
NK and T Cell Immunological Signatures in Hospitalized Patients with COVID-19

 ,
,  ,
,  ,
,  , ,
, , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Preparation and Storage of PBMCs
2.3. Antibodies
2.4. Flow Cytometry Analysis of NK and T Cells
2.5. Immunoassay
3. Statistical Analysis
4. Results
4.1. Data of Patients
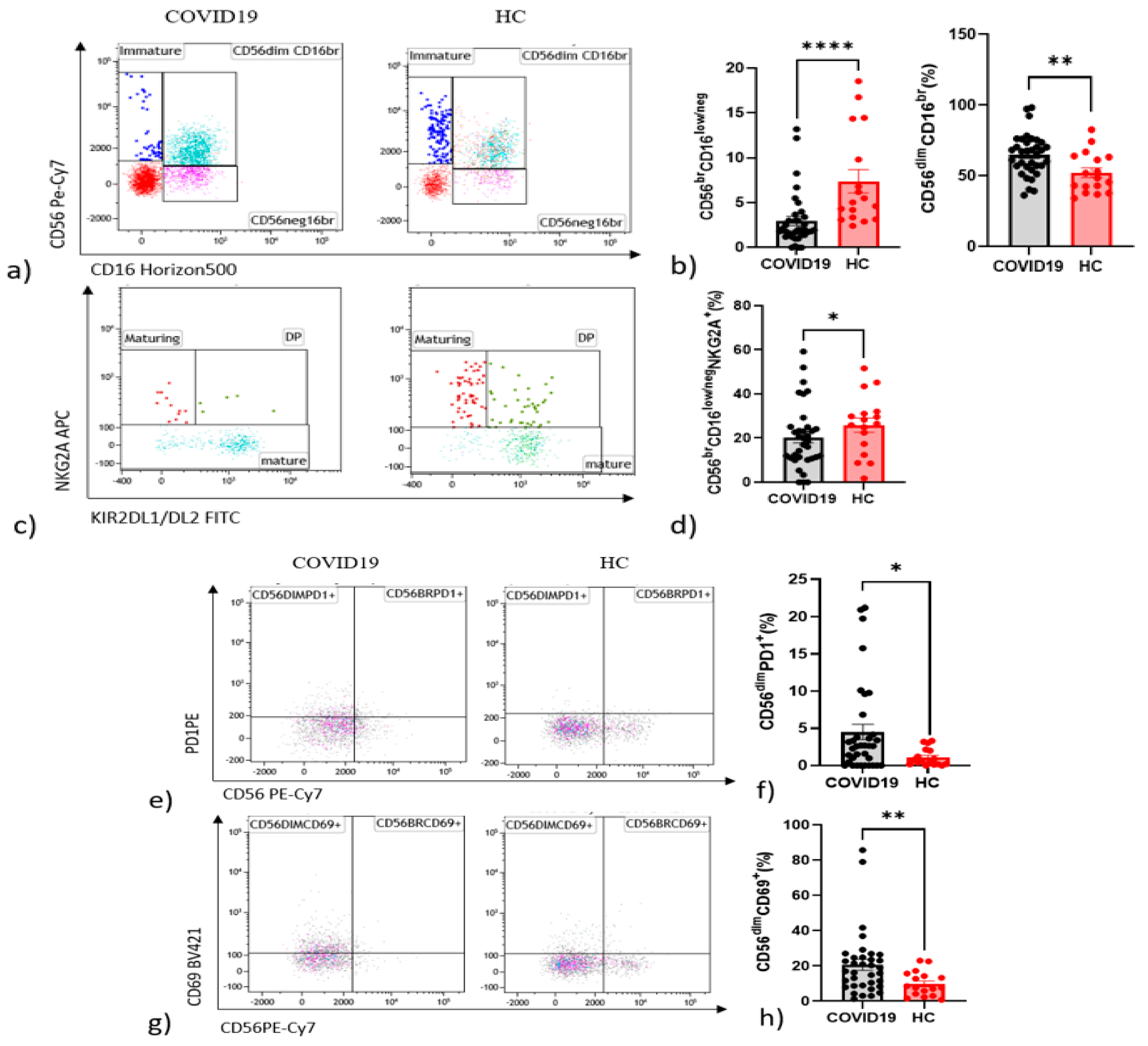
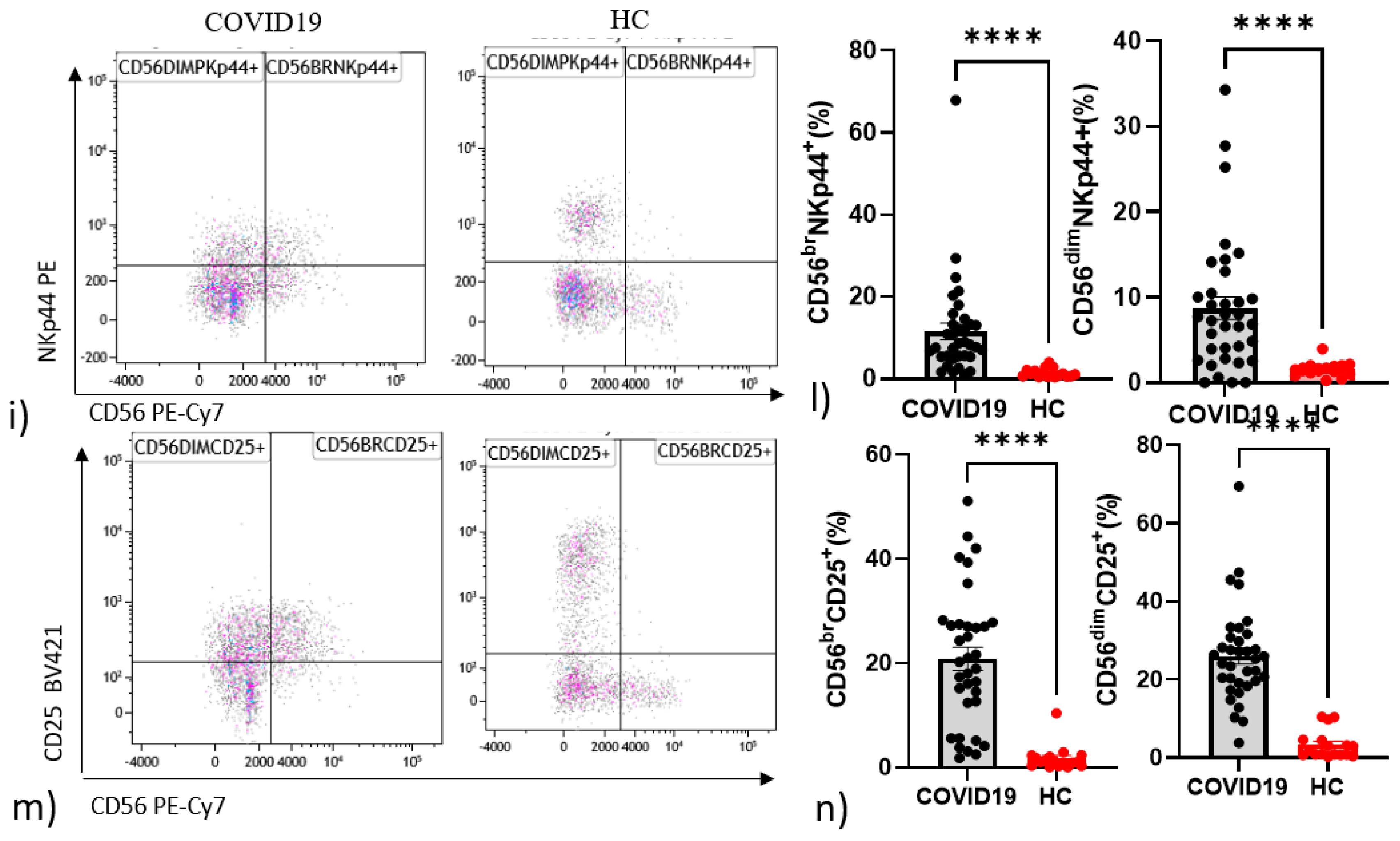
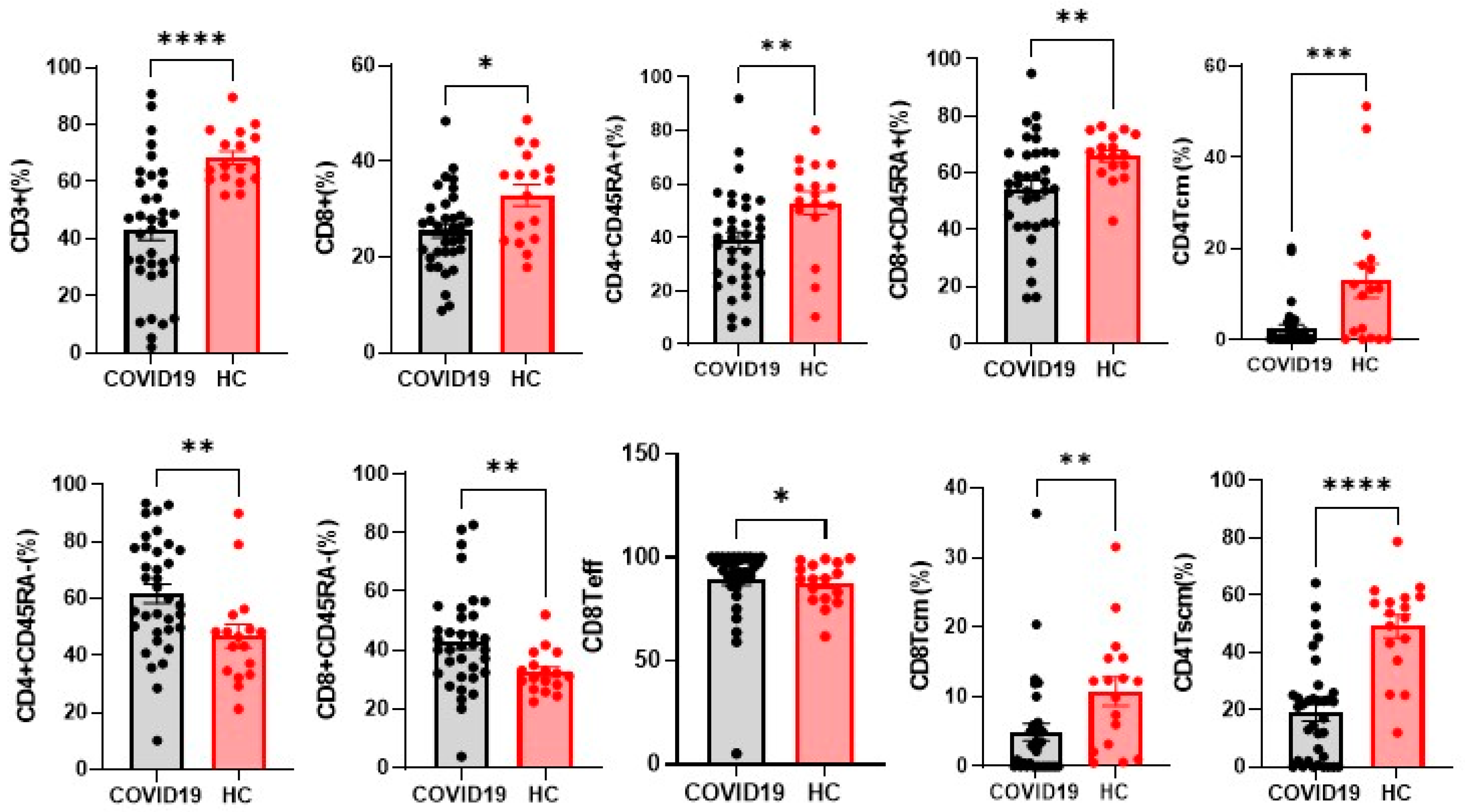
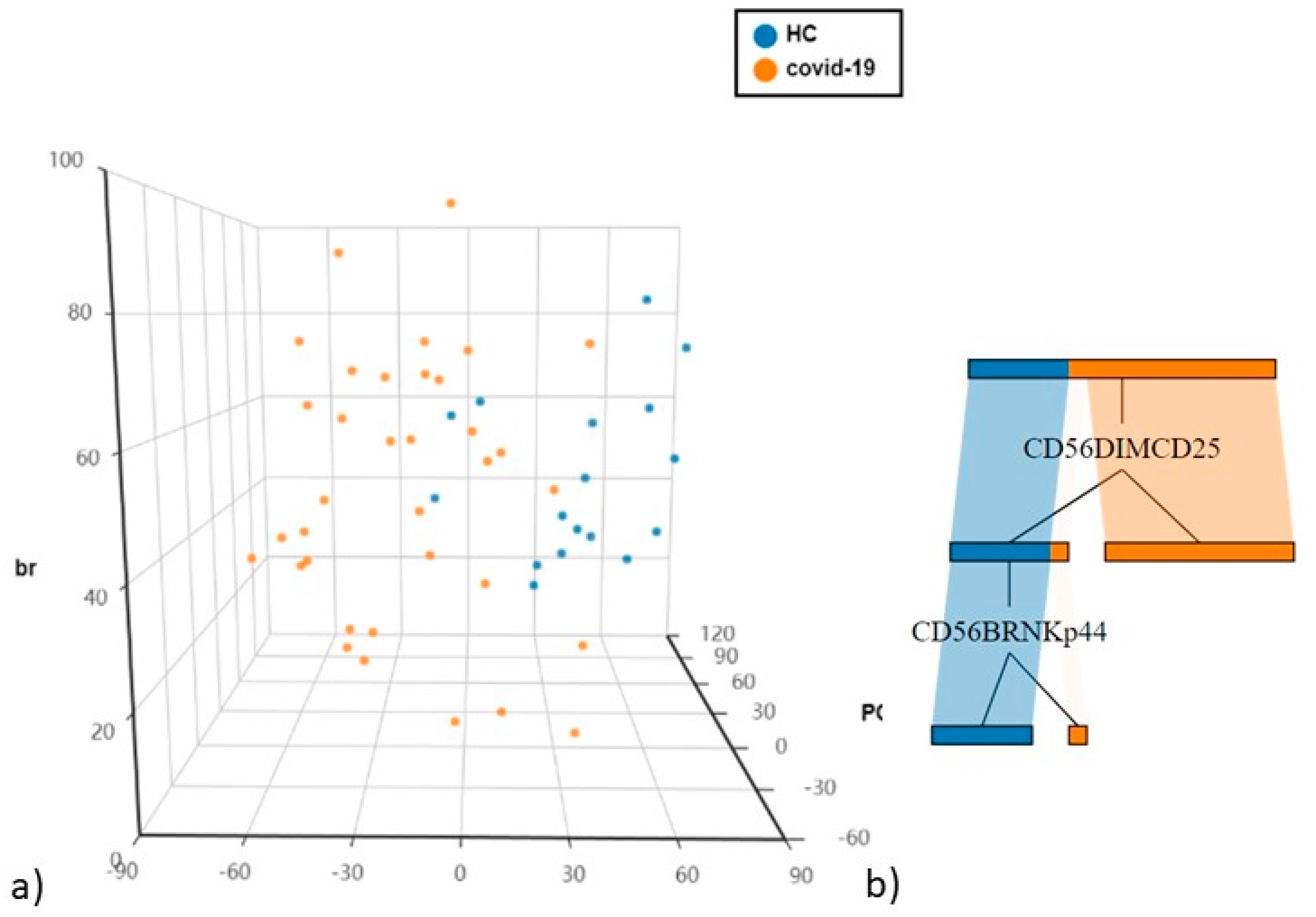
4.2. Differences in NK and T Cells in Hospitalised Patients with COVID-19 and Controls
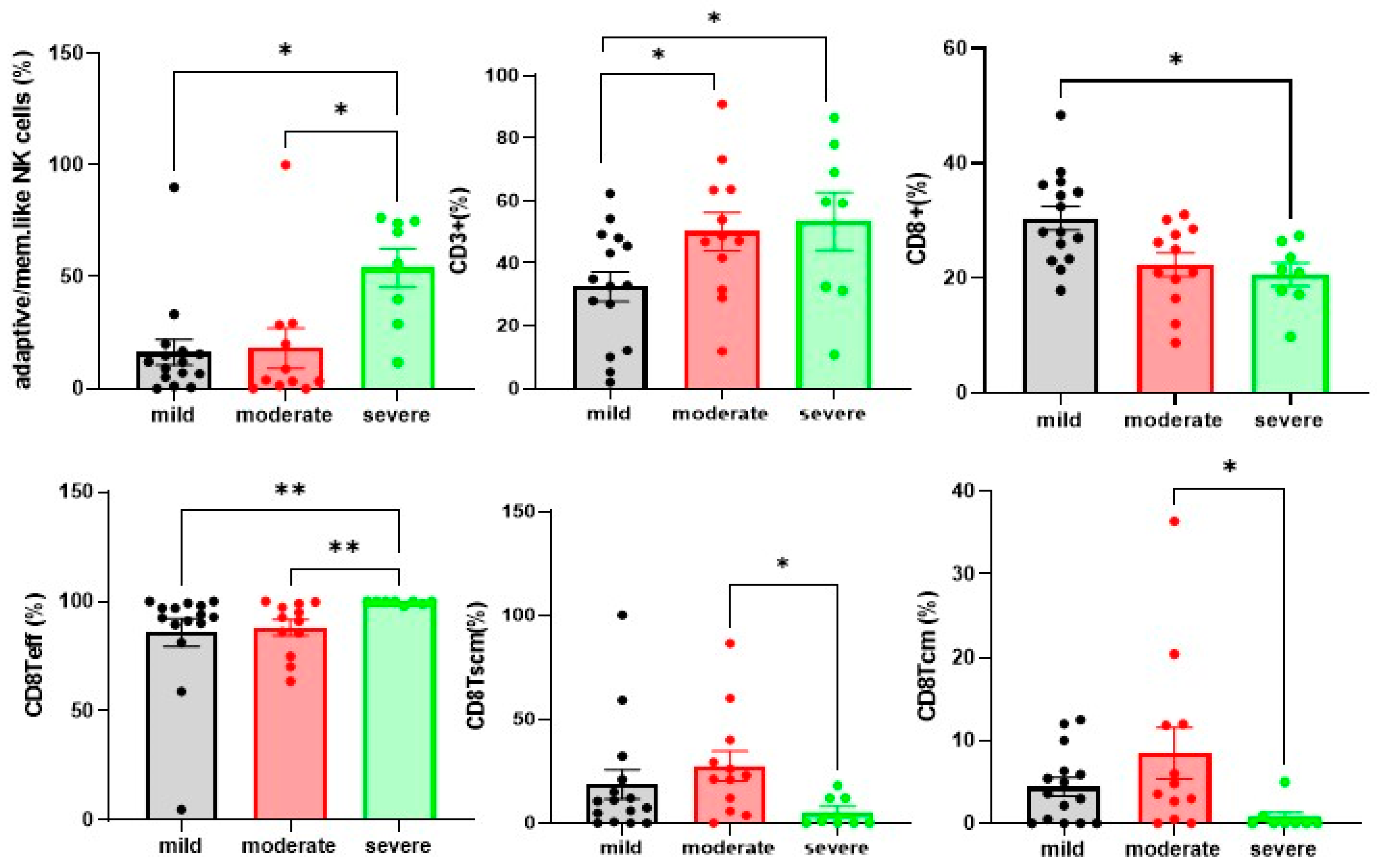
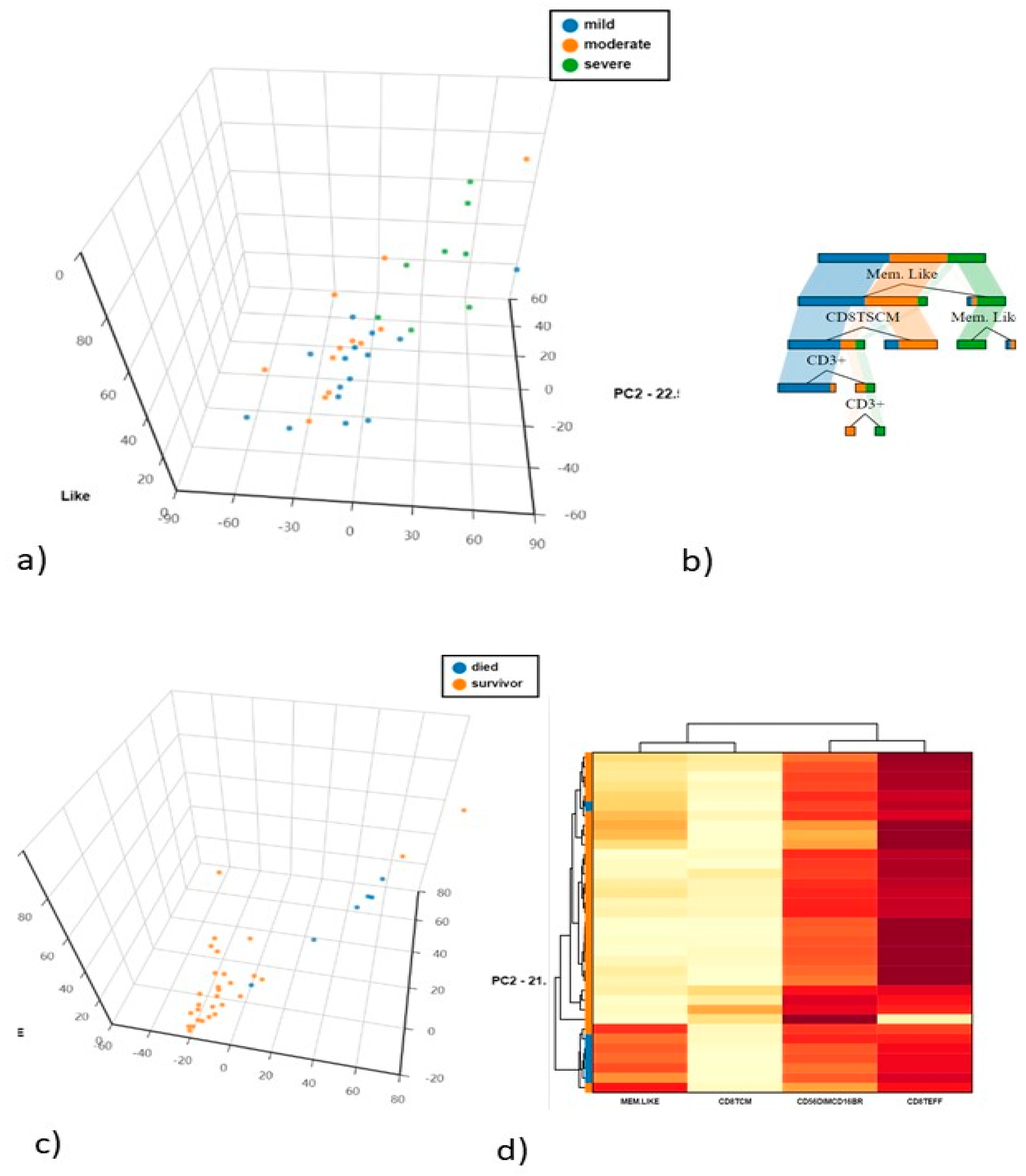
4.3. Differences in NK and T Cells in Mild. Moderate and Severe Hospitalised Patients with COVID-19
4.4. Differences in NK and T Cells in Hospitalised Patients in Relation to Death and Survival
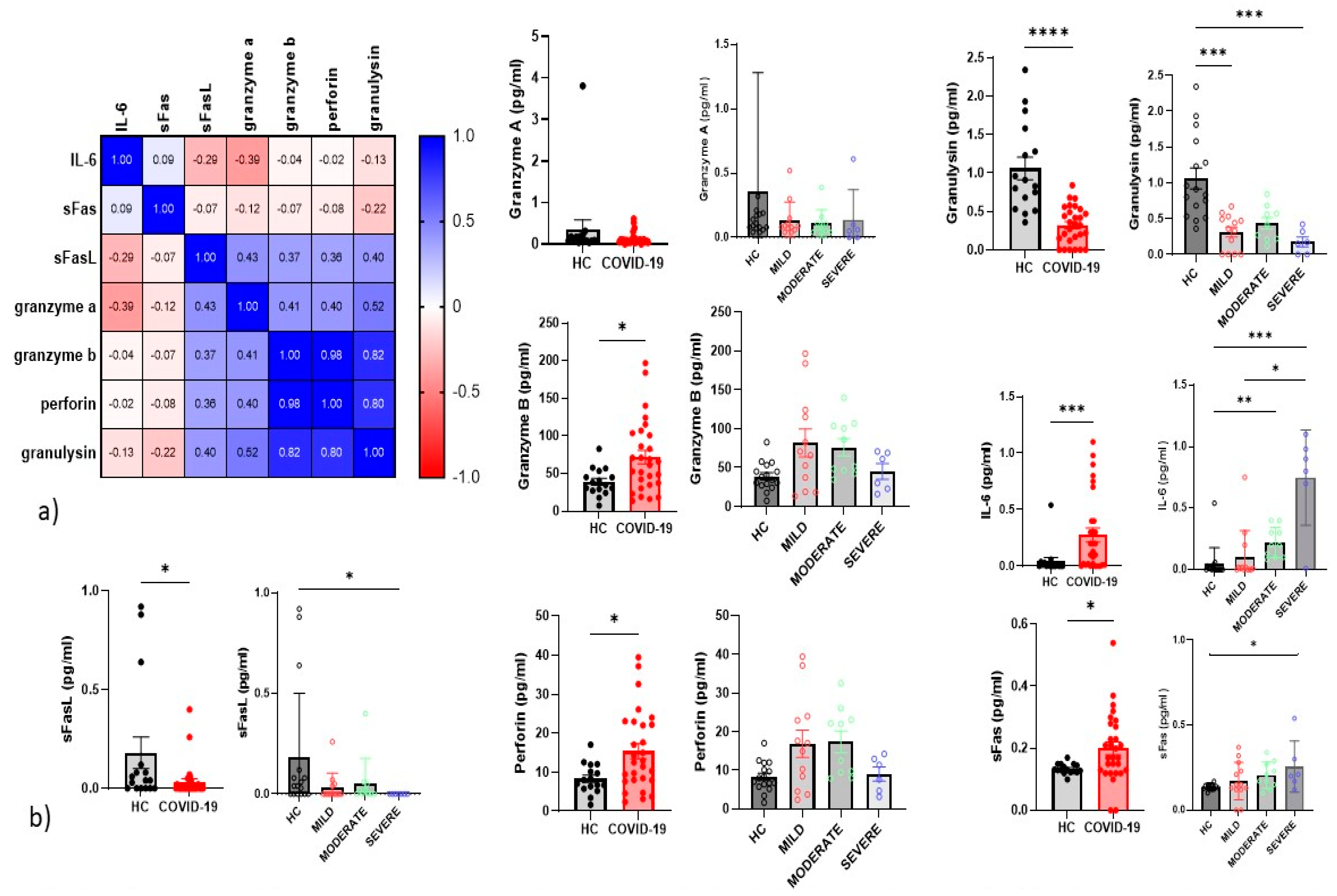
4.5. Serum Inflammatory Cytokines
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bergantini, L.; Bargagli, E.; d’Alessandro, M.; Refini, R.M.; Cameli, P.; Galasso, L.; Scapellato, C.; Montagnani, F.; Scolletta, S.; Franchi, F.; et al. Prognostic bioindicators in severe COVID-19 patients. Cytokine 2021, 141, 155455. [Google Scholar] [CrossRef]
- Maucourant, C.; Filipovic, I.; Ponzetta, A.; Aleman, S.; Cornillet, M.; Hertwig, L.; Strunz, B.; Lentini, A.; Reinius, B.; Brownlie, D.; et al. Natural Killer Cell Activation Related to Clinical Outcome of COVID-19. 2020. Available online: https://www.medrxiv.org/content/10.1101/2020.07.07.20148478v1 (accessed on 15 October 2021).
- Guan, W.; Ni, Z.; Hu, Y.; Liang, W.; Ou, C.; He, J.; Liu, L.; Shan, H.; Lei, C.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020. Available online: https://www.nejm.org/doi/10.1056/nejmoa2002032 (accessed on 15 October 2021).
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider cytokine storm syndromes and immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.-E.; Katsaounou, P.; et al. Complex Immune Dysregulation in COVID-19 Patients with Severe Respiratory Failure. Cell Host Microbe 2020, 27, 992–1000.e3. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameli, P.; Bergantini, L.; d’Alessandro, M.; Bargagli, E. High-dose steroids for the treatment of severe COVID-19: A new therapeutic tool? Intern. Emerg. Med. 2021, 16, 1329–1330. [Google Scholar] [CrossRef] [PubMed]
- d’Alessandro, M.; Cameli, P.; Refini, R.M.; Bergantini, L.; Alonzi, V.; Lanzarone, N.; Bennett, D.; Rana, G.D.; Montagnani, F.; Scolletta, S.; et al. Serum KL-6 concentrations as a novel biomarker of severe COVID-19. J. Med. Virol. 2020, 92, 2216–2220. [Google Scholar] [CrossRef]
- Ghasemzadeh, M.; Ghasemzadeh, A.; Hosseini, E. Exhausted NK cells and cytokine storms in COVID-19: Whether NK celtherapy could be a therapeutic choice. Hum. Immunol. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Hervier, B.; Russick, J.; Cremer, I.; Vieillard, V. NK Cells in the Human Lungs. Front. Immunol. 2019, 10, 1263. [Google Scholar] [CrossRef] [Green Version]
- Del Zotto, G.; Antonini, F.; Pesce, S.; Moretta, F.; Moretta, L.; Marcenaro, E. Comprehensive Phenotyping of Human PB NK Cells by Flow Cytometry. Cytometry A 2020, 97, 891–899. [Google Scholar] [CrossRef]
- Bergantini, L.; d’Alessandro, M.; Cameli, P.; Otranto, A.; Finco, T.; Curatola, G.; Sestini, P.; Bargagli, E. Prognostic role of NK cell percentages in bronchoalveolar lavage from patients with different fibrotic interstitial lung diseases. Clin. Immunol. 2021, 230, 108827. [Google Scholar] [CrossRef]
- Bergantini, L.; Cameli, P.; d’Alessandro, M.; Vagaggini, C.; Refini, R.M.; Landi, C.; Pieroni, M.G.; Sestini, P.; Bargagli, E. NK and NKT-like cells in granulomatous and fibrotic lung diseases. Clin. Exp. Med. 2019, 19, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Balato, A.; Unutmaz, D.; Gaspari, A.A. Natural killer T cells: An unconventional T-cell subset with diverse effector and regulatory functions. J. Investig. Dermatol. 2009, 129, 1628–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitale, M.; Bottino, C.; Sivori, S.; Sanseverino, L.; Castriconi, R.; Marcenaro, E.; Augugliaro, R.; Moretta, L.; Moretta, A. NKp44, a novel triggering surface molecule specifically expressed by activated natural killer cells, is involved in non-major histocompatibility complex-restricted tumor cell lysis. J. Exp. Med. 1998, 187, 2065–2072. [Google Scholar] [CrossRef]
- Pesce, S.; Belgrano, V.; Greppi, M.; Carlomagno, S.; Squillario, M.; Barla, A.; Della Chiesa, M.; Di Domenico, S.; Mavilio, D.; Moretta, L.; et al. Different Features of Tumor-Associated NK Cells in Patients with Low-Grade or High-Grade Peritoneal Carcinomatosis. Front. Immunol. 2019, 10, 1963. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.C.; Lanier, L.L. NK cell development, homeostasis and function: Parallels with CD8+ T cells. Nat. Rev. Immunol. 2011, 11, 645–657. [Google Scholar] [CrossRef]
- Brandt, C.S.; Baratin, M.; Yi, E.C.; Kennedy, J.; Gao, Z.; Fox, B.; Haldeman, B.; Ostrander, C.D.; Kaifu, T.; Chabannon, C.; et al. The B7 family member B7-H6 is a tumor cell ligand for the activating natural killer cell receptor NKp30 in humans. J. Exp. Med. 2009, 206, 1495–1503. [Google Scholar] [CrossRef] [Green Version]
- Gaggero, S.; Bruschi, M.; Petretto, A.; Parodi, M.; Del Zotto, G.; Lavarello, C.; Prato, C.; Santucci, L.; Barbuto, A.; Bottino, C.; et al. Nidogen-1 is a novel extracellular ligand for the NKp44 activating receptor. Oncoimmunology 2018, 7, e1470730. [Google Scholar] [CrossRef]
- Narni-Mancinelli, E.; Gauthier, L.; Baratin, M.; Guia, S.; Fenis, A.; Deghmane, A.-E.; Rossi, B.; Fourquet, P.; Escalière, B.; Kerdiles, Y.M.; et al. Complement factor P is a ligand for the natural killer cell-activating receptor NKp46. Sci. Immunol. 2017, 2, eaam9628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- d’Alessandro, M.; Bennett, D.; Montagnani, F.; Cameli, P.; Perrone, A.; Bergantini, L.; Franchi, F.; Scolletta, S.; Mazzei, M.; Volterrani, L.; et al. Peripheral lymphocyte subset monitoring in COVID-19 Italian patients. Minerva Med. 2021, 112, 423–426. [Google Scholar] [CrossRef]
- Cook, K.D.; Waggoner, S.N.; Whitmire, J.K. NK cells and their ability to modulate T cells during virus infections. Crit. Rev. Immunol. 2014, 34, 359–388. [Google Scholar] [CrossRef] [PubMed]
- Barrow, A.D.; Edeling, M.A.; Trifonov, V.; Luo, J.; Goyal, P.; Bohl, B.; Bando, J.K.; Kim, A.H.; Walker, J.; Andahazy, M.; et al. Natural Killer Cells Control Tumor Growth by Sensing a Growth Factor. Cell 2018, 172, 534–548.e19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demaria, O.; Carvelli, J.; Batista, L.; Thibult, M.-L.; Morel, A.; André, P.; Morel, Y.; Vély, F.; Vivier, E. Identification of druggable inhibitory immune checkpoints on Natural Killer cells in COVID-19. Cell Mol. Immunol. 2020, 17, 995–997. [Google Scholar] [CrossRef] [PubMed]
- Daga, S.; Fallerini, C.; Baldassarri, M.; Fava, F.; Valentino, F.; Doddato, G.; Benetti, E.; Furini, S.; Giliberti, A.; Tita, R.; et al. Employing a systematic approach to biobanking and analyzing clinical and genetic data for advancing COVID-19 research. Eur. J. Hum. Genet. 2021, 29, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; d’Alessandro, M.; Cameli, P.; Pianigiani, T.; Fanetti, M.; Sestini, P.; Bargagli, E. Follicular T Helper and Breg Cell Balance in Severe Allergic Asthma Before and After Omalizumab Therapy. Mol. Diagn. Ther. 2021, 25, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; d’Alessandro, M.; Cameli, P.; Bono, C.; Perruzza, M.; Biagini, M.; Pini, L.; Bigliazzi, C.; Sestini, P.; Dotta, F.; et al. Regulatory T cell monitoring in severe eosinophilic asthma patients treated with mepolizumab. Scand. J. Immunol. 2021, 94, e13031. [Google Scholar] [CrossRef]
- Bergantini, L.; d’Alessandro, M.; Cameli, P.; Bianchi, F.; Sestini, P.; Bargagli, E.; Refini, R.M. Personalized Approach of Severe Eosinophilic Asthma Patients Treated with Mepolizumab and Benralizumab. IAA 2020, 181, 746–753. [Google Scholar] [CrossRef] [PubMed]
- d’Alessandro, M.; Bergantini, L.; Cameli, P.; Mezzasalma, F.; Refini, R.M.; Pieroni, M.; Sestini, P.; Bargagli, E. Adaptive immune system in pulmonary sarcoidosis-Comparison of peripheral and alveolar biomarkers. Clin. Exp. Immunol. 2021, 205, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Bergantini, L.; d’Alessandro, M.; De Vita, E.; Perillo, F.; Fossi, A.; Luzzi, L.; Paladini, P.; Perrone, A.; Rottoli, P.; Sestini, P.; et al. Regulatory and Effector Cell Disequilibrium in Patients with Acute Cellular Rejection and Chronic Lung Allograft Dysfunction after Lung Transplantation: Comparison of Peripheral and Alveolar Distribution. Cells 2021, 10, 780. [Google Scholar] [CrossRef]
- Maecker, H.T. Multiparameter Flow Cytometry Monitoring of T Cell Responses. Methods Mol. Biol. 2009, 485, 375–391. [Google Scholar]
- Breadth of Concomitant Immune Responses Prior to Patient Recovery: A Case Report of Non-Severe COVID-19. Nat. Med. 2020, 26, 453–455. Available online: https://www.nature.com/articles/s41591-020-0819-2 (accessed on 4 November 2021).
- Zhang, Y.; Wang, X.; Li, X.; Xi, D.; Mao, R.; Wu, X.; Cheng, S.; Sun, X.; Yi, C.; Ling, Z.; et al. Potential contribution of increased soluble IL-2R to lymphopenia in COVID-19 patients. Cell Mol. Immunol. 2020, 17, 878–880. [Google Scholar] [CrossRef]
- Xie, M.; Yunis, J.; Yao, Y.; Shi, J.; Yang, Y.; Zhou, P.; Liang, K.; Wan, Y.; Mehdi, A.; Chen, Z.; et al. High levels of soluble CD25 in COVID-19 severity suggest a divergence between anti-viral and pro-inflammatory T-cell responses. Clin. Transl. Immunol. 2021, 10, e1251. [Google Scholar] [CrossRef]
- Michel, T.; Poli, A.; Cuapio, A.; Briquemont, B.; Iserentant, G.; Ollert, M.; Zimmer, J. Human CD56bright NK Cells: An Update. J. Immunol. 2016, 196, 2923–2931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carson, W.E.; Fehniger, T.A.; Caligiuri, M.A. CD56bright natural killer cell subsets: Characterization of distinct functional responses to interleukin-2 and the c-kit ligand. Eur. J. Immunol. 1997, 27, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Guo, W.; Dong, Y.; Wang, X.; Dai, D.; Liu, X.; Wu, Y.; Li, M.; Zhang, W.; Zhou, H.; et al. Elevated Exhaustion Levels of NK and CD8+ T Cells as Indicators for Progression and Prognosis of COVID-19 Disease. Front. Immunol. 2020, 11, 2681. [Google Scholar] [CrossRef]
- Lorente, L.; Martín, M.M.; González-Rivero, A.F.; Pérez-Cejas, A.; Argueso, M.; Perez, A.; Ramos-Gómez, L.; Solé-Violán, J.; Marcos Y Ramos, J.A.; Ojeda, N.; et al. Blood concentrations of proapoptotic sFas and antiapoptotic Bcl2 and COVID-19 patient mortality. Expert Rev. Mol. Diagn. 2021, 21, 837–844. [Google Scholar] [CrossRef]
- Cifaldi, L.; Prencipe, G.; Caiello, I.; Bracaglia, C.; Locatelli, F.; De Benedetti, F.; Strippoli, R. Inhibition of natural killer cell cytotoxicity by interleukin-6: Implications for the pathogenesis of macrophage activation syndrome. Arthritis Rheumatol. 2015, 67, 3037–3046. [Google Scholar] [CrossRef]
- Mazzoni, A.; Salvati, L.; Maggi, L.; Capone, M.; Vanni, A.; Spinicci, M.; Mencarini, J.; Caporale, R.; Peruzzi, B.; Antonelli, A.; et al. Impaired immune cell cytotoxicity in severe COVID-19 is IL-6 dependent. J. Clin. Investig. 2020, 130, 4694–4703. [Google Scholar] [CrossRef] [PubMed]
- Borrego, F.; Robertson, M.J.; Ritz, J.; Peña, J.; Solana, R. CD69 is a stimulatory receptor for natural killer cell and its cytotoxic effect is blocked by CD94 inhibitory receptor. Immunology 1999, 97, 159–165. [Google Scholar] [CrossRef]
- Gumá, M.; Angulo, A.; Vilches, C.; Gómez-Lozano, N.; Malats, N.; López-Botet, M. Imprint of human cytomegalovirus infection on the NK cell receptor repertoire. Blood 2004, 104, 3664–3671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleimanian, S.; Yaghobi, R. Harnessing Memory NK Cell to Protect Against COVID-19. Front. Pharmacol. 2020, 11, 1309. [Google Scholar] [CrossRef]
- Maucourant, C.; Filipovic, I.; Ponzetta, A.; Aleman, S.; Cornillet, M.; Hertwig, L.; Strunz, B.; Lentini, A.; Reinius, B.; Brownlie, D.; et al. Natural killer cell immunotypes related to COVID-19 disease severity. Sci. Immunol. 2020, 5, eabd6832. [Google Scholar] [CrossRef]
- Le Balc’h, P.; Pinceaux, K.; Pronier, C.; Seguin, P.; Tadié, J.-M.; Reizine, F. Herpes simplex virus and cytomegalovirus reactivations among severe COVID-19 patients. Crit. Care 2020, 24, 530. [Google Scholar] [CrossRef]
- Wherry, E.J.; Kurachi, M. Molecular and cellular insights into T cell exhaustion. Nat. Rev. Immunol. 2015, 15, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Aghbash, P.S.; Eslami, N.; Shamekh, A.; Entezari-Maleki, T.; Baghi, H.B. SARS-CoV-2 infection: The role of PD-1/PD-L1 and CTLA-4 axis. Life Sci. 2021, 270, 119124. [Google Scholar] [CrossRef]
- McKinney, E.F.; Lee, J.C.; Jayne, D.R.W.; Lyons, P.A.; Smith, K.G.C. T-cell exhaustion, co-stimulation and clinical outcome in autoimmunity and infection. Nature 2015, 523, 612–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parodi, M.; Favoreel, H.; Candiano, G.; Gaggero, S.; Sivori, S.; Mingari, M.C.; Moretta, L.; Vitale, M.; Cantoni, C. NKp44-NKp44 Ligand Interactions in the Regulation of Natural Killer Cells and Other Innate Lymphoid Cells in Humans. Front. Immunol. 2019, 10, 719. [Google Scholar] [CrossRef] [PubMed]
- Scharenberg, M.; Vangeti, S.; Kekäläinen, E.; Bergman, P.; Al-Ameri, M.; Johansson, N.; Sondén, K.; Falck-Jones, S.; Färnert, A.; Ljunggren, H.-G.; et al. Influenza A Virus Infection Induces Hyperresponsiveness in Human Lung Tissue-Resident and Peripheral Blood NK Cells. Front. Immunol. 2019, 10, 1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jost, S.; Lucar, O.; Yoder, T.; Werner, A.; Ghofrani, J.; Reeves, R.K. Human influenza-specific adaptive NK cells mediate potent HLA-E-restricted responses against antigenically distinct influenza strains. J. Immunol. 2020, 204 (Suppl. S1), 148.5. [Google Scholar]
- Uzhachenko, R.V.; Shanker, A. CD8+ T Lymphocyte and NK Cell Network: Circuitry in the Cytotoxic Domain of Immunity. Front. Immunol. 2019, 10, 1906. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Wei, X.; Guan, J.; Qin, S.; Wang, Z.; Lu, H.; Qian, J.; Wu, L.; Chen, Y.; Chen, Y.; et al. COVID-19 pneumonia: CD8+ T and NK cells are decreased in number but compensatory increased in cytotoxic potential. Clin. Immunol. 2020, 218, 108516. [Google Scholar] [CrossRef]
- Khwatenge, C.N.; Pate, M.; Miller, L.C.; Sang, Y. Immunometabolic Dysregulation at the Intersection of Obesity and COVID-19. Front. Immunol. 2021, 12, 732913. [Google Scholar] [CrossRef] [PubMed]
- Ronderos, D.; Omar, A.M.S.; Abbas, H.; Makker, J.; Baiomi, A.; Sun, H.; Mantri, N.; Choi, Y.; Fortuzi, K.; Shin, D.; et al. Chronic hepatitis-C infection in COVID-19 patients is associated with in-hospital mortality. World J. Clin. Cases 2021, 9, 8749–8762. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC (n = 17) | COVID-19 (n = 35) | p Values | Mild (n = 15) | Moderate (n = 12) | Severe (n = 8) | p Values | |
|---|---|---|---|---|---|---|---|
| AGE (years) | 65.3 ± 13.7 | 68.2 ± 16.3 | ns | 70.8 ± 14 | 58 ± 7 | 71.5 ± 120 | ns |
| GENDER (f/m) | 6/11 | 7/28 | ns | 2/13 | 4/8 | 1/7 | ns |
| SMOKING HABITS (current/never/former) | 6/6/5 | 11/9/15 | ns | 3/4/8 | 4/3/5 | 4/2/2 | ns |
| Chest X-ray (monolateral/bilateral pneumoniae) | 10/25 | 5/10 | 4/8 | 1/7 | ns | ||
| Comorbidities: yes/no diabetes arterial hypertension other lung diseases cancer haematological disorders | 0/35 | 31/4 | 12/3 6 8 5 2 1 | 11/1 4 9 3 4 0 | 8/0 4 7 4 3 1 | ns | |
| CRP (mg/L) (median (IQR)) | 4 (2.2–9.9) | 3.8 (2.1–7.2) | 3.1(2–4.4) | 5.4(3.8–7.3) | 0.012 | ||
| Blood counts: RBC WBC PLT | 4.4 (4.1–4.8) 6 (4.1–7.7) 197.5 (158–244) | 4.2 (4.1–4.4) 6 (4.5–7.5) 197 (162–244) | 3.9 (3–5.1) 6.7 (4.5–8.7) 217 (190–247) | 3.5 (3–4.3) 5.3 (4.5–6.4) 190 (161–251) | ns ns ns | ||
| Leucocytes counts (%): Lymphocytes Neutrophils Eosinophils Monocytes | 15.9 (9.3–19.7) 77.6 (70.9–88.5) 0 (0–0.2) 5.3 (6.1–10.1) | 16.8 (9.5–28.9) 77.6 (56.5–80) 0 (0–0.4) 8.8 (4.1–12.8) | 17 (10.5–32.9) 78.7 (52.1–82) 0 (0–0.4) 8.8 (4.1–12.8) | 11.1 (7–14.6) 64 (48.3–83.2) 0 (0–0.6) 7.6 (5.1–11.1) | 0.002 0.03 ns ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergantini, L.; d'Alessandro, M.; Cameli, P.; Cavallaro, D.; Gangi, S.; Cekorja, B.; Sestini, P.; Bargagli, E. NK and T Cell Immunological Signatures in Hospitalized Patients with COVID-19. Cells 2021, 10, 3182. https://doi.org/10.3390/cells10113182
Bergantini L, d'Alessandro M, Cameli P, Cavallaro D, Gangi S, Cekorja B, Sestini P, Bargagli E. NK and T Cell Immunological Signatures in Hospitalized Patients with COVID-19. Cells. 2021; 10(11):3182. https://doi.org/10.3390/cells10113182
Chicago/Turabian StyleBergantini, Laura, Miriana d'Alessandro, Paolo Cameli, Dalila Cavallaro, Sara Gangi, Behar Cekorja, Piersante Sestini, and Elena Bargagli. 2021. "NK and T Cell Immunological Signatures in Hospitalized Patients with COVID-19" Cells 10, no. 11: 3182. https://doi.org/10.3390/cells10113182
APA StyleBergantini, L., d'Alessandro, M., Cameli, P., Cavallaro, D., Gangi, S., Cekorja, B., Sestini, P., & Bargagli, E. (2021). NK and T Cell Immunological Signatures in Hospitalized Patients with COVID-19. Cells, 10(11), 3182. https://doi.org/10.3390/cells10113182