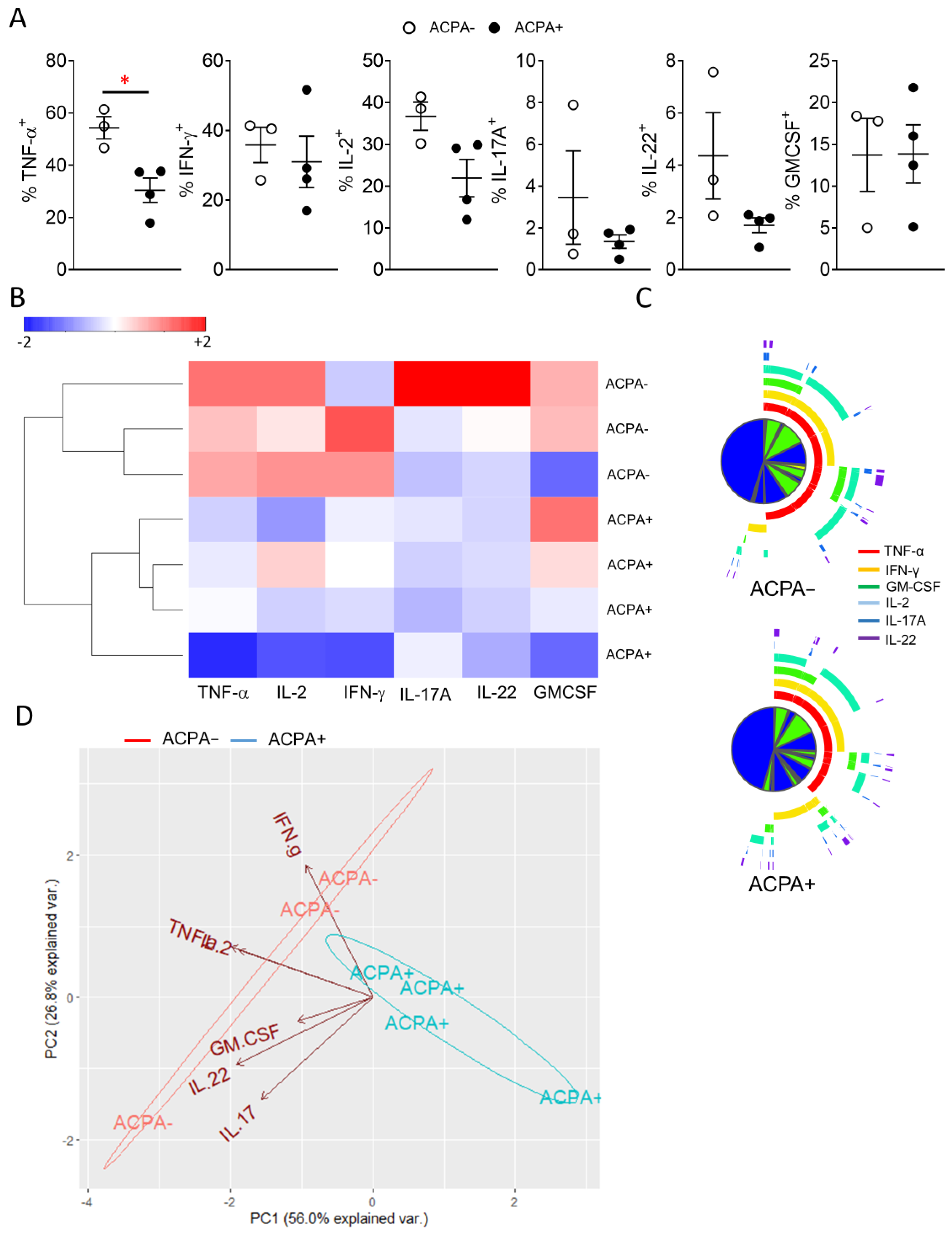
3.4. Synovial CD4 T Cell Proinflammatory Cytokine Differences Lead to Distinct Grouping of ACPA− and ACPA+ RA Patients
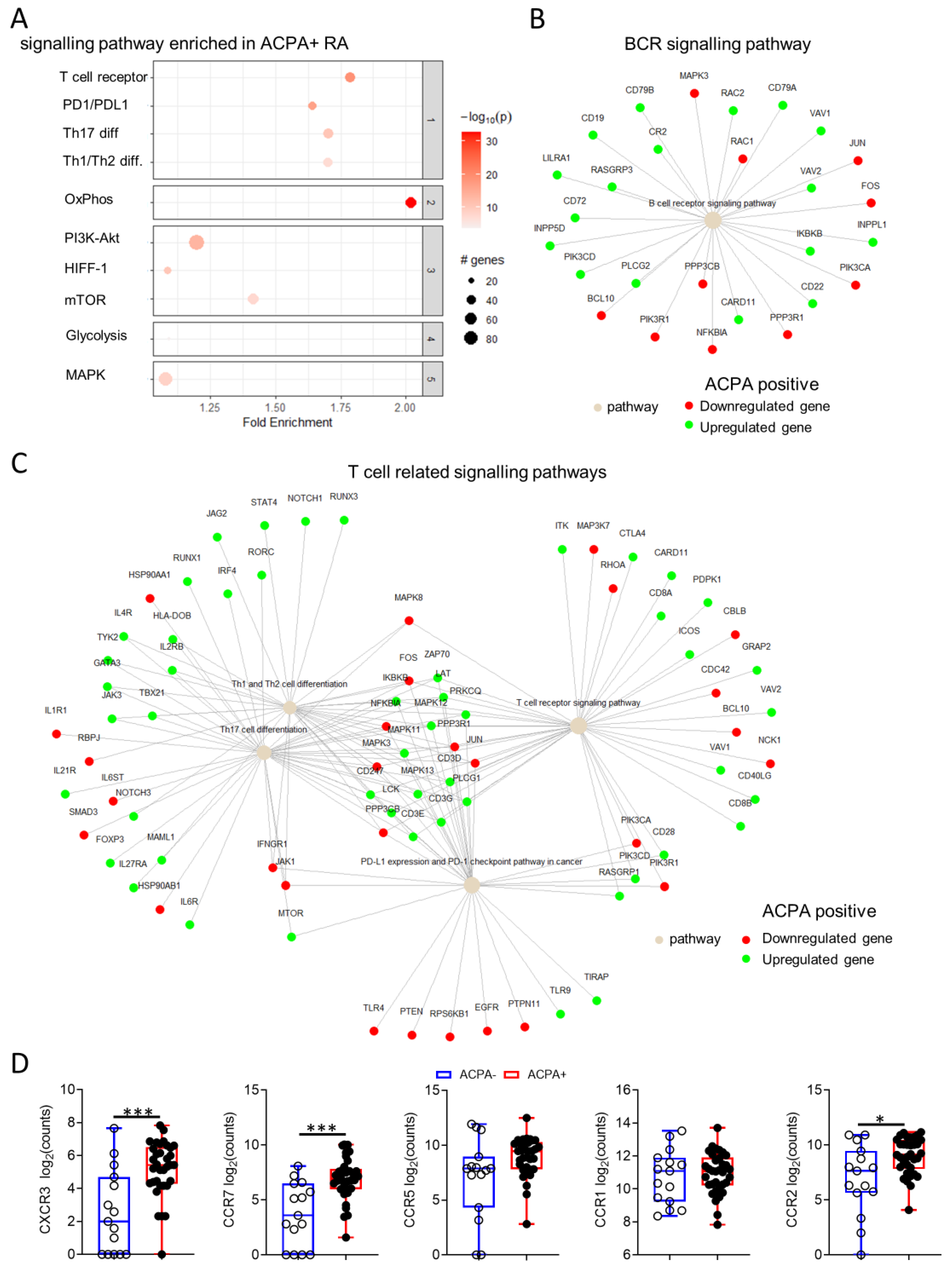
3.5. Distinct Synovial Tissue Gene Signalling Pathways Are Implicated in ACPA+ Compared to ACPA− RA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Veale, D.J.; Orr, C.; Fearon, U. Cellular and molecular perspectives in rheumatoid arthritis. Semin. Immunopathol. 2017, 39, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Avouac, J.; Gossec, L.; Dougados, M. Diagnostic and predictive value of anti-cyclic citrullinated protein antibodies in rheumatoid arthritis: A systematic literature review. Ann. Rheum. Dis. 2006, 65, 845–851. [Google Scholar] [CrossRef]
- van Zanten, A.; Arends, S.; Roozendaal, C.; Limburg, P.C.; Maas, F.; Trouw, L.A.; Toes, R.E.M.; Huizinga, T.W.J.; Bootsma, H.; Brouwer, E. Presence of anticitrullinated protein antibodies in a large population-based cohort from the Netherlands. Ann. Rheum. Dis. 2017, 76, 1184–1190. [Google Scholar] [CrossRef]
- Padyukov, L.; Seielstad, M.; Ong, R.T.; Ding, B.; Ronnelid, J.; Seddighzadeh, M.; Alfredsson, L.; Klareskog, L. Epidemiological Investigation of Rheumatoid Arthritis study, g., A genome-wide association study suggests contrasting associations in ACPA-positive versus ACPA-negative rheumatoid arthritis. Ann. Rheum. Dis. 2011, 70, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Grosse, J.; Allado, E.; Roux, C.; Pierreisnard, A.; Couderc, M.; Clerc-Urmes, I.; Remen, T.; Albuisson, E.; De Carvalho-Bittencourt, M.; Chary-Valckenaere, I.; et al. ACPA-positive versus ACPA-negative rheumatoid arthritis: Two distinct erosive disease entities on radiography and ultrasonography. Rheumatol. Int. 2020, 40, 615–624. [Google Scholar] [CrossRef]
- Mouterde, G.; Rincheval, N.; Lukas, C.; Daien, C.; Saraux, A.; Dieude, P.; Morel, J.; Combe, B. Outcome of patients with early arthritis without rheumatoid factor and ACPA and predictors of rheumatoid arthritis in the ESPOIR cohort. Arthritis Res. Ther. 2019, 21, 140. [Google Scholar] [CrossRef] [PubMed]
- Burgers, L.E.; van Steenbergen, H.W.; Ten Brinck, R.M.; Huizinga, T.W.; van der Helm-van Mil, A.H. Differences in the symptomatic phase preceding ACPA-positive and ACPA-negative RA: A longitudinal study in arthralgia during progression to clinical arthritis. Ann. Rheum. Dis. 2017, 76, 1751–1754. [Google Scholar] [CrossRef] [PubMed]
- Seegobin, S.D.; Ma, M.H.; Dahanayake, C.; Cope, A.P.; Scott, D.L.; Lewis, C.M.; Scott, I.C. ACPA-positive and ACPA-negative rheumatoid arthritis differ in their requirements for combination DMARDs and corticosteroids: Secondary analysis of a randomized controlled trial. Arthritis Res. Ther. 2014, 16, R13. [Google Scholar] [CrossRef]
- Paulissen, S.M.; van Hamburg, J.P.; Davelaar, N.; Vroman, H.; Hazes, J.M.; de Jong, P.H.; Lubberts, E. CCR6+ Th cell populations distinguish ACPA positive from ACPA negative rheumatoid arthritis. Arthritis Res. Ther. 2015, 17, 344. [Google Scholar] [CrossRef]
- Alvandpur, N.; Tabatabaei, R.; Tahamoli-Roudsari, A.; Basiri, Z.; Behzad, M.; Rezaeepoor, M.; Roshanaei, G.; Hajilooi, M.; Solgi, G. Circulating IFN-gamma producing CD4+ T cells and IL-17A producing CD4+ T cells, HLA-shared epitope and ACPA may characterize the clinical response to therapy in rheumatoid arthritis patients. Hum. Immunol. 2020, 81, 228–236. [Google Scholar] [CrossRef]
- Orr, C.; Najm, A.; Biniecka, M.; McGarry, T.; Ng, C.T.; Young, F.; Fearon, U.; Veale, D.J. Synovial Immunophenotype and Anti-Citrullinated Peptide Antibodies in Rheumatoid Arthritis Patients: Relationship to Treatment Response and Radiologic Prognosis. Arthritis Rheumatol. 2017, 69, 2114–2123. [Google Scholar] [CrossRef]
- Yancey, C.L.; Zmijewski, C.; Athreya, B.H.; Doughty, R.A. Arthropathy of Down’s syndrome. Arthritis Rheum. 1984, 27, 929–934. [Google Scholar] [CrossRef]
- van Oosterhout, M.; Bajema, I.; Levarht, E.W.; Toes, R.E.; Huizinga, T.W.; van Laar, J.M. Differences in synovial tissue infiltrates between anti-cyclic citrullinated peptide-positive rheumatoid arthritis and anti-cyclic citrullinated peptide-negative rheumatoid arthritis. Arthritis Rheum. 2008, 58, 53–60. [Google Scholar] [CrossRef]
- Gomez-Puerta, J.A.; Celis, R.; Hernandez, M.V.; Ruiz-Esquide, V.; Ramirez, J.; Haro, I.; Canete, J.D.; Sanmarti, R. Differences in synovial fluid cytokine levels but not in synovial tissue cell infiltrate between anti-citrullinated peptide/protein antibody-positive and -negative rheumatoid arthritis patients. Arthritis Res. Ther. 2013, 15, R182. [Google Scholar] [CrossRef] [PubMed]
- Aldridge, J.; Ekwall, A.H.; Mark, L.; Bergstrom, B.; Andersson, K.; Gjertsson, I.; Lundell, A.C.; Rudin, A. T helper cells in synovial fluid of patients with rheumatoid arthritis primarily have a Th1 and a CXCR3+Th2 phenotype. Arthritis Res. Ther. 2020, 22, 245. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.T.; Biniecka, M.; Kennedy, A.; McCormick, J.; Fitzgerald, O.; Bresnihan, B.; Buggy, D.; Taylor, C.T.; O’Sullivan, J.; Fearon, U.; et al. Synovial tissue hypoxia and inflammation in vivo. Ann. Rheum. Dis. 2010, 69, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Floudas, A.N.N.; Marzaioli, V.; Murray, K.; Moran, B.; Monaghan, M.; Low, C.; Mullan, R.H.; Rao, N.L.; Krishna, V.; Nagpal, S.; et al. Pathogenic, glycolytic PD-1+ B cells accumulate in the hypoxic RA joint. JCI Insight 2020, 5, e139032. [Google Scholar] [CrossRef] [PubMed]
- Wade, S.M.; Canavan, M.; McGarry, T.; Low, C.; Wade, S.C.; Mullan, R.H.; Veale, D.J.; Fearon, U. Association of synovial tissue polyfunctional T-cells with DAPSA in psoriatic arthritis. Ann. Rheum. Dis. 2019, 78, 350–354. [Google Scholar] [CrossRef]
- Guo, Y.; Walsh, A.M.; Fearon, U.; Smith, M.D.; Wechalekar, M.D.; Yin, X.; Cole, S.; Orr, C.; McGarry, T.; Canavan, M.; et al. CD40L-Dependent Pathway Is Active at Various Stages of Rheumatoid Arthritis Disease Progression. J. Immunol. 2017, 198, 4490–4501. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Iwata, Y.; Matsushita, T.; Horikawa, M.; Dilillo, D.J.; Yanaba, K.; Venturi, G.M.; Szabolcs, P.M.; Bernstein, S.H.; Magro, C.M.; Williams, A.D.; et al. Characterization of a rare IL-10-competent B-cell subset in humans that parallels mouse regulatory B10 cells. Blood 2011, 117, 530–541. [Google Scholar] [CrossRef]
- Rao, D.A.; Gurish, M.F.; Marshall, J.L.; Slowikowski, K.; Fonseka, C.Y.; Liu, Y.; Donlin, L.T.; Henderson, L.A.; Wei, K.; Mizoguchi, F.; et al. Pathologically expanded peripheral T helper cell subset drives B cells in rheumatoid arthritis. Nature 2017, 542, 110–114. [Google Scholar] [CrossRef]
- Bautista-Caro, M.B.; Arroyo-Villa, I.; Castillo-Gallego, C.; de Miguel, E.; Peiteado, D.; Plasencia-Rodriguez, C.; Villalba, A.; Sanchez-Mateos, P.; Puig-Kroger, A.; Martin-Mola, E.; et al. Decreased frequencies of circulating follicular helper T cell counterparts and plasmablasts in ankylosing spondylitis patients Naive for TNF blockers. PLoS ONE 2014, 9, e107086. [Google Scholar]
- Velu, V.; Mylvaganam, G.H.; Gangadhara, S.; Hong, J.J.; Iyer, S.S.; Gumber, S.; Ibegbu, C.C.; Villinger, F.; Amara, R.R. Induction of Th1-Biased T Follicular Helper (Tfh) Cells in Lymphoid Tissues during Chronic Simian Immunodeficiency Virus Infection Defines Functionally Distinct Germinal Center Tfh Cells. J. Immunol. 2016, 197, 1832–1842. [Google Scholar] [CrossRef]
- Lingampalli, N.; Sokolove, J.; Lahey, L.J.; Edison, J.D.; Gilliland, W.R.; Holers, V.M.; Deane, K.D.; Robinson, W.H. Combination of anti-citrullinated protein antibodies and rheumatoid factor is associated with increased systemic inflammatory mediators and more rapid progression from preclinical to clinical rheumatoid arthritis. Clin. Immunol. 2018, 195, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Santos-Moreno, P.; Sanchez, G.; Castro, C. Rheumatoid factor as predictor of response to treatment with anti-TNF alpha drugs in patients with rheumatoid arthritis: Results of a cohort study. Medicine 2019, 98, e14181. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Ulgen, E.; Ozisik, O.; Sezerman, O.U. pathfindR: An R Package for Comprehensive Identification of Enriched Pathways in Omics Data Through Active Subnetworks. Front. Genet. 2019, 10, 858. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.R.; McDavid, A.N.; Kongpachith, S.; Lingampalli, N.; Glanville, J.; Ju, C.H.; Gottardo, R.; Robinson, W.H. T Cell-Dependent Affinity Maturation and Innate Immune Pathways Differentially Drive Autoreactive B Cell Responses in Rheumatoid Arthritis. Arthritis Rheumatol. 2018, 70, 1732–1744. [Google Scholar] [CrossRef]
- Ma, J.; Zhu, C.; Ma, B.; Tian, J.; Baidoo, S.E.; Mao, C.; Wu, W.; Chen, J.; Tong, J.; Yang, M.; et al. Increased frequency of circulating follicular helper T cells in patients with rheumatoid arthritis. Clin. Dev. Immunol. 2012, 2012, 827480. [Google Scholar] [CrossRef]
- Wang, X.; Yang, C.; Xu, F.; Qi, L.; Wang, J.; Yang, P. Imbalance of circulating Tfr/Tfh ratio in patients with rheumatoid arthritis. Clin. Exp. Med. 2019, 19, 55–64. [Google Scholar] [CrossRef]
- Basdeo, S.A.; Moran, B.; Cluxton, D.; Canavan, M.; McCormick, J.; Connolly, M.; Orr, C.; Mills, K.H.; Veale, D.J.; Fearon, U.; et al. Polyfunctional, Pathogenic CD161+ Th17 Lineage Cells Are Resistant to Regulatory T Cell-Mediated Suppression in the Context of Autoimmunity. J. Immunol. 2015, 195, 528–540. [Google Scholar] [CrossRef]
- Nielen, M.M.; van Schaardenburg, D.; Reesink, H.W.; van de Stadt, R.J.; van der Horst-Bruinsma, I.E.; de Koning, M.H.; Habibuw, M.R.; Vandenbroucke, J.P.; Dijkmans, B.A. Specific autoantibodies precede the symptoms of rheumatoid arthritis: A study of serial measurements in blood donors. Arthritis Rheum. 2004, 50, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, P.; Eder, R.; Boeld, T.J.; Doser, K.; Piseshka, B.; Andreesen, R.; Edinger, M. Only the CD45RA+ subpopulation of CD4+CD25high T cells gives rise to homogeneous regulatory T-cell lines upon in vitro expansion. Blood 2006, 108, 4260–4267. [Google Scholar] [CrossRef]
- Booth, N.J.; McQuaid, A.J.; Sobande, T.; Kissane, S.; Agius, E.; Jackson, S.E.; Salmon, M.; Falciani, F.; Yong, K.; Rustin, M.H.; et al. Different proliferative potential and migratory characteristics of human CD4+ regulatory T cells that express either CD45RA or CD45RO. J. Immunol. 2010, 184, 4317–4326. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, A.; Joshua, V.; Haj Hensvold, A.; Jin, T.; Sun, M.; Vivar, N.; Ytterberg, A.J.; Engstrom, M.; Fernandes-Cerqueira, C.; Amara, K.; et al. Identification of a novel chemokine-dependent molecular mechanism underlying rheumatoid arthritis-associated autoantibody-mediated bone loss. Ann. Rheum. Dis. 2016, 75, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Moschovakis, G.L.; Bubke, A.; Friedrichsen, M.; Ristenpart, J.; Back, J.W.; Falk, C.S.; Kremmer, E.; Forster, R. The chemokine receptor CCR7 is a promising target for rheumatoid arthritis therapy. Cell. Mol. Immunol. 2019, 16, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Lebre, M.C.; Vergunst, C.E.; Choi, I.Y.; Aarrass, S.; Oliveira, A.S.; Wyant, T.; Horuk, R.; Reedquist, K.A.; Tak, P.P. Why CCR2 and CCR5 blockade failed and why CCR1 blockade might still be effective in the treatment of rheumatoid arthritis. PLoS ONE 2011, 6, e21772. [Google Scholar] [CrossRef] [PubMed]
- Bakheet, S.A.; Ansari, M.A.; Nadeem, A.; Attia, S.M.; Alhoshani, A.R.; Gul, G.; Al-Qahtani, Q.H.; Albekairi, N.A.; Ibrahim, K.E.; Ahmad, S.F. CXCR3 antagonist AMG487 suppresses rheumatoid arthritis pathogenesis and progression by shifting the Th17/Treg cell balance. Cell. Signal. 2019, 64, 109395. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Floudas, A.; Canavan, M.; McGarry, T.; Mullan, R.; Nagpal, S.; Veale, D.J.; Fearon, U. ACPA Status Correlates with Differential Immune Profile in Patients with Rheumatoid Arthritis. Cells 2021, 10, 647. https://doi.org/10.3390/cells10030647
Floudas A, Canavan M, McGarry T, Mullan R, Nagpal S, Veale DJ, Fearon U. ACPA Status Correlates with Differential Immune Profile in Patients with Rheumatoid Arthritis. Cells. 2021; 10(3):647. https://doi.org/10.3390/cells10030647
Chicago/Turabian StyleFloudas, Achilleas, Mary Canavan, Trudy McGarry, Ronan Mullan, Sunil Nagpal, Douglas J. Veale, and Ursula Fearon. 2021. "ACPA Status Correlates with Differential Immune Profile in Patients with Rheumatoid Arthritis" Cells 10, no. 3: 647. https://doi.org/10.3390/cells10030647
APA StyleFloudas, A., Canavan, M., McGarry, T., Mullan, R., Nagpal, S., Veale, D. J., & Fearon, U. (2021). ACPA Status Correlates with Differential Immune Profile in Patients with Rheumatoid Arthritis. Cells, 10(3), 647. https://doi.org/10.3390/cells10030647