
Dunking into the Lipid Bilayer: How Direct Membrane Binding of Nucleoporins Can Contribute to Nuclear Pore Complex Structure and Assembly



Abstract
1. Introduction
2. Modes of NPC Assembly
3. Lipid Composition of the Nuclear Envelope
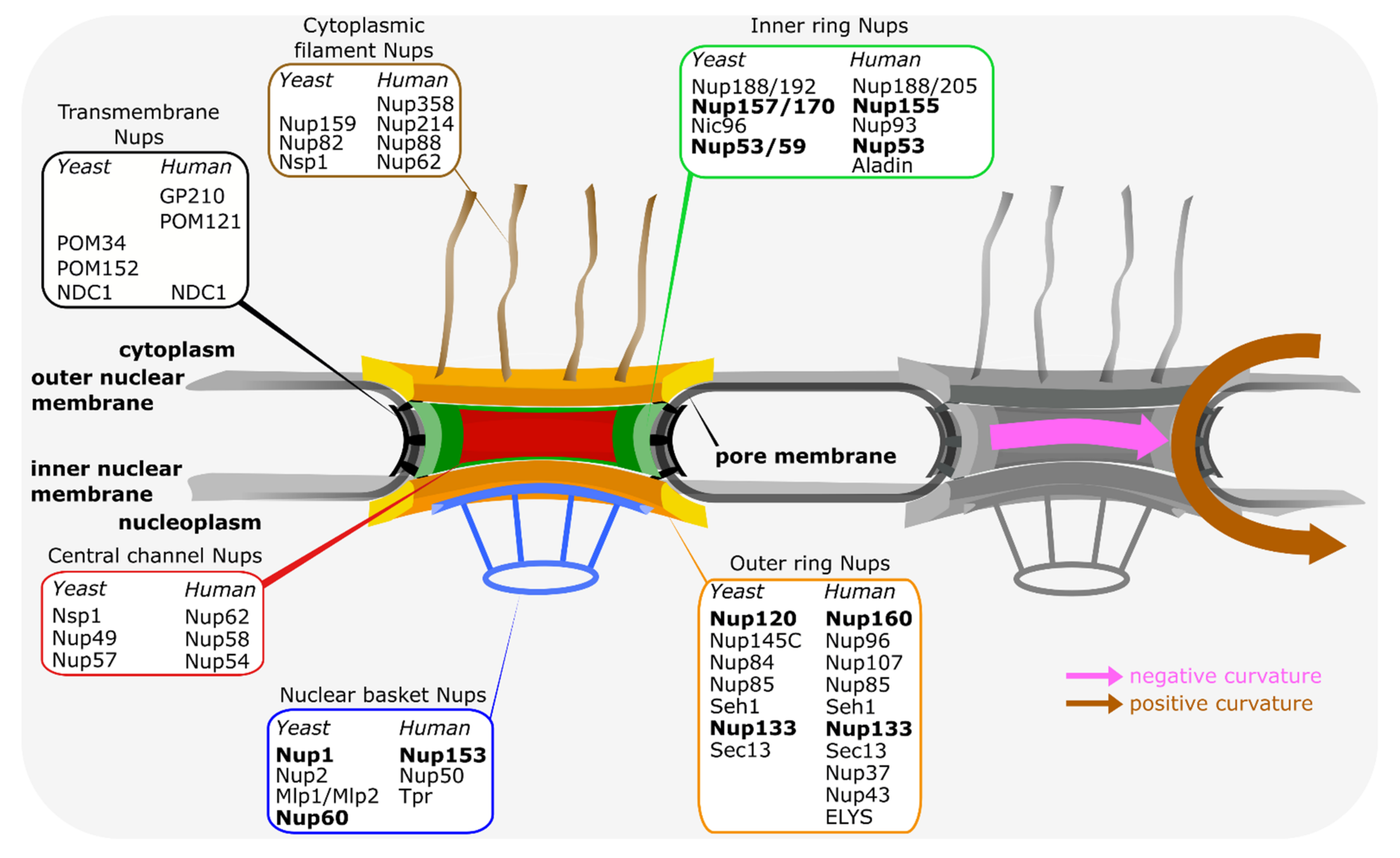
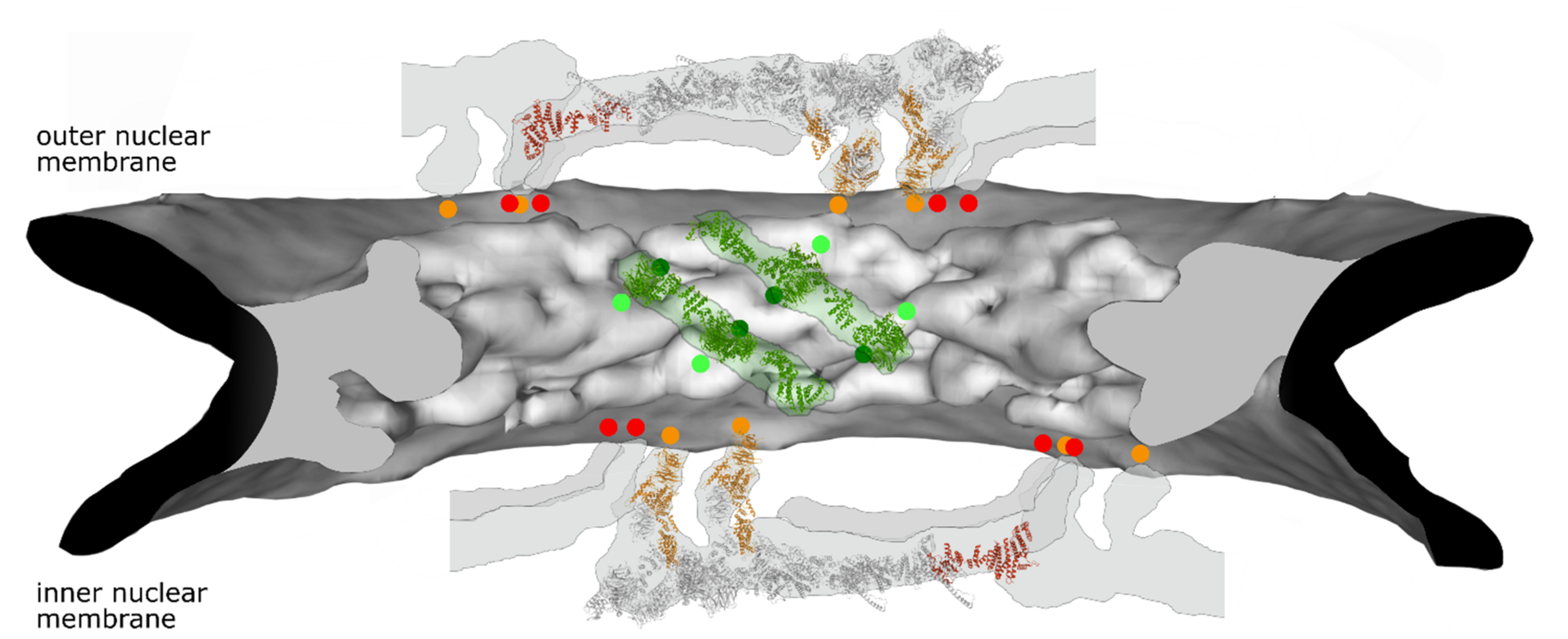
4. Nucleoporin Interactions with the Nuclear Pore Membrane
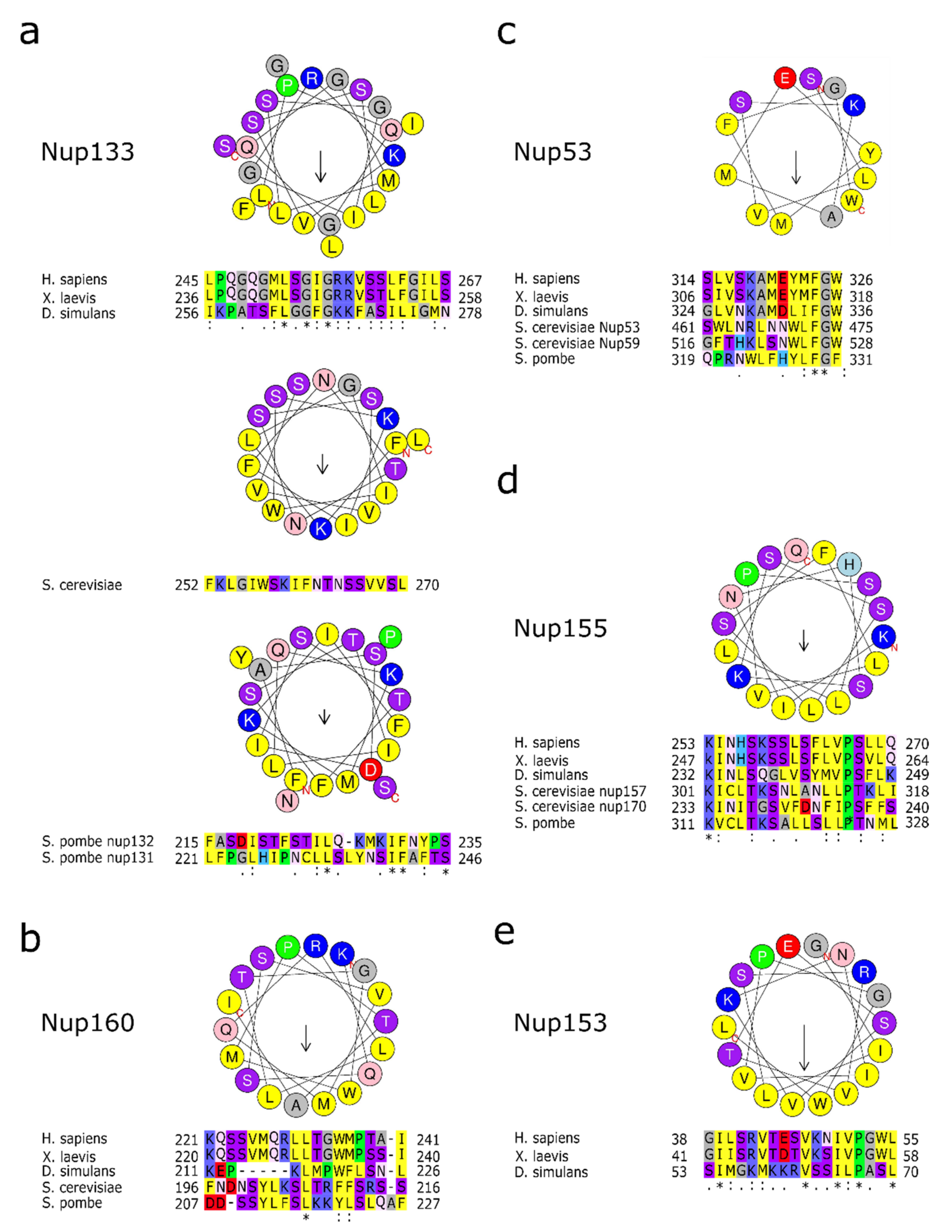
4.1. Y-Complexes Interact with the Pore Membrane Via ALPS Motifs
4.2. Nup53 and Nup59
4.3. Nup155 and Its Orthologues Nup157 and Nup170
4.4. Nup153 and Relatives, Membrane Interacting Nucleoporins of the Nuclear Basket
4.5. Nup50 and Nup2, Nuclear Basket Nucleoporins as Phosphoinositide Binders
5. Membrane Interactions for Curvature Stabilization or Induction?
6. Assays for Membrane Binding
7. Conclusions and Outlook
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Paci, G.; Caria, J.; Lemke, E.A. Cargo transport through the nuclear pore complex at a glance. J. Cell Sci. 2021, 134, jcs247874. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Hetzer, M.W. Functional interactions between nucleoporins and chromatin. Curr. Opin. Cell Biol. 2011, 23, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Garcia, P.; Capelson, M. The nuclear pore complex and the genome: Organizing and regulatory principles. Curr. Opin. Genet. Dev. 2021, 67, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Hampoelz, B.; Andres-Pons, A.; Kastritis, P.; Beck, M. Structure and Assembly of the Nuclear Pore Complex. Annu. Rev. Biophys. 2019, 48, 515–536. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.H.; Hoelz, A. The Structure of the Nuclear Pore Complex(An Update). Annu. Rev. Biochem. 2019, 88, 725–783. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.; Mosalaganti, S.; Kosinski, J. From the resolution revolution to evolution: Structural insights into the evolutionary relationships between vesicle coats and the nuclear pore. Curr. Opin. Struct. Biol. 2018, 52, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Brohawn, S.G.; Leksa, N.C.; Spear, E.D.; Rajashankar, K.R.; Schwartz, T.U. Structural evidence for common ancestry of the nuclear pore complex and vesicle coats. Science 2008, 322, 1369–1373. [Google Scholar] [CrossRef]
- Devos, D.; Dokudovskaya, S.; Alber, F.; Williams, R.; Chait, B.T.; Sali, A.; Rout, M.P. Components of coated vesicles and nuclear pore complexes share a common molecular architecture. PLoS Biol. 2004, 2, e380. [Google Scholar] [CrossRef]
- Mans, B.J.; Anantharaman, V.; Aravind, L.; Koonin, E.V. Comparative genomics, evolution and origins of the nuclear envelope and nuclear pore complex. Cell Cycle 2004, 3, 1612–1637. [Google Scholar] [CrossRef]
- DeGrasse, J.A.; DuBois, K.N.; Devos, D.; Siegel, T.N.; Sali, A.; Field, M.C.; Rout, M.P.; Chait, B.T. Evidence for a shared nuclear pore complex architecture that is conserved from the last common eukaryotic ancestor. Mol. Cell. Proteom. 2009, 8, 2119–2130. [Google Scholar] [CrossRef]
- Devos, D.; Dokudovskaya, S.; Williams, R.; Alber, F.; Eswar, N.; Chait, B.T.; Rout, M.P.; Sali, A. Simple fold composition and modular architecture of the nuclear pore complex. Proc. Natl. Acad. Sci. USA 2006, 103, 2172–2177. [Google Scholar] [CrossRef]
- Knockenhauer, K.E.; Schwartz, T.U. The Nuclear Pore Complex as a Flexible and Dynamic Gate. Cell 2016, 164, 1162–1171. [Google Scholar] [CrossRef]
- Makarov, A.A.; Padilla-Mejia, N.E.; Field, M.C. Evolution and diversification of the nuclear pore complex. Biochem. Soc. Trans. 2021, 49, 1601–1619. [Google Scholar] [CrossRef]
- Neumann, N.; Lundin, D.; Poole, A.M. Comparative genomic evidence for a complete nuclear pore complex in the last eukaryotic common ancestor. PLoS ONE 2010, 5, e13241. [Google Scholar] [CrossRef]
- Iwamoto, M.; Osakada, H.; Mori, C.; Fukuda, Y.; Nagao, K.; Obuse, C.; Hiraoka, Y.; Haraguchi, T. Compositionally distinct nuclear pore complexes of functionally distinct dimorphic nuclei in the ciliate Tetrahymena. J. Cell Sci. 2017, 130, 1822–1834. [Google Scholar] [CrossRef]
- Kutay, U.; Juhlen, R.; Antonin, W. Mitotic disassembly and reassembly of nuclear pore complexes. Trends Cell Biol. 2021, 31, 1019–1033. [Google Scholar] [CrossRef]
- Dultz, E.; Zanin, E.; Wurzenberger, C.; Braun, M.; Rabut, G.; Sironi, L.; Ellenberg, J. Systematic kinetic analysis of mitotic dis- and reassembly of the nuclear pore in living cells. J. Cell Biol. 2008, 180, 857–865. [Google Scholar] [CrossRef]
- Dultz, E.; Ellenberg, J. Live imaging of single nuclear pores reveals unique assembly kinetics and mechanism in interphase. J. Cell Biol. 2010, 191, 15–22. [Google Scholar] [CrossRef]
- Otsuka, S.; Ellenberg, J. Mechanisms of nuclear pore complex assembly - two different ways of building one molecular machine. FEBS Lett. 2018, 592, 475–488. [Google Scholar] [CrossRef]
- Weberruss, M.; Antonin, W. Perforating the nuclear boundary - how nuclear pore complexes assemble. J. Cell Sci. 2016, 129, 4439–4447. [Google Scholar] [CrossRef]
- Pawar, S.; Kutay, U. The Diverse Cellular Functions of Inner Nuclear Membrane Proteins. Cold Spring Harb. Perspect. Biol. 2021, 13, a040477. [Google Scholar] [CrossRef]
- Franke, W.W.; Deumling, B.; Ermen, B.; Jarasch, E.D.; Kleinig, H. Nuclear membranes from mammalian liver. I. Isolation procedure and general characterization. J. Cell Biol. 1970, 46, 379–395. [Google Scholar] [CrossRef]
- Khandwala, A.S.; Kasper, C.B. The fatty acid composition of individual phospholipids from rat liver nuclear membrane and nuclei. J. Biol. Chem. 1971, 246, 6242–6246. [Google Scholar] [CrossRef]
- van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Zinser, E.; Daum, G. Isolation and biochemical characterization of organelles from the yeast, Saccharomyces cerevisiae. Yeast 1995, 11, 493–536. [Google Scholar] [CrossRef]
- Bigay, J.; Antonny, B. Curvature, lipid packing, and electrostatics of membrane organelles: Defining cellular territories in determining specificity. Dev. Cell 2012, 23, 886–895. [Google Scholar] [CrossRef]
- Lutzmann, M.; Kunze, R.; Buerer, A.; Aebi, U.; Hurt, E. Modular self-assembly of a Y-shaped multiprotein complex from seven nucleoporins. EMBO J. 2002, 21, 387–397. [Google Scholar] [CrossRef]
- Belgareh, N.; Rabut, G.; Bai, S.W.; van Overbeek, M.; Beaudouin, J.; Daigle, N.; Zatsepina, O.V.; Pasteau, F.; Labas, V.; Fromont-Racine, M.; et al. An evolutionarily conserved NPC subcomplex, which redistributes in part to kinetochores in mammalian cells. J. Cell Biol. 2001, 154, 1147–1160. [Google Scholar] [CrossRef]
- Loiodice, I.; Alves, A.; Rabut, G.; Van Overbeek, M.; Ellenberg, J.; Sibarita, J.B.; Doye, V. The entire Nup107–160 complex, including three new members, is targeted as one entity to kinetochores in mitosis. Mol. Biol. Cell 2004, 15, 3333–3344. [Google Scholar] [CrossRef]
- Allegretti, M.; Zimmerli, C.E.; Rantos, V.; Wilfling, F.; Ronchi, P.; Fung, H.K.H.; Lee, C.W.; Hagen, W.; Turonova, B.; Karius, K.; et al. In-cell architecture of the nuclear pore and snapshots of its turnover. Nature 2020, 586, 796–800. [Google Scholar] [CrossRef]
- Kim, S.J.; Fernandez-Martinez, J.; Nudelman, I.; Shi, Y.; Zhang, W.; Raveh, B.; Herricks, T.; Slaughter, B.D.; Hogan, J.A.; Upla, P.; et al. Integrative structure and functional anatomy of a nuclear pore complex. Nature 2018, 555, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Bui, K.H.; von Appen, A.; Diguilio, A.L.; Ori, A.; Sparks, L.; Mackmull, M.T.; Bock, T.; Hagen, W.; Andres-Pons, A.; Glavy, J.S.; et al. Integrated structural analysis of the human nuclear pore complex scaffold. Cell 2013, 155, 1233–1243. [Google Scholar] [CrossRef] [PubMed]
- Mosalaganti, S.; Kosinski, J.; Albert, S.; Schaffer, M.; Strenkert, D.; Salome, P.A.; Merchant, S.S.; Plitzko, J.M.; Baumeister, W.; Engel, B.D.; et al. In situ architecture of the algal nuclear pore complex. Nat. Commun. 2018, 9, 2361. [Google Scholar] [CrossRef] [PubMed]
- Zimmerli, C.E.; Allegretti, M.; Rantos, V.; Goetz, S.K.; Obarska-Kosinska, A.; Zagoriy, I.; Halavatyi, A.; Hummer, G.; Mahamid, J.; Kosinski, J.; et al. Nuclear pores dilate and constrict in cellulo. Science 2021, 374, eabd9776. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, H.; Kojidani, T.; Yang, H.J.; Ohtsuki, C.; Osakada, H.; Matsuda, A.; Iwamoto, M.; Chikashige, Y.; Nagao, K.; Obuse, C.; et al. Asymmetrical localization of Nup107–160 subcomplex components within the nuclear pore complex in fission yeast. PLoS Genet. 2019, 15, e1008061. [Google Scholar] [CrossRef] [PubMed]
- Faini, M.; Beck, R.; Wieland, F.T.; Briggs, J.A. Vesicle coats: Structure, function, and general principles of assembly. Trends Cell Biol. 2013, 23, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.M.; Mansfeld, J.; Capitanio, J.; Kutay, U.; Wozniak, R.W. Pom121 links two essential subcomplexes of the nuclear pore complex core to the membrane. J. Cell Biol. 2010, 191, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Soderqvist, H.; Hallberg, E. The large C-terminal region of the integral pore membrane protein, POM121, is facing the nuclear pore complex. Eur. J. Cell Biol. 1994, 64, 186–191. [Google Scholar]
- Mosalaganti, S.; Obarska-Kosinska, A.; Siggel, M.; Turonova, B.; Zimmerli, C.E.; Buczak, K.; Schmidt, F.H.; Margiotta, E.; Mackmull, M.-T.; Hagen, W.; et al. Artificial intelligence reveals nuclear pore complexity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Schuller, A.P.; Wojtynek, M.; Mankus, D.; Tatli, M.; Kronenberg-Tenga, R.; Regmi, S.G.; Dip, P.V.; Lytton-Jean, A.K.R.; Brignole, E.J.; Dasso, M.; et al. The cellular environment shapes the nuclear pore complex architecture. Nature 2021, 598, 667–671. [Google Scholar] [CrossRef]
- von Appen, A.; Kosinski, J.; Sparks, L.; Ori, A.; DiGuilio, A.L.; Vollmer, B.; Mackmull, M.T.; Banterle, N.; Parca, L.; Kastritis, P.; et al. In situ structural analysis of the human nuclear pore complex. Nature 2015, 526, 140–143. [Google Scholar] [CrossRef]
- Drin, G.; Casella, J.F.; Gautier, R.; Boehmer, T.; Schwartz, T.U.; Antonny, B. A general amphipathic alpha-helical motif for sensing membrane curvature. Nat. Struct. Mol. Biol. 2007, 14, 138–146. [Google Scholar] [CrossRef]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific alpha-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef]
- Berke, I.C.; Boehmer, T.; Blobel, G.; Schwartz, T.U. Structural and functional analysis of Nup133 domains reveals modular building blocks of the nuclear pore complex. J. Cell Biol. 2004, 167, 591–597. [Google Scholar] [CrossRef]
- Kim, S.J.; Fernandez-Martinez, J.; Sampathkumar, P.; Martel, A.; Matsui, T.; Tsuruta, H.; Weiss, T.M.; Shi, Y.; Markina-Inarrairaegui, A.; Bonanno, J.B.; et al. Integrative structure-function mapping of the nucleoporin Nup133 suggests a conserved mechanism for membrane anchoring of the nuclear pore complex. Mol. Cell. Proteom. 2014, 13, 2911–2926. [Google Scholar] [CrossRef]
- Nordeen, S.A.; Turman, D.L.; Schwartz, T.U. Yeast Nup84-Nup133 complex structure details flexibility and reveals conservation of the membrane anchoring ALPS motif. Nat. Commun. 2020, 11, 6060. [Google Scholar] [CrossRef]
- Doucet, C.M.; Esmery, N.; de Saint-Jean, M.; Antonny, B. Membrane Curvature Sensing by Amphipathic Helices Is Modulated by the Surrounding Protein Backbone. PLoS ONE 2015, 10, e0137965. [Google Scholar] [CrossRef]
- Doucet, C.M.; Talamas, J.A.; Hetzer, M.W. Cell cycle-dependent differences in nuclear pore complex assembly in metazoa. Cell 2010, 141, 1030–1041. [Google Scholar] [CrossRef]
- Bilir, Ş.; Kojidani, T.; Mori, C.; Osakada, H.; Kobayashi, S.; Koujin, T.; Hiraoka, Y.; Haraguchi, T. Roles of Nup133, Nup153 and membrane fenestrations in assembly of the nuclear pore complex at the end of mitosis. Genes Cells 2019, 24, 338–353. [Google Scholar] [CrossRef]
- Lupu, F.; Alves, A.; Anderson, K.; Doye, V.; Lacy, E. Nuclear pore composition regulates neural stem/progenitor cell differentiation in the mouse embryo. Dev. Cell 2008, 14, 831–842. [Google Scholar] [CrossRef]
- Souquet, B.; Freed, E.; Berto, A.; Andric, V.; Auduge, N.; Reina-San-Martin, B.; Lacy, E.; Doye, V. Nup133 Is Required for Proper Nuclear Pore Basket Assembly and Dynamics in Embryonic Stem Cells. Cell Rep. 2018, 23, 2443–2454. [Google Scholar] [CrossRef] [PubMed]
- Doye, V.; Wepf, R.; Hurt, E.C. A novel nuclear pore protein Nup133p with distinct roles in poly(A)+ RNA transport and nuclear pore distribution. EMBO J. 1994, 13, 6062–6075. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.W.; Rouquette, J.; Umeda, M.; Faigle, W.; Loew, D.; Sazer, S.; Doye, V. The fission yeast Nup107–120 complex functionally interacts with the small GTPase Ran/Spi1 and is required for mRNA export, nuclear pore distribution, and proper cell division. Mol. Cell. Biol. 2004, 24, 6379–6392. [Google Scholar] [CrossRef] [PubMed]
- Osmani, A.H.; Davies, J.; Liu, H.L.; Nile, A.; Osmani, S.A. Systematic deletion and mitotic localization of the nuclear pore complex proteins of Aspergillus nidulans. Mol. Biol. Cell 2006, 17, 4946–4961. [Google Scholar] [CrossRef]
- Kanamori, N.; Madsen, L.H.; Radutoiu, S.; Frantescu, M.; Quistgaard, E.M.; Miwa, H.; Downie, J.A.; James, E.K.; Felle, H.H.; Haaning, L.L.; et al. A nucleoporin is required for induction of Ca2+ spiking in legume nodule development and essential for rhizobial and fungal symbiosis. Proc. Natl. Acad. Sci. USA 2006, 103, 359–364. [Google Scholar] [CrossRef]
- Galy, V.; Mattaj, I.W.; Askjaer, P. Caenorhabditis elegans nucleoporins Nup93 and Nup205 determine the limit of nuclear pore complex size exclusion in vivo. Mol. Biol. Cell 2003, 14, 5104–5115. [Google Scholar] [CrossRef]
- Fernandez-Martinez, J.; Phillips, J.; Sekedat, M.D.; Diaz-Avalos, R.; Velazquez-Muriel, J.; Franke, J.D.; Williams, R.; Stokes, D.L.; Chait, B.T.; Sali, A.; et al. Structure-function mapping of a heptameric module in the nuclear pore complex. J. Cell Biol. 2012, 196, 419–434. [Google Scholar] [CrossRef]
- Kosinski, J.; Mosalaganti, S.; von Appen, A.; Teimer, R.; DiGuilio, A.L.; Wan, W.; Bui, K.H.; Hagen, W.J.; Briggs, J.A.; Glavy, J.S.; et al. Molecular architecture of the inner ring scaffold of the human nuclear pore complex. Science 2016, 352, 363–365. [Google Scholar] [CrossRef]
- Vollmer, B.; Antonin, W. The diverse roles of the Nup93/Nic96 complex proteins - structural scaffolds of the nuclear pore complex with additional cellular functions. Biol. Chem. 2014, 395, 515–528. [Google Scholar] [CrossRef]
- Mansfeld, J.; Guttinger, S.; Hawryluk-Gara, L.A.; Pante, N.; Mall, M.; Galy, V.; Haselmann, U.; Muhlhausser, P.; Wozniak, R.W.; Mattaj, I.W.; et al. The conserved transmembrane nucleoporin NDC1 is required for nuclear pore complex assembly in vertebrate cells. Mol. Cell 2006, 22, 93–103. [Google Scholar] [CrossRef]
- Stavru, F.; Hulsmann, B.B.; Spang, A.; Hartmann, E.; Cordes, V.C.; Gorlich, D. NDC1: A crucial membrane-integral nucleoporin of metazoan nuclear pore complexes. J. Cell Biol. 2006, 173, 509–519. [Google Scholar] [CrossRef]
- Hawryluk-Gara, L.A.; Platani, M.; Santarella, R.; Wozniak, R.W.; Mattaj, I.W. Nup53 is required for nuclear envelope and nuclear pore complex assembly. Mol. Biol. Cell 2008, 19, 1753–1762. [Google Scholar] [CrossRef]
- Sachdev, R.; Sieverding, C.; Flotenmeyer, M.; Antonin, W. The C-terminal domain of Nup93 is essential for assembly of the structural backbone of nuclear pore complexes. Mol. Biol. Cell 2012, 23, 740–749. [Google Scholar] [CrossRef]
- Chug, H.; Trakhanov, S.; Hulsmann, B.B.; Pleiner, T.; Gorlich, D. Crystal structure of the metazoan Nup62*Nup58*Nup54 nucleoporin complex. Science 2015, 350, 106–110. [Google Scholar] [CrossRef]
- Grandi, P.; Dang, T.; Pane, N.; Shevchenko, A.; Mann, M.; Forbes, D.; Hurt, E. Nup93, a vertebrate homologue of yeast Nic96p, forms a complex with a novel 205-kDa protein and is required for correct nuclear pore assembly. Mol. Biol. Cell 1997, 8, 2017–2038. [Google Scholar] [CrossRef]
- Eisenhardt, N.; Redolfi, J.; Antonin, W. Interaction of Nup53 with Ndc1 and Nup155 is required for nuclear pore complex assembly. J. Cell Sci. 2014, 127, 908–921. [Google Scholar] [CrossRef]
- Hawryluk-Gara, L.A.; Shibuya, E.K.; Wozniak, R.W. Vertebrate Nup53 interacts with the nuclear lamina and is required for the assembly of a Nup93-containing complex. Mol. Biol. Cell 2005, 16, 2382–2394. [Google Scholar] [CrossRef]
- Yavuz, S.; Santarella-Mellwig, R.; Koch, B.; Jaedicke, A.; Mattaj, I.W.; Antonin, W. NLS-mediated NPC functions of the nucleoporin Pom121. FEBS Lett. 2010, 584, 3292–3298. [Google Scholar] [CrossRef]
- Marelli, M.; Aitchison, J.D.; Wozniak, R.W. Specific binding of the karyopherin Kap121p to a subunit of the nuclear pore complex containing Nup53p, Nup59p, and Nup170p. J. Cell Biol. 1998, 143, 1813–1830. [Google Scholar] [CrossRef]
- Onischenko, E.; Stanton, L.H.; Madrid, A.S.; Kieselbach, T.; Weis, K. Role of the Ndc1 interaction network in yeast nuclear pore complex assembly and maintenance. J. Cell Biol. 2009, 185, 475–491. [Google Scholar] [CrossRef]
- Stuwe, T.; Bley, C.J.; Thierbach, K.; Petrovic, S.; Schilbach, S.; Mayo, D.J.; Perriches, T.; Rundlet, E.J.; Jeon, Y.E.; Collins, L.N.; et al. Architecture of the fungal nuclear pore inner ring complex. Science 2015, 350, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Uetz, P.; Giot, L.; Cagney, G.; Mansfield, T.A.; Judson, R.S.; Knight, J.R.; Lockshon, D.; Narayan, V.; Srinivasan, M.; Pochart, P.; et al. A comprehensive analysis of protein-protein interactions in Saccharomyces cerevisiae. Nature 2000, 403, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Lusk, C.P.; Makhnevych, T.; Marelli, M.; Aitchison, J.D.; Wozniak, R.W. Karyopherins in nuclear pore biogenesis: A role for Kap121p in the assembly of Nup53p into nuclear pore complexes. J. Cell Biol. 2002, 159, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Alber, F.; Dokudovskaya, S.; Veenhoff, L.M.; Zhang, W.; Kipper, J.; Devos, D.; Suprapto, A.; Karni-Schmidt, O.; Williams, R.; Chait, B.T.; et al. Determining the architectures of macromolecular assemblies. Nature 2007, 450, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Fahrenkrog, B.; Hubner, W.; Mandinova, A.; Pante, N.; Keller, W.; Aebi, U. The yeast nucleoporin Nup53p specifically interacts with Nic96p and is directly involved in nuclear protein import. Mol. Biol. Cell 2000, 11, 3885–3896. [Google Scholar] [CrossRef][Green Version]
- Marelli, M.; Lusk, C.P.; Chan, H.; Aitchison, J.D.; Wozniak, R.W. A link between the synthesis of nucleoporins and the biogenesis of the nuclear envelope. J. Cell Biol. 2001, 153, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Rexach, M.F. Discovering novel interactions at the nuclear pore complex using bead halo: A rapid method for detecting molecular interactions of high and low affinity at equilibrium. Mol. Cell. Proteom. 2008, 7, 121–131. [Google Scholar] [CrossRef]
- Vollmer, B.; Schooley, A.; Sachdev, R.; Eisenhardt, N.; Schneider, A.M.; Sieverding, C.; Madlung, J.; Gerken, U.; Macek, B.; Antonin, W. Dimerization and direct membrane interaction of Nup53 contribute to nuclear pore complex assembly. EMBO J. 2012, 31, 4072–4084. [Google Scholar] [CrossRef]
- Rodenas, E.; Klerkx, E.P.; Ayuso, C.; Audhya, A.; Askjaer, P. Early embryonic requirement for nucleoporin Nup35/NPP-19 in nuclear assembly. Dev. Biol. 2009, 327, 399–409. [Google Scholar] [CrossRef]
- Handa, N.; Kukimoto-Niino, M.; Akasaka, R.; Kishishita, S.; Murayama, K.; Terada, T.; Inoue, M.; Kigawa, T.; Kose, S.; Imamoto, N.; et al. The crystal structure of mouse Nup35 reveals atypical RNP motifs and novel homodimerization of the RRM domain. J. Mol. Biol. 2006, 363, 114–124. [Google Scholar] [CrossRef]
- Linder, M.I.; Kohler, M.; Boersema, P.; Weberruss, M.; Wandke, C.; Marino, J.; Ashiono, C.; Picotti, P.; Antonin, W.; Kutay, U. Mitotic Disassembly of Nuclear Pore Complexes Involves CDK1- and PLK1-Mediated Phosphorylation of Key Interconnecting Nucleoporins. Dev. Cell 2017, 43, 141–156.e7. [Google Scholar] [CrossRef]
- Whittle, J.R.; Schwartz, T.U. Architectural nucleoporins Nup157/170 and Nup133 are structurally related and descend from a second ancestral element. J. Biol. Chem. 2009, 284, 28442–28452. [Google Scholar] [CrossRef]
- De Magistris, P.; Tatarek-Nossol, M.; Dewor, M.; Antonin, W. A self-inhibitory interaction within Nup155 and membrane binding are required for nuclear pore complex formation. J. Cell Sci. 2018, 131, jcs208538. [Google Scholar] [CrossRef]
- Lin, D.H.; Stuwe, T.; Schilbach, S.; Rundlet, E.J.; Perriches, T.; Mobbs, G.; Fan, Y.; Thierbach, K.; Huber, F.M.; Collins, L.N.; et al. Architecture of the symmetric core of the nuclear pore. Science 2016, 352, aaf1015. [Google Scholar] [CrossRef]
- Aksenova, V.; Smith, A.; Lee, H.; Bhat, P.; Esnault, C.; Chen, S.; Iben, J.; Kaufhold, R.; Yau, K.C.; Echeverria, C.; et al. Nucleoporin TPR is an integral component of the TREX-2 mRNA export pathway. Nat. Commun. 2020, 11, 4577. [Google Scholar] [CrossRef]
- Bastos, R.; Lin, A.; Enarson, M.; Burke, B. Targeting and function in mRNA export of nuclear pore complex protein Nup153. J. Cell Biol. 1996, 134, 1141–1156. [Google Scholar] [CrossRef]
- Cordes, V.C.; Reidenbach, S.; Kohler, A.; Stuurman, N.; van Driel, R.; Franke, W.W. Intranuclear filaments containing a nuclear pore complex protein. J. Cell Biol. 1993, 123, 1333–1344. [Google Scholar] [CrossRef]
- Hase, M.E.; Cordes, V.C. Direct interaction with nup153 mediates binding of Tpr to the periphery of the nuclear pore complex. Mol. Biol. Cell 2003, 14, 1923–1940. [Google Scholar] [CrossRef]
- Smitherman, M.; Lee, K.; Swanger, J.; Kapur, R.; Clurman, B.E. Characterization and targeted disruption of murine Nup50, a p27(Kip1)-interacting component of the nuclear pore complex. Mol. Cell. Biol. 2000, 20, 5631–5642. [Google Scholar] [CrossRef]
- Vasu, S.; Shah, S.; Orjalo, A.; Park, M.; Fischer, W.H.; Forbes, D.J. Novel vertebrate nucleoporins Nup133 and Nup160 play a role in mRNA export. J. Cell Biol. 2001, 155, 339–354. [Google Scholar] [CrossRef]
- Vollmer, B.; Lorenz, M.; Moreno-Andres, D.; Bodenhofer, M.; De Magistris, P.; Astrinidis, S.A.; Schooley, A.; Flotenmeyer, M.; Leptihn, S.; Antonin, W. Nup153 Recruits the Nup107–160 Complex to the Inner Nuclear Membrane for Interphasic Nuclear Pore Complex Assembly. Dev. Cell 2015, 33, 717–728. [Google Scholar] [CrossRef]
- Meszaros, N.; Cibulka, J.; Mendiburo, M.J.; Romanauska, A.; Schneider, M.; Kohler, A. Nuclear pore basket proteins are tethered to the nuclear envelope and can regulate membrane curvature. Dev. Cell 2015, 33, 285–298. [Google Scholar] [CrossRef]
- Souquet, B.; Doye, V. Bending or Building: Multifaceted Functions of Amphipathic Helices in Basket Nucleoporins. Dev. Cell 2015, 33, 626–628. [Google Scholar] [CrossRef]
- Moore, M.S. Npap60: A new player in nuclear protein import. Trends Cell Biol. 2003, 13, 61–64. [Google Scholar] [CrossRef]
- Swaminathan, S.; Melchior, F. Nucleocytoplasmic transport: More than the usual suspects. Dev. Cell 2002, 3, 304–306. [Google Scholar] [CrossRef]
- Vonkova, I.; Saliba, A.E.; Deghou, S.; Anand, K.; Ceschia, S.; Doerks, T.; Galih, A.; Kugler, K.G.; Maeda, K.; Rybin, V.; et al. Lipid Cooperativity as a General Membrane-Recruitment Principle for PH Domains. Cell Rep. 2015, 12, 1519–1530. [Google Scholar] [CrossRef]
- Lewis, A.E.; Sommer, L.; Arntzen, M.; Strahm, Y.; Morrice, N.A.; Divecha, N.; D’Santos, C.S. Identification of nuclear phosphatidylinositol 4,5-bisphosphate-interacting proteins by neomycin extraction. Mol. Cell. Proteom. 2011, 10, S1–S15. [Google Scholar] [CrossRef]
- Castano, E.; Yildirim, S.; Fáberová, V.; Krausová, A.; Uličná, L.; Paprčková, D.; Sztacho, M.; Hozák, P. Nuclear Phosphoinositides-Versatile Regulators of Genome Functions. Cells 2019, 8, 649. [Google Scholar] [CrossRef]
- Rabut, G.; Doye, V.; Ellenberg, J. Mapping the dynamic organization of the nuclear pore complex inside single living cells. Nat. Cell. Biol. 2004, 6, 1114–1121. [Google Scholar] [CrossRef]
- Buchwalter, A.L.; Liang, Y.; Hetzer, M.W. Nup50 is required for cell differentiation and exhibits transcription-dependent dynamics. Mol. Biol. Cell 2014, 25, 2472–2484. [Google Scholar] [CrossRef]
- Holzer, G.; De Magistris, P.; Gramminger, C.; Sachdev, R.; Magalska, A.; Schooley, A.; Scheufen, A.; Lennartz, B.; Tatarek-Nossol, M.; Lue, H.; et al. The nucleoporin Nup50 activates the Ran guanine nucleotide exchange factor RCC1 to promote NPC assembly at the end of mitosis. EMBO J. 2021, 40, e108788. [Google Scholar] [CrossRef] [PubMed]
- Antonin, W.; Ellenberg, J.; Dultz, E. Nuclear pore complex assembly through the cell cycle: Regulation and membrane organization. FEBS Lett. 2008, 582, 2004–2016. [Google Scholar] [CrossRef] [PubMed]
- Antonin, W.; Mattaj, I.W. Nuclear pore complexes: Round the bend? Nat. Cell. Biol. 2005, 7, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Shemesh, T.; Kasthuri, N.; Klemm, R.W.; Schalek, R.; Hayworth, K.J.; Hand, A.R.; Yankova, M.; Huber, G.; Lichtman, J.W.; et al. Stacked endoplasmic reticulum sheets are connected by helicoidal membrane motifs. Cell 2013, 154, 285–296. [Google Scholar] [CrossRef]
- McMahon, H.T.; Gallop, J.L. Membrane curvature and mechanisms of dynamic cell membrane remodelling. Nature 2005, 438, 590–596. [Google Scholar] [CrossRef]
- Ford, M.G.; Mills, I.G.; Peter, B.J.; Vallis, Y.; Praefcke, G.J.; Evans, P.R.; McMahon, H.T. Curvature of clathrin-coated pits driven by epsin. Nature 2002, 419, 361–366. [Google Scholar] [CrossRef]
- Campelo, F.; Fabrikant, G.; McMahon, H.T.; Kozlov, M.M. Modeling membrane shaping by proteins: Focus on EHD2 and N-BAR domains. FEBS Lett. 2010, 584, 1830–1839. [Google Scholar] [CrossRef]
- Campelo, F.; McMahon, H.T.; Kozlov, M.M. The hydrophobic insertion mechanism of membrane curvature generation by proteins. Biophys. J. 2008, 95, 2325–2339. [Google Scholar] [CrossRef]
- Bley, C.J.; Nie, S.; Mobbs, G.W.; Petrovic, S.; Gres, A.T.; Liu, X.; Mukherjee, S.; Harvey, S.; Huber, F.M.; Lin, D.H.; et al. Architecture of the cytoplasmic face of the nuclear pore. bioRxiv 2021. [Google Scholar] [CrossRef]
- Petrovic, S.; Samanta, D.; Perriches, T.; Bley, C.J.; Thierbach, K.; Brown, B.; Nie, S.; Mobbs, G.W.; Stevens, T.A.; Liu, X.; et al. Architecture of the linker-scaffold in the nuclear pore. bioRxiv 2021. [Google Scholar] [CrossRef]
- Chen, X.Q.; Du, X.; Liu, J.; Balasubramanian, M.K.; Balasundaram, D. Identification of genes encoding putative nucleoporins and transport factors in the fission yeast Schizosaccharomyces pombe: A deletion analysis. Yeast 2004, 21, 495–509. [Google Scholar] [CrossRef]
- Roux, A.; Koster, G.; Lenz, M.; Sorre, B.; Manneville, J.B.; Nassoy, P.; Bassereau, P. Membrane curvature controls dynamin polymerization. Proc. Natl. Acad. Sci. USA 2010, 107, 4141–4146. [Google Scholar] [CrossRef]
- Chou, Y.Y.; Upadhyayula, S.; Houser, J.; He, K.; Skillern, W.; Scanavachi, G.; Dang, S.; Sanyal, A.; Ohashi, K.G.; Di Caprio, G.; et al. Inherited nuclear pore substructures template post-mitotic pore assembly. Dev. Cell 2021, 56, 1786–1803.e9. [Google Scholar] [CrossRef]
- Dawson, T.R.; Lazarus, M.D.; Hetzer, M.W.; Wente, S.R. ER membrane-bending proteins are necessary for de novo nuclear pore formation. J. Cell Biol. 2009, 184, 659–675. [Google Scholar] [CrossRef]
- Golchoubian, B.; Brunner, A.; Bragulat-Teixidor, H.; Neuner, A.; Akarlar, B.A.; Ozlu, N.; Schlaitz, A.L. Reticulon-like REEP4 at the inner nuclear membrane promotes nuclear pore complex formation. J. Cell Biol. 2022, 221, e202101049. [Google Scholar] [CrossRef]
- Zhang, W.; Khan, A.; Vitale, J.; Neuner, A.; Rink, K.; Luchtenborg, C.; Brugger, B.; Sollner, T.H.; Schiebel, E. A short perinuclear amphipathic alpha-helix in Apq12 promotes nuclear pore complex biogenesis. Open Biol. 2021, 11, 210250. [Google Scholar] [CrossRef]
- Floch, A.G.; Tareste, D.; Fuchs, P.F.; Chadrin, A.; Naciri, I.; Leger, T.; Schlenstedt, G.; Palancade, B.; Doye, V. Nuclear pore targeting of the yeast Pom33 nucleoporin depends on karyopherin and lipid binding. J. Cell Sci. 2015, 128, 305–316. [Google Scholar] [CrossRef]
- Heath, C.V.; Copeland, C.S.; Amberg, D.C.; Del Priore, V.; Snyder, M.; Cole, C.N. Nuclear pore complex clustering and nuclear accumulation of poly(A)+ RNA associated with mutation of the Saccharomyces cerevisiae RAT2/NUP120 gene. J. Cell Biol. 1995, 131, 1677–1697. [Google Scholar] [CrossRef]
- Franz, C.; Askjaer, P.; Antonin, W.; Iglesias, C.L.; Haselmann, U.; Schelder, M.; de Marco, A.; Wilm, M.; Antony, C.; Mattaj, I.W. Nup155 regulates nuclear envelope and nuclear pore complex formation in nematodes and vertebrates. EMBO J. 2005, 24, 3519–3531. [Google Scholar] [CrossRef]
- Makio, T.; Stanton, L.H.; Lin, C.C.; Goldfarb, D.S.; Weis, K.; Wozniak, R.W. The nucleoporins Nup170p and Nup157p are essential for nuclear pore complex assembly. J. Cell Biol. 2009, 185, 459–473. [Google Scholar] [CrossRef]
- Rout, M.P.; Aitchison, J.D.; Suprapto, A.; Hjertaas, K.; Zhao, Y.; Chait, B.T. The yeast nuclear pore complex: Composition, architecture, and transport mechanism. J. Cell Biol. 2000, 148, 635–651. [Google Scholar] [CrossRef]
- Van de Vosse, D.W.; Wan, Y.; Lapetina, D.L.; Chen, W.M.; Chiang, J.H.; Aitchison, J.D.; Wozniak, R.W. A role for the nucleoporin Nup170p in chromatin structure and gene silencing. Cell 2013, 152, 969–983. [Google Scholar] [CrossRef]
- Radu, A.; Blobel, G.; Moore, M.S. Identification of a protein complex that is required for nuclear protein import and mediates docking of import substrate to distinct nucleoporins. Proc. Natl. Acad. Sci. USA 1995, 92, 1769–1773. [Google Scholar] [CrossRef]
- Walther, T.C.; Fornerod, M.; Pickersgill, H.; Goldberg, M.; Allen, T.D.; Mattaj, I.W. The nucleoporin Nup153 is required for nuclear pore basket formation, nuclear pore complex anchoring and import of a subset of nuclear proteins. EMBO J. 2001, 20, 5703–5714. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Nucleoporin | Main Functions | Nucleoporin Interactions |
|---|---|---|
| Nup133 | part of the Y-complex [27] mRNA export [52,90] interphase NPC assembly [48] neural differentiation [50] proper NPC spacing in budding yeast [52,57] | binds within the Y-complex Nup84 and is also important for inter-Y-complex interactions [4,5]. The Y-complex acts as a central NPC building block and shows within NPCs manifold interactions, e.g., with Nup98, Nup153, POM121 [37,68,90] |
| Nup160 (Nup120 in budding yeast) | part of the Y-complex [27] mRNA export [90,118] proper NPC spacing in budding yeast [57,118] | binds within the Y-complex Nup85 and Nup96 (Nup145C in budding yeast) [4,5] |
| Nup53 (Nup53 and Nup59 in budding yeast) | part of the inner ring complex [4,5] mitotic and interphase NPC assembly [62,78] | NDC1, Nup155, Nup93 (NDC1, Nup157/Nup170 and Nic96 in budding yeast) [60,62,63,66,67,69,70,72,75,110] |
| Nup155 (Nup157 and Nup170 in budding yeast) | part of the inner ring complex [4,5] NPC assembly [119,120] mRNA export [121] gene silencing [122] | NDC1, POM121, Nup53, Nup93 (NDC1, Nup53/Nup59 and Nic96 in budding yeast) [37,66,67,69,70,72,110] |
| Nup153 (corresponds to Nup1 and Nup60 in budding yeast) | part of the nuclear basket [4] interphase NPC assembly [91] mRNA export [85,86] nuclear import [123] proper NPC spacing [124] | Y-complex [90] Nup50, TPR [88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamed, M.; Antonin, W. Dunking into the Lipid Bilayer: How Direct Membrane Binding of Nucleoporins Can Contribute to Nuclear Pore Complex Structure and Assembly. Cells 2021, 10, 3601. https://doi.org/10.3390/cells10123601
Hamed M, Antonin W. Dunking into the Lipid Bilayer: How Direct Membrane Binding of Nucleoporins Can Contribute to Nuclear Pore Complex Structure and Assembly. Cells. 2021; 10(12):3601. https://doi.org/10.3390/cells10123601
Chicago/Turabian StyleHamed, Mohamed, and Wolfram Antonin. 2021. "Dunking into the Lipid Bilayer: How Direct Membrane Binding of Nucleoporins Can Contribute to Nuclear Pore Complex Structure and Assembly" Cells 10, no. 12: 3601. https://doi.org/10.3390/cells10123601
APA StyleHamed, M., & Antonin, W. (2021). Dunking into the Lipid Bilayer: How Direct Membrane Binding of Nucleoporins Can Contribute to Nuclear Pore Complex Structure and Assembly. Cells, 10(12), 3601. https://doi.org/10.3390/cells10123601