
Comparative Genomics and Physiological Investigation of a New Arthrospira/Limnospira Strain O9.13F Isolated from an Alkaline, Winter Freezing, Siberian Lake

 ,
,  ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Strain Isolation and Growth Conditions
2.2. Phenotypic Characteristics of Arthrospira sp. O9.13F
2.2.1. Acclimation of Arthrospira Strains to Long-Term Stress of Low Temperature
2.2.2. The Survivability of Arthrospira Strains in Different Stress Conditions
2.2.3. Fatty Acids Methyl Ester Analysis (FAME)
2.2.4. Proteins Content Analysis
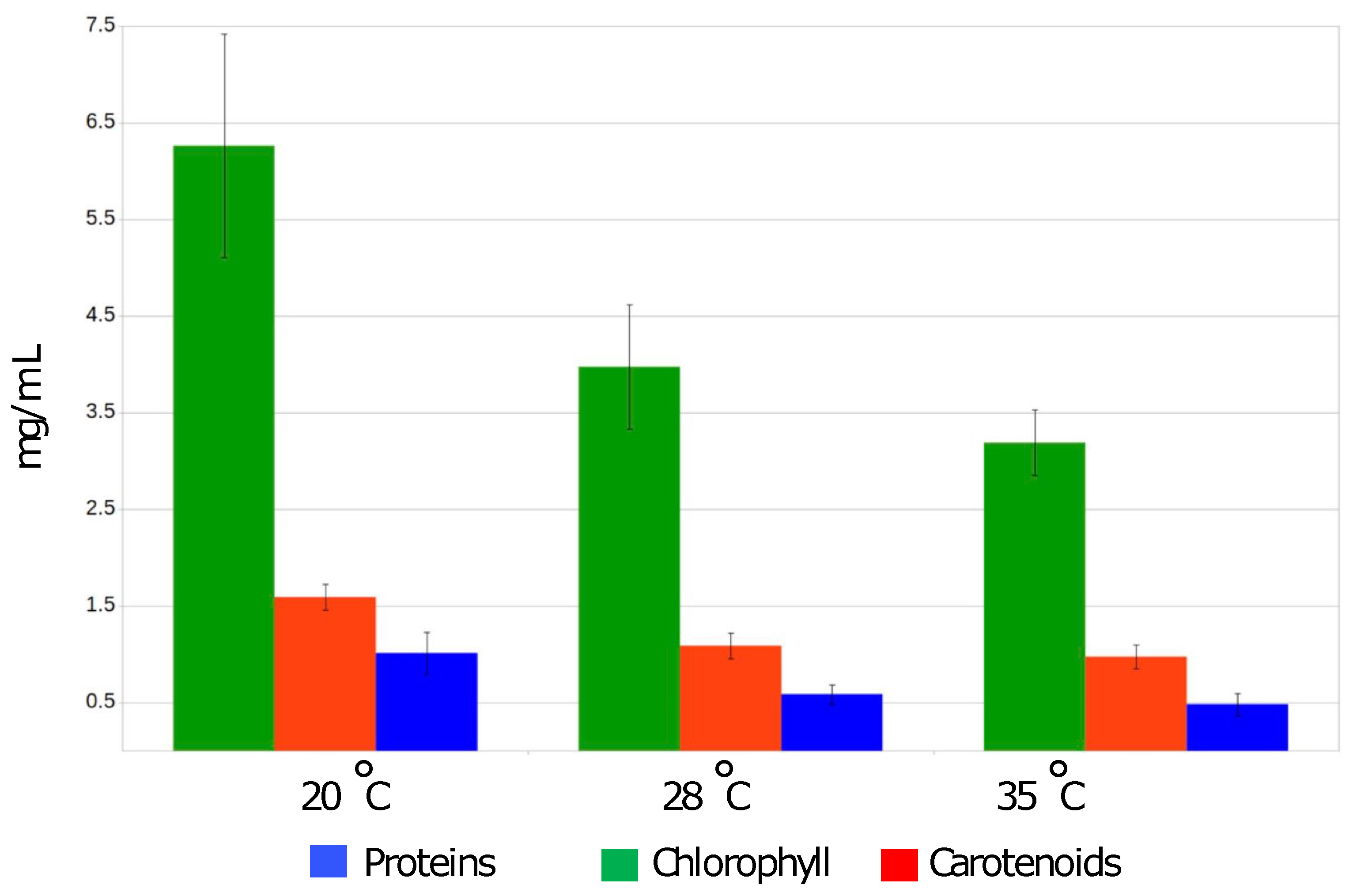
2.2.5. The Measurements of Photosynthetic Dyes Content
2.2.6. Statistical Analysis
2.3. Microscopic Observations
2.3.1. Light Microscopy
2.3.2. Transmission Electron Microscopy (TEM)
2.4. Molecular Characteristics
2.4.1. DNA Extraction
2.4.2. PCR Amplification Sequencing and Phylogenetic Analysis
2.4.3. Whole-Genome Sequencing and Annotation
2.4.4. Phylogenomic Analyses
3. Results
3.1. Phenotypic Characteristics of Arthrospira sp. O9.13F
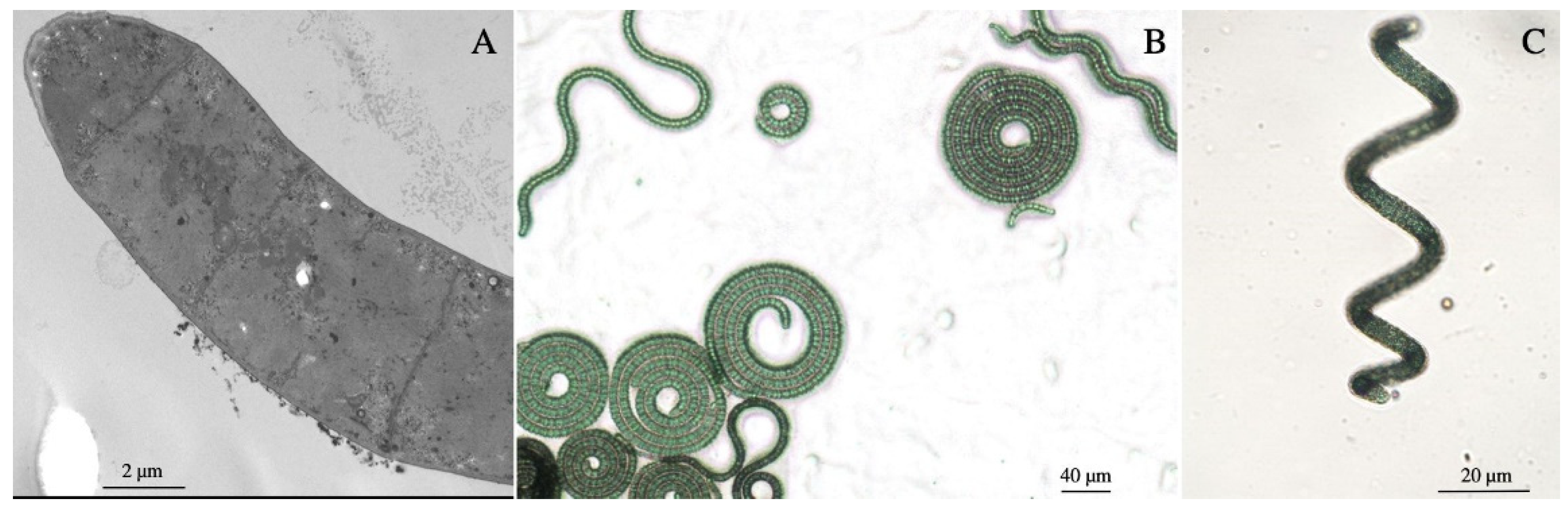
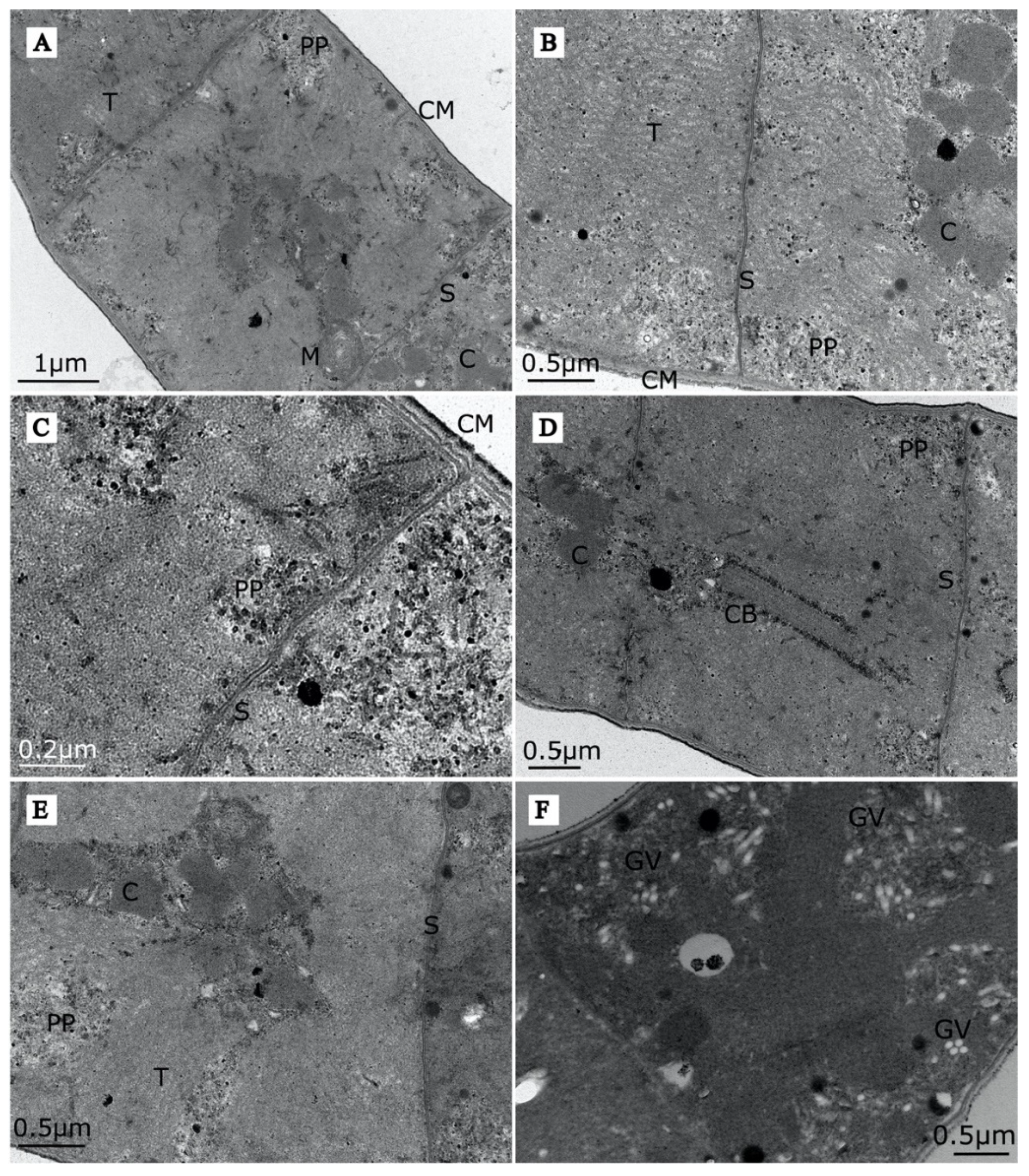
3.1.1. Morphology of Arthrospira sp. O9.13F
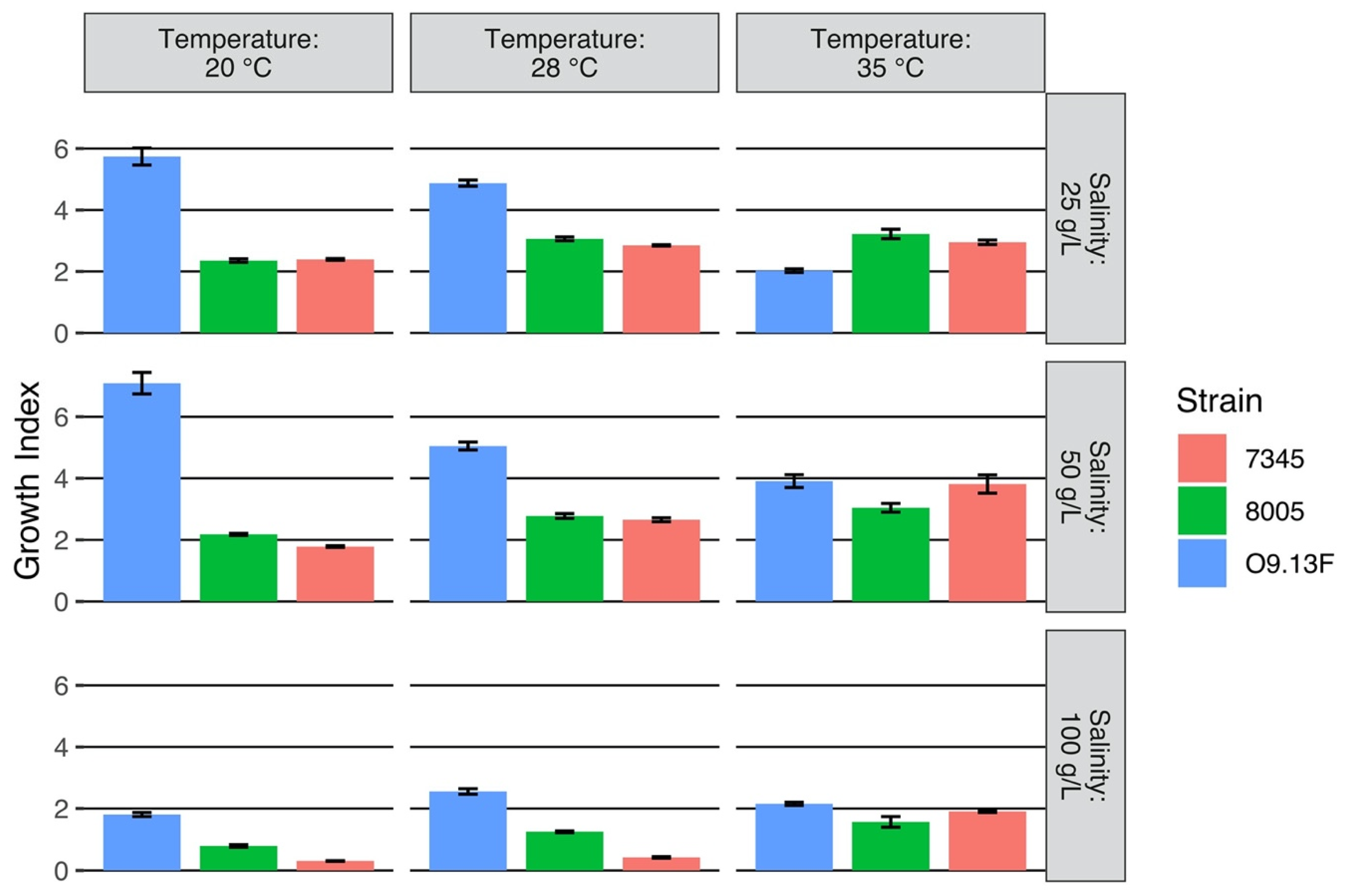
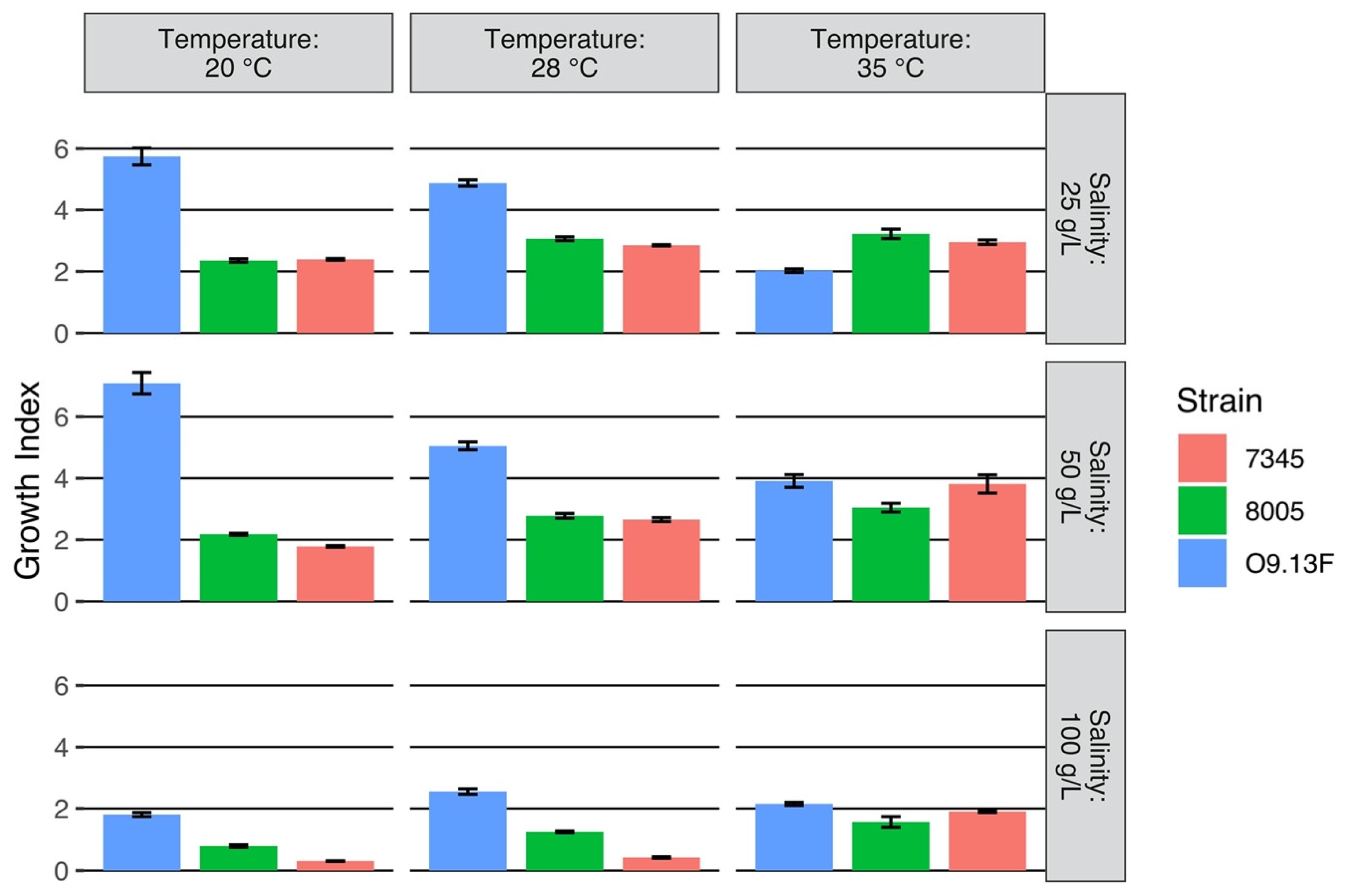
3.1.2. Growth Conditions
3.1.3. Acclimation to Prolonged Cold Stress
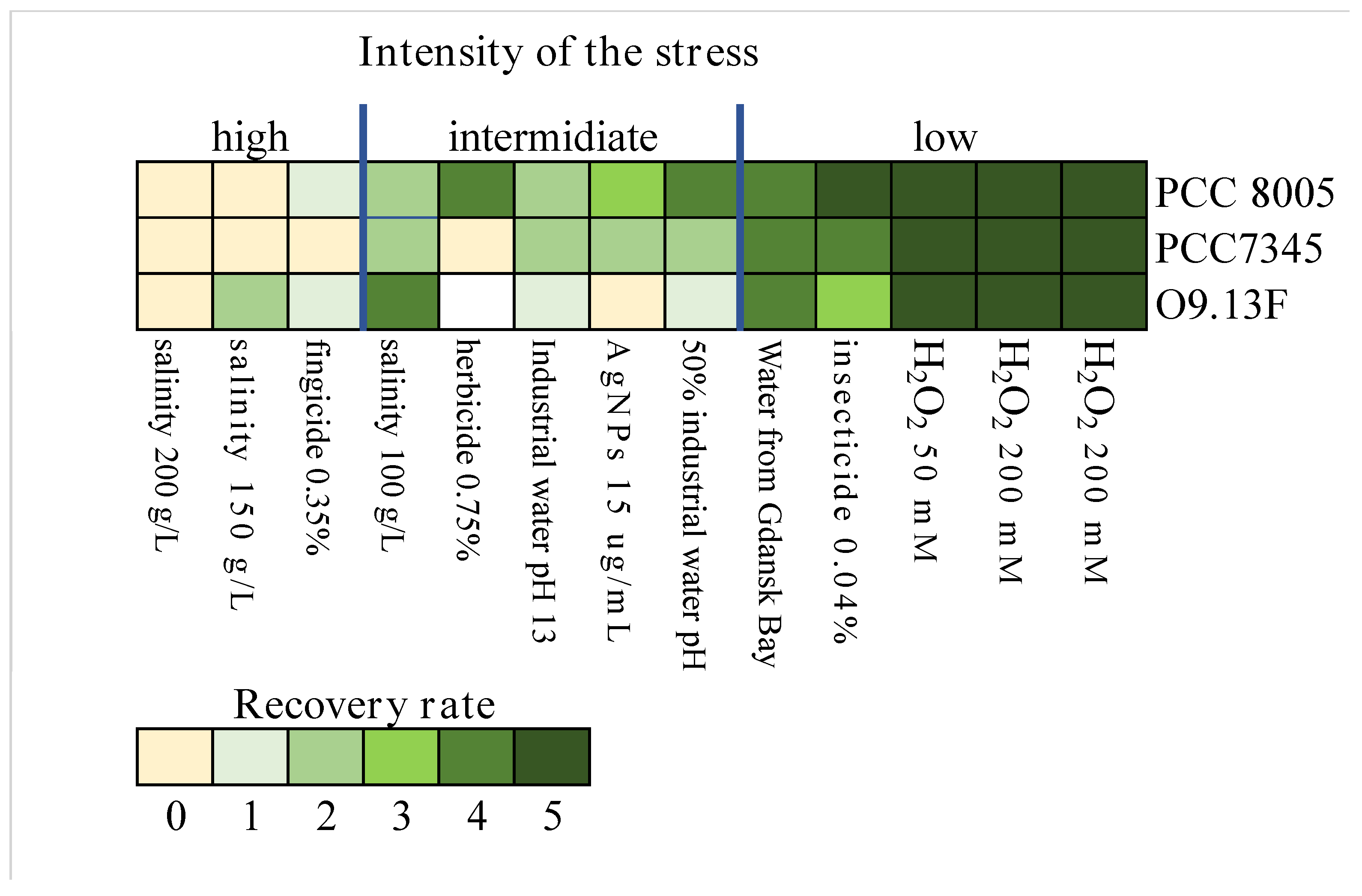
3.1.4. The Sensitivity of Arthrospira sp. O9.13F to Various Stressors
3.1.5. Fatty Acid Composition
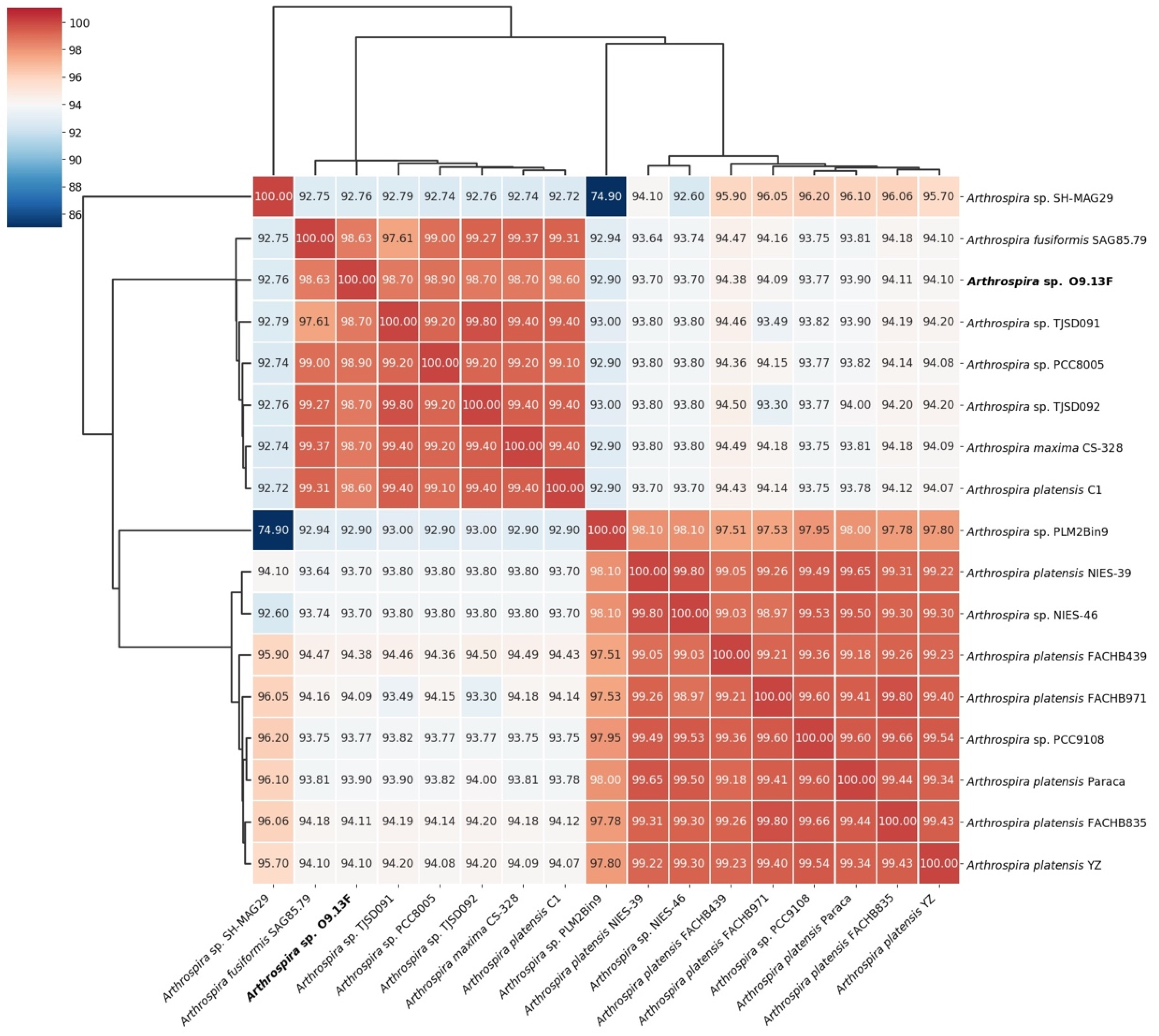
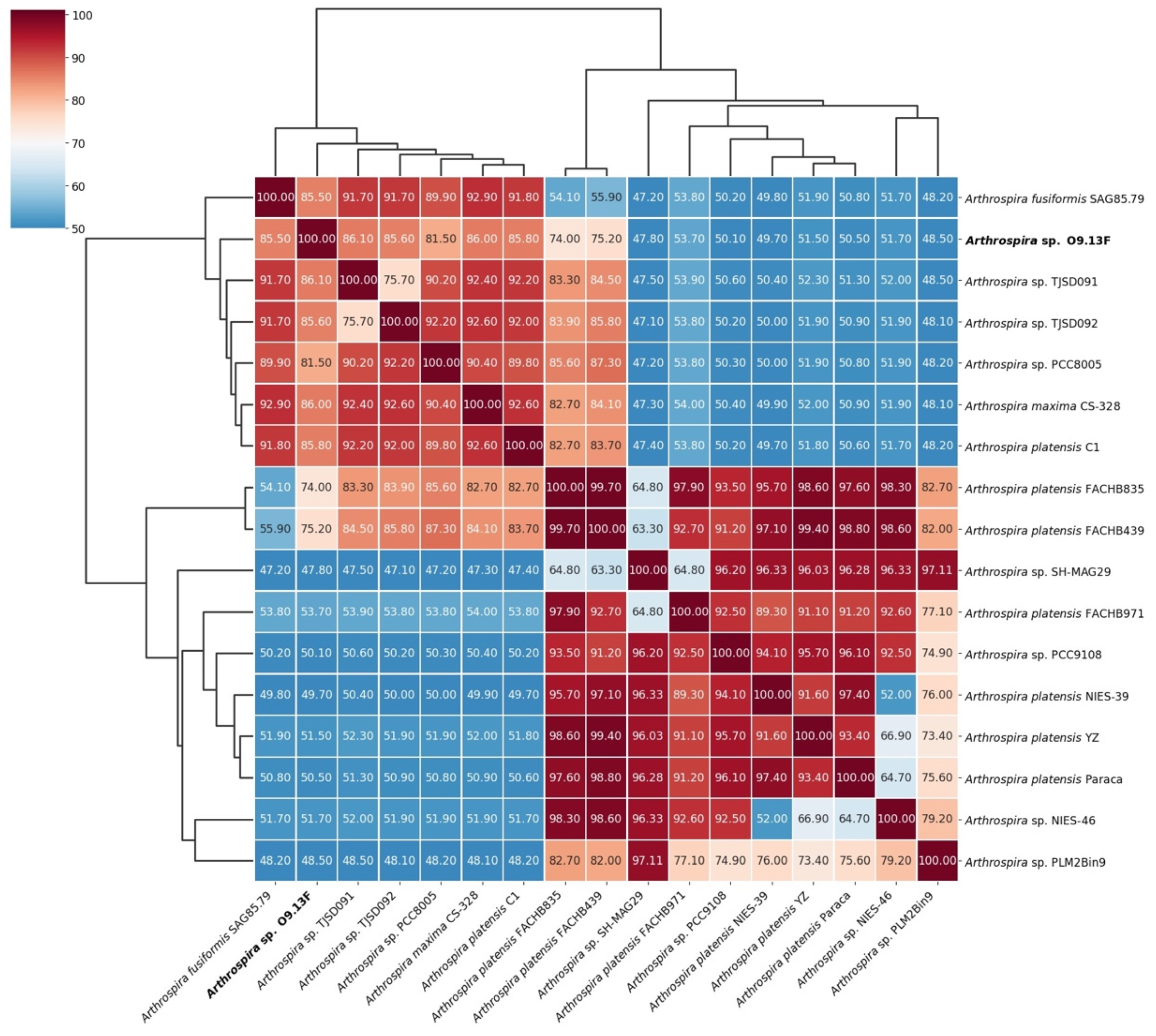
3.2. Taxonomy of Arthrospira sp. Strain O9.13F
3.3. General Features of the Arthrospira sp. O9.13F Genome
3.4. Comparative Genome Analyses
3.4.1. Genes Responsible for the Stress Adaptation
3.4.2. Arthrospira sp. O9.13F Unique Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castenholz, R.W.; Rippka, R.; Herdman, M.; Wilmotte, A. Form- Arthrospira. In Bergey’s Manual of Systematic Bacteriology; Staley, J.T., Bryant, M.P., Pfenning, N., Holt, J.G., Eds.; William & Wilkins: Baltimore, MD, USA, 1989; Volume 3, pp. 1774–1775. ISBN 978-1-118-96060-8. [Google Scholar]
- Nowicka-Krawczyk, P.; Muhlsteinova, R.; Hauer, T. Detailed Characterization of the Arthrospira Type Species Separating Commercially Grown Taxa into the New Genus Limnospira (Cyanobacteria). Sci. Rep. 2019, 9, 694. [Google Scholar] [CrossRef]
- Geitler, L. Dr. L. Rabenhorst’s Kryptogamen-Flora von Deutschland, und der Schweiz. Zweite, vollständig neu bearbeitete Auflage; Vierzehnter Band. Die Algen herausgegeben von Prof. Dr. R. Kolkwitz, Berlin, Cyanophyceae von Europa unter Berücksichtigung der anderen Kontinente/von Dr. Lothar Geitler; Kryptogamen-Flora von Deutschland, und der Schweiz. Zweite, vollständig neu bearbeitete Auflage, 2nd ed.; Akademische Verlagsgesellschaft m. b. H.: Leipzig, Germany, 1930; Volume 14. [Google Scholar]
- Ciferri, O. Spirulina, the Edible Microorganism. Microbiol. Rev. 1984, 47, 551–578. [Google Scholar] [CrossRef]
- Dadheech, P.K.; Ballot, A.; Casper, P.; Kotut, K.; Novelo, E.; Lemma, B.; Pröschold, T.; Krienitz, L. Phylogenetic Relationship and Divergence among Planktonic Strains of Arthrospira (Oscillatoriales, Cyanobacteria) of African, Asian and American Origin Deduced by 16S–23S ITS and Phycocyanin Operon Sequences. Phycologia 2010, 49, 361–372. [Google Scholar] [CrossRef]
- Linhart, C.; Schagerl, M. Seasonal Succession of the Travertine-Forming Desmid Oocardium Stratum. J. Phycol. 2015, 51, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Bazhenova, O.; Konovalova, O. Phytoplankton of Lake Solenoye (Omsk) as a Promising Source of Bioresources. Contemp. Probl. Ecol. 2012, 5, 275–280. [Google Scholar] [CrossRef]
- Leboulanger, C.; Agogué, H.; Bernard, C.; Bouvy, M.; Carré, C.; Cellamare, M.; Duval, C.; Fouilland, E.; Got, P.; Intertaglia, L.; et al. Microbial Diversity and Cyanobacterial Production in Dziani Dzaha Crater Lake, a Unique Tropical Thalassohaline Environment. PLoS ONE 2017, 12, e0168879. [Google Scholar] [CrossRef] [PubMed]
- Castro-Severyn, J.; Pardo-Esté, C.; Mendez, K.; Fortt, J.; Marquez, S.; Molina, F.; Castro-Nallar, E.; Remonsellez, F.; Saavedra, C. Living to the High Extreme: Unraveling the Composition, Structure, and Functional Insights of Bacterial Communities Thriving in the Arsenic-Rich Salar de Huasco Altiplanic Ecosystem. Microbiol. Spectr. 2021, 9, e0044421. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; Berben, T.; Melton, E.D.; Overmars, L.; Vavourakis, C.D.; Muyzer, G. Microbial Diversity and Biogeochemical Cycling in Soda Lakes. Extremophiles 2014, 18, 791–809. [Google Scholar] [CrossRef] [Green Version]
- Fužinato, S.; Fodora, A.; Subakov-Simić, G. Arthrospira Fusiformis (Voronichin) Komarek et Lund (Cyanoprokaryota) A New Species for Europe. Algol. Stud. 2010, 134, 17–24. [Google Scholar] [CrossRef]
- Vavourakis, C.; Mehrshad, M.; Balkema, C.; van Hall, R.; Andrei, A.-S.; Ghai, R.; Sorokin, D.; Muyzer, G. Metagenomes and Metatranscriptomes Shed New Light on the Microbial-Mediated Sulfur Cycle in a Siberian Soda Lake. BMC Biol. 2019, 17, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorz, J.; Sharp, C.; Kleiner, M.; Gordon, P.; Pon, R.; Dong, X.; Strous, M. A Shared Core Microbiome in Soda Lakes Separated by Large Distances. Nat. Commun. 2019, 10, 4230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, W.D.; Sorokin, D.Y. Distribution and Diversity of Soda Lake Alkaliphiles. In Extremophiles Handbook; Horikoshi, K., Ed.; Springer: Tokyo, Japan, 2011; pp. 27–54. ISBN 978-4-431-53898-1. [Google Scholar]
- Furmaniak, M.; Misztak, A.; Franczuk, M.; Wilmotte, A.; Waleron, M.; Waleron, K. Edible Cyanobacterial Genus Arthrospira: Actual State of the Art in Cultivation Methods, Genetics, and Application in Medicine. Front. Microbiol. 2017, 8, 2541. [Google Scholar] [CrossRef]
- Ananyev, G.; Carrieri, D.; Dismukes, G. Optimization of Metabolic Capacity and Flux through Environmental Cues to Maximize Hydrogen Production by the Cyanobacterium “Arthrospira (Spirulina) Maxima”. Appl. Environ. Microbiol. 2008, 74, 6102–6113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ainas, M.; Hasnaoui, S.; Bouarab, R.; Abdi, N.; Drouiche, N.; Mameri, N. Hydrogen Production with the Cyanobacterium Spirulina Platensis. Int. J. Hydrogen Energy 2017, 42, 4902–4907. [Google Scholar] [CrossRef]
- Krishnan, A.; Qian, X.; Ananyev, G.; Lun, D.; Dismukes, G. Rewiring of Cyanobacterial Metabolism for Hydrogen Production: Synthetic Biology Approaches and Challenges. Adv. Exp. Med. Biol. 2018, 1080, 171–213. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Yamaguchi, H.; Kawachi, M. The Draft Genome of a Hydrogen-Producing Cyanobacterium, Arthrospira Platensis NIES-46. J. Genom. 2019, 7, 56–59. [Google Scholar] [CrossRef]
- Badri, H.; Monsieurs, P.; Coninx, I.; Ruddy, W.; Leys, N. Molecular Investigation of the Radiation Resistance of Edible Cyanobacterium Arthrospira Sp. PCC 8005. MicrobiologyOpen 2015, 4, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, L.; De Wever, H.; Hermans, V.; Mastroleo, F.; Morin, N.; Wilmotte, A.; Janssen, P.; Mergeay, M. Microbial Ecology of the Closed Artificial Ecosystem MELiSSA (Micro-Ecological Life Support System Alternative): Reinventing and Compartmentalizing the Earth’s Food and Oxygen Regeneration System for Long-Haul Space Exploration Missions. Res. Microbiol. 2006, 157, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Gugliemi, G.; Rippka, R.; Tandeau De Marsac, N. Main Properties That Justify the Different Taxonomic Position of Spirulina spp. and Arthrospira spp. among Cyanobacteria. Bull. Inst. Oceanogr. Monaco 1993, 12, 13–23. [Google Scholar]
- Tomaselli, L. Morphology, Ultrastructure and Taxonomy of Arthrospira (Spirulina Maxima and Arthrospira (Spirulina) Platensis. In Spirulina platensis (Arthrospira): Physiology, Cell-Biology and Biotechnology; Vonshak, A., Ed.; Taylor & Francis Ltd.: London, UK, 1997; pp. 1–15. ISBN 978-0-429-07994-8. [Google Scholar]
- Komárek, J.; Lund, J.W.G. What Is «Spirulina Platensis» in Fact? Algol. Stud. Hydrobiol. Suppl. Vol. 1990, 58, 1–13. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Süßwasserflora von Mitteleuropa, Bd. 19/2: Cyanoprokaryota: Bd. 2/Part 2: Oscillatoriales; Elsveire GmbH: Munchen, Germany, 2005; ISBN 3-8274-0919-5. [Google Scholar]
- Bai, N.; Seshadri, C.V. On Coiling and Uncoiling of Trichomes in the Genus Spirulina. Algol. Stud. 1980, 26, 32–47. [Google Scholar]
- Bai, N. Competitive Exclusion or Morphological Transformation? A Case Study with Spirulina Fusiformis. Arch. Hydrobiol. 1985, 71, 191–199. [Google Scholar]
- Mühling, M.; Harris, N.; Belay, A.; Whitton, B. Reversal of Helix Orientation in the Cyanobacterium Arthrospira. J. Phycol. 2003, 39, 360–367. [Google Scholar] [CrossRef]
- Scheldeman, P.; Baurain, D.; Bouhy, R.; Scott, M.; Mühling, M.; Whitton, B.; Belay, A.; Wilmotte, A. Arthrospira (‘Spirulina’) Strains from Four Continents Are Resolved into Only Two Clusters, Based on Amplified Ribosomal DNA Restriction Analysis of the Internally Transcribed Spacer. FEMS Microbiol. Lett. 1999, 172, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Baurain, D.; Renquin, L.; Grubisic, S.; Scheldeman, P.; Belay, A.; Wilmotte, A. Remarkable Conservation of Internally Transcribed Spacer Sequences of Arthrospira (“Spirulina”) (Cyanophyceae, Cyanobacteria) Strains from Four Continents and of Recent and 30-Year-Old Dried Samples from Africa. J. Phycol. 2002, 38, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Hicks, M.; Tran-Dao, T.-K.; Mulroney, L.; Bernick, D. De-Novo Assembly of Limnospira Fusiformis Using Ultra-Long Reads. Front. Microbiol. 2021, 12, 657995. [Google Scholar] [CrossRef] [PubMed]
- Manen, J.-F.; Falquet, J. The CpcB-CpcA Locus as a Tool for the Genetic Characterization of the Genus Arthrospira (Cyanobacteria): Evidence for Horizontal Transfer. Int. J. Syst. Evol. Microbiol. 2002, 52, 861–867. [Google Scholar] [CrossRef] [Green Version]
- Ballot, A.; Dadheech, P.; Krienitz, L. Phylogenetic Relationship of Arthrospira, Phormidium and Spirulina Strains from Kenyan and Indian Waterbodies. Algol. Stud. 2004, 113, 37–56. [Google Scholar] [CrossRef]
- Papapanagiotou, G.; Gkelis, S. Taxonomic Revision of Commercially Used Arthrospira (Cyanobacteria) Strains: A Polyphasic Approach. Eur. J. Phycol. 2019, 54, 595–608. [Google Scholar] [CrossRef]
- Mühling, M.; Somerfield, P.; Harris, N.; Belay, A.; Whitton, B. Phenotypic Analysis of Arthrospira (Spirulina) Strains (Cyanobacteria). Phycologia 2006, 45, 148–157. [Google Scholar] [CrossRef]
- Ogato, T.; Kifle, D.; Fetahi, T.; Sitotaw, B. Evaluation of Growth and Biomass Production of Arthrospira (Spirulina) Fusiformis in Laboratory Cultures Using Waters from the Ethiopian Soda Lakes Chitu and Shala. J. Appl. Phycol. 2014, 26, 2273–2282. [Google Scholar] [CrossRef]
- Janssen, P.; Morin, N.; Mergeay, M.; Leroy, B.; Ruddy, W.; Vallaeys, T.; Waleron, K.; Waleron, M.; Wilmotte, A.; Quillardet, P.; et al. Genome Sequence of the Edible Cyanobacterium Arthrospira Sp. PCC 8005. J. Bacteriol. 2010, 192, 2465–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Narikawa, R.; Okamoto, S.; Ehira, S.; Yoshimura, H.; Suzuki, I.; Masuda, T.; Mochimaru, M.; Takaichi, S.; Awai, K.; et al. Genomic Structure of an Economically Important Cyanobacterium, Arthrospira (Spirulina) Platensis NIES-39. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2010, 17, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheevadhanarak, S.; Paithoonrangsarid, K.; Prommeenate, P.; Kaewngam, W.; Musigkain, A.; Tragoonrung, S.; Tabata, S.; Kaneko, T.; Chaijaruwanich, J.; Sangsrakru, D.; et al. Draft Genome Sequence of Arthrospira Platensis C1 (PCC9438). Stand. Genom. Sci. 2012, 6, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrieri, D.; Ananyev, G.; Lenz, O.; Bryant, D.; Dismukes, G. Contribution of a Sodium Ion Gradient to Energy Conservation during Fermentation in the Cyanobacterium Arthrospira (Spirulina) Maxima CS-328. Appl. Environ. Microbiol. 2011, 77, 7185–7194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, F.; Calmin, G.; Crovadore, J.; Falquet, J.; Hurni, J.-P.; Østerås, M.; Haldemann, F.; Farinelli, L. Whole-Genome Shotgun Sequence of Arthrospira Platensis Strain Paraca, a Cultivated and Edible Cyanobacterium. Genome Announc. 2014, 2, e00751-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, T.; Qin, S.; Hu, Y.; Song, Z.; Ying, J.; Li, P.; Dong, W.; Zhao, F.; Yang, H.; Bao, Q. Whole Genomic DNA Sequencing and Comparative Genomic Analysis of Arthrospira Platensis: High Genome Plasticity and Genetic Diversity. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2016, 23, 325–338. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Chen, J.; Wang, S.; Wu, Y.; Hou, H.; Li, M.; Yan, C. Draft Genome Sequence of Cyanobacteria Arthrospira Sp. TJSD091 Isolated from Seaside Wetland. Mar. Genom. 2015, 24, 197–198. [Google Scholar] [CrossRef] [PubMed]
- Sharp, C.; Urschel, S.; Dong, X.; Brady, A.; Slater, G.; Strous, M. Robust, High-Productivity Phototrophic Carbon Capture at High PH and Alkalinity Using Natural Microbial Communities. Biotechnol. Biofuels 2017, 10, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.-Y.; Teng, W.; Zhao, L.; Hu, C.-X.; Zhou, Y.; Han, B.-P.; Song, L.-R.; Shu, W.-S. Comparative Genomics Reveals Insights into Cyanobacterial Evolution and Habitat Adaptation. ISME J. 2020, 15, 211–227. [Google Scholar] [CrossRef] [PubMed]
- Maghembe, R.; Angelina, M.; Harish, A.; Nyandoro, S.; Lyantagaye, S.; Hati-Kaul, R. Draft Genome Sequence of Limnospira Sp. Strain BM01, Isolated from a Hypersaline Lake of the Momela Ecosystem in Tanzania. Microbiol. Resour. Announc. 2021, 10, e00132-21. [Google Scholar] [CrossRef] [PubMed]
- Zarrouk, C.; Zarrouk, C. Contribution à l´Étude d´une Cyanophycée Influence de Divers Facteurs Physiques et Chimiques sur la Croissance et la Photosynthése de Spirulina Maxima Geitler; Academie de Paris: Paris, France, 1966. [Google Scholar]
- Rippka, R. Isolation and Purification of Cyanobacteria. Methods Enzymol. 1988, 167, 3–27. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.L.G.; Dyer, W.J.A. A Rapid Method of Total Lipid Extraction And Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ritchie, R.J. Consistent Sets of Spectrophotometric Chlorophyll Equations for Acetone, Methanol and Ethanol Solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Wellburn, A. The Spectral Determination of Chlorophylls a and b, as Well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-153, 2021. Available online: https://CRAN.R-project.org/package=nlme (accessed on 1 October 2020).
- Eykelenburg, C. The Ultrastructure of Spirulina Platensis in Relation to Temperature and Light Intensity. Antonie Leeuwenhoek 1979, 45, 369–390. [Google Scholar] [CrossRef] [PubMed]
- Hayat, M.A. Principles and Techniques of Electron Microscopy: Biological Applications; Edward Arnold: London, UK, 1981; ISBN 978-0-7131-2830-7. [Google Scholar]
- William, S.; Feil, H.; Copeland, A. Bacterial Genomic DNA Isolation Using CTAB 2012. Available online: http://1ofdmq2n8tc36m6i46scovo2e.wpengine.netdna-cdn.com/wp-content/uploads/2014/02/JGI-Bacterial-DNA-isolation-CTAB-Protocol-2012.pdf (accessed on 1 October 2019).
- Waleron, M.; Waleron, K.; Vincent, W.F.; Wilmotte, A. Allochthonous Inputs of Riverine Picocyanobacteria to Coastal Waters in the Arctic Ocean. FEMS Microbiol. Ecol. 2007, 59, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Hrouzek, P.; Ventura, S.; Lukesova, A.; Mugnai, M.; Turicchia, S.; Komárek, J. Diversity of Soil Nostoc Strains: Phylogenetic and Phenotypic Variability. Algol. Stud. 2005, 117, 251–264. [Google Scholar] [CrossRef]
- Taton, A.; Grubisic, S.; Brambilla, E.; De Wit, R.; Wilmotte, A. Cyanobacterial Diversity in Natural and Artificial Microbial Mats of Lake Fryxell (McMurdo Dry Valleys, Antarctica): A Morphological and Molecular Approach. Appl. Environ. Microbiol. 2003, 69, 5157–5169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilmotte, A.; Van der Auwera, G.; Wachter, R. Structure of the 16 S Ribosomal RNA of the Thermophilic Cyanobacterium Chlorogloeopsis HTF (‘Mastigocladus Laminosus HTF’) Strain PCC7518, and Phylogenetic Analysis. FEBS Lett. 1993, 317, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinform. Oxf. Engl. 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.; Salzberg, S.; Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast Metagenomic Sequence Classification Using Exact Alignment. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinform. Oxf. Engl. 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikhi, R.; Medvedev, P. Informed and Automated K-Mer Size Selection for Genome Assembly. Bioinform. Oxf. Engl. 2013, 30, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.; Dvorkin, M.; Kulikov, A.; Lesin, V.; Nikolenko, S.; Pham, S.; Prjibelski, A.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbino, D.; Birney, E. Velvet: Algorithms for De Novo Short Read Assembly Using De Bruijn Graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Chevreux, B.; Wetter, T.; Suhai, S. Genome Sequence Assembly Using Trace Signals and Additional Sequence Information. In Proceedings of the Computer Science and Biology: Proceedings of the German Conference on Bioinformatics (GCB’99), Braunschweig, Germany, 7–10 October 1999; Volume 99, p. 56. [Google Scholar]
- Boisvert, S.; Laviolette, F.; Corbeil, J. Ray: Simultaneous Assembly of Reads from a Mix of High-Throughput Sequencing Technologies. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2010, 17, 1519–1533. [Google Scholar] [CrossRef] [PubMed]
- Boetzer, M.; Henkel, C.; Jansen, H.; Butler, D.; Pirovano, W. Scaffolding Pre-Assembled Contigs Using SSPACE. Bioinform. Oxf. Engl. 2011, 27, 578–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinform. Oxf. Engl. 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.; Borodovsky, M.; Ostell, J. NCBI Prokaryotic Genome Annotation Pipeline. Nucleic Acids Res. 2016, 44, gkw569. [Google Scholar] [CrossRef] [PubMed]
- Haft, D.; DiCuccio, M.; Badretdin, A.; Brover, V.; Chetvernin, V.; O’Neill, K.; Li, W.; Chitsaz, F.; Derbyshire, M.; Gonzales, N.; et al. RefSeq: An Update on Prokaryotic Genome Annotation and Curation. Nucleic Acids Res. 2017, 46, D851–D860. [Google Scholar] [CrossRef] [PubMed]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.; Kanehisa, M.; Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An Automatic Genome Annotation and Pathway Reconstruction Server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koski, L.; Gray, M.; Lang, B.; Burger, G. AutoFACT: An Automatic Functional Annotation and Classification Tool. BMC Bioinform. 2005, 6, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Käll, L.; Krogh, A.; Sonnhammer, E. Advantages of Combined Transmembrane Topology and Signal Peptide Prediction—The Phobius Web Server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, R.; Vincze, T.; Posfai, J.; Macelis, D. REBASE-A Database for DNA Restriction and Modification: Enzymes, Genes and Genomes. Nucleic Acids Res. 2015, 43, D298–D299. [Google Scholar] [CrossRef] [PubMed]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A Web Tool to Identify Clustered Regularly Interspaced Short Palindromic Repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallenet, D.; Calteau, A.; Dubois, M.; Amours, P.; Bazin, A.; Beuvin, M.; Burlot, L.; Bussell, X.; Fouteau, S.; Gautreau, G.; et al. MicroScope: An Integrated Platform for the Annotation and Exploration of Microbial Gene Functions through Genomic, Pangenomic and Metabolic Comparative Analysis. Nucleic Acids Res. 2019, 48, D579–D589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arndt, D.; Grant, J.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D. PHASTER: A Better, Faster Version of the PHAST Phage Search Tool. Nucleic Acids Res. 2016, 44, gkw387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Larsen, M.; Lund, O.; Villa, L.; Aarestrup, F.; Hasman, H. In Silico Detection and Typing of Plasmids Using PlasmidFinder and Plasmid Multilocus Sequence Typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, T.Y.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.; Müller, R.; Wohlleben, W.; et al. AntiSMASH 3.0-A Comprehensive Resource for the Genome Mining of Biosynthetic Gene Clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blin, K.; Medema, M.; Kottmann, R.; Lee, S.Y.; Weber, T. The AntiSMASH Database, a Comprehensive Database of Microbial Secondary Metabolite Biosynthetic Gene Clusters. Nucleic Acids Res. 2016, 45, gkw960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segata, N.; Börnigen, D.; Morgan, X.; Huttenhower, C. PhyloPhlAn Is a New Method for Improved Phylogenetic and Taxonomic Placement of Microbes. Nat. Commun. 2013, 4, 2304. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, L.; Glover, R.; Humphris, S.; Elphinstone, J.; Toth, I. Genomics and Taxonomy in Diagnostics for Food Security: Soft-Rotting Enterobacterial Plant Pathogens. Anal. Methods 2015, 8, 12–24. [Google Scholar] [CrossRef]
- Kurtz, S.; Phillippy, A.; Delcher, A.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S. Versatile and Open Software for Comparing Large Genomes. Genome Biol. 2004, 5, R12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.; Auch, A.; Klenk, H.-P.; Göker, M. Genome Sequence-Based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comte, K.; Coursin, T.; Carre-Mlouka, A. A New Genotype in the Genus Arthrospira (Oscillatoriales, Cyanobacteria) Revealed by a Mosaic-like Structure of the 16S-23SrRNA Intergenic Spacer Region in Strain PCC 9901. Phycologia 2013, 52, 333–337. [Google Scholar] [CrossRef]
- Burns, B.; Saker, M.; Moffitt, M.; Neilan, B. Molecular Detection of Genes Responsible for Cyanobacterial Toxin Production in the Genera Microcystis, Nodularia, and Cylindrospermopsis. Methods Mol. Biol. 2004, 268, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Waleron, M.; Misztak, A.; Waleron, M.; Furmaniak, M.; Mrozik, A.; Waleron, K. Arthrospiribacter Ruber Gen. Nov., Sp. Nov., a Novel Bacterium Isolated from Arthrospira Cultures. Syst. Appl. Microbiol. 2020, 43, 126072. [Google Scholar] [CrossRef] [PubMed]
- Hoiczyk, E.; Baumeister, W. Oscillin, an Extracellular, Ca2+-Binding Glycoprotein Essential for the Gliding Motility of Cyanobacteria. Mol. Microbiol. 1997, 26, 699–708. [Google Scholar] [CrossRef]
- Miklaszewska, M.; Waleron, M.; Morin, N.; Calusinska, M.; Wilmotte, A.; Marsac, N.; Rippka, R.; Waleron, K. Elucidation of the Gas Vesicle Gene Clusters in Cyanobacteria of the Genus Arthrospira (Oscillatoriales, Cyanophyta) and Correlation with ITS Phylogeny. Eur. J. Phycol. 2012, 47, 233–244. [Google Scholar] [CrossRef]
- Carre-Mlouka, A.; Comte, K.; Castets, A.-M.; Bouchier, C.; Marsac, N. The Gas Vesicle Gene Cluster from Microcystis Aeruginosa and DNA Rearrangements That Lead to Loss of Cell Buoyancy. J. Bacteriol. 2004, 186, 2355–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagemann, M. Molecular Biology of Cyanobacterial Salt Acclimation. FEMS Microbiol. Rev. 2011, 35, 87–123. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M. Chapter Two—Genomics of Salt Acclimation: Synthesis of Compatible Solutes among Cyanobacteria. In Advances in Botanical Research; Chauvat, F., Cassier-Chauvat, C., Eds.; Genomics of Cyanobacteria; Academic Press: Cambridge, MA, USA, 2013; Volume 65, pp. 27–55. [Google Scholar]
- Matelska, D.; Ginalski, K. Comprehensive Classification of the PIN Domain-like Superfamily. Nucleic Acids Res. 2017, 45, 6995–7020. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dong, C.; Li, L.; Wasney, G.; Min, J. Structural Insights into the Modulatory Role of the Accessory Protein WYL1 in the Type VI-D CRISPR-Cas System. Nucleic Acids Res. 2019, 47, 5420–5428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.; Zhang, F. Coupling Immunity and Programmed Cell Suicide in Prokaryotes: Life-or-Death Choices. BioEssays 2016, 39, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stappen, G.; Litvinenko, L.; Litvinenko, A.; Boyko, E.; Marden, B.; Sorgeloos, P. A Survey of Artemia Resources of Southwest Siberia (Russian Federation). Rev. Fish. Sci. 2009, 17, 117–148. [Google Scholar] [CrossRef]
- Mühling, M.; Belay, A.; Whitton, B. Variation in Fatty Acid Composition of Arthrospira (Spirulina) Strains. J. Appl. Phycol. 2005, 17, 137–146. [Google Scholar] [CrossRef]
- Colla, L.M.; Bertolin, T.E.; Costa, J.A.V. Fatty Acids Profile of Spirulina Platensis Grown Under Different Temperatures and Nitrogen Concentrations. Z. Nat. C 2004, 59, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Shiraishi, H. Cryopreservation of the Edible Alkalophilic Cyanobacterium Arthrospira Platensis. Biosci. Biotechnol. Biochem. 2016, 80, 2051–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | O9.13F | PCC 8005 | PCC 7345 |
|---|---|---|---|
| 16:0 | 36.71 ± 3.93 | 30.81 ± 1.2 | 31.64 ± 3.87 |
| 16:1 | 8.80 ± 1.1 | 10.29 ± 0.65 | 9.97 ± 1.13 |
| 18:0 | 1.12 ± 0.33 | 0.87 ± 0.16 | 1.62 ± 0.68 |
| 18:1n9c | 2.40 ± 0.97 | 2.59 ± 0.32 | 1.54 ± 0.66 |
| 18:1n11c | 1.67 ± 0.3 | 1.29 ± 0.04 | 1.32 ± 0.35 |
| 18:2n6c | 18.86 ± 0.34 | 18.78 ± 0.9 | 19.66 ± 2.43 |
| 18:3n6 | 30.45 ± 4.45 | 35.37 ± 1.98 | 34.26 ± 6.16 |
| Attribute | Value | % of Total |
|---|---|---|
| Genome size (bp) | 4,945,448 | 100 |
| DNA coding (bp) | 3,759,108 | 76.01 |
| DNA G + C (bp) | 2,198,162 | 44.5 |
| DNA scaffolds | 928 | - |
| Total genes | 5355 | 100 |
| Protein coding genes | 4688 | 87.5 |
| RNA genes | 42 | 0.78 |
| Pseudogenes | 625 | 11.67 |
| Genes in internal clusters | - | - |
| Genes with function prediction | 3363 | 62.80 |
| Genes assigned to COGs | 3278 | 69.92 |
| Genes with Pfam domains | 3237 | 69.05 |
| Genes with signal peptides | 400 | 8.53 |
| Genes with transmembrane helices | 862 | 18.39 |
| CRISPR repeats | 3 | 0.06 |
| Code | Value | % of Total * | Description |
|---|---|---|---|
| J | 146 | 4.45 | Translation, ribosomal structure and biogenesis |
| A | 0 | 0 | RNA processing and modification |
| K | 145 | 4.42 | Transcription |
| L | 311 | 9.49 | Replication, recombination and repair |
| B | 2 | 0.06 | Chromatin structure and dynamics |
| D | 25 | 0.76 | Cell cycle control, cell division, chromosome partitioning |
| V | 84 | 2.56 | Defence mechanisms |
| T | 308 | 9.4 | Signal transduction mechanisms |
| M | 209 | 6.38 | Cell wall/membrane biogenesis |
| N | 39 | 1.19 | Cell motility |
| U | 44 | 1.34 | Intracellular trafficking and secretion |
| O | 156 | 4.76 | Posttranslational modification, protein turnover, chaperones |
| C | 174 | 5.31 | Energy production and conversion |
| G | 140 | 4.27 | Carbohydrate transport and metabolism |
| E | 195 | 5.95 | Amino acid transport and metabolism |
| F | 58 | 1.77 | Nucleotide transport and metabolism |
| H | 115 | 3.51 | Coenzyme transport and metabolism |
| I | 73 | 2.23 | Lipid transport and metabolism |
| P | 138 | 4.21 | Inorganic ion transport and metabolism |
| Q | 95 | 2.90 | Secondary metabolites biosynthesis, transport and catabolism |
| R | 499 | 15.22 | General function prediction only |
| S | 322 | 9.82 | Function unknown |
| - | 1811 | 33.82 | Not in COGs |
| ORF | Description | The Most Similar Sequence in Genbank | Identity | |
|---|---|---|---|---|
| 1 | B9S53_26655 | hypothetical protein | - | - |
| 2 | B9S53_26345 | hypothetical protein | AI-2E family transporter [Natronoflexus pectinivorans] WP_165921875.1 | 61% |
| 3 | B9S53_26040 | IS91 family transposase | transposase [Lentisphaerae bacterium] MBM4144559.1 | 73% |
| 4 | B9S53_26035 | hypothetical protein | WYL domain-containing protein [Lentisphaerae bacterium] MBM4165248.1 | 83% |
| 5 | B9S53_25925 | hypothetical protein | - | - |
| 6 | B9S53_25920 | RNA-binding protein | RNA-binding S4 domain-containing protein [Flavobacteriales bacterium] TVQ76998.1 | 78% |
| 7 | B9S53_25790 | hypothetical protein | - | - |
| 8 | B9S53_25595 | four helix bundle protein | four helix bundle protein [Chloroflexi bacterium] HFC09004.1 | 67% |
| 9 | B9S53_25590 | hypothetical protein | four helix bundle protein [Chloroflexi bacterium] HFC09004.1 | 53% |
| 10 | B9S53_25580 | four helix bundle protein | - | |
| 11 | B9S53_24895 | hypothetical protein | N-6 DNA methylase [Sphaerospermopsis sp. FACHB-1094] WP_190648063.1 | 59% |
| 12 | B9S53_23130 | hypothetical protein | ROK family protein [Pseudanabaena sp. FACHB-1277] WP_190353199.1 | 43% |
| 13 | B9S53_22765 | hypothetical protein | - | - |
| 14 | B9S53_21915 | hypothetical protein | - | - |
| 15 | B9S53_21010 | IS4 family transposase | transposase [Phormidium sp. FACHB-1136] WP_199324687.1 | 75% |
| 16 | B9S53_20985 | hypothetical protein | - | - |
| 17 | B9S53_20435 | hypothetical protein | hypothetical protein [Cylindrospermopsis raciborskii] WP_071249329.1 | 67% |
| 18 | B9S53_19635 | hypothetical protein | - | - |
| 19 | B9S53_12695 | hypothetical protein | - | - |
| 20 | B9S53_12545 | hypothetical protein | - | - |
| 21 | B9S53_11425 | hypothetical protein | aldolase [Chloroflexi bacterium] MBT7081734.1 | 50% |
| 22 | B9S53_11330 | IS4 family transposase | transposase, partial [Leptolyngbya sp. PCC 6406] WP_155834780.1 | 75% |
| 23 | B9S53_06535 | DUF2281 domain-containing protein | DUF2281 domain-containing protein [Geitlerinema sp. P-1104] NMG59147.1 | 94% |
| 24 | B9S53_01945 | hypothetical protein | hypothetical protein [Nodularia sp. LEGE 06071] WP_193996843.1 | 32% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Misztak, A.E.; Waleron, M.; Furmaniak, M.; Waleron, M.M.; Bazhenova, O.; Daroch, M.; Waleron, K.F. Comparative Genomics and Physiological Investigation of a New Arthrospira/Limnospira Strain O9.13F Isolated from an Alkaline, Winter Freezing, Siberian Lake. Cells 2021, 10, 3411. https://doi.org/10.3390/cells10123411
Misztak AE, Waleron M, Furmaniak M, Waleron MM, Bazhenova O, Daroch M, Waleron KF. Comparative Genomics and Physiological Investigation of a New Arthrospira/Limnospira Strain O9.13F Isolated from an Alkaline, Winter Freezing, Siberian Lake. Cells. 2021; 10(12):3411. https://doi.org/10.3390/cells10123411
Chicago/Turabian StyleMisztak, Agnieszka E., Malgorzata Waleron, Magda Furmaniak, Michal M. Waleron, Olga Bazhenova, Maurycy Daroch, and Krzysztof F. Waleron. 2021. "Comparative Genomics and Physiological Investigation of a New Arthrospira/Limnospira Strain O9.13F Isolated from an Alkaline, Winter Freezing, Siberian Lake" Cells 10, no. 12: 3411. https://doi.org/10.3390/cells10123411
APA StyleMisztak, A. E., Waleron, M., Furmaniak, M., Waleron, M. M., Bazhenova, O., Daroch, M., & Waleron, K. F. (2021). Comparative Genomics and Physiological Investigation of a New Arthrospira/Limnospira Strain O9.13F Isolated from an Alkaline, Winter Freezing, Siberian Lake. Cells, 10(12), 3411. https://doi.org/10.3390/cells10123411