
Based on a Self-Feeder Layer, a Novel 3D Culture Model of Human ADSCs Facilitates Trans-Differentiation of the Spheroid Cells into Neural Progenitor-Like Cells Using siEID3 with a Laminin/Poly-d-lysine Matrix

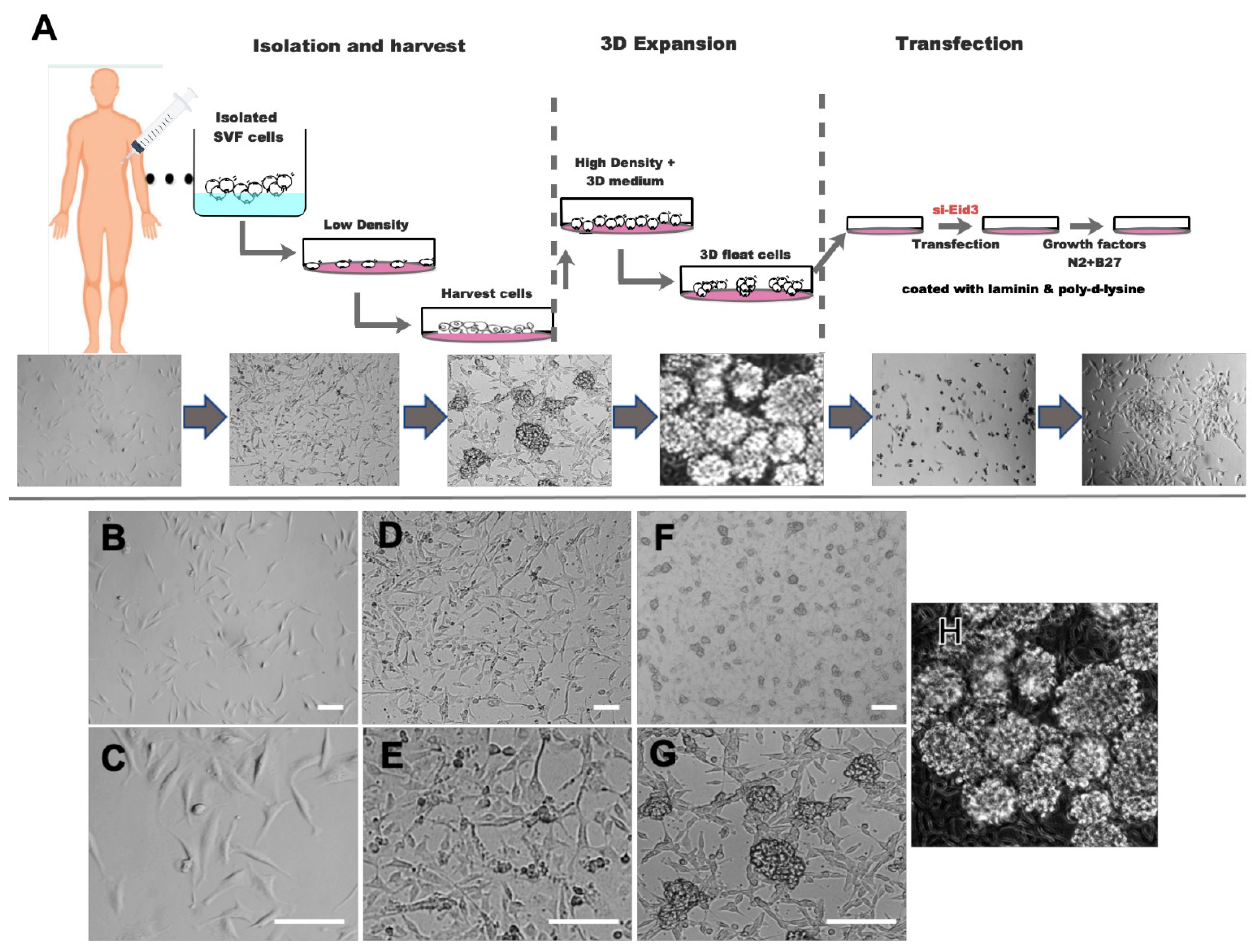{kind=link}
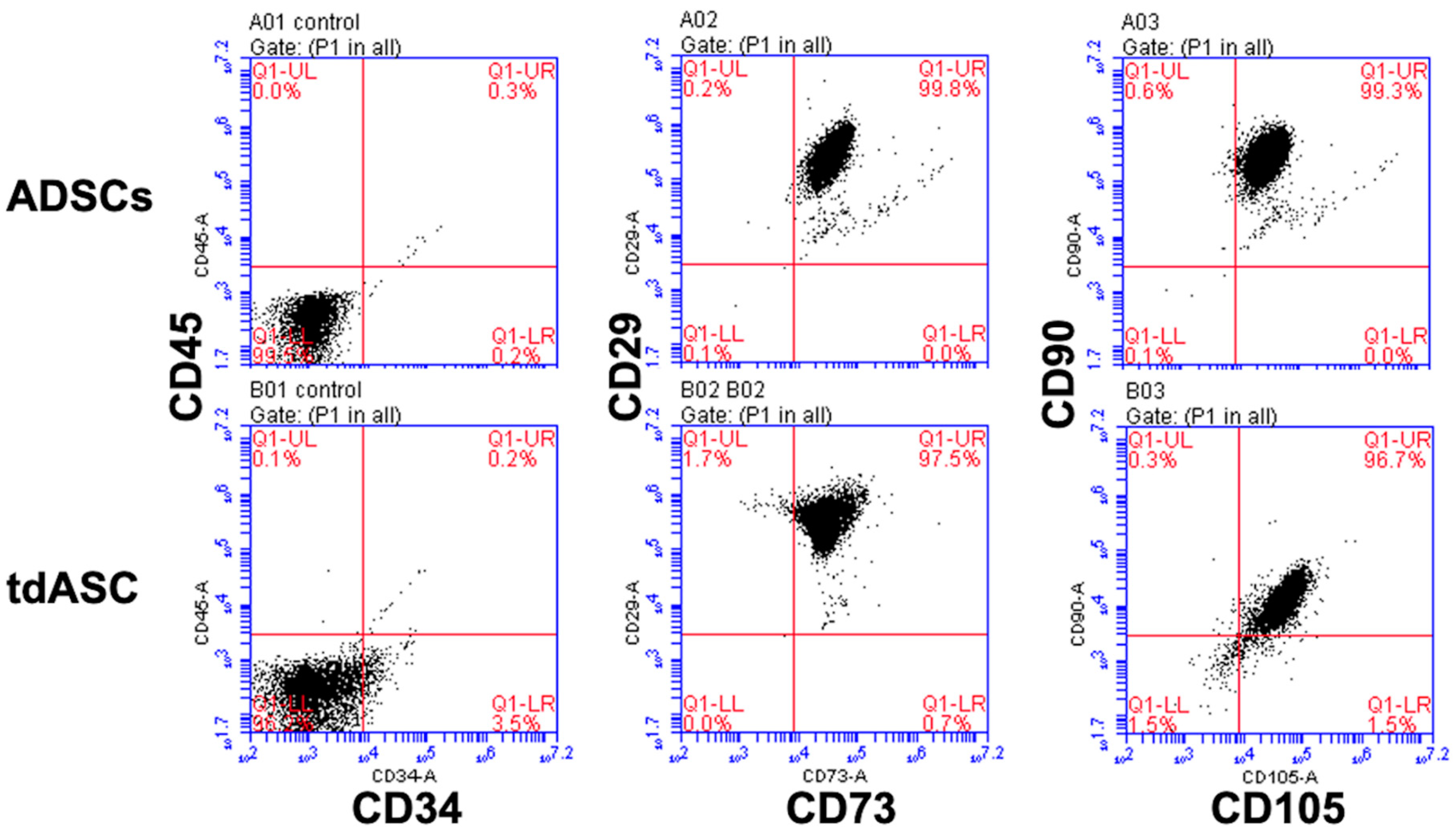{kind=link}
{kind=link}
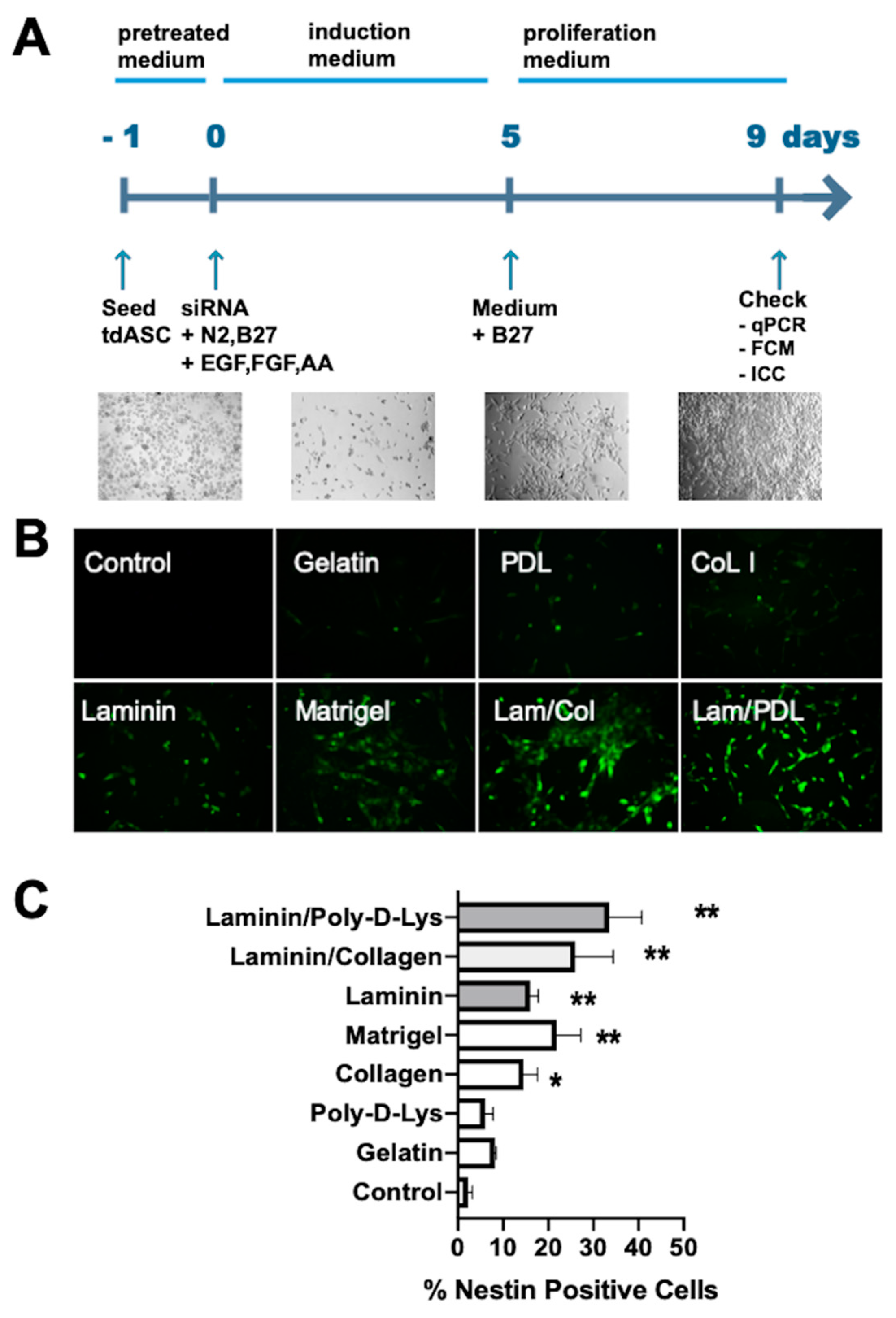{kind=link}
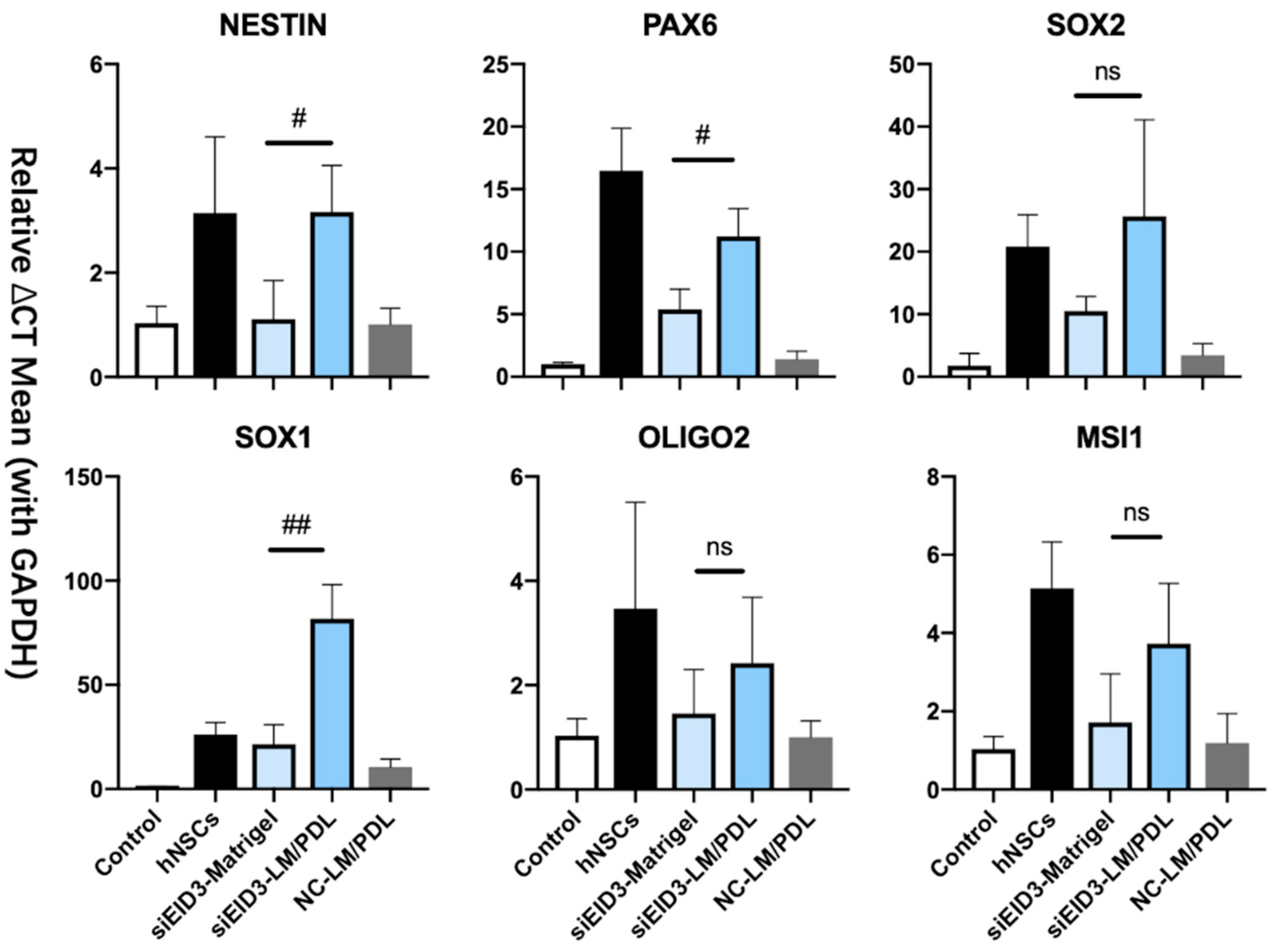{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Monolayer Culture of Human ADSCs and Human Neural Stem Cells (NSCs) Culture
2.2. Three-Dimension Adipose Stem Cells (tdASC) Spheroid Culture
2.3. Cell Counting Kit-8 (CCK-8) Assays
2.4. Proliferation Assays
2.5. Alkaline Phosphatase (ALP) Staining and Alkaline Phosphatase Activity
2.6. Immunophenotypic Characterization by Flow Cytometry
2.7. siEID3 Transfect and Generation of NPC-Like Cells from tdASC
2.8. Substrates Preparation
2.9. Neural Stem Cell Differentiation
2.10. Immunocytochemistry and Confocal Microscopy
2.11. RNA Extraction, Quantitative Real-Time RT-PCR Analysis
2.12. Statistical Analysis
3. Results
3.1. Three-Dimensional ASC Spheroid (tdASC) Expansion and Differentiation
3.2. tdASC Cell Proliferation and Population Senescence Analysis
3.3. tdASC Stemness Gene Expression
3.4. Identification of the Optimal Matrix for tdASC Induction
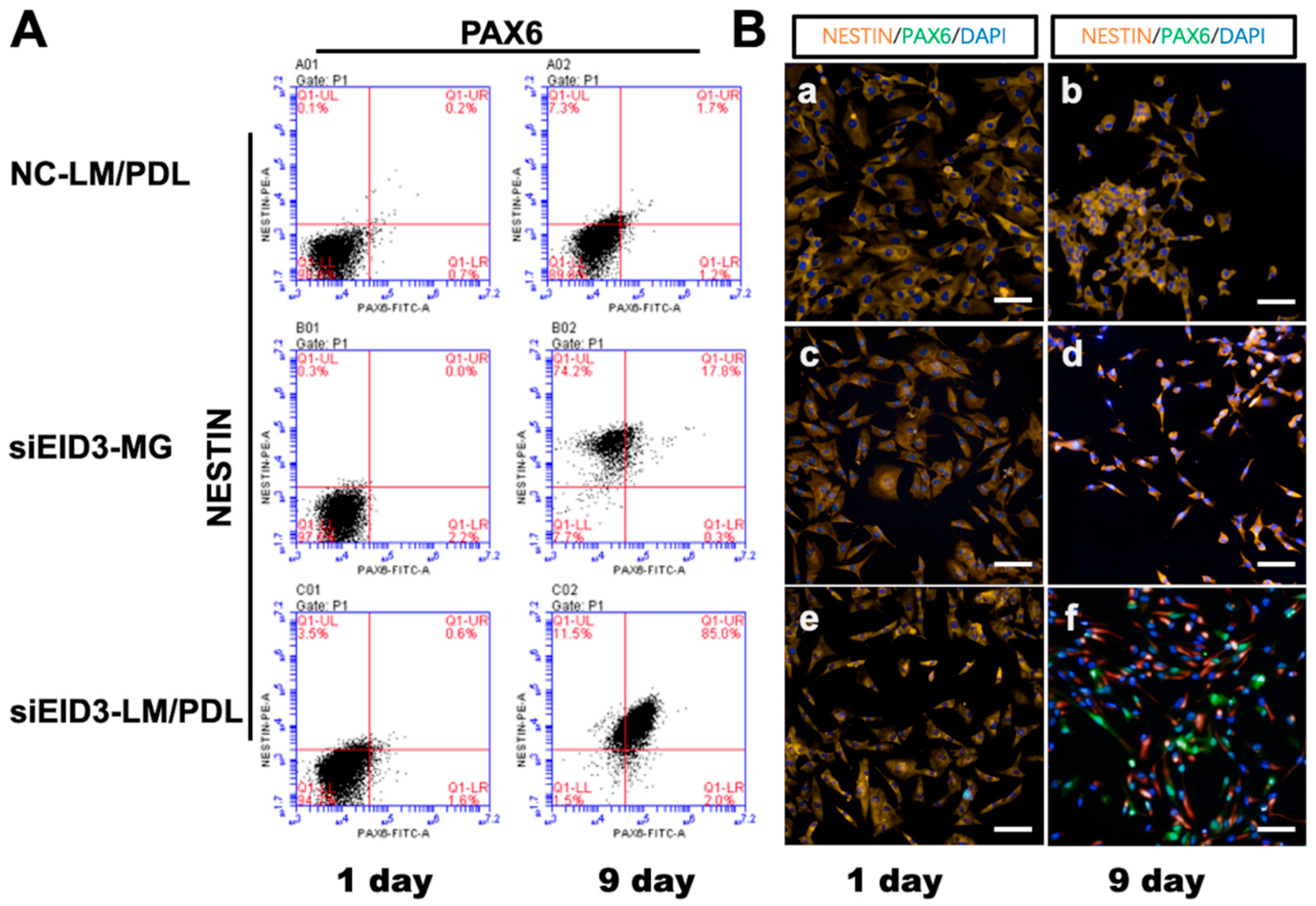
3.5. Generation and Characterization of iNPLCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef]
- Si, Z.; Wang, X.; Sun, C.; Kang, Y.; Xu, J.; Wang, X.; Hui, Y. Adipose-Derived Stem Cells: Sources, Potency, and Implications for Regenerative Therapies. Biomed. Pharmacother. 2019, 114, 108765. [Google Scholar] [CrossRef] [PubMed]
- Bateman, M.E.; Strong, A.L.; Gimble, J.M.; Bunnell, B.A. Concise Review: Using Fat to Fight Disease: A Systematic Review of Nonhomologous Adipose-Derived Stromal/Stem Cell Therapies. STEM CELLS Transl. Med. 2018, 1311–1328. [Google Scholar] [CrossRef] [PubMed]
- Rondini, E.A.; Granneman, J.G. Single Cell Approaches to Address Adipose Tissue Stromal Cell Heterogeneity. Biochem. J. 2020, 477, 583–600. [Google Scholar] [CrossRef] [PubMed]
- Francis, M.P.; Sachs, P.C.; Elmore, L.W.; Holt, S.E. Isolating Adipose-Derived Mesenchymal Stem Cells from Lipoaspirate Blood and Saline Fraction. Organogenesis 2010, 6, 11–14. [Google Scholar] [CrossRef]
- Bacakova, L.; Zarubova, J.; Travnickova, M.; Musilkova, J.; Pajorova, J.; Slepicka, P.; Kasalkova, N.S.; Svorcik, V.; Kolska, Z.; Motarjemi, H.; et al. Stem Cells: Their Source, Potency and Use in Regenerative Therapies with Focus on Adipose-Derived Stem Cells—A Review. Biotechnol. Adv. 2018, 36, 1111–1126. [Google Scholar] [CrossRef]
- Sekiya, I.; Larson, B.L.; Smith, J.R.; Pochampally, R.; Cui, J.-G.; Prockop, D.J. Expansion of Human Adult Stem Cells from Bone Marrow Stroma: Conditions That Maximize the Yields of Early Progenitors and Evaluate Their Quality. STEM CELLS 2002, 20, 530–541. [Google Scholar] [CrossRef]
- Seo, Y.; Shin, T.-H.; Kim, H.-S. Current Strategies to Enhance Adipose Stem Cell Function: An Update. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Prieto González, E.A. Heterogeneity in Adipose Stem Cells. In Stem Cells Heterogeneity—Novel Concepts; Advances in Experimental Medicine and Biology; Birbrair, A., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 119–150. ISBN 978-3-030-11096-3. [Google Scholar]
- Ryu, N.-E.; Lee, S.-H.; Park, H. Spheroid Culture System Methods and Applications for Mesenchymal Stem Cells. Cells 2019, 8. [Google Scholar] [CrossRef]
- Ravi, M.; Paramesh, V.; Kaviya, S.R.; Anuradha, E.; Solomon, F.P. 3D Cell Culture Systems: Advantages and Applications. J. Cell. Physiol. 2015, 230, 16–26. [Google Scholar] [CrossRef]
- Muller, S.; Ader, I.; Creff, J.; Leménager, H.; Achard, P.; Casteilla, L.; Sensebé, L.; Carrière, A.; Deschaseaux, F. Human Adipose Stromal-Vascular Fraction Self-Organizes to Form Vascularized Adipose Tissue in 3D Cultures. Sci. Rep. 2019, 9, 7250. [Google Scholar] [CrossRef] [PubMed]
- Sung, T.-C.; Heish, C.-W.; Lee, H.H.-C.; Hsu, J.-Y.; Wang, C.-K.; Wang, J.-H.; Zhu, Y.-R.; Jen, S.-H.; Hsu, S.-T.; Hirad, A.H.; et al. 3D Culturing of Human Adipose-Derived Stem Cells Enhances Their Pluripotency and Differentiation Abilities. J. Mater. Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Yin, Q.; Xu, N.; Xu, D.; Dong, M.; Shi, X.; Wang, Y.; Hao, Z.; Zhu, S.; Zhao, D.; Jin, H.; et al. Comparison of Senescence-Related Changes between Three- and Two-Dimensional Cultured Adipose-Derived Mesenchymal Stem Cells. Stem Cell Res. Ther. 2020, 11, 226. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Chen, W.-J.; Yin, J.Q.; Xu, R.-X. EID3 Directly Associates with DNMT3A during Transdifferentiation of Human Umbilical Cord Mesenchymal Stem Cells to NPC-like Cells. Sci. Rep. 2017, 7, 40463. [Google Scholar] [CrossRef]
- Chakravarti, D.; Ogryzko, V.; Kao, H.-Y.; Nash, A.; Chen, H.; Nakatani, Y.; Evans, R.M. A Viral Mechanism for Inhibition of P300 and PCAF Acetyltransferase Activity. Cell 1999, 96, 393–403. [Google Scholar] [CrossRef]
- Båvner, A.; Matthews, J.; Sanyal, S.; Gustafsson, J.-A.; Treuter, E. EID3 Is a Novel EID Family Member and an Inhibitor of CBP-Dependent Co-Activation. Nucleic Acids Res. 2005, 33, 3561–3569. [Google Scholar] [CrossRef]
- Zhang, W.; Bai, X.; Zhao, B.; Li, Y.; Zhang, Y.; Li, Z.; Wang, X.; Luo, L.; Han, F.; Zhang, J.; et al. Cell-Free Therapy Based on Adipose Tissue Stem Cell-Derived Exosomes Promotes Wound Healing via the PI3K/Akt Signaling Pathway. Exp. Cell Res. 2018, 370, 333–342. [Google Scholar] [CrossRef]
- Lund, P.; Pilgaard, L.; Duroux, M.; Fink, T.; Zachar, V. Effect of Growth Media and Serum Replacements on the Proliferation and Differentiation of Adipose-Derived Stem Cells. Cytotherapy 2009, 11, 189–197. [Google Scholar] [CrossRef]
- Cristofalo, V.J.; Allen, R.G.; Pignolo, R.J.; Martin, B.G.; Beck, J.C. Relationship between Donor Age and the Replicative Lifespan of Human Cells in Culture: A Reevaluation. Proc. Natl. Acad. Sci. USA 1998, 95, 10614–10619. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.-R.; Shin, J.-H.; Kim, J.-J.; Koog, M.G.; Lee, J.Y.; Choi, S.W.; Kim, H.-S.; Seo, Y.; Lee, S.; Shin, T.; et al. Rapid and Efficient Direct Conversion of Human Adult Somatic Cells into Neural Stem Cells by HMGA2/Let-7b. Cell Rep. 2015, 10, 441–452. [Google Scholar] [CrossRef]
- Zhang, R.; Li, Y.; Hu, B.; Lu, Z.; Zhang, J.; Zhang, X. Traceable Nanoparticle Delivery of Small Interfering RNA and Retinoic Acid with Temporally Release Ability to Control Neural Stem Cell Differentiation for Alzheimer’s Disease Therapy. Adv. Mater. 2016, 28, 6345–6352. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Ouyang, K.; Huang, J.; Zhou, Y.; Ouyang, H.; Li, H.; Wang, G.; Wu, Q.; Wei, C.; Bi, Y. Direct Conversion of Fibroblasts to Neurons by Reprogramming PTB-Regulated MicroRNA Circuits. Cell 2013, 152, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, X.; Josey, B.; Chou, C.J.; Tan, Y.; Zhang, N.; Wen, X. Short Laminin Peptide for Improved Neural Stem Cell Growth. STEM CELLS Transl. Med. 2014, 3, 662–670. [Google Scholar] [CrossRef]
- Wilems, T.; Vardhan, S.; Wu, S.; Sakiyama-Elbert, S. The Influence of Microenvironment and Extracellular Matrix Molecules in Driving Neural Stem Cell Fate within Biomaterials. Brain Res. Bull. 2019, 148, 25–33. [Google Scholar] [CrossRef]
- Kleinman, H.K.; Martin, G.R. Matrigel: Basement Membrane Matrix with Biological Activity. Semin. Cancer Biol. 2005, 15, 378–386. [Google Scholar] [CrossRef]
- Wang, J.; Chu, R.; Ni, N.; Nan, G. The Effect of Matrigel as Scaffold Material for Neural Stem Cell Transplantation for Treating Spinal Cord Injury. Sci. Rep. 2020, 10, 2576. [Google Scholar] [CrossRef]
- Wang, Y.-H.; Guo, Y.-C.; Wang, D.-R.; Liu, J.-Y.; Pan, J. Adipose Stem Cell-Based Clinical Strategy for Neural Regeneration: A Review of Current Opinion. Stem Cells Int. 2019, 2019, 8502370. [Google Scholar] [CrossRef]
- Fesharaki, M.; Razavi, S.; Ghasemi-Mobarakeh, L.; Behjati, M.; Yarahmadian, R.; Kazemi, M.; Hossein, H. Differentiation of Human Scalp Adipose-Derived Mesenchymal Stem Cells into Mature Neural Cells on Electrospun Nanofibrous Scaffolds for Nerve Tissue Engineering Applications. Cell J. 2018, 20, 168–176. [Google Scholar] [CrossRef]
- Luca, M.D.; Aiuti, A.; Cossu, G.; Parmar, M.; Pellegrini, G.; Robey, P.G. Advances in Stem Cell Research and Therapeutic Development. Nat. Cell Biol. 2019, 21, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Hu, D.-H.; Yin, J.Q.; Xu, R.-X. Molecular Mechanisms of Transdifferentiation of Adipose-Derived Stem Cells into Neural Cells: Current Status and Perspectives. Stem Cells Int. 2018, 2018, 5630802. [Google Scholar] [CrossRef]
- Cholewa, D.; Stiehl, T.; Schellenberg, A.; Bokermann, G.; Joussen, S.; Koch, C.; Walenda, T.; Pallua, N.; Marciniak-Czochra, A.; Suschek, C.V. Expansion of Adipose Mesenchymal Stromal Cells Is Affected by Human Platelet Lysate and Plating Density. Cell Transplant. 2011, 20, 1409–1422. [Google Scholar] [CrossRef]
- Houlihan, D.D.; Mabuchi, Y.; Morikawa, S.; Niibe, K.; Araki, D.; Suzuki, S.; Okano, H.; Matsuzaki, Y. Isolation of Mouse Mesenchymal Stem Cells on the Basis of Expression of Sca-1 and PDGFR-α. Nat. Protoc. 2012, 7, 2103. [Google Scholar] [CrossRef] [PubMed]
- Falcon, N.D.; Riley, G.P.; Saeed, A. Induction of Tendon-Specific Markers in Adipose-Derived Stem Cells in Serum-Free Culture Conditions. Tissue Eng. Part C Methods 2019, 25, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Legzdina, D.; Romanauska, A.; Nikulshin, S.; Kozlovska, T.; Berzins, U. Characterization of Senescence of Culture-Expanded Human Adipose-Derived Mesenchymal Stem Cells. Int. J. Stem Cells 2016, 9, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Horn, P.; Castoldi, M.; Diehlmann, A.; Bork, S.; Saffrich, R.; Benes, V.; Blake, J.; Pfister, S.; Eckstein, V.; et al. Replicative Senescence of Mesenchymal Stem Cells: A Continuous and Organized Process. PLoS ONE 2008, 3, e2213. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Zhao, Y.; Wei, J.; Han, J.; Chen, L.; Xiao, Z.; Chen, B.; Dai, J. The Effect of Forced Growth of Cells into 3D Spheres Using Low Attachment Surfaces on the Acquisition of Stemness Properties. Biomaterials 2013, 34, 3215–3222. [Google Scholar] [CrossRef]
- Toma, J.G.; Akhavan, M.; Fernandes, K.J.L.; Barnabé-Heider, F.; Sadikot, A.; Kaplan, D.R.; Miller, F.D. Isolation of Multipotent Adult Stem Cells from the Dermis of Mammalian Skin. Nat. Cell Biol. 2001, 3, 778–784. [Google Scholar] [CrossRef]
- Hunsberger, J.G.; Rao, M.; Kurtzberg, J.; Bulte, J.W.M.; Atala, A.; LaFerla, F.M.; Greely, H.T.; Sawa, A.; Gandy, S.; Schneider, L.S.; et al. Accelerating Stem Cell Trials for Alzheimer’s Disease. Lancet Neurol. 2016, 15, 219–230. [Google Scholar] [CrossRef]
- Jakel, R.J.; Schneider, B.L.; Svendsen, C.N. Using Human Neural Stem Cells to Model Neurological Disease. Nat. Rev. Genet. 2004, 5, 136–144. [Google Scholar] [CrossRef]
- Flanagan, L.A.; Rebaza, L.M.; Derzic, S.; Schwartz, P.H.; Monuki, E.S. Regulation of Human Neural Precursor Cells by Laminin and Integrins. J. Neurosci. Res. 2006, 83, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Viale-Bouroncle, S.; Gosau, M.; Morsczeck, C. Laminin Regulates the Osteogenic Differentiation of Dental Follicle Cells via Integrin-A2/-Β1 and the Activation of the FAK/ERK Signaling Pathway. Cell Tissue Res. 2014, 357, 345–354. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, L.; Zhang, W.; Chen, W.; Fu, X.; Wang, X.; Xu, R.; Hu, D. Based on a Self-Feeder Layer, a Novel 3D Culture Model of Human ADSCs Facilitates Trans-Differentiation of the Spheroid Cells into Neural Progenitor-Like Cells Using siEID3 with a Laminin/Poly-d-lysine Matrix. Cells 2021, 10, 493. https://doi.org/10.3390/cells10030493
Luo L, Zhang W, Chen W, Fu X, Wang X, Xu R, Hu D. Based on a Self-Feeder Layer, a Novel 3D Culture Model of Human ADSCs Facilitates Trans-Differentiation of the Spheroid Cells into Neural Progenitor-Like Cells Using siEID3 with a Laminin/Poly-d-lysine Matrix. Cells. 2021; 10(3):493. https://doi.org/10.3390/cells10030493
Chicago/Turabian StyleLuo, Liang, Wei Zhang, Wenjin Chen, Xiaojun Fu, Xujie Wang, Ruxiang Xu, and Dahai Hu. 2021. "Based on a Self-Feeder Layer, a Novel 3D Culture Model of Human ADSCs Facilitates Trans-Differentiation of the Spheroid Cells into Neural Progenitor-Like Cells Using siEID3 with a Laminin/Poly-d-lysine Matrix" Cells 10, no. 3: 493. https://doi.org/10.3390/cells10030493
APA StyleLuo, L., Zhang, W., Chen, W., Fu, X., Wang, X., Xu, R., & Hu, D. (2021). Based on a Self-Feeder Layer, a Novel 3D Culture Model of Human ADSCs Facilitates Trans-Differentiation of the Spheroid Cells into Neural Progenitor-Like Cells Using siEID3 with a Laminin/Poly-d-lysine Matrix. Cells, 10(3), 493. https://doi.org/10.3390/cells10030493