
KLF4, Slug and EMT in Head and Neck Squamous Cell Carcinoma

 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Samples for RNA Isolation
2.2. RNA Isolation, Reverse Transcription, PCR and Sequencing
2.3. Immunohistochemistry and Immunofluorescence Staining, Image Acquisition
2.4. Cell Lines, Cell Culture and Treatment Conditions
2.5. Western Blot
3. Results
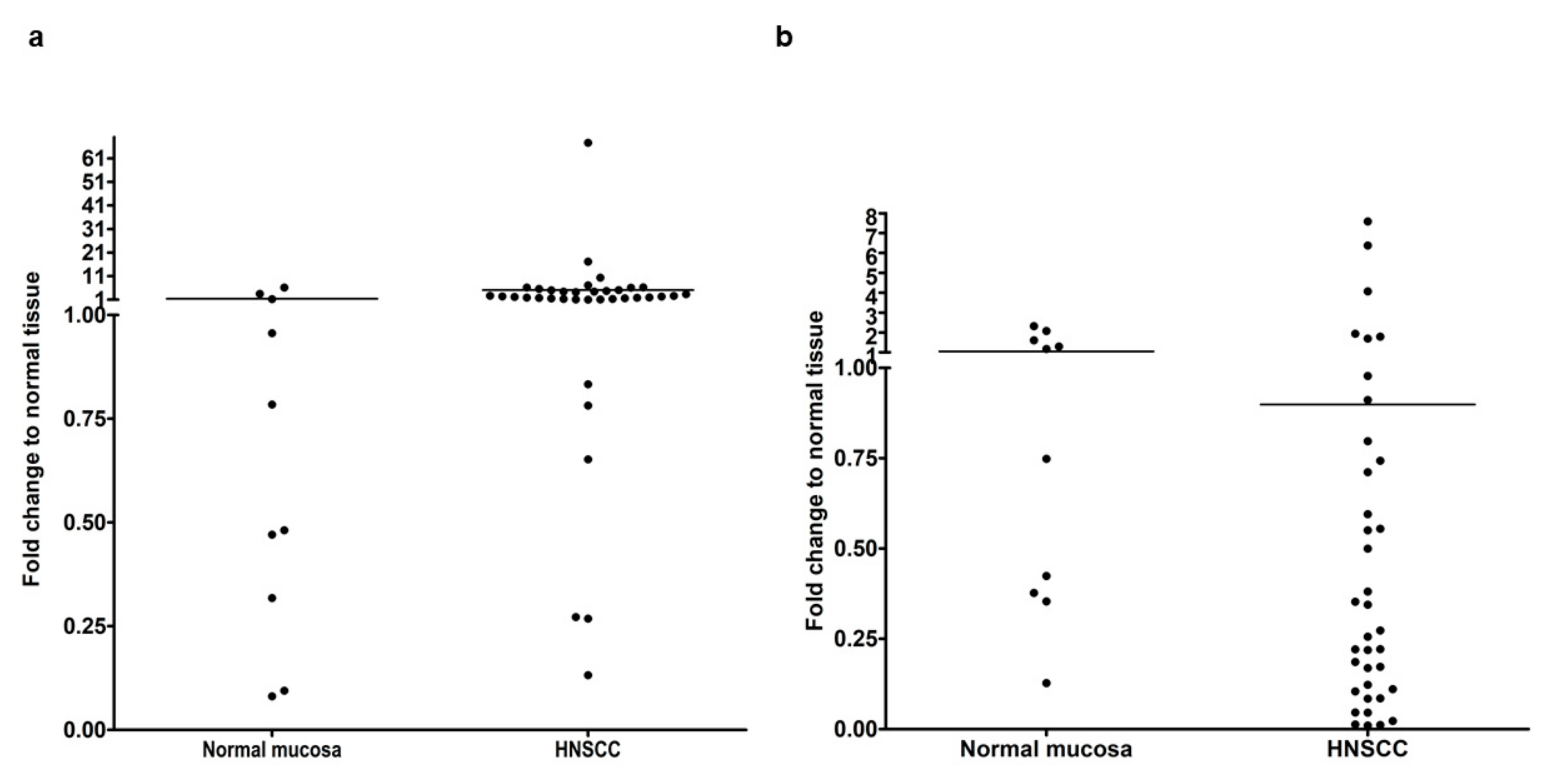
3.1. Gene Expression Analysis of EMT-Related Transcription Factors in HNSCC Compared with Normal Oral Mucosa
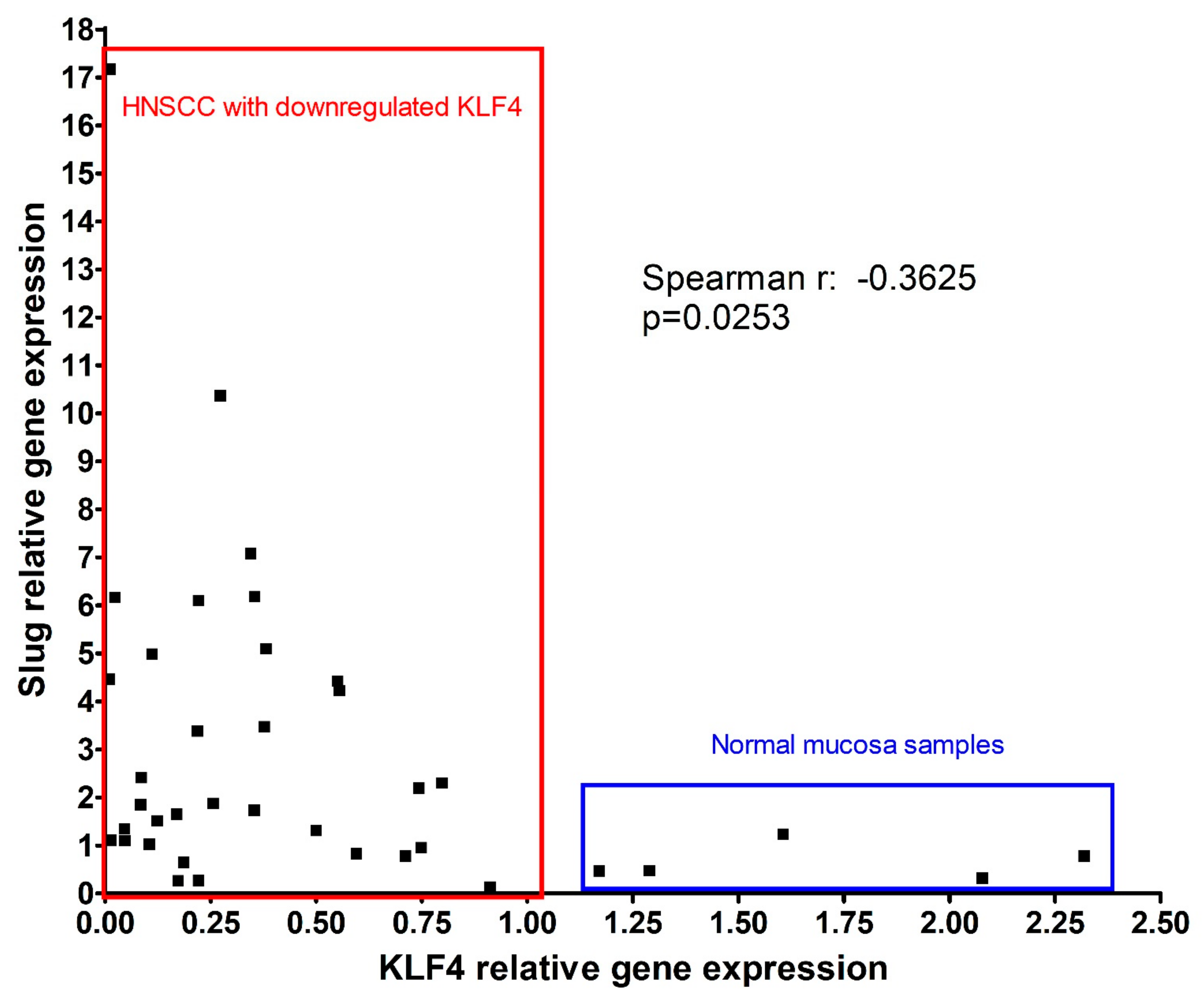
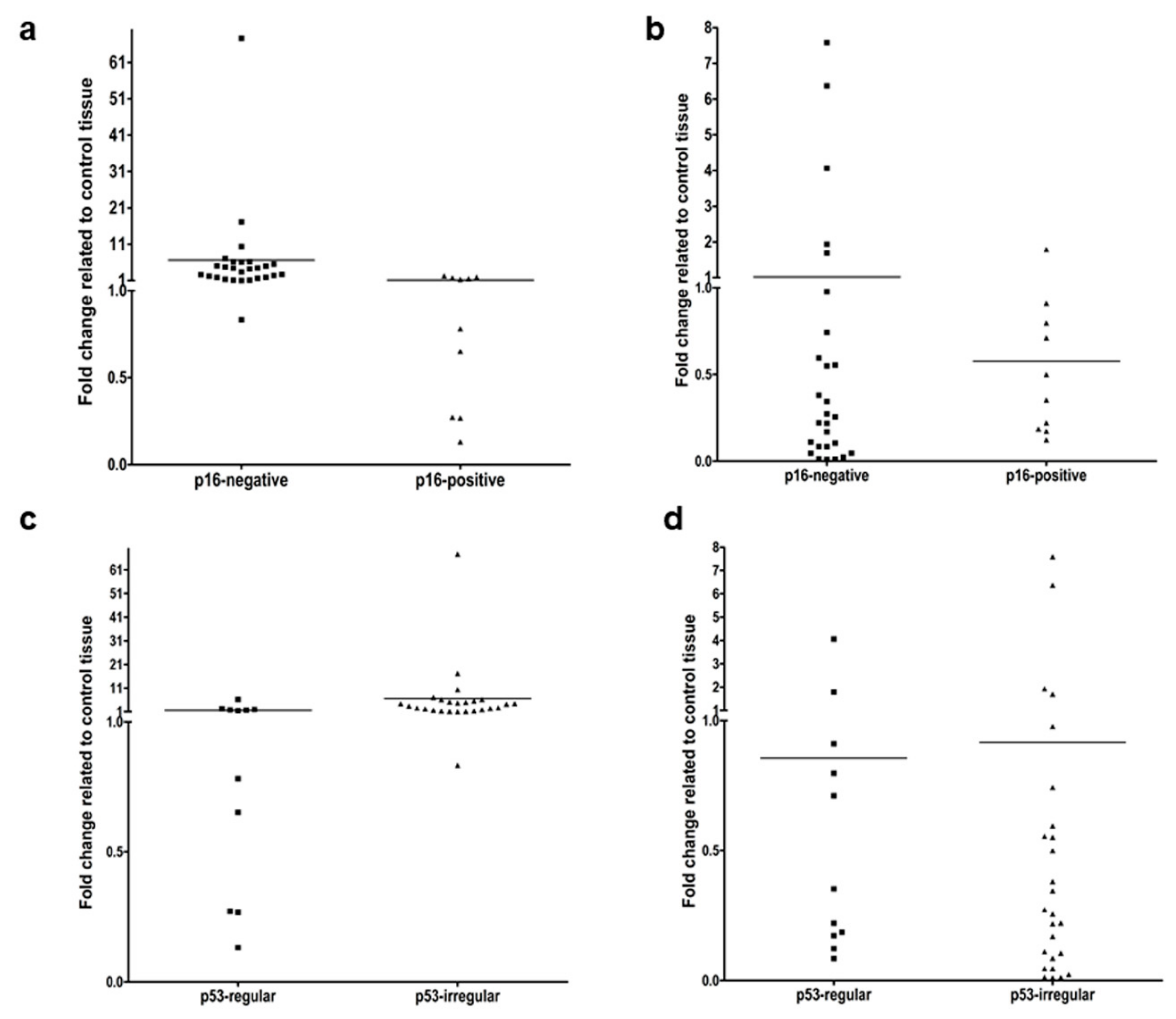
3.2. Gene Expression of Slug and KLF4 in HNSCC in Relation to HPV and p53 Background
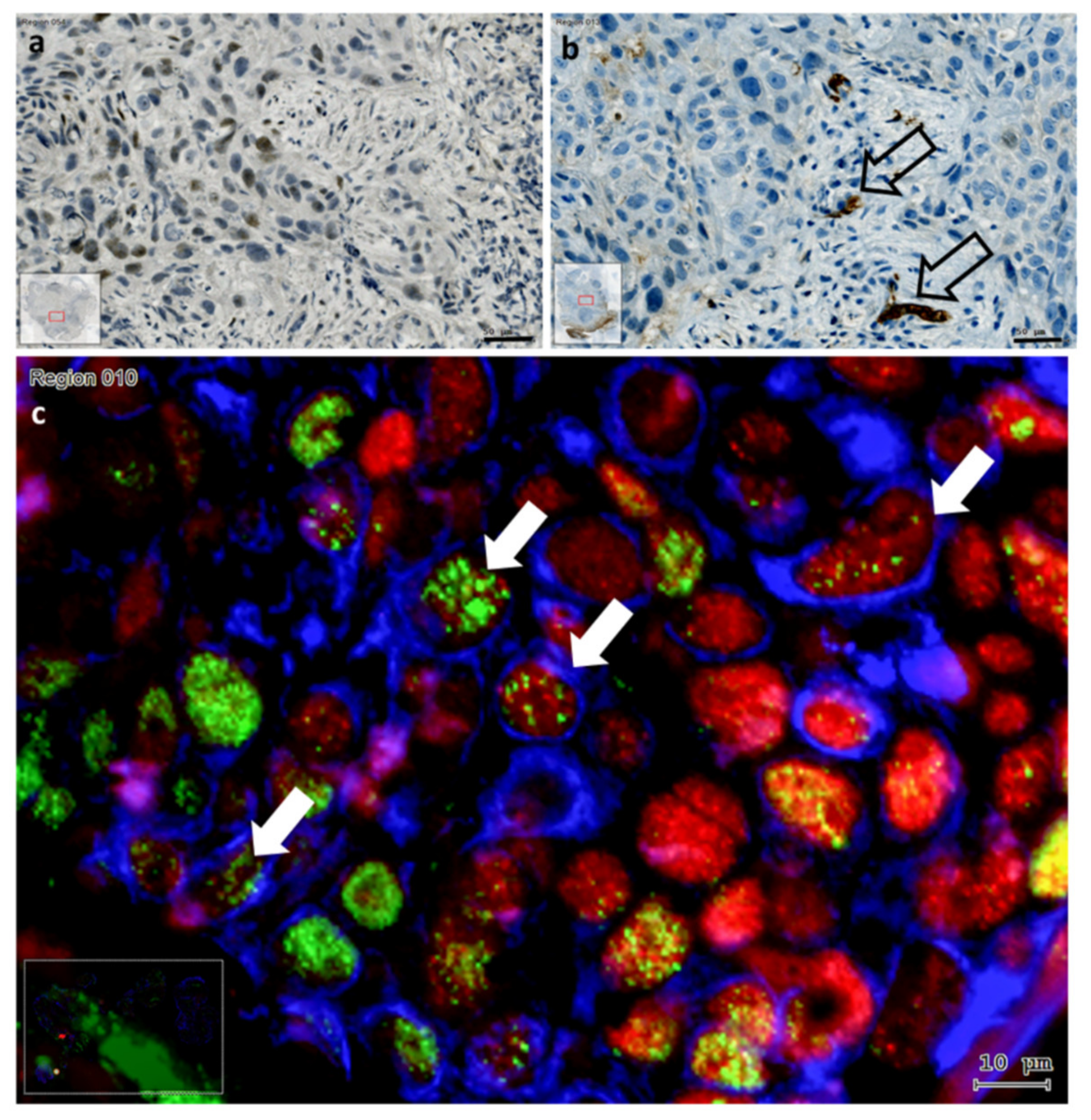
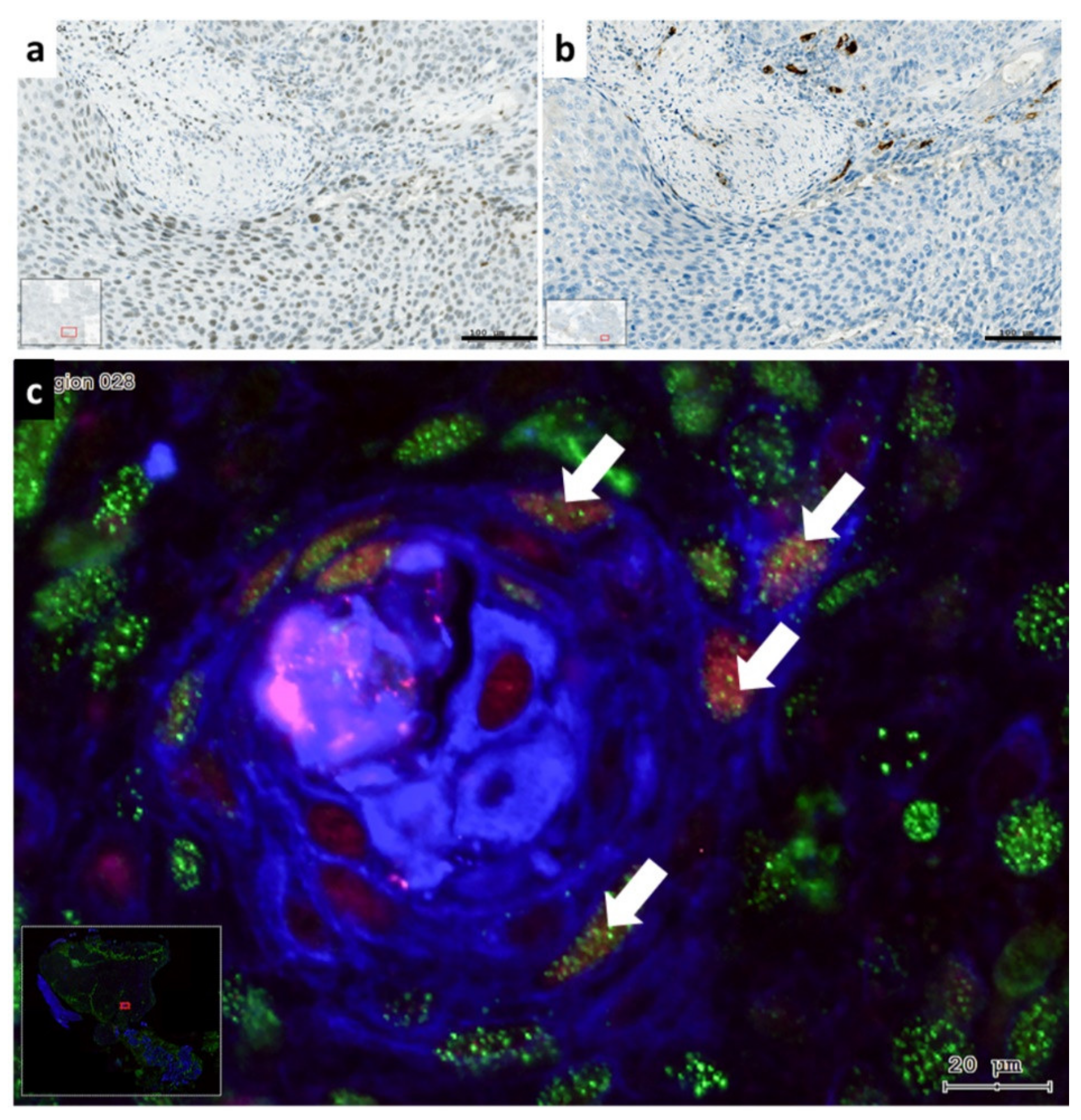
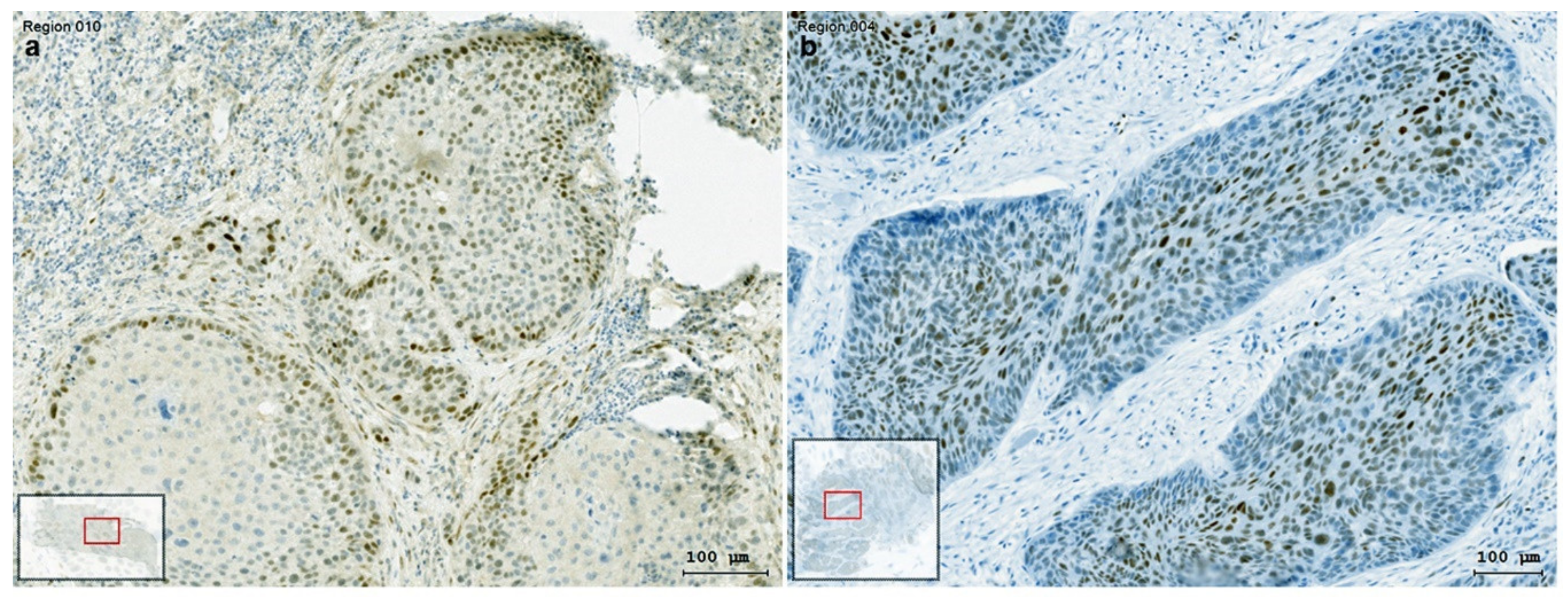
3.3. Distribution of Slug KLF4 and TGF-β1 in HNSCC Tumor Tissue
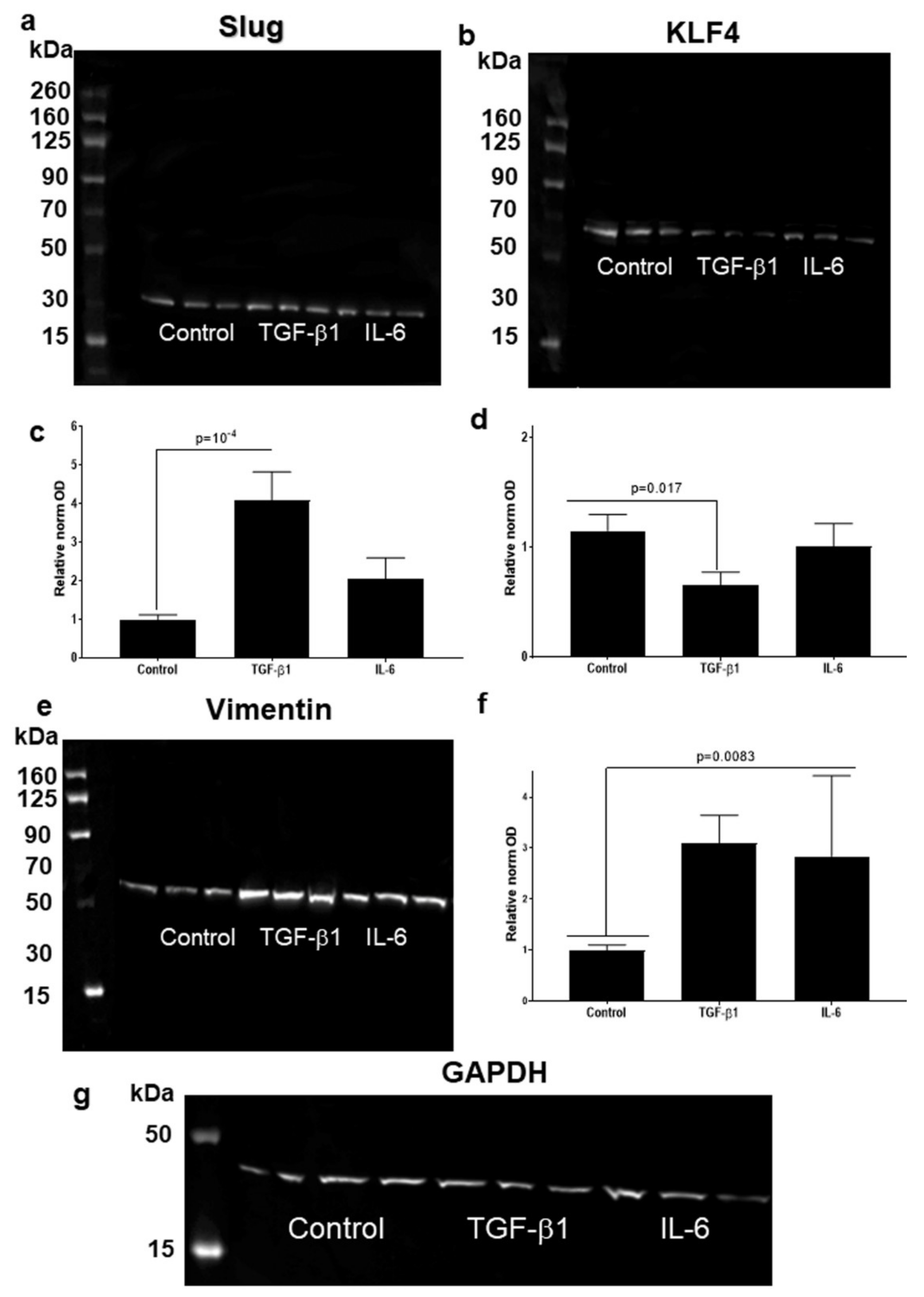
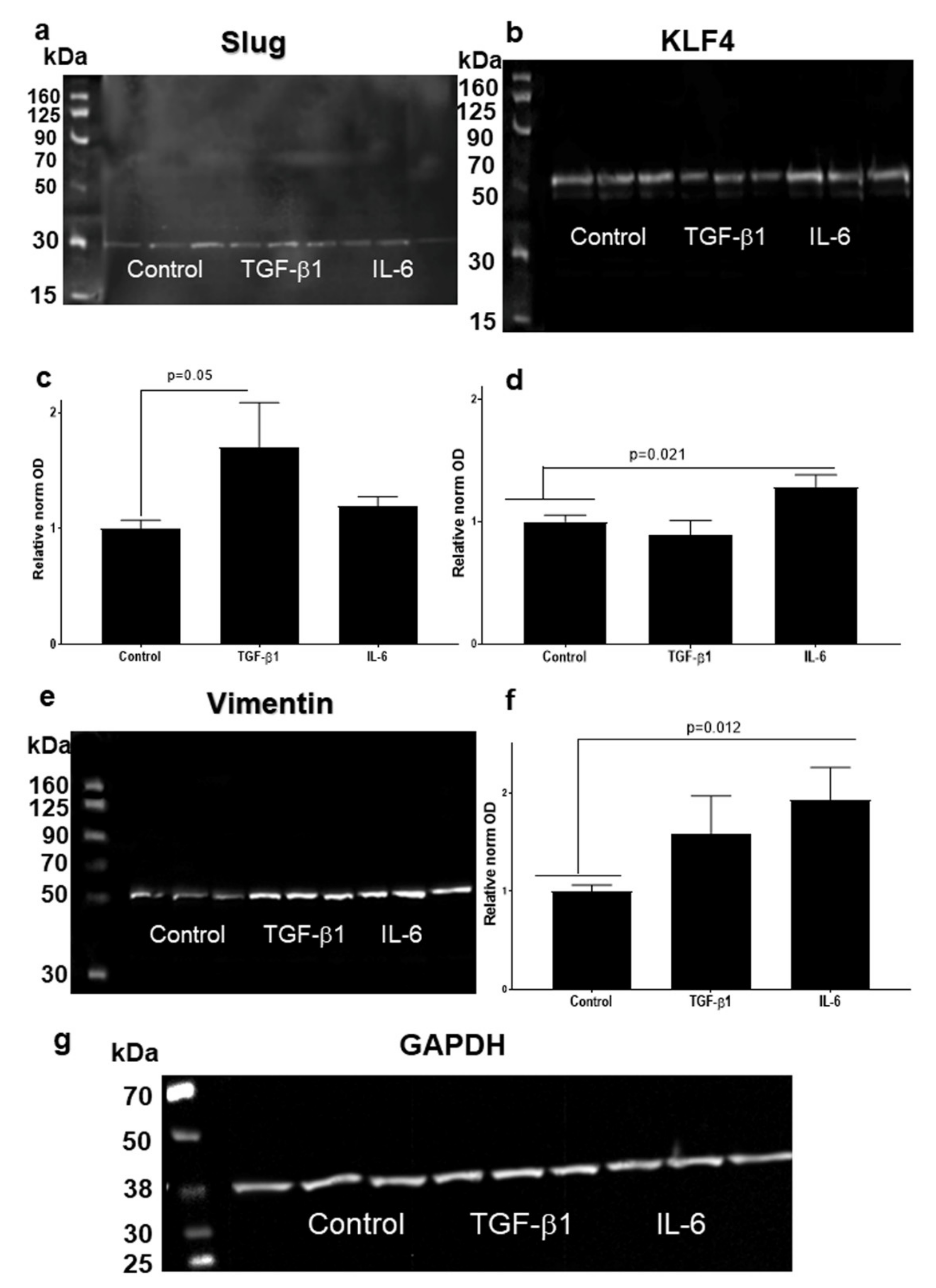
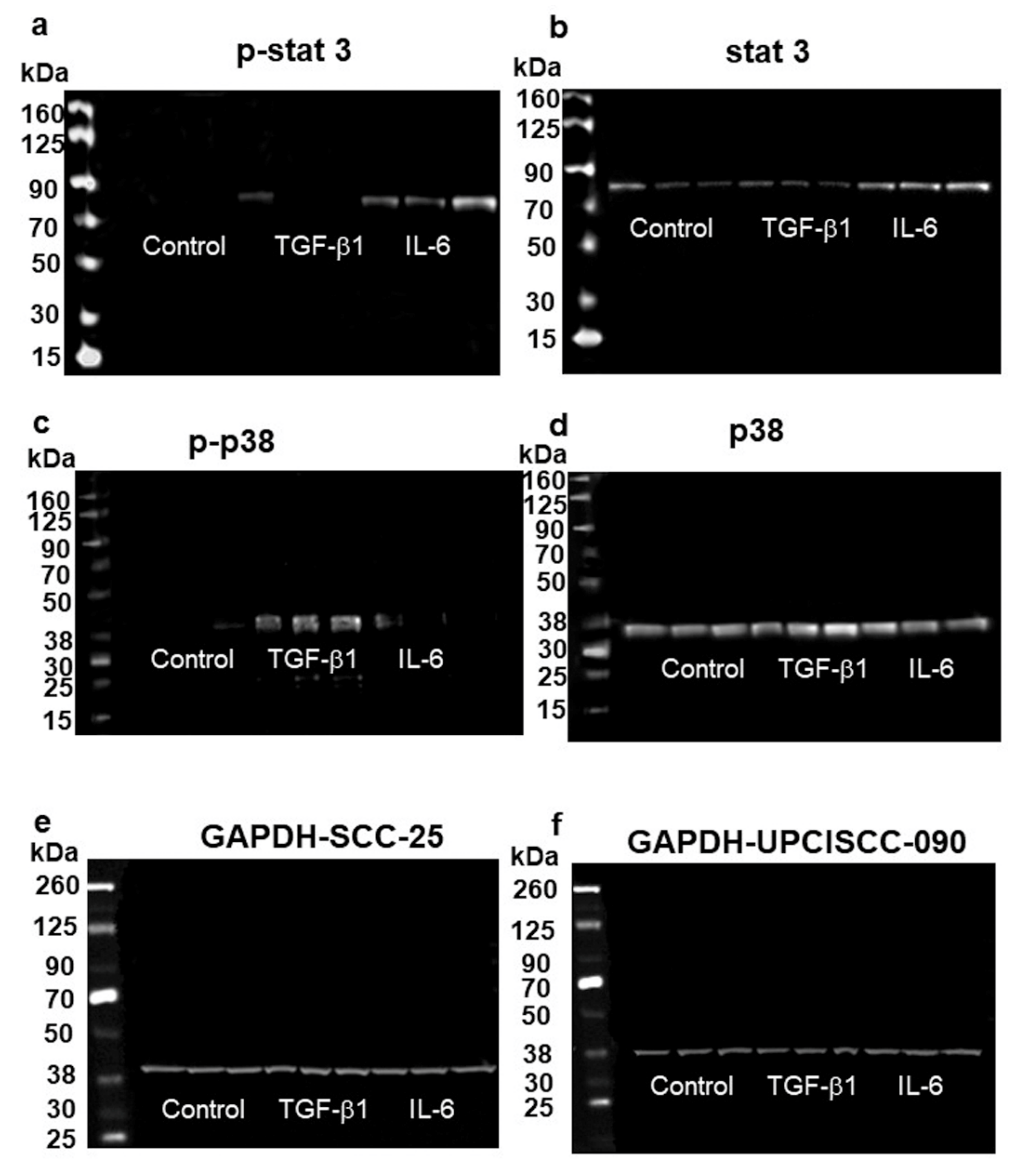
3.4. Higher Slug, and Vimentin, and Decreased KLF4 Protein Levels in HNSCC Cell Line Models Treated with TGF-β1
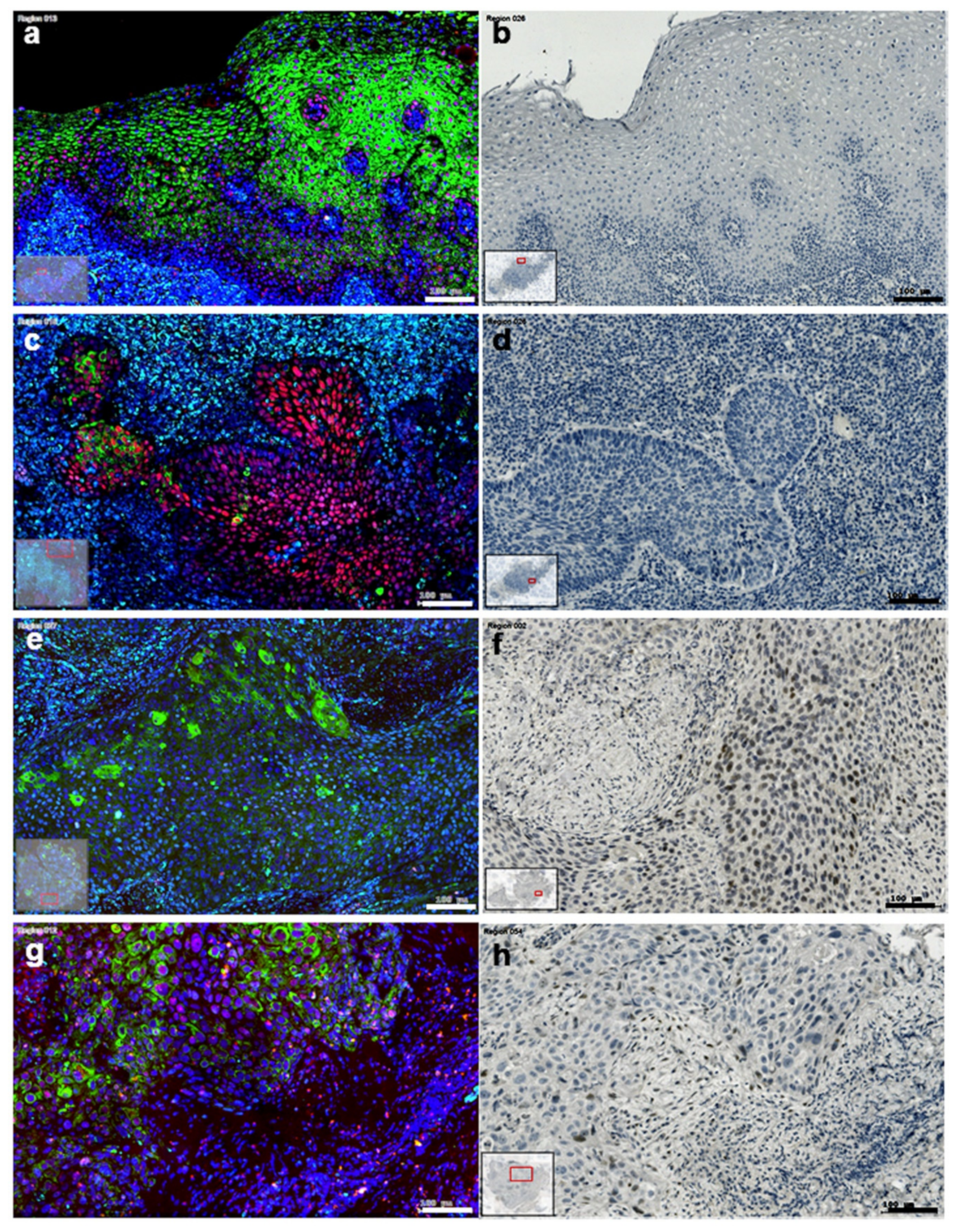
3.5. The Presentation of EMT-Factors KLF4 and Slug Is Focally Modified by Stroma in HNSCC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ASA score | American Society of Anesthesiologists classification |
| CT | Threshold Cycle |
| CHT | Chemotherapy |
| CAFs | Cancer-associated fibroblasts |
| CK | Cytokeratin |
| EMT | Epithelial-to-mesenchymal transition |
| GAPDH | Glycerin aldehyde dehydrogenase |
| HNSCC | Head and neck squamous cell carcinoma |
| HPV | Human papilloma virus |
| IL-6 | Interleukin-6 |
| KLFs | Krüppel-like factors |
| KLF4 | Krüppel-like-factor-4 |
| SLUG | Transcription Factor Slug |
| RT | Radiotherapy |
| RCHT | Radio-chemotherapy |
| RIT | Radioimmuntherapy |
| TF | Transcription Factor |
| TME | Tumor microenvironment |
| TGF-β1 | Transforming-growth-factor-beta-1 |
| UPPP | Uvulopalatopharyngoplasty |
| VIM | Vimentin |
References
- Hay, E.D. Organization and fine structure of epithelium and mesenchyme in the developing chick embryo. In Epithelial-Mesenchymal Interactions; Williams & Wilkins Co.: Baltimore, MD, USA, 1968; pp. 31–55. [Google Scholar]
- Nawshad, A.; Lagamba, D.; Polad, A.; Hay, E.D. Transforming growth factor-beta signaling during epithelial-mesenchymal transformation: Implications for embryogenesis and tumor metastasis. Cells Tissues Organs 2005, 179, 11–23. [Google Scholar] [CrossRef]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat. Rev. Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef]
- Derynck, R.; Weinberg, R.A. EMT and Cancer: More Than Meets the Eye. Dev. Cell 2019, 49, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, T. EMT and MET in metastasis: Where are the cancer stem cells? Cancer Cell 2012, 22, 699–701. [Google Scholar] [CrossRef]
- Taube, J.H.; Herschkowitz, J.I.; Komurov, K.; Zhou, A.Y.; Gupta, S.; Yang, J.; Hartwell, K.; Onder, T.T.; Gupta, P.B.; Evans, K.W.; et al. Core epithelial-to-mesenchymal transition interactome gene-expression signature is associated with claudin-low and metaplastic breast cancer subtypes. Proc. Natl. Acad. Sci. USA 2010, 107, 15449–15454. [Google Scholar] [CrossRef]
- Ye, X.; Tam, W.L.; Shibue, T.; Kaygusuz, Y.; Reinhardt, F.; Ng Eaton, E.; Weinberg, R.A. Distinct EMT programs control normal mammary stem cells and tumour-initiating cells. Nature 2015, 525, 256–260. [Google Scholar] [CrossRef]
- Moon, J.H.; Lee, S.H.; Koo, B.S.; Kim, J.M.; Huang, S.; Cho, J.H.; Eun, Y.G.; Shin, H.A.; Lim, Y.C. Slug is a novel molecular target for head and neck squamous cell carcinoma stem-like cells. Oral Oncol. 2020, 111, 104948. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Peng, Z.; Tang, H.; Xie, D.; Jia, Z.; Zhong, L.; Zhao, S.; Ma, Z.; Gao, Y.; Zeng, L.; et al. Loss of KLF4 and consequential downregulation of Smad7 exacerbate oncogenic TGF-beta signaling in and promote progression of hepatocellular carcinoma. Oncogene 2017, 36, 2957–2968. [Google Scholar] [CrossRef]
- McConnell, B.B.; Ghaleb, A.M.; Nandan, M.O.; Yang, V.W. The diverse functions of Kruppel-like factors 4 and 5 in epithelial biology and pathobiology. Bioessays 2007, 29, 549–557. [Google Scholar] [CrossRef]
- Li, W.; Liu, M.; Su, Y.; Zhou, X.; Liu, Y.; Zhang, X. The Janus-faced roles of Kruppel-like factor 4 in oral squamous cell carcinoma cells. Oncotarget 2015, 6, 44480–44494. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ghaleb, A.M.; Yang, V.W. Kruppel-like factor 4 (KLF4): What we currently know. Gene 2017, 611, 27–37. [Google Scholar] [CrossRef]
- Tiwari, N.; Meyer-Schaller, N.; Arnold, P.; Antoniadis, H.; Pachkov, M.; van Nimwegen, E.; Christofori, G. Klf4 is a transcriptional regulator of genes critical for EMT, including Jnk1 (Mapk8). PLoS ONE 2013, 8, e57329. [Google Scholar] [CrossRef]
- Curry, J.M.; Sprandio, J.; Cognetti, D.; Luginbuhl, A.; Bar-ad, V.; Pribitkin, E.; Tuluc, M. Tumor microenvironment in head and neck squamous cell carcinoma. Semin. Oncol. 2014, 41, 217–234. [Google Scholar] [CrossRef]
- Katafiasz, D.; Smith, L.M.; Wahl, J.K., 3rd. Slug (SNAI2) expression in oral SCC cells results in altered cell-cell adhesion and increased motility. Cell Adh. Migr. 2011, 5, 315–322. [Google Scholar] [CrossRef]
- Tai, S.K.; Yang, M.H.; Chang, S.Y.; Chang, Y.C.; Li, W.Y.; Tsai, T.L.; Wang, Y.F.; Chu, P.Y.; Hsieh, S.L. Persistent Kruppel-like factor 4 expression predicts progression and poor prognosis of head and neck squamous cell carcinoma. Cancer Sci. 2011, 102, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Saleh, K.; Eid, R.; Haddad, F.G.; Khalife-Saleh, N.; Kourie, H.R. New developments in the management of head and neck cancer—Impact of pembrolizumab. Ther. Clin. Risk Manag. 2018, 14, 295–303. [Google Scholar] [CrossRef]
- Gottgens, E.L.; Ostheimer, C.; Bussink, J.; Span, P.N.; Hammond, E. HPV, hypoxia and radiation response in head and neck cancer. Br. J. Radiol. 2018, 92, 20180047. [Google Scholar] [CrossRef]
- Salazar, C.R.; Smith, R.V.; Garg, M.K.; Haigentz, M., Jr.; Schiff, B.A.; Kawachi, N.; Anayannis, N.; Belbin, T.J.; Prystowsky, M.B.; Burk, R.D.; et al. Human papillomavirus-associated head and neck squamous cell carcinoma survival: A comparison by tumor site and initial treatment. Head Neck Pathol. 2014, 8, 77–87. [Google Scholar] [CrossRef]
- Devaraja, K.; Aggarwal, S.; Verma, S.S.; Gupta, S.C. Clinico-pathological peculiarities of human papilloma virus driven head and neck squamous cell carcinoma: A comprehensive update. Life Sci. 2020, 245, 117383. [Google Scholar] [CrossRef]
- Sisk, E.A.; Soltys, S.G.; Zhu, S.; Fisher, S.G.; Carey, T.E.; Bradford, C.R. Human papillomavirus and p53 mutational status as prognostic factors in head and neck carcinoma. Head Neck 2002, 24, 841–849. [Google Scholar] [CrossRef]
- Lin, Y.; Mallen-St Clair, J.; Luo, J.; Sharma, S.; Dubinett, S.; St John, M. p53 modulates NF-kappaB mediated epithelial-to-mesenchymal transition in head and neck squamous cell carcinoma. Oral Oncol. 2015, 51, 921–928. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Dudas, J.; Ingruber, J.; Glueckert, R.; Sprung, S.; Fleischer, F.; Cidlinsky, N.; Dejaco, D.; Kofler, B.; Giotakis, A.I.; et al. Slug Is A Surrogate Marker of Epithelial to Mesenchymal Transition (EMT) in Head and Neck Cancer. J. Clin. Med. 2020, 9, 2061. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Bender, B.; Giotakis, A.I.; Dejaco, D.; Url, C.; Riechelmann, H. Comparison of two surgical suture techniques in uvulopalatopharyngoplasty and expansion sphincter pharyngoplasty. Eur. Arch. Otorhinolaryngol. 2018, 275, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ferris, R.L.; Martinez, I.; Sirianni, N.; Wang, J.; Lopez-Albaitero, A.; Gollin, S.M.; Johnson, J.T.; Khan, S. Human papillomavirus-16 associated squamous cell carcinoma of the head and neck (SCCHN): A natural disease model provides insights into viral carcinogenesis. Eur. J. Cancer 2005, 41, 807–815. [Google Scholar] [CrossRef]
- Dudás, J.; Bitsche, M.; Schartinger, V.; Falkeis, C.; Sprinzl, G.M.; Riechelmann, H. Fibroblasts produce brain-derived neurotrophic factor and induce mesenchymal transition of oral tumor cells. Oral Oncol. 2011, 47, 98–103. [Google Scholar] [CrossRef]
- Dudás, J.; Fullár, A.; Romani, A.; Pritz, C.; Kovalszky, I.; Hans Schartinger, V.; Mathias Sprinzl, G.; Riechelmann, H. Curcumin targets fibroblast-tumor cell interactions in oral squamous cell carcinoma. Exp. Cell Res. 2013, 319, 800–809. [Google Scholar] [CrossRef]
- Fullár, A.; Kovalszky, I.; Bitsche, M.; Romani, A.; Schartinger, V.H.; Sprinzl, G.M.; Riechelmann, H.; Dudás, J. Tumor cell and carcinoma-associated fibroblast interaction regulates matrix metalloproteinases and their inhibitors in oral squamous cell carcinoma. Exp. Cell Res. 2012, 318, 1517–1527. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Metzler, V.; Pritz, C.; Riechelmann, H.; Dudas, J. Tumor-associated fibroblast-conditioned medium induces CDDP resistance in HNSCC cells. Oncotarget 2015, 7, 2508. [Google Scholar] [CrossRef]
- Dudás, J.; Fullár, A.; Bitsche, M.; Schartinger, V.; Kovalszky, I.; Sprinzl, G.M.; Riechelmann, H. Tumor-produced, active interleukin-1β regulates gene expression in carcinoma-associated fibroblasts. Exp. Cell Res. 2011, 317, 2222–2229. [Google Scholar] [CrossRef]
- Steinbichler, T.B.; Alshaimaa, A.; Maria, M.V.; Daniel, D.; Herbert, R.; Jozsef, D.; Ira-Ida, S. Epithelial-mesenchymal crosstalk induces radioresistance in HNSCC cells. Oncotarget 2018, 9, 3641–3652. [Google Scholar] [CrossRef]
- Metzler, V.M.; Pritz, C.; Riml, A.; Romani, A.; Tuertscher, R.; Steinbichler, T.; Dejaco, D.; Riechelmann, H.; Dudas, J. Separation of cell survival, growth, migration, and mesenchymal transdifferentiation effects of fibroblast secretome on tumor cells of head and neck squamous cell carcinoma. Tumour Biol. 2017, 39, 1010428317705507. [Google Scholar] [CrossRef]
- Dudas, J.; Dietl, W.; Romani, A.; Reinold, S.; Glueckert, R.; Schrott-Fischer, A.; Dejaco, D.; Johnson Chacko, L.; Tuertscher, R.; Schartinger, V.H.; et al. Nerve Growth Factor (NGF)-Receptor Survival Axis in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2018, 19, 1771. [Google Scholar] [CrossRef]
- Kofler, B.; Borena, W.; Manzl, C.; Dudas, J.; Wegscheider, A.S.; Jansen-Durr, P.; Schartinger, V.; Riechelmann, H. Sensitivity of tumor surface brushings to detect human papilloma virus DNA in head and neck cancer. Oral Oncol. 2017, 67, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Dudas, J.; Riml, A.; Tuertscher, R.; Pritz, C.; Steinbichler, T.B.; Schartinger, V.H.; Sprung, S.; Glueckert, R.; Schrott-Fischer, A.; Johnson Chacko, L.; et al. Brain-Derived Neurotrophin and TrkB in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2019, 20, 272. [Google Scholar] [CrossRef] [PubMed]
- Bouchalova, P.; Nenutil, R.; Muller, P.; Hrstka, R.; Appleyard, M.V.; Murray, K.; Jordan, L.B.; Purdie, C.A.; Quinlan, P.; Thompson, A.M.; et al. Mutant p53 accumulation in human breast cancer is not an intrinsic property or dependent on structural or functional disruption but is regulated by exogenous stress and receptor status. J. Pathol. 2014, 233, 238–246. [Google Scholar] [CrossRef]
- Wang, S.P.; Wang, W.L.; Chang, Y.L.; Wu, C.T.; Chao, Y.C.; Kao, S.H.; Yuan, A.; Lin, C.W.; Yang, S.C.; Chan, W.K.; et al. p53 controls cancer cell invasion by inducing the MDM2-mediated degradation of Slug. Nat. Cell Biol. 2009, 11, 694–704. [Google Scholar] [CrossRef]
- Wu, M.; Chen, G.; Li, Y.P. TGF-beta and BMP signaling in osteoblast, skeletal development, and bone formation, homeostasis and disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sex | male | 36 | 97% |
| female | 1 | 3% | |
| age | ≤50 | 1 | 3% |
| 51–60 | 7 | 19% | |
| 61–70 | 21 | 57% | |
| 71–80 | 4 | 11% | |
| >80 | 4 | 11% | |
| ASA score | ASA I/II | 26 | 70% |
| ASA III/IV | 11 | 30% | |
| tumor site | lips/oral cavity | 4 | 11% |
| oropharynx | 16 | 43% | |
| hypopharynx | 7 | 19% | |
| Larynx | 10 | 27% | |
| histology | squamous cell carcinoma | 37 | 100% |
| HPV/tumor localization | P16-positive oropharynx SCC | 9 | 56.25% |
| P16-negative oropharynx SCC | 7 | 43.75% | |
| P16-positive larynx SCC | 1 | 10% | |
| P16-negative larynx SCC | 9 | 90% | |
| P16-negative SCC of lips/oral cavity | 4 | 100% | |
| P16-negative SCC of hypopharynx | 7 | 100% | |
| UICC stage | stage 1 | 4 | 11% |
| stage 2 | 2 | 5% | |
| stage 3 | 11 | 30% | |
| stage 4a | 17 | 46% | |
| stage 4b | 2 | 5% | |
| stage 4c | 1 | 3% | |
| HPV status | negative (<70%) | 27 | 73% |
| positive (≥70%) | 10 | 27% | |
| P53 status | regular | 11 | 30% |
| irregular | 26 | 70% | |
| treatment modality | surgery | 9 | 24% |
| surgery & RT | 7 | 19% | |
| surgery & RCHT/RIT | 1 | 3% | |
| RCHT/RIT | 18 | 49% | |
| RT | 1 | 3% | |
| CHT | 1 | 3% |
| Antibody | Isotype | Catalogue Number | Manufacturer | Dilution |
|---|---|---|---|---|
| pan-cytokeratin | IgG1 | 760-2595 | Roche Ventana, Mannheim, Germany | pre-diluted |
| anti-vimentin, (clone VI-10) | IgM | EX-11-460-C100 | Exbio, Prague, Czech Republic | 1:200 |
| anti-KLF4 | IgG | ab215036 | Abcam, Cambridge, UK | 1:2000 |
| anti-slug | IgG1 | 564614 | BD Pharmingen Austria, Vienna Austria | 1:400 |
| anti-TGFβ1 | IgG | AB190503 | Abcam, Cambridge, UK | 1:100 |
| Isotype-control mouse | IgG1 | 11-632-C100 | Exbio, Prague, Czech Republic | 1:100 |
| Isotype-control mouse | IgM | 11-803-C100 | Exbio, Prague, Czech Republic | 1:100 |
| Isotype-control rabbit | IgG | 02-6102 | Invitrogen Life Technologies, Darmstadt, Germany | 1:400 |
| Antibody | Isotyp | Catalogue Number | kDa | Manufacturer | Dilution |
|---|---|---|---|---|---|
| anti-Slug | IgG1 | 9585 | 30 | Cell Signalling Technologies, Danvers, MA, USA | 1:1000 |
| anti-Slug | IgG1,K | 564614 | 30 | BD Pharmingen, Szabo Scandic, Vienna Austria | 1:400 |
| anti-vimentin (clone SP-20) | IgG | M3202 | 53 | Spring Biotech, Linaris, Germany | 1:100 |
| anti-GAPDH | IgG1 | ab8245 | 36 | Abcam, Cambridge, UK | 1:5000 |
| anti-KLF4 | IgG1 | ab215036 | 54 | Abcam, Cambridge, UK | 1:1000 |
| anti-p38 MAPK | IgG | 8690S | 40 | Cell Signalling Technologies, Danvers, MA, USA | 1:1000 |
| anti-phospho-p38 MAPK | IgG | 4511S | 43 | Cell Signalling Technologies, Danvers, MA, USA | 1:1000 |
| anti-Smad2/3 | IgG | 3102 | 52.6 | Cell Signalling Technologies, Danvers, MA, USA | 1:1000 |
| anti-phospho-Smad2 | IgG | 3108 | 60 | Cell Signalling Technologies, Danvers, MA, USA | 1:1000 |
| anti-Stat3 | IgG | 12640 | 79.86 | Cell Signalling Technologies, Danvers, MA, USA | 1:1000 |
| anti-phospho-Stat3 | IgG | 9145 | 79.86 | Cell Signalling Technologies, Danvers, MA, USA | 1:2000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ingruber, J.; Savic, D.; Steinbichler, T.B.; Sprung, S.; Fleischer, F.; Glueckert, R.; Schweigl, G.; Skvortsova, I.-I.; Riechelmann, H.; Dudás, J. KLF4, Slug and EMT in Head and Neck Squamous Cell Carcinoma. Cells 2021, 10, 539. https://doi.org/10.3390/cells10030539
Ingruber J, Savic D, Steinbichler TB, Sprung S, Fleischer F, Glueckert R, Schweigl G, Skvortsova I-I, Riechelmann H, Dudás J. KLF4, Slug and EMT in Head and Neck Squamous Cell Carcinoma. Cells. 2021; 10(3):539. https://doi.org/10.3390/cells10030539
Chicago/Turabian StyleIngruber, Julia, Dragana Savic, Teresa Bernadette Steinbichler, Susanne Sprung, Felix Fleischer, Rudolf Glueckert, Gabriele Schweigl, Ira-Ida Skvortsova, Herbert Riechelmann, and József Dudás. 2021. "KLF4, Slug and EMT in Head and Neck Squamous Cell Carcinoma" Cells 10, no. 3: 539. https://doi.org/10.3390/cells10030539
APA StyleIngruber, J., Savic, D., Steinbichler, T. B., Sprung, S., Fleischer, F., Glueckert, R., Schweigl, G., Skvortsova, I.-I., Riechelmann, H., & Dudás, J. (2021). KLF4, Slug and EMT in Head and Neck Squamous Cell Carcinoma. Cells, 10(3), 539. https://doi.org/10.3390/cells10030539