
Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview


 , and
, and 
Abstract
1. Introduction
2. Microalgae Nutrient Composition
2.1. Role of Nitrogen
2.2. Role of Phosphorus
3. Microalgae Cultivation Medium
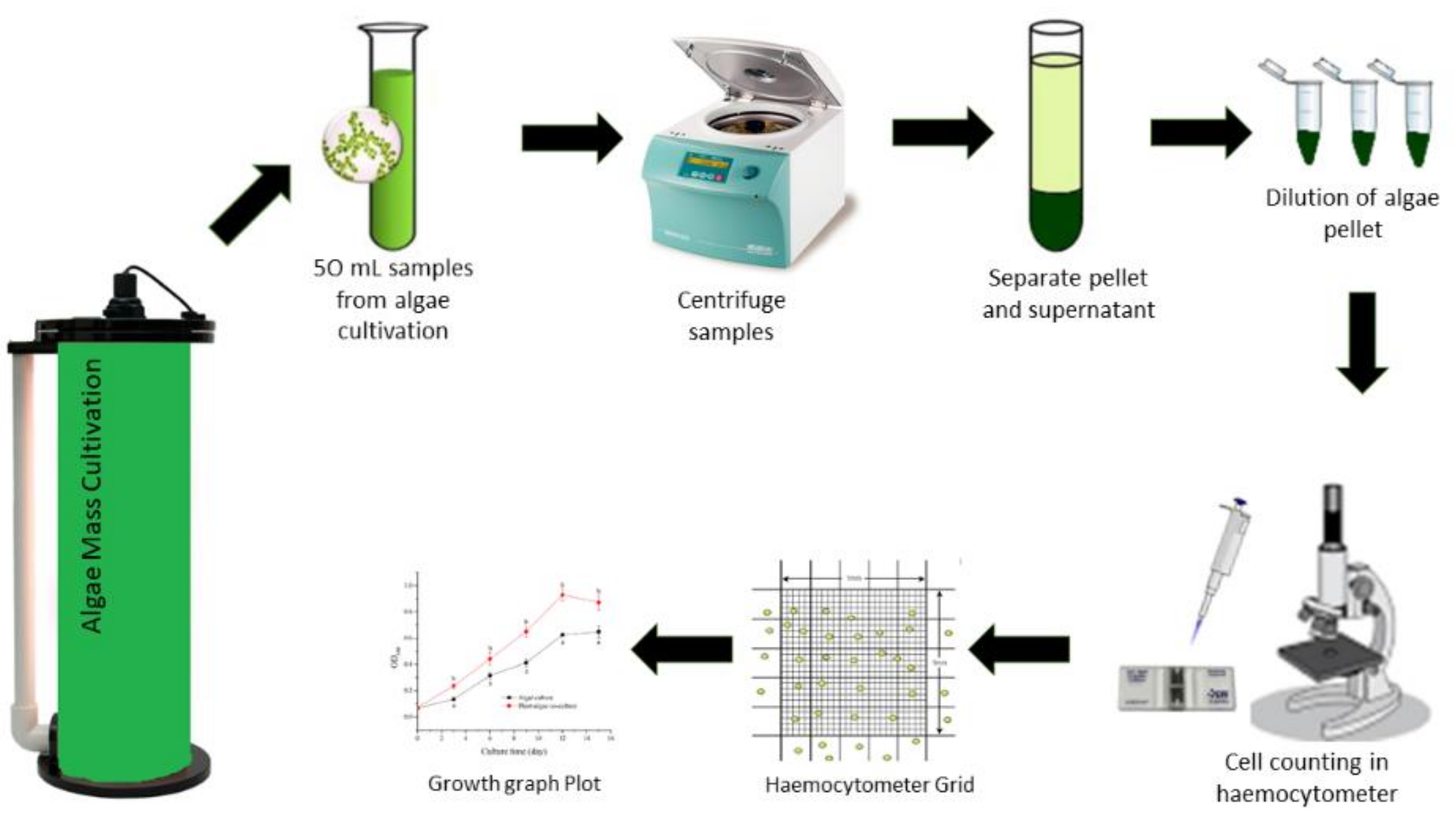
3.1. Growth Rates
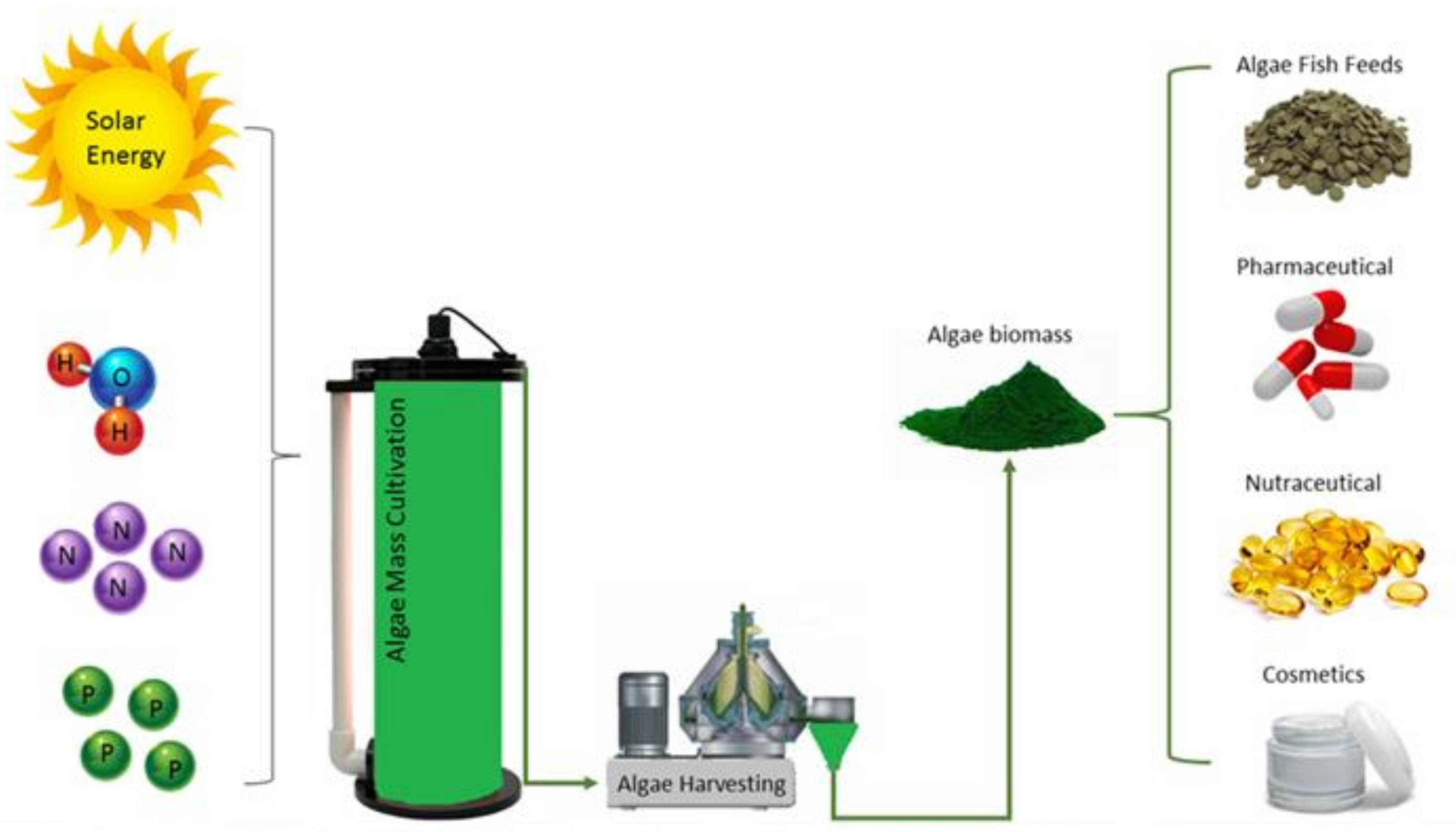
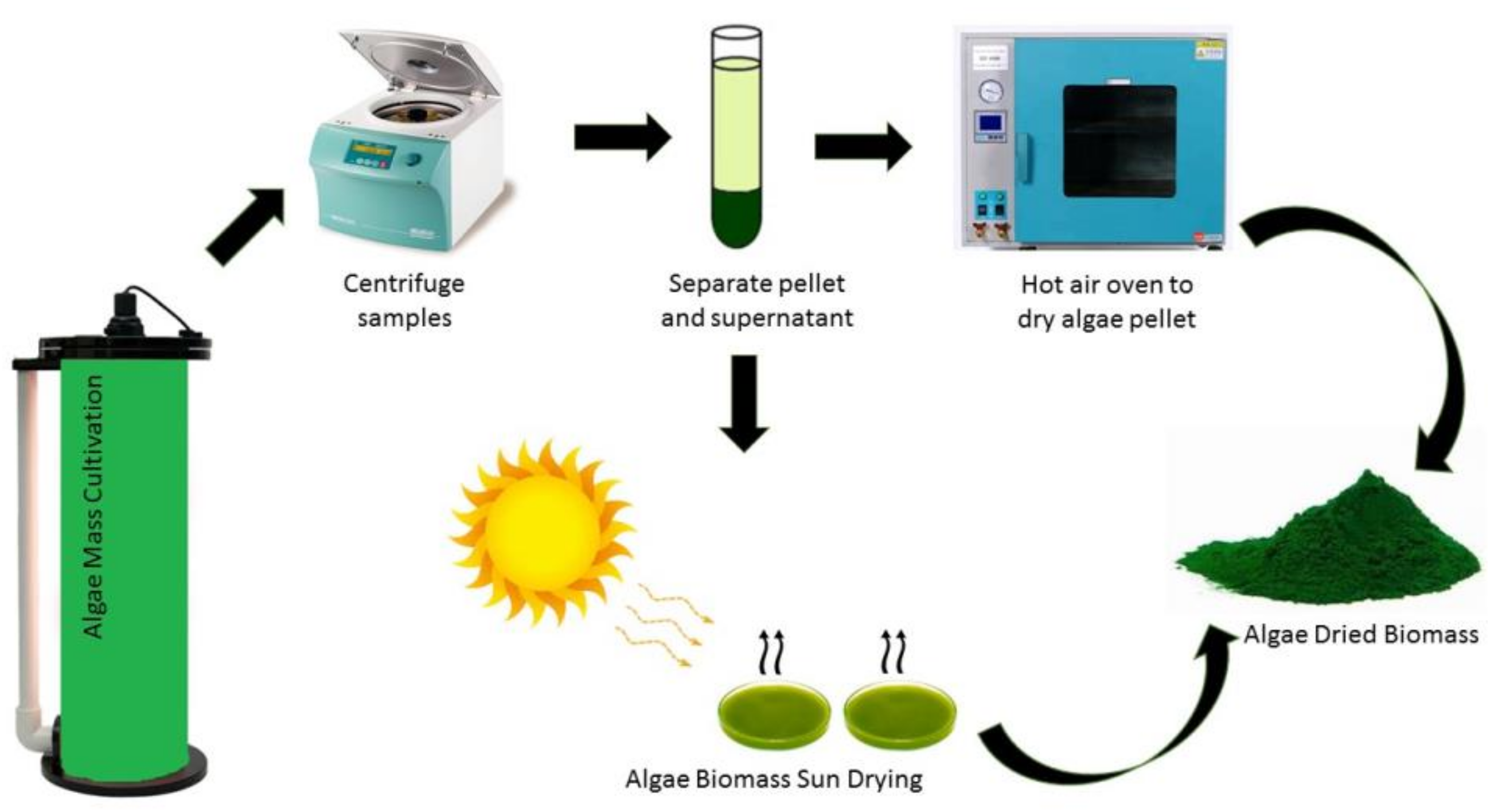
3.2. Biomass Quality
4. Microalgae Lipids and Fatty Acids Extraction Methods
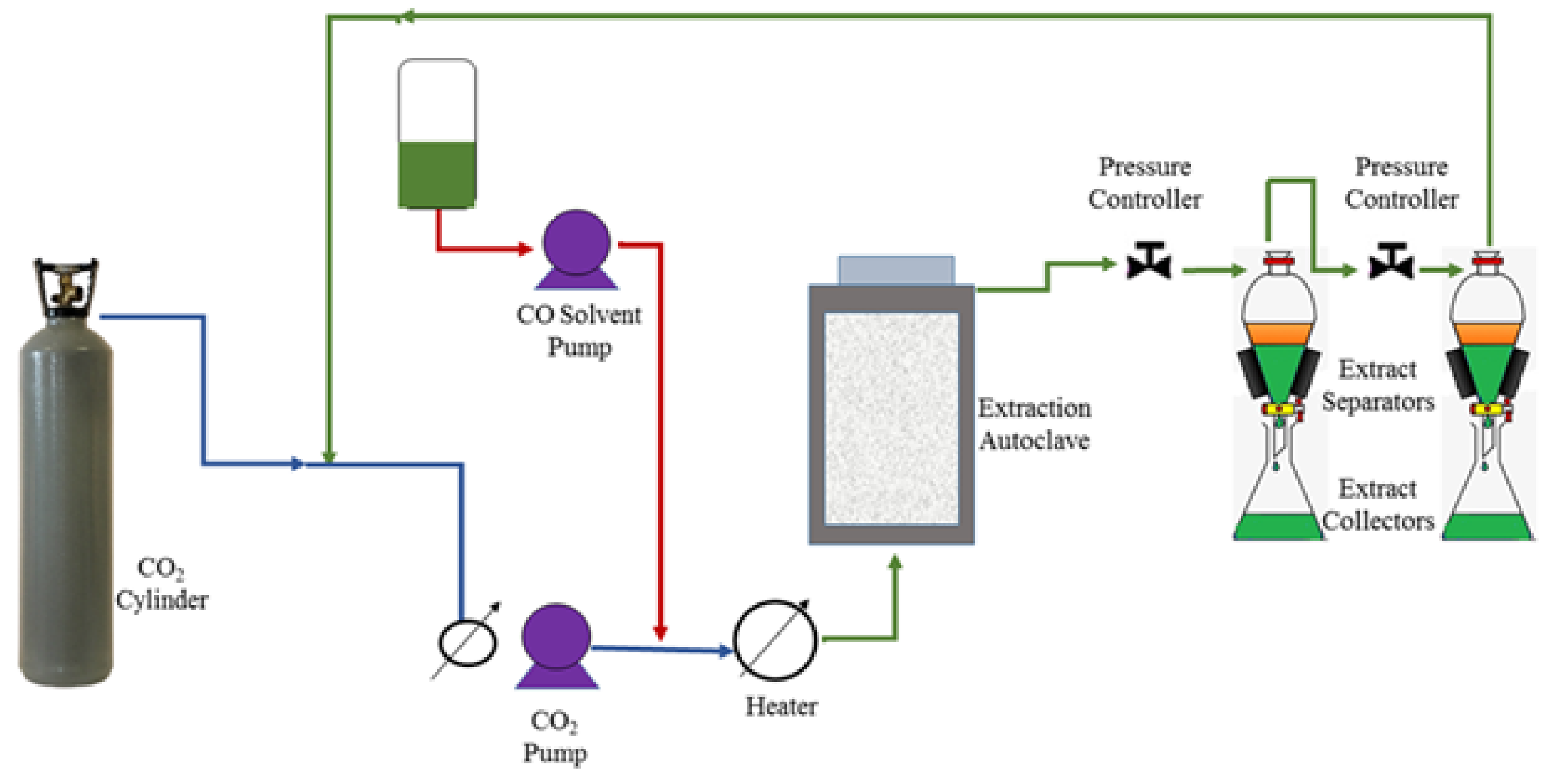
4.1. Lipids Extraction Methods
4.2. Fatty Acids Extraction Method
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sirakov, I.; Velichkova, K.; Stoyanova, S.; Staykov, Y. The importance of microalgae for aquaculture industry. Review. Int. J. Fish. Aquat. Stud. 2015, 2, 81–84. [Google Scholar]
- Maizatul, A.Y.; Mohamed, R.M.S.R.; Al-Gheethi, A.A.; Hashim, M.A. An overview of the utilisation of microalgae biomass derived from nutrient recycling of wetmarket wastewater and slaughterhouse wastewater. Int. Aquat. Res. 2017, 9, 177–193. [Google Scholar] [CrossRef]
- Zhu, S.; Huang, W.; Xu, J.; Wang, Z.; Xu, J.; Yuan, Z. Metabolic changes of starch and lipid triggered by nitrogen starvation in the microalga Chlorella zofingiensis. Bioresour. Technol. 2014, 152, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, J.; Qin, S.; Zeng, M.; Jiang, Y.; Hu, L.; Xiao, P.; Hao, W.; Hu, Z.; Lei, A.; et al. Growth and lipid accumulation by different nutrients in the microalga Chlamydomonas reinhardtii. Biotechnol. Biofuels 2008, 11, 40. [Google Scholar] [CrossRef]
- Yodsuwan, N.; Sawayama, S.; Sirisansaneeyakul, S. Effect of nitrogen concentration on growth, lipid production and fatty acid profiles of the marine diatom Phaeodactylum tricornutum. Agric. Nat. Resour. 2017, 51, 190–197. [Google Scholar] [CrossRef]
- Yang, F.F.; Xiang, W.; Li, T.; Long, L. Transcriptome analysis for phosphorus starvation-induced lipid accumulation in Scenedesmus sp. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Zarrinmehr, M.J.; Farhadian, O.; Heyrati, F.P.; Keramat, J.; Koutra, E.; Kornaros, M.; Daneshvar, E. Effect of nitrogen concentration on the growth rate and biochemical composition of the microalga, Isochrysis galbana. Egypt. J. Aquat. Res. 2019, 46, 1687–4285. [Google Scholar] [CrossRef]
- Zienkiewicz, A.; Zienkiewicz, K.; Poliner, E.; Pulman, J.A.; Du, Z.-Y.; Stefano, G.; Tsai, C.H.; Horn, P.; Feussner, I.; Farre, E.M.; et al. The microalga Nannochloropsis during transition from quiescence to qutotrophy in response to nitrogen availability. Plant Physiol. 2020, 182, 819–839. [Google Scholar] [CrossRef] [PubMed]
- Jais, N.M.; Mohamed, R.M.S.R.; Al-Gheethi, A.A.; Hashim, A. Dual role of phycoremediation of wet market wastewater for nutrients and heavy metals removal and microalgae biomass production. Clean. Technol. Environ. Policy. 2016, 19, 37–52. [Google Scholar] [CrossRef]
- Yaakob, M.A.; Mohamed, R.M.S.R.; Al-Gheethi, A.; Athirah, A.; Kassim, A.H.M. Optimising of Scenedesmus sp. biomass production in chicken slaughterhouse wastewater using response surface methodology and potential utilisation as fish feeds. Environ. Sci. Pollut. Res. 2019, 26, 12089–12108. [Google Scholar] [CrossRef]
- Kim, M.K.; Park, J.W.; Park, C.S.; Kim, S.J.; Jeune, K.H.; Chang, M.U.; Acreman, J. Enhanced production of Scenedesmus spp. (green microalgae) using a new medium containing fermented swine wastewater. Bioresour. Technol. 2007, 98, 2220–2228. [Google Scholar] [CrossRef]
- Abou-Shanab, R.A.I.; Ji, M.K.; Kim, H.C.; Paeng, K.J.; Jeon, B.H. Microalgal species growing on piggery wastewater as a valuable candidate for nutrient removal and biodiesel production. J. Environ. Manage. 2013, 115, 257–264. [Google Scholar] [CrossRef]
- Brown, M.R. Nutritional value of microalgae for aquaculture. In Advances in Aquaculture nutrition, Proceedings of the VI International Symposium on Nutrition Aquaculture, Cancun, Quintana Roo, Mexico 3–6 September 2002; Cruz-Suarez, L.E., Ricque-Marie, D., Tapia-Salazar, M., Gaxiola-Cortés, M.G., Simoes, N., Eds.; Universidad Autónoma De Nuevo León: Monterey, Mexico, 2002. [Google Scholar]
- Hemaiswarya, S.; Raja, R.; Kumar, R.R.; Ganesan, V.; Anbazhagan, C. Microalgae: A sustainable feed source for aquaculture. World J. Microbiol. Biotechnol. 2011, 27, 1737–1746. [Google Scholar] [CrossRef]
- Spolaore, P.; Joannis-Cassan, C.; Duran, E.; Isambert, A. Commercial applications of microalgae. J. Biosci. Bioeng. 2006, 101, 87–96. [Google Scholar] [CrossRef]
- Hakalin, N.L.S.; Paz, A.P.; Aranda, D.A.G.; Moraes, L.M.P. Enhancement of cell growth and lipid content of a freshwater microalga Scenedesmus sp. by optimizing nitrogen, phosphorus and vitamin concentrations for biodiesel production. Nat. Sci. 2014, 6, 1044–1054. [Google Scholar] [CrossRef]
- Baharuddin, N.N.D.E.; Azizi, N.S.; Sohif, H.N.; Karim, W.A.B.; Al-Obaidi, J.R.; Basiran, M.N. Marine microalgae flocculation using plant: The case of Nannochloropsis oculata and Moringa oleifera. Pak. J. Bot. 2016, 48, 831–840. [Google Scholar]
- Xin, L.; Hong-ying, H.; Ke, G.; Ying-xue, S. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494–5500. [Google Scholar] [CrossRef]
- Singh, J.; Gu, S. Commercialization potential of microalgae for biofuels production. Renew. Sustain. Energy Rev. 2010, 14, 2586–2610. [Google Scholar] [CrossRef]
- Zhang, Q.; Hong, Y. Comparison of growth and lipid accumulation properties of two oleaginous microalgae under different nutrient conditions. Front. Environ. Sci. Eng. 2014, 8, 703–709. [Google Scholar] [CrossRef]
- Arkronrat, W.; Deemark, P.; Oniam, V. Growth performance and proximate composition of mixed cultures of marine microalgae (Nannochloropsis sp. and Tetraselmis sp.) with monocultures. Songklanakarin J. Sci. Technol. 2016, 38, 1–5. [Google Scholar]
- Munoz, R.; Guieysse, B. Algal–bacterial processes for the treatment of hazardous contaminants: A review. Water Res. 2006, 40, 2799–2815. [Google Scholar] [CrossRef]
- Mansour, M.P.; Frampton, D.M.F.; Nichols, P.D.; Volkman, J.K.; Blackburn, S.I. Lipid and fatty acid yield of nine stationary-phase microalgae: Applications and unusual C24–C28 polyunsaturated fatty acids. J. Appl. Phycol. 2005, 17, 287–300. [Google Scholar] [CrossRef]
- Vooren, G.V.; Grand, F.L.; Legrand, J.; Cuiné, S.; Peltier, G.; Pruvost, J. Investigation of fatty acids accumulation in Nannochloropsis oculata for biodiesel application. Bioresour. Technol. 2012, 124, 421–432. [Google Scholar] [CrossRef]
- Xu, N.; Zhang, X.; Fan, X.; Han, L.; Zeng, C. Effects of nitrogen source and concentration on growth rate and fatty acid composition of Ellipsoidion sp. (Eustigmatophyta). J. Appl. Phycol. 2001, 13, 463–469. [Google Scholar] [CrossRef]
- Procházková, G.; Brányiková, I.; Zachleder, V.; Brányik, T. Effect of nutrient supply status on biomass composition of eukaryotic green microalgae. J. Appl. Phycol. 2013, 26, 1359–1377. [Google Scholar] [CrossRef]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef]
- Chu, F.F.; Chu, P.N.; Cai, P.J.; Li, W.W.; Lam, P.K.S.; Zeng, R.J. Phosphorus plays an important role in enhancing biodiesel productivity of Chlorella vulgaris under nitrogen deficiency. Bioresour. Technol. 2013, 134, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Pancha, I.; Chokshi, K.; George, B.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Nitrogen stress triggered biochemical and morphological changes in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2014, 156, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Han, D.; Gerken, H.G.; Li, Y.; Sommerfeld, M.; Hu, Q.; Xu, J. Molecular mechanisms for photosynthetic carbon partitioning into storage neutral lipids in Nannochloropsis oceanica under nitrogen-depletion conditions. Algal Res. 2015, 7, 66–77. [Google Scholar] [CrossRef]
- Paes, C.R.P.S.; Faria, G.R.; Tinoco, N.A.B.; Castro, D.J.F.A.; Barbarino, E.; Lourenço, S.O. Growth, nutrient uptake and chemical composition of Chlorella sp. and Nannochloropsis oculata under nitrogen starvation. Lat. Am. J. Aquat. Res. 2016, 44, 275–292. [Google Scholar] [CrossRef]
- Patel, A.; Pruthi, V.; Pruthi, P.A. Synchronized nutrient stress conditions trigger the diversion of CDP-DG pathway of phospholipids synthesis towards de novo TAG synthesis in oleaginous yeast escalating biodiesel production. Energy 2017, 139, 962–974. [Google Scholar] [CrossRef]
- An, M.; Gao, L.; Zhao, W.; Chen, W.; Li, M. Effects of nitrogen forms and supply mode on lipid production of microalga Scenedesmus obliquus. Energies 2020, 13, 697. [Google Scholar] [CrossRef]
- Feng, P.; Deng, Z.; Fan, L.; Hu, Z. Lipid accumulation and growth characteristics of Chlorella zofingiensis under different nitrate and phosphate concentrations. J. Biosci. Bioeng. 2012, 114, 405–410. [Google Scholar] [CrossRef]
- Breuer, G.; Lamers., P.P.; Martens, D.E.; Draaisma, R.B.; Wijffels, R.H. The impact of nitrogen starvation on the dynamics of triacylglycerol accumulation in nine microalgae strains. Bioresour. Technol. 2012, 124, 217–226. [Google Scholar] [CrossRef]
- Anand, J.; Arumugam, M. Enhanced lipid accumulation and biomass yield of Scenedesmus quadricauda under nitrogen starved condition. Bioresour. Technol. 2015, 188, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Guldhe, A.; Kumari, S.; Rawat, I.; Bux, F. Investigation of combined effect of nitrogen, phosphorus and iron on lipid productivity of microalgae Ankistrodesmus falcatus KJ671624 using response surface methodology. Biochem. Eng J. 2015, 94, 22–29. [Google Scholar] [CrossRef]
- Cointet, E.; Wielgosz-Collin, G.; Bougaran, G.; Rabesaotra, V.; Goncalves, O.; Meleder, V. Effects of light and nitrogen availability on photosynthetic efficiency and fatty acid content of three original benthic diatom strains. PLoS ONE 2019, 14, e0224701. [Google Scholar] [CrossRef] [PubMed]
- Latsos, C.; Houcke, J.V.; Timmermans, K.R. The effect of nitrogen starvation on biomass yield and biochemical constituents of Rhodomonas sp. Front. Mar. Sci. 2020, 7, 563333. [Google Scholar] [CrossRef]
- Delgado, R.T.; Guarieiro, M.D.S.; Cassini, S.T.; Terreros, H.M.; Fernandes, V.D.O. Effect of nitrogen limitation on growth, biochemical composition and cell ultrastructure of the microalga Picocystis Salinarum. Preprint 2020. [Google Scholar] [CrossRef]
- Savvidou, M.G.; Boli, E.; Logothetis, D.; Lymperopoulou, T.; Ferraro, A.; Louli, V.; Mamma, D.; Kekos, D.; Magoulas, K.; Kolisis, F.N. A study on the effect of macro- and micro-nutrients on Nannochloropsis oceanica growth, fatty acid composition and magnetic harvesting efficiency. Plants 2020, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Ota, S.; Yoshihara, M.; Yamazaki, T.; Takeshita, T.; Hirata, A.; Konomi, M.; Oshima, K.; Hattori, M.; Bisova, K.; Zachleder, V.; et al. Deciphering the relationship among phosphate dynamics, electron-dense body and lipid accumulation in the green alga Parachlorella kessleri. Sci. Rep. 2016, 6, 25731. [Google Scholar] [CrossRef] [PubMed]
- Hannon, M.; Gimpel, J.; Tran, M.; Rasala, B.; Mayfield, S. Biofuels from algae: Challenges and potential. Biofuels 2010, 1, 763–784. [Google Scholar] [CrossRef] [PubMed]
- Atiku, A.; Mohamed, R.M.S.R.; Al-Gheethi, A.A.; Wurochekke, A.A.; Kassim, A.H. Harvesting microalgae biomass from the phycoremediation process of greywater. Environ. Sci. Pollut. Res. 2016, 23, 24624–24641. [Google Scholar] [CrossRef]
- Solovchenko, A.; Gorelova, O.; Karpova, O.; Selyakh, I.; Semenova, L.; Chivkunova, O.; Baulina, O.; Vinogradova, E.; Pugacheva, T.; Scherbakov, P.; et al. Phosphorus feast and famine in Cyanobacteria: Is luxury uptake of the nutrient just a consequence of acclimation to its shortage. Cells 2020, 9, 1933. [Google Scholar] [CrossRef]
- Goldberg, I.K.; Cohen, Z. The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water eustigmatophyte Monodus subterraneus. Phytochemistry 2006, 67, 696–701. [Google Scholar] [CrossRef]
- Roopnarain, A.; Gray, V.M.; Sym, S.D. Phosphorus limitation and starvation effects on cell growth and lipid accumulation in Isochrysis galbana U4 for biodiesel production. Bioresour. Technol. 2014, 156, 408–411. [Google Scholar] [CrossRef]
- Jais, N.M.; Mohamed, R.M.S.R.; Apandi, W.A.W.M.; Peralta., H.M.M. Removal of nutrients and selected heavy metals in wet market wastewater by using microalgae Scenedesmus sp. Appl. Mech. Mater. 2015, 1210–1214. [Google Scholar] [CrossRef]
- Satpati, G.G.; Gorain, P.C.; Pal, R. Efficacy of EDTA and phosphorous on biomass yield and total lipid accumulation in two green microalgae with special emphasis on neutral lipid detection by flow cytometry. Adv. Biol. 2016, 2016, 12. [Google Scholar] [CrossRef]
- Lorenzen, J.; Igl, N.; Tippelt, M.; Stege, A.; Qoura, F.; Sohling, U.; Brück, T. Extraction of microalgae derived lipids with supercritical carbon dioxide in an industrial relevant pilot plant. Bioprocess Biosyst. Eng. 2017, 40, 911–918. [Google Scholar] [CrossRef]
- Anto, S.; Pugazhendhi, A.; Mathimani, T. Lipid enhancement through nutrient starvation in Chlorella sp. and its fatty acid profiling for appropriate bioenergy feedstock. Biocatal. Agric. Biotechnol. 2019, 20, 101179. [Google Scholar] [CrossRef]
- Mulbry, W.; Kondrad, S.; Pizarro, C.; Kebede-Westhead, E. Treatment of dairy manure effluent using freshwater algae: Algal productivity and recovery of manure nutrients using pilot-scale algal turf scrubbers. Bioresour. Technol. 2008, 99, 8137–8142. [Google Scholar] [CrossRef] [PubMed]
- Solovchenko, A.; Verschoor, A.M.; Jablonowski, N.D.; Nedbal, L. Phosphorus from wastewater to crops: An alternative path involving microalgae. Biotechnol. Adv. 2016, 34, 550–564. [Google Scholar] [CrossRef]
- Salem, O.M.A.; El-Ardy, O.; Abd El-Rahman, M.A. Effect of Nitrogen and Phosphorus Concentrations in Growth Medium and Salt Stress on Growth, Lipid Content, and Biodiesel Producing Ability of Microalgae; Helwan University: Helwan, Egypt, 2013; pp. 17–18. [Google Scholar]
- Liang, K.; Zhang, Q.; Gu, M.; Cong, W. Effect of phosphorus on lipid accumulation in freshwater microalga Chlorella sp. J. Appl. Phycol. 2013, 25, 311–318. [Google Scholar] [CrossRef]
- Qari, H.A.; Oves, M. Fatty acid synthesis by Chlamydomonas reinhardtii in phosphorus limitation. J. Bioenerg. Biomembr. 2020, 52, 27–38. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, M.; Ding, W.; Liu, J. Novel insights into phosphorus deprivation-boosted lipid synthesis in the marine alga Nannochloropsis oceanica without compromising biomass production. J. Agric. Food Chem. 2020, 68, 11488–11502. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Limits to growth. In Wastewater Treatment with Algae, 1st ed.; Wong, Y.S., Tam, N.F.Y., Eds.; Springer: Berlin/Heidelberg, Germany, 1997; pp. 203–226. [Google Scholar]
- Chen, J.; Wang, Y.; Benemann, J.R.; Zhang, X.; Hu, H.; Qin, S. Microalgal industry in China: Challenges and prospects. J. Appl. Phycol. 2016, 28, 715–725. [Google Scholar] [CrossRef]
- Costa, J.A.V.; de Morais, M.G. An open pond system for microalgal cultivation. In Biofuels from Algae, 1st ed.; Pandey, A., Lee, D.J., Chisti, Y., Soccol, C.R., Eds.; Elsevier: Saint Luis, PHL, USA, 2013; pp. 1–22. [Google Scholar]
- Sun, Z.; Liu, J.; Zhou, Z.G. Algae for biofuels: An emerging feedstock. In Handbook of Biofuels Production, 2nd ed.; Luque, R., Lin, C.S.K., Wilson, K., Clark, J., Eds.; Woodhead publishing: Cambridge, UK, 2016; pp. 673–698. [Google Scholar]
- Khatoon, N.; Pal, R. Microalgae in biotechnological application: A commercial approach. In Plant Biology and Biotechnology; Bahadur, B., Venkat Rajam, M., Sahijram, L., Krishnmurhty, K.V., Eds.; Springer: New Delhi, India, 2015; Volume 2, pp. 22–47. [Google Scholar] [CrossRef]
- Arumugam, M.; Agarwal, A.; Arya, M.C.; Ahmed, Z. Influence of nitrogen sources on biomass productivity of microalgae Scenedesmus bijugatus. Bioresour. Technol. 2013, 131, 246–249. [Google Scholar] [CrossRef]
- Kumar, R.R.; Rao, P.H.; Arumugam, M. Lipid extraction methods from microalgae: A comprehensive review. Front. Energy Res. 2015, 2, 61. [Google Scholar]
- James, I.; Yoon, L.W.; Chow, Y.H. Effect of phosphorus-limited nutrients on growth and glucose production from microalgae. In Proceedings of the International Engineering Research Conference, Subang Jaya, Malaysia, 3–4 July 2019; AIP Publishing: College Park, MD, USA, 2019; Volume 2137, p. 020005. [Google Scholar]
- Ji, C.F.; Yu, X.J.; Chen, Z.A.; Xue, S.; Legrand, J.; Zhang, W. Effects of nutrient deprivation on biochemical compositions and photo-hydrogen production of Tetraselmis subcordiformis. Int. J. Hydrogen Energy. 2011, 36, 5817–5821. [Google Scholar] [CrossRef]
- Lin, H.; Shavezipur, M.; Yousef, A.; Maleky, F. Prediction of growth of Pseudomonas fluorescens in milk during storage under fluctuating temperature. J. Dairy Sci. 2016, 99, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Přibyl, P.; Cepák, V.; Zachleder, V. Oil overproduction by means of microalgae. In Algal Biorefineries; Bajpal, R., Prokop, A., Zappi, M., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 1, pp. 241–273. [Google Scholar]
- FAO. Code of Conduct for Responsible Fisheries; Food and Agricultural Organization of the United Nations: Rome, Italy, 1996. [Google Scholar]
- Zhu, L.D.; Li, Z.H.; Hiltunen, E. Strategies for lipid production improvement in microalgae as a biodiesel feedstock. Biomed. Res. Int. 2016, 8792548. [Google Scholar] [CrossRef]
- Zachleder, V.; Brányiková, I. Starch overproduction by means of algae. In Algal Biorefineries; Bajpai, R., Prokop, A., Zappi, M., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 1, pp. 217–240. [Google Scholar]
- Vítová, M.; Bisová, K.; Kawano, S.; Zachleder, V. Accumulation of energy reserves in algae: From cell cycles to biotechnological applications. Biotechnol.Adv. 2015, 33, 1204–1218. [Google Scholar] [CrossRef]
- Fernandes, B.; Teixeira, J.; Dragone, G.; Vicente, A.A.; Kawano, S.; Bišová, K.; Přibyl, P.; Zachleder, V.; Vítová, M. Relationship between starch and lipid accumulation induced by nutrient depletion and replenishment in the microalga Parachlorella kessleri. Bioresour. Technol. 2013, 144, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.W.M.; Lee, Y.K. The dilemma for lipid productivity in green microalgae: Importance of substrate provision in improving oil yield without sacrificing growth. Biotechnol. Biofuels. 2016, 9, 255. [Google Scholar] [CrossRef] [PubMed]
- Yoo, G.; Park, W.K.; Kim, C.W.; Choi, Y.E.; Yang, J.W. Direct lipid extraction from wet Chlamydomonas reinhardtii biomass using osmotic shock. Bioresour. Technol. 2012, 123, 717–722. [Google Scholar] [CrossRef] [PubMed]
- King, J.W. Supercritical fluid technology for lipid extraction, fractionation and reactions. In Lipid Biotechnology, 1st ed.; Kuo, T.M., Gardner, H., Eds.; Marcel Dekker Inc.: New York, NY, USA, 2002; pp. 663–687. [Google Scholar]
- Santana, A.; Jesus, S.; Larrayoz, A.; Filho, R.M. Supercritical carbon dioxide extraction of algal lipids for the biodiesel production. Procedia Eng. 2012, 42, 1755–1761. [Google Scholar] [CrossRef]
- Patil, P.D.; Dandamudi, K.P.R.; Wang, J.; Deng, Q.; Deng, S. Extraction of bio-oils from algae with supercritical carbon dioxide and co-solvents. J. Supercrit. Fluids. 2018, 135, 60–68. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- González-González, L.M.; Astals, S.; Pratt, S.; Jensen, P.D.; Schenk, P.M. Impact of osmotic shock pre-treatment on microalgae lipid extraction and subsequent methane production. Bioresour. Technol. Rep. 2019, 7, 100214. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Maneein, S.; Harvey, P.J. A brief review of anaerobic digestion of algae for bioenergy. Energies 2019, 12, 1166. [Google Scholar] [CrossRef]
- Klassen, V.; Blifernez-Klassen, O.; Wibberg, D.; Winkler, A.; Kalinowski, J.; Posten, C.; Kruse, O. Highly efficient methane generation from untreated microalgae biomass. Biotechnol. Biofuels 2017, 10, 186. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhang, Y.; Rosenberg, J.N.; Betenbaugh, M.J.; Wang, F. Optimization of one-step in situ transesterification method for accurate quantification of EPA in Nannochloropsis gaditana. Appl. Sci. 2016, 6, 343. [Google Scholar] [CrossRef]
- Rafael, L.; Carol, S.K.L.; Karen, W.; James, C. Handbook of Biofuels Production, 2nd ed.; Woodhead publishing: Cambridge, UK, 2016; pp. 1–770. [Google Scholar]
- Praveenkumar, R.; Shameera, K.; Mahalakshmi, G.; Akbarsha, M.A.; Thajuddin, N. Influence of nutrient deprivations on lipid accumulation in a dominant indigenous microalga Chlorella sp., BUM11008: Evaluation for biodiesel production. Biomass Bioenerg. 2012, 37, 60–66. [Google Scholar]
- Prommuak, C.; Pavasant, P.; Quitain, A.T.; Goto, M.; Shotipruk, A. Microalgal lipid extraction and evaluation of single-step biodiesel production. Eng. J. 2012, 16, 157–166. [Google Scholar] [CrossRef]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids. MIDI; Technical Note #101: Newark, DE, USA, 1990. [Google Scholar]
- Glucksman, A.M.; Skipper, H.D.; Brigmon, R.L.; Santo Domingo, J.W. Use of the MIDI-FAME technique to characterize groundwater communities. J. Appl. Microbiol. 2000, 88, 711–719. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
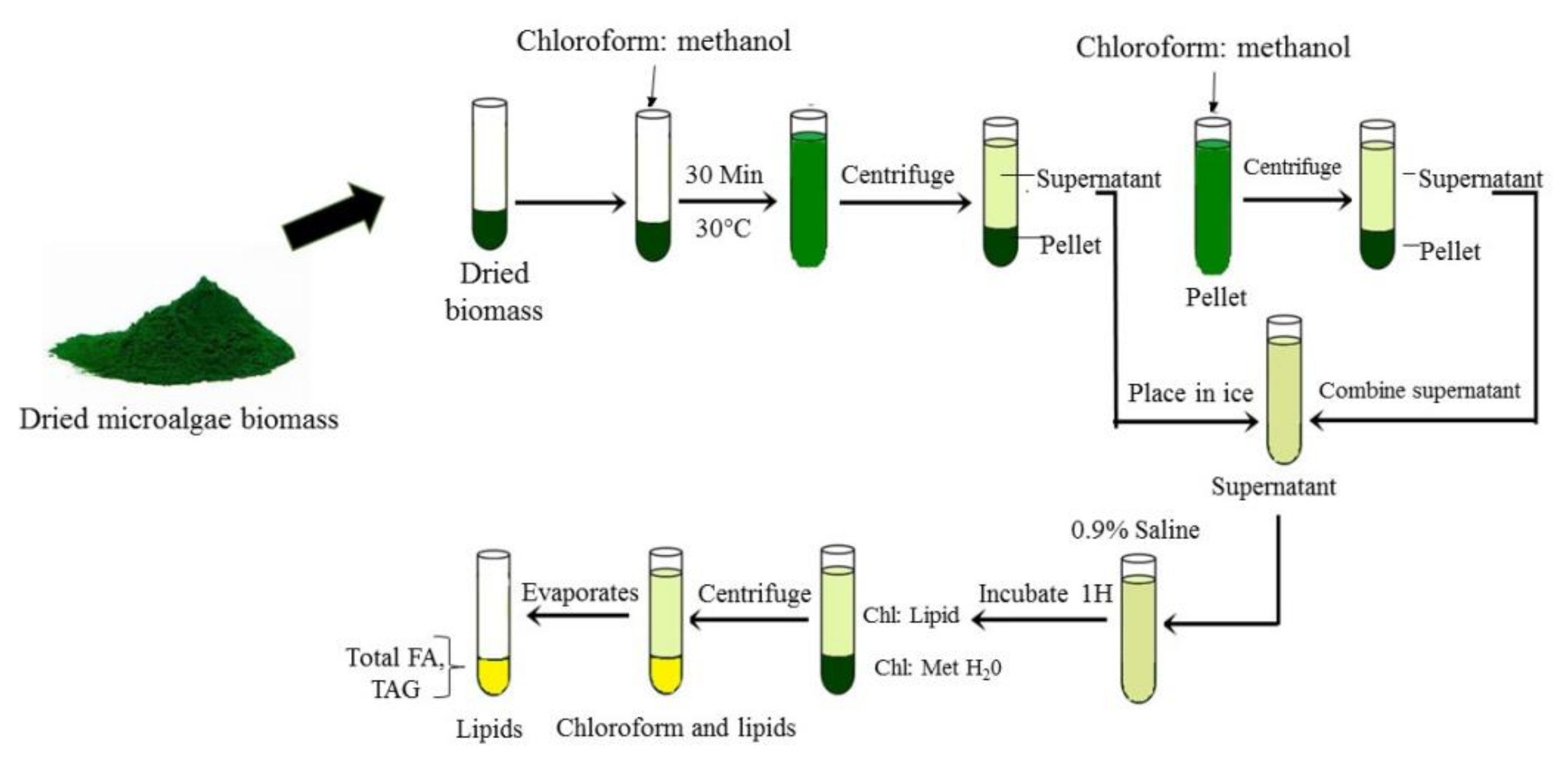{kind=link}
{kind=link}
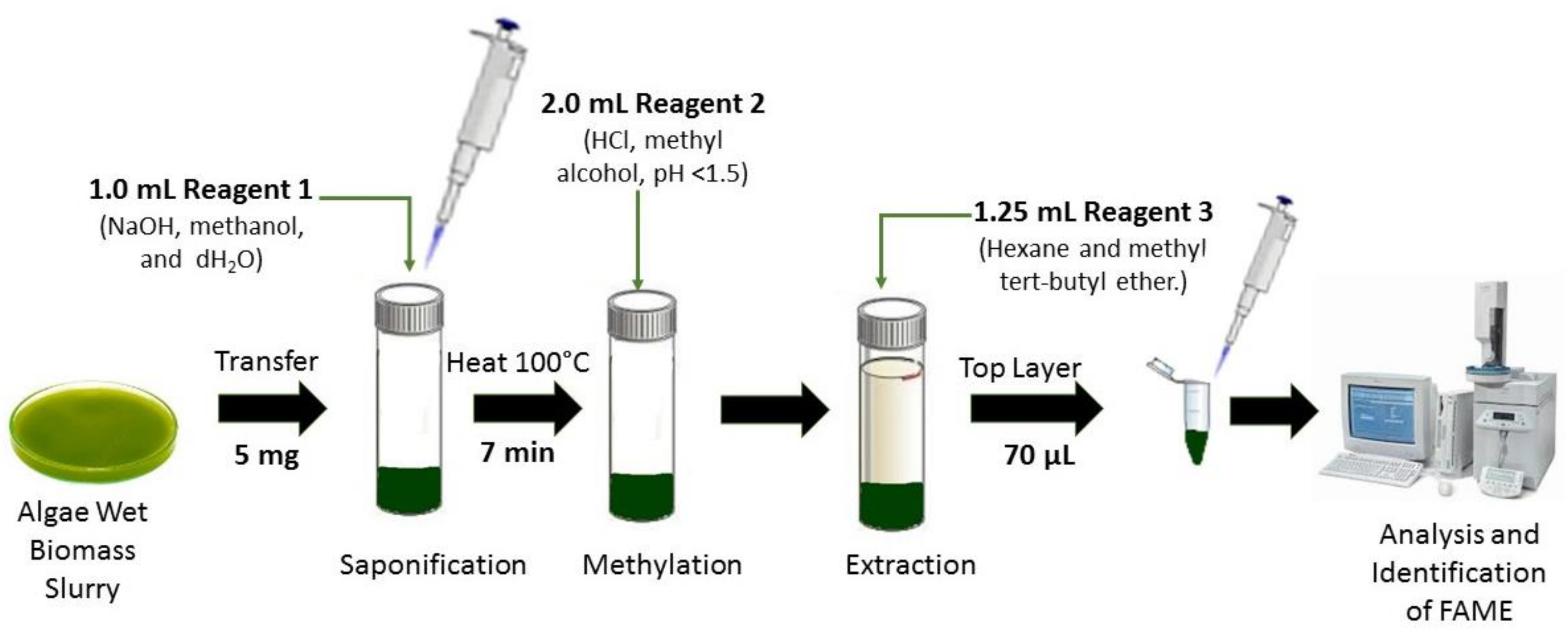{kind=link}
| Microalgae Types | Experiment | Outcomes Growth and Biochemical Composition | Reference |
|---|---|---|---|
| Nannochloropsis sp. | Transferred from nitrogen-sufficient to -deficient condition | Lipid content increased from 32% to 60% | [27] |
| Chlorella vulgaris | Lipid production under nitrogen starvation conditions | Lipid productivity was 58.39 mg/L/day. | [28] |
| Chlorella zofingiensis | 10 days cultivation in two conditions: nitrogen starvation and nitrogen repletion (0 g/L and 1.1 g/L, respectively) | Rapid growth in nitrogen-sufficient culture medium, whereas growth inhibition under nitrogen starvation condition | [3] |
| Scenedesmus sp. | Effects of nitrogen limitation and starvation on morphological and biochemical changes | Nitrate starvation generates 27% lipid accumulation | [29] |
| Nannochloropsis oceanica | 14 days under nitrogen-deplete and nitrogen-replete conditions | Regulation of metabolic pathways along with acetyl CoA synthesis and lipid degradation for TAG synthesis | [30] |
| Chlorella sp. and Nannochloropsis oculata | 13 days in Conway nutrient batch cultures without nitrogen in the 7th day of growth | 15.3 ±1.0% lipid in Chlorella sp., 33.7 ± 2.8% lipid in N. oculata under nitrogen limitation | [31] |
| Phaeodactylum tricornutum | Lipid accumulation in nitrogen-deficient medium | High lipid content (53.04 ± 3.26%) | [5] |
| C. reinhardtii | Effect of different nutrients on standard TAP, TAP nitrogen deficiency (T-N), TAP without nitrogen and phosphorus (T-N-P), TAP nitrogen deficiency with additional phosphorus 1 M(K)PO4 (T-N+P) on growth and lipid accumulation | Lipid contents in cells increased as a result of nitrogen-deficient conditions; the highest lipid content was 104.7% in T-N-P condition and the lowest lipid values were 49.9% in T-N condition | [32] |
| I. galbana | Effect of different nitrogen concentrations(0, 36, 72, 144 and 288 mg/L) on the growth rate and biochemical composition | Growth decreased in nitrogen-deficient conditions; in contrast, carbohydrates and fatty acids showed the highest value, 47% and 75%, respectively | [7] |
| Scenedesmus obliquus | Various nitrogen concentrations’ effect on growth and lipid production | Lipid production increased with increasing nitrogen concentration; the highest cell density (1.7 × 107 cells/mL) and lipid production (242.4 mg/L) | [33] |
| Microalgae Strain | Initial Nitrogen Available | Final Nitrogen Starvation | Cultivation of Algae (Days) | Biomass Yield | Lipid Content | Fatty Acid | Higher Fatty Acids | Reference |
|---|---|---|---|---|---|---|---|---|
| S. obliquus | 0 mM | -- | 14 | 719 mg/L/d | 35% | 44.7 ± 1.7% | C18:1 | [35] |
| C. zofingiensis | 0 mg/L | -- | 10 | 0.7 g/L | 24.5% | 12.10 × 10−7 µg 10.49 × 10−7 µg 6.25 × 10−7 µg | C16:0, C18:1, C18:2 | [3] |
| Scenedesmus quadricauda | 0 mg/L | -- | 30 | 265 × 104 cells/mL | 226 mg/L | 15.16%, 25.08%, 15.78% | C18:0, C18:3 C20:0 | [36] |
| Ankistrodesmus falcatus | 750 mg/L | -- | 14 | 1740 mg/L | 59.6% | 24.68 ± 0.13%, 19.2 ± 0.66%, 16.64 ± 0.33%, 11.77 ± 0.73% | C16:0, C18:1, C18:2, C18:3 | [37] |
| Phaeodactylum tricornutum | 0 mg/L | -- | 21 | 68.57 ± 7.57 mg/L | 53.04 ± 3.26% | 17.73 ± 8.40%, 13.07 ± 6.03%, 1.81 ± 1.25% | C16:0, C16:1, C20:5 | [5] |
| Entomoneis paludosa | 83.56 μM | 0.14 μM | 35 | 2.00 ± 0.11 cells/mL | 20.68 ± 2.62% | 36.7 ± 6.9%, 13.5 ± 2.0%, 20.1 ± 3.2% | C16:0, C18:0, C18:1 | [38] |
| Rhodomonas sp. | 1.5 g/L | 113 mg/L | 8 | 2.5 × 106 cells/mL | 30.3% | 6.1%, 5.5%, 3.7% | C16:0, C18:1, C18:3 | [39] |
| Nannochloropsis oceanica | 0 g/L | NR | 3 | 20 × 106 cells/mL | 58% | 48%, 40%, 5% | C16:0, C16:1, C18:1 | [8] |
| Picocystis salinarum | 9.375 mg/L | -- | 20 | 0.7 g/L | 33.87% | 72.13 µg/g, 923.95 µg/g, 19.87 µg/g | C14:0, C16:0, C18:2 | [40] |
| Microalgae Types | Experiment | Outcomes | Reference |
|---|---|---|---|
| Eustigmatophyte sp. | Phosphorus limitation condition | Triacylglycerols, TAG content increased from 12.9% to 15.1% in absence of phosphorus condition | [46] |
| C. zofingiensis | Affects phosphate on lipid accumulation as a biodiesel feedstock | Lipid content obtained from phosphate-deficient was 44.7%, relatively higher than cells grown in full medium (33.5%). | [34] |
| C. vulgaris | Biodiesel production under phosphorus-sufficient conditions | Lipid yield of C. vulgaris for biodiesel production was 58.39 mg/L/day in phosphate-sufficient conditions | [28] |
| I. galbana | Varied levels of phosphorus in F/2 medium for biodiesel production | Phosphorus starvation stimulated lipid accumulation up to 50% w/w | [47] |
| Scenedesmus sp. | Phosphorus-rich medium from public market wastewater treated for 8 days | Total phosphorus in medium reduced about 82.17% | [48] |
| Chlorella ellipsoidea and Chlorococcum infusionum | Phosphorous starvation culture for 30 days | Total lipid content of Chlorella (41.8 ± 1.9% at 16 days of incubation period) and Chlorococcum (31.3 ± 1.0% at 18 days of incubation period) | [49] |
| Scenedesmus obtusiusculus | Cultivated in BG-11 medium under non-limiting conditions | 6.4% w/w lipid yield after SCCO2 extraction | [50] |
| Scenedesmus sp. | Resuspension in modified SE medium with 2 mg/L NaH2PO4·2H2O for 9 days | The highest lipid production (350 mg/L) and lipid content (41.0%) were obtained by addition of 2 mg/L NaH2PO4·2H2O every 2 days | [6] |
| Chlorella sp. | Lipid enhancement through nutrient starvation | Under phosphate starvation, the lipid content was 13.9%. | [51] |
| Microalgae Strain | Initial Phosphorus Available | Final Phosphorus Starvation | Cultivation of Algae (Days) | Biomass Yield | Lipid Content | Fatty Acid | Higher Fatty Acids | Ref |
|---|---|---|---|---|---|---|---|---|
| Phormidium sp. | 0.02 g/L | -- | 15 | 0.130 g | 27.9% | 34.667%, 11.266%, 27.912%, | C16:0, C18:0 C20:2, | [54] |
| Chlorella sp. | 32 μM | -- | 22 | 2 g/L | 23.60% | 35.48%, 33.90% | C16:0, C18:0 | [55] |
| C. ellipsoidea, C. infusionum | 1.5 g/L | -- | 30 | 1.56 ± 0.06 g/L, 2.17 ± 0.12 g/L | 41.8 ± 1.9%, 31.3 ± 1.0% | 21.62 ± 0.94%, 30.32 ± 2.68% | C18:1, | [49] |
| Rhodosporidium kratochvilovae | 0.05 g/L | -- | 10 | 12.65 ± 0.12 g/L | 51.7 ± 0.81% | 10.36 ± 0.59% 66.79 ± 0.21% 10.48 ± 0.43% | C18:0, C18:1, C18:2 | [32] |
| C. reinhardtii | 0 μg/mg | -- | 7 | 1.1 g/L | 105.00 μg/mg | 35.86 ± 0.25 μg/mg, 22.12 ± 0.12 μg/mg, 22.84 ± 0.08 μg/mg | C16:0, C18:2, C18:3 | [4] |
| Scenedesmus sp. | 2mg/L | -- | 9 | 0.6 g/L | 41.0% | NR | C16:0, C18:1 | [6] |
| C. reinhardtii | 0.4 mg/L | -- | 14 | 0.08 mg/L | NR | 56% 28.8% | C16:0, C18:3 | [56] |
| Nannochloropsis oceanica | 0 g/L | -- | 6 | 2.5 mg/L | 337 mg/g | 37.3 ± 0.1%, 30.8 ± 0.3%, 10.7 ± 0.8%. | C16:0, C16:1, C20:5 | [57] |
| N. oceanica | 0 g/L | -- | 14 | 0.29 g/L | 23.7% | 13.93%, 26.31% | C18:1, C16:1. | [41] |
| Types of Extraction Method | Advantages/Efficiency | Key Methodology | Drawbacks |
|---|---|---|---|
| Mechanical Approach | |||
| Expeller press | Simple and effective crushing method | High mechanical pressure to crush the cells and squeeze oil out from microalgae biomass | Pressure decreased lipid recovery, increased heat; expensive and time-consuming |
| Ultrasound-assisted extraction | Economical and eco-friendly, completed in a short time, high reproducibility, does not require the addition of beads or chemicals | Cavitation produces heat shock waves to disrupt microalgae cells with less thermal denaturation of biomolecules | Expensive, leads to free radicals and detrimental to the oil quality |
| Bead beating | Moderate efficiency | Disruption of cells using high-speed spinning beads | Moderate efficiency |
| Microwaves irradiation method | Short reaction time, low costs, efficient extraction and quick oil recovery | Extraction and transesterification of the oils into biodiesel by intracellular heating, which disrupts the cells | High maintenance cost on a large scale. |
| Electroporation | High efficiency | Altered cell membranes and cell walls to improved lipid extraction | High initial and maintenance costs |
| Chemical Approach | |||
| Supercritical fluid extraction method (SEF) | Rapid, safe and economical method and does not require dewatering of biomass | Using high pressure to extract lipids from cells | High cost and environmental and safety issues |
| Bligh and Dyer method | Rapid and easy processing of large number of samples | Extracting total lipids from microalgae using ratio 2:1 methanol: chloroform | Less sensitive |
| Pressurized solvent extraction | High efficiency | Using solvent and pressurized nitrogen for extraction | High cost, fire, health and environmental hazards |
| Folch method | Rapid and easy processing of large number of samples | Combination of different solvents such as chloroform–methanol in the ratio of 2:1 by volume | Less sensitive |
| Soxhlet extraction method | Easy processing of a large number of samples and moderate efficiency | Normally used to quantify high quality lipids such as fatty acids and triglycerides using hexane as a solvent | Environmental and health risks |
| Accelerated solvent extraction (ASE) | Short process time and can recover solvent for reuse | Using heat or pressure to achieve better lipid recovery | Less efficient in a larger scale |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaakob, M.A.; Mohamed, R.M.S.R.; Al-Gheethi, A.; Aswathnarayana Gokare, R.; Ambati, R.R. Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview. Cells 2021, 10, 393. https://doi.org/10.3390/cells10020393
Yaakob MA, Mohamed RMSR, Al-Gheethi A, Aswathnarayana Gokare R, Ambati RR. Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview. Cells. 2021; 10(2):393. https://doi.org/10.3390/cells10020393
Chicago/Turabian StyleYaakob, Maizatul Azrina, Radin Maya Saphira Radin Mohamed, Adel Al-Gheethi, Ravishankar Aswathnarayana Gokare, and Ranga Rao Ambati. 2021. "Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview" Cells 10, no. 2: 393. https://doi.org/10.3390/cells10020393
APA StyleYaakob, M. A., Mohamed, R. M. S. R., Al-Gheethi, A., Aswathnarayana Gokare, R., & Ambati, R. R. (2021). Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview. Cells, 10(2), 393. https://doi.org/10.3390/cells10020393