
Disruption of PAK3 Signaling in Social Interaction Induced cFos Positive Cells Impairs Social Recognition Memory


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Doxycycline Administration
2.2. Behavioral Tests
2.3. Immunohistochemistry and Fluorescent Imaging
2.4. Statistical Analyses
3. Results
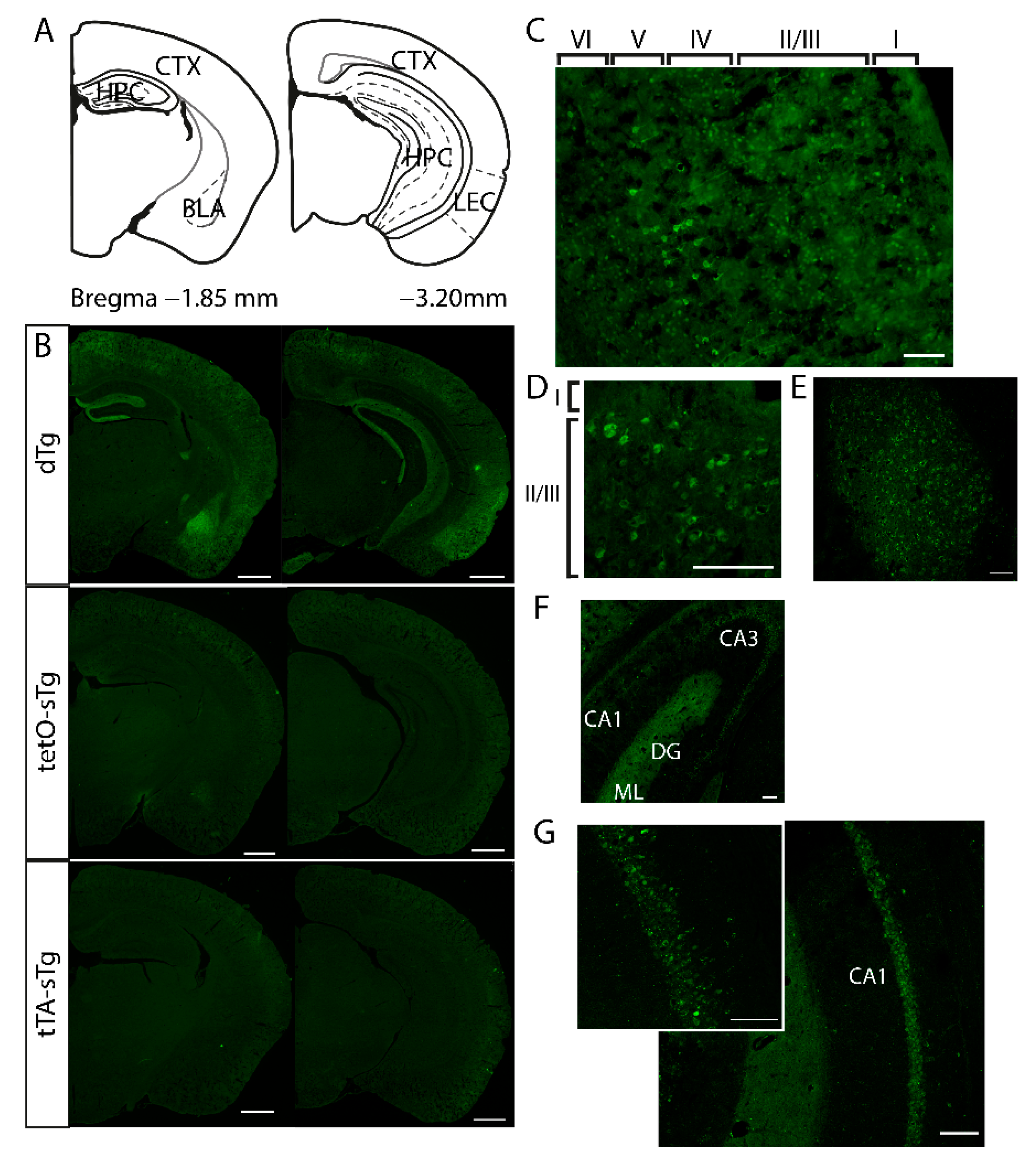
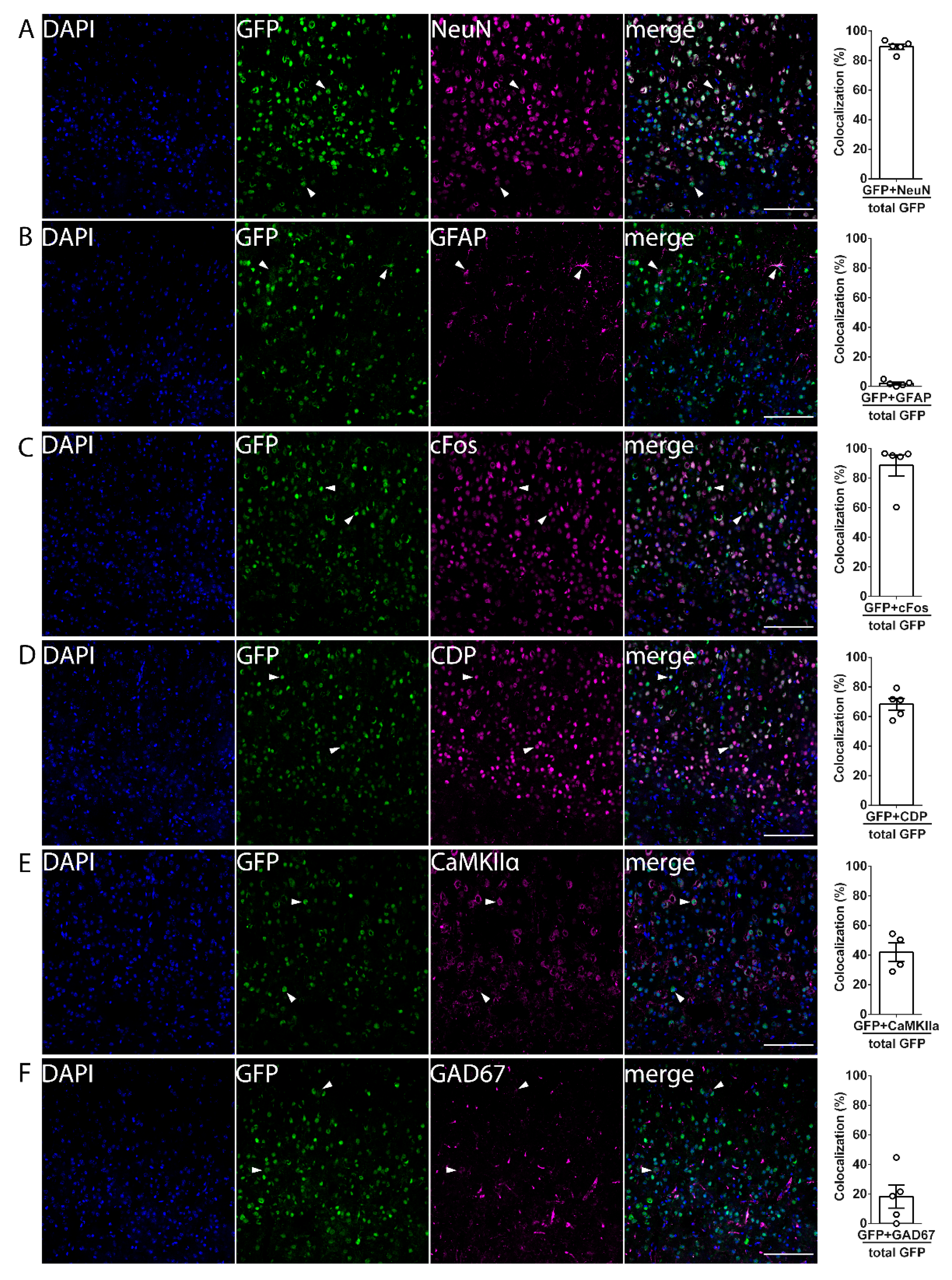
3.1. mPAK3-GFP Transgene Is Expressed in cFos-Expressing Excitatory and Inhibitory Neurons under Basal Conditions
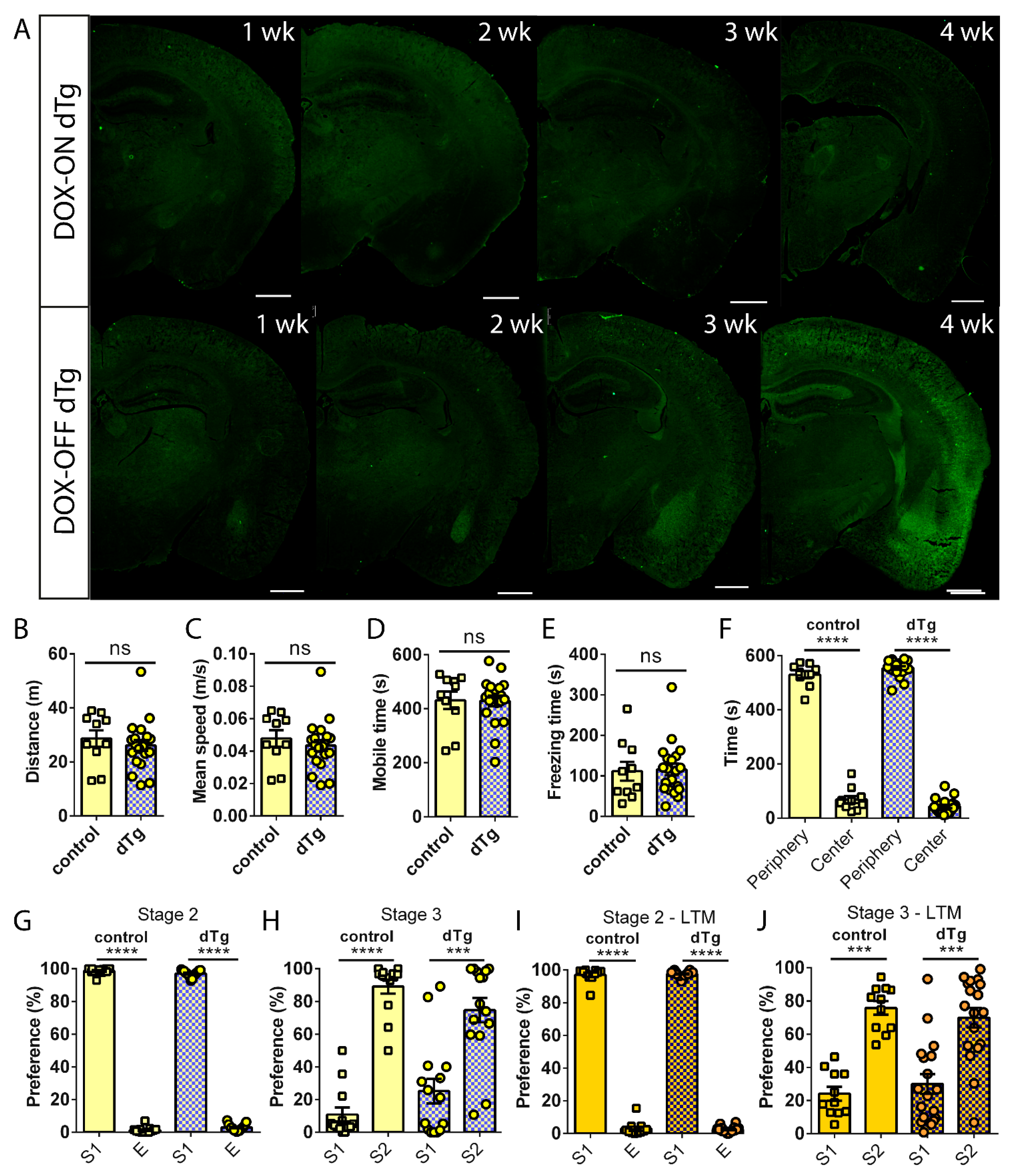
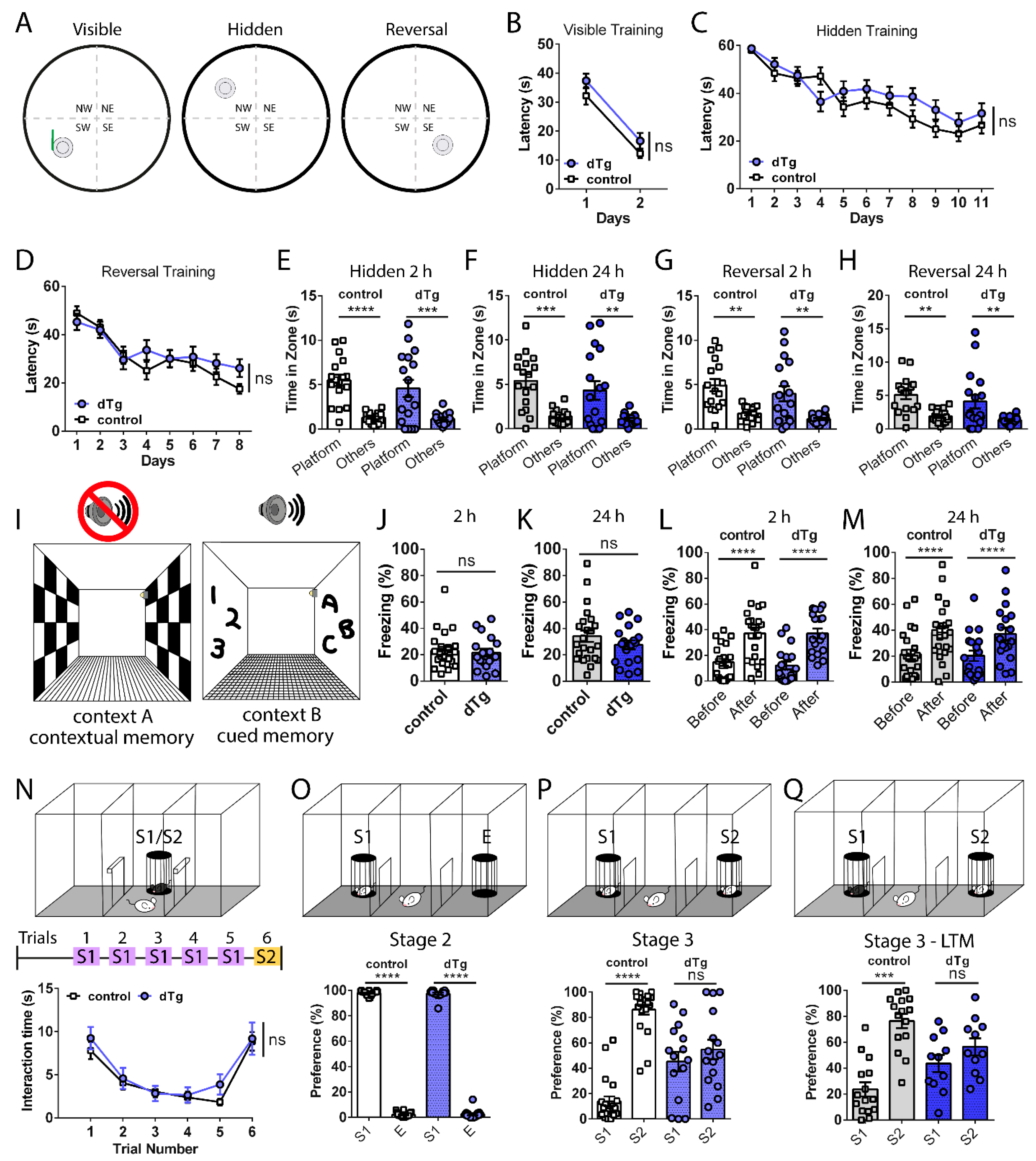
3.2. Basal Expression of mPAK3-GFP Proteins in cFos-Positive Cells Impairs Social Recognition Memory
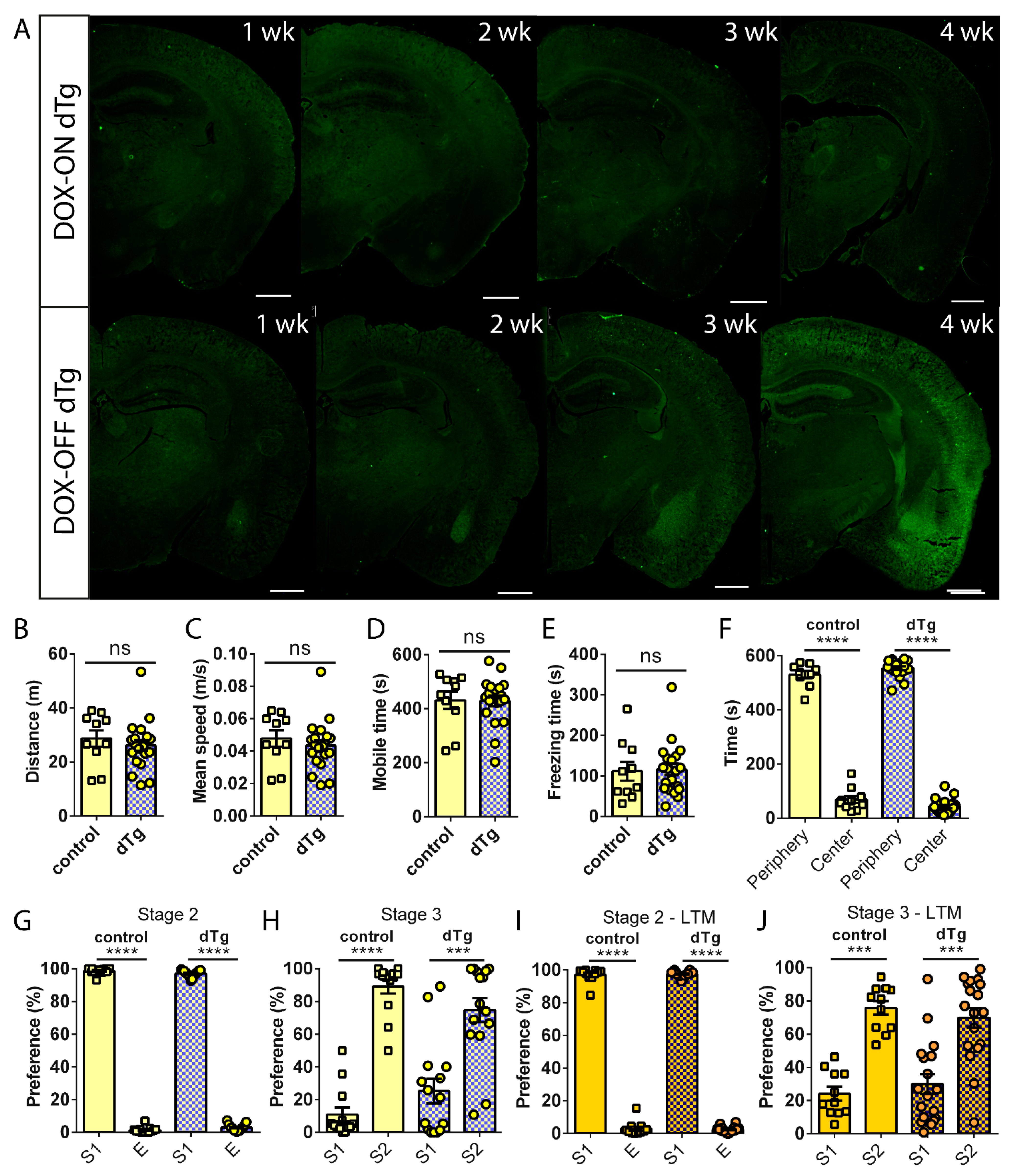
3.3. Administration of DOX Reverses Behavioral Deficits Seen in dTg Animals
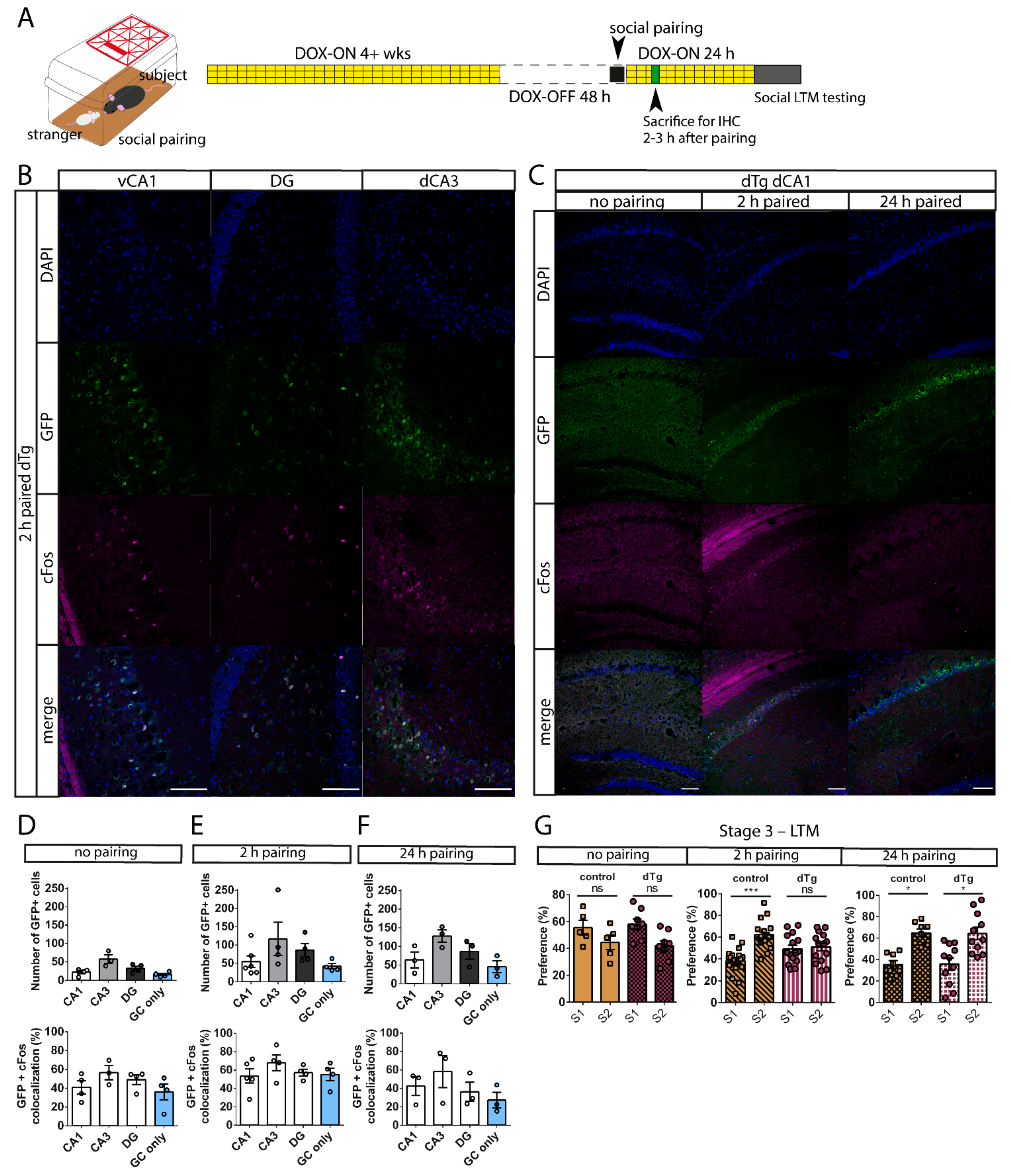
3.4. dTg Animals Can Be Used for a Social Pairing Protocol to Label cFos-Expressing Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartup, W.W. Social Relationships and Their Developmental Significance. Am. Psychol. 1989, 44, 120–126. [Google Scholar] [CrossRef]
- Cohen, S. Social Relationships and Health. Am. Psychol. 2004, 59, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Boen, C.; Gerken, K.; Li, T.; Schorpp, K.; Harris, K.M. Social Relationships and Physiological Determinants of Longevity across the Human Life Span. Proc. Natl. Acad. Sci. USA 2016, 113, 578–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijker, A.; Van Alphen, L.; Bos, A.; van den Borne, B.; Curfs, L. Social Integration of People with Intellectual Disability: Insights from a Social Psychological Research Programme. J. Intellect. Disabil. Res. 2011, 55, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Green, M.F.; Horan, W.P.; Lee, J. Social Cognition in Schizophrenia. Nat. Rev. Neurosci. 2015, 16, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Happé, F.; Cook, J.L.; Bird, G. The Structure of Social Cognition: In(Ter)Dependence of Sociocognitive Processes. Annu. Rev. Psychol. 2017, 68, 243–267. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, L.; Ayub, M.; Vincent, J.B. The Genetic Basis of Non-Syndromic Intellectual Disability: A Review. J. Neurodev. Disord. 2010, 2, 182–209. [Google Scholar] [CrossRef] [Green Version]
- Tejada, M.I.; Ibarluzea, N. Non-Syndromic X Linked Intellectual Disability: Current Knowledge in Light of the Recent Advances in Molecular and Functional Studies. Clin. Genet. 2020, 97, 677–687. [Google Scholar] [CrossRef]
- Allen, K.M.; Gleeson, J.G.; Bagrodia, S.; Partington, M.W.; MacMillan, J.C.; Cerione, R.A.; Mulley, J.C.; Walsh, C.A. PAK3 Mutation in Nonsyndromic X-Linked Mental Retardation. Nat. Genet. 1998, 20, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Bienvenu, T.; Des Portes, V.; McDonell, N.; Carrié, A.; Zemni, R.; Couvert, P.; Ropers, H.H.; Moraine, C.; Van Bokhoven, H.; Fryns, J.P.; et al. Missense Mutation in PAK3, R67C, Causes X-Linked Nonspecific Mental Retardation. Am. J. Med. Genet. 2000, 93, 294–298. [Google Scholar] [CrossRef] [Green Version]
- Magini, P.; Pippucci, T.; Tsai, I.C.; Coppola, S.; Stellacci, E.; Bartoletti-Stella, A.; Turchetti, D.; Graziano, C.; Cenacchi, G.; Neri, I.; et al. A Mutation in PAK3 with a Dual Molecular Effect Deregulates the RAS/MAPK Pathway and Drives an X-Linked Syndromic Phenotype. Hum. Mol. Genet. 2014, 23, 3607–3617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.L.; Choi, S.Y.; Shankaranarayana Rao, B.S.; Jung, H.Y.; Lee, H.K.; Zhang, D.; Chattarji, S.; Kirkwood, A.; Tonegawa, S. Altered Cortical Synaptic Morphology and Impaired Memory Consolidation in Forebrain-Specific Dominant-Negative PAK Transgenic Mice. Neuron 2004, 42, 773–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, J.; Meng, Y.; Hanna, A.; Janus, C.; Jia, Z. Abnormal Long-Lasting Synaptic Plasticity and Cognition in Mice Lacking the Mental Retardation Gene Pak3. J. Neurosci. 2005, 25, 6641–6650. [Google Scholar] [CrossRef] [Green Version]
- Kreis, P.; Thévenot, E.; Rousseau, V.; Boda, B.; Muller, D.; Barnier, J.-V. The P21-Activated Kinase 3 Implicated in Mental Retardation Regulates Spine Morphogenesis Through a Cdc42-Dependent Pathway. J. Biol. Chem. 2007, 282, 21497–21506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillon, C.; Gonzalez, L.; Domenichini, F.; Guyon, S.; Da Silva, K.; Durand, C.; Lestaevel, P.; Vaillend, C.; Laroche, S.; Barnier, J.-V.; et al. The Intellectual Disability PAK3 R67C Mutation Impacts Cognitive Functions and Adult Hippocampal Neurogenesis. Hum. Mol. Genet. 2020, 29, 1950–1968. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhou, Z.; Asrar, S.; Henkelman, M.; Xie, W.; Jia, Z. P21-Activated Kinases 1 and 3 Control Brain Size through Coordinating Neuronal Complexity and Synaptic Properties. Mol. Cell. Biol. 2011, 31, 388–403. [Google Scholar] [CrossRef] [Green Version]
- Leung, C.; Cao, F.; Nguyen, R.; Iii, O.C.S.; Kim, J.C.; Jia, Z. Activation of Entorhinal Cortical Projections to the Dentate Gyrus Underlies Social Memory Retrieval Article Activation of Entorhinal Cortical Projections to the Dentate Gyrus Underlies Social Memory Retrieval. Cell Rep. 2018, 23, 2379–2391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagrodia, S.; Bailey, D.; Lenard, Z.; Hart, M.; Guan, J.L.; Premont, R.T.; Taylor, S.J.; Cerione, R.A. A Tyrosine-Phosphorylated Protein That Binds to an Important Regulatory Region on the Cool Family of P21-Activated Kinase-Binding Proteins. J. Biol. Chem. 1999, 274, 22393–22400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, N.K.; Thomas, G.M.; Luo, J.; Huganir, R.L. Regulation of AMPA Receptor Subunit GluA1 Surface Expression by PAK3 Phosphorylation. Proc. Natl. Acad. Sci. USA 2015, 112, 5883–5890. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Sanawar, R.; Li, X.; Li, F. Structure, Biochemistry, and Biology of PAK Kinases. Gene 2017, 605, 20–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manser, E.; Chong, C.; Zhao, Z.S.; Leung, T.; Michael, G.; Hall, C.; Lim, L. Molecular Cloning of a New Member of the P21-Cdc42/Rac-Activated Kinase (PAK) Family. J. Biol. Chem. 1995, 270, 25070–25078. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; King, A.J.; Diaz, H.B.; Marshall, M.S. Regulation of the Protein Kinase Raf-1 by Oncogenic Ras through Phosphatidylinositol 3-Kinase, Cdc42/Rac and Pak. Curr. Biol. 2000, 10, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Thévenot, E.; Moreau, A.W.; Rousseau, V.; Combeau, G.; Domenichini, F.; Jacquet, C.; Goupille, O.; Amar, M.; Kreis, P.; Fossier, P.; et al. P21-Activated Kinase 3 (PAK3) Protein Regulates Synaptic Transmission through Its Interaction with the Nck2/Grb4 Protein Adaptor. J. Biol. Chem. 2011, 286, 40044–40059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, M.; Homma, K.; Saito, J.; Inoue, A.; Ikebe, R.; Ikebe, M. Dual Regulation of Mammalian Myosin VI Motor Function. J. Biol. Chem. 2001, 276, 39600–39607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combeau, G.; Kreis, P.; Domenichini, F.; Amar, M.; Fossier, P.; Rousseau, V.; Barnier, J.-V. The P21-Activated Kinase PAK3 Forms Heterodimers With PAK1 in Brain Implementing Trans-Regulation of PAK3 Activity. J. Biol. Chem. 2012, 287, 30084–30096. [Google Scholar] [CrossRef] [Green Version]
- Boda, B.; Jourdain, L.; Muller, D. Distinct, but Compensatory Roles of PAK1 and PAK3 in Spine Morphogenesis. Hippocampus 2008, 18, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Boda, B.; Alberi, S.; Nikonenko, I.; Node-Langlois, R.; Jourdain, P.; Moosmayer, M.; Parisi-Jourdain, L.; Muller, D. The Mental Retardation Protein PAK3 Contributes to Synapse Formation and Plasticity in Hippocampus. J. Neurosci. 2004, 24, 10816–10825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreis, P.; Rousseau, V.; Thévenot, E.; Combeau, G.; Barnier, J.V. The Four Mammalian Splice Variants Encoded by the P21-Activated Kinase 3 Gene Have Different Biological Properties. J. Neurochem. 2008, 106, 1184–1197. [Google Scholar] [CrossRef]
- Hainmueller, T.; Bartos, M. Parallel Emergence of Stable and Dynamic Memory Engrams in the Hippocampus. Nature 2018, 558, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, T.; Ogawa, S.K.; Roy, D.S.; Okuyama, T.; Morrissey, M.D.; Smith, L.M.; Redondo, R.L.; Tonegawa, S. Engrams and Circuits Crucial for Systems Consolidation of a Memory. Science 2017, 356, 73–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asok, A.; Leroy, F.; Rayman, J.B.; Kandel, E.R. Molecular Mechanisms of the Memory Trace. Trends Neurosci. 2019, 42, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Nader, K.; Schafe, G.E.; Le Doux, J.E. Fear Memories Require Protein Synthesis in the Amygdala for Reconsolidation after Retrieval. Nature 2000, 406, 722–726. [Google Scholar] [CrossRef] [PubMed]
- Tonegawa, S.; Pignatelli, M.; Roy, D.S.; Ryan, T.J. Memory Engram Storage and Retrieval. Curr. Opin. Neurobiol. 2015, 35, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, M.; Hayashi, Y. Catching the Engram: Strategies to Examine the Memory Trace. Mol. Brain 2012, 5, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayford, M. The Search for a Hippocampal Engram. Philos. Trans. R. Soc. B 2014, 369, 20130161. [Google Scholar] [CrossRef] [Green Version]
- Abraham, W.C.; Dragunow, M.; Tate, W.P. The Role of Immediate Early Genes in the Stabilization of Long-Term Potentiation. Mol. Neurobiol. 1991, 5, 297–314. [Google Scholar] [CrossRef]
- Hoffman, G.E.; Smith, M.S.; Verbalis, J.G. C-Fos and Related Immediate Early Gene Products as Markers of Activity in Neuroendocrine Systems. Front. Neuroendocrinol. 1993, 14, 173–213. [Google Scholar] [CrossRef]
- Sauvage, M.M.; Nakamura, N.H.; Beer, Z. Mapping Memory Function in the Medial Temporal Lobe with the Immediate-Early Gene Arc. Behav. Brain Res. 2013, 254, 22–33. [Google Scholar] [CrossRef]
- Garner, A.R.; Rowland, D.C.; Hwang, S.Y.; Baumgaertel, K.; Roth, B.L.; Kentros, C.; Mayford, M. Generation of a Synthetic Memory Trace. Science 2012, 335, 1513–1516. [Google Scholar] [CrossRef] [Green Version]
- Reijmers, L.G.; Perkins, B.L.; Matsuo, N.; Mayford, M. Localization of a Stable Neural Correlate of Associative Memory. Science 2007, 317, 1230–1234. [Google Scholar] [CrossRef] [Green Version]
- Cao, F.; Liu, J.J.; Zhou, S.; Cortez, M.A.; Snead, O.C.; Han, J.; Jia, Z. Neuroligin 2 Regulates Absence Seizures and Behavioral Arrests through GABAergic Transmission within the Thalamocortical Circuitry. Nat. Commun. 2020, 11, 3744. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Crawley, J.N. Simple Behavioral Assessment of Mouse Olfaction. Curr. Protoc. Neurosci. 2009, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Vorhees, C.V.; Williams, M.T. Morris Water Maze: Procedures for Assessing Spatial and Related Forms of Learning and Memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Liu, A.; Xia, S.; Leung, C.; Qi, J.; Meng, Y.; Xie, W.; Park, P.; Collingridge, G.L.; Jia, Z. The C-Terminal Tails of Endogenous GluA1 and GluA2 Differentially Contribute to Hippocampal Synaptic Plasticity and Learning. Nat. Neurosci. 2018, 21, 50–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curzon, P.; Rustay, N.R.; Browman, K.E. Cued and contextual fear conditioning for rodents. In Methods of Behavior Analysis in Neuroscience, 2nd ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2000. [Google Scholar] [CrossRef]
- Neunuebel, J.P.; Knierim, J.J. Spatial Firing Correlates of Physiologically Distinct Cell Types of the Rat Dentate Gyrus. J. Neurosci. 2012, 32, 3848–3858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamantaki, M.; Frey, M.; Berens, P.; Preston-Ferrer, P.; Burgalossi, A. Sparse Activity of Identified Dentate Granule Cells during Spatial Exploration. Elife 2016, 5, e20252. [Google Scholar] [CrossRef]
- Baron, U.; Bujard, H. Tet Repressor-Based System for Regulated Gene Expression in Eukaryotic Cells: Principles and Advances. Methods Enzymol. 2000, 327, 401–421. [Google Scholar] [CrossRef]
- Delerue, F.; White, M.; Ittner, L.M. Inducible, Tightly Regulated and Non-Leaky Neuronal Gene Expression in Mice. Transgenic Res. 2014, 23, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Jung, M.W. Neural Circuits and Mechanisms Involved in Pavlovian Fear Conditioning: A Critical Review. Neurosci. Biobehav. Rev. 2006, 30, 188–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, D.S.; Kitamura, T.; Okuyama, T.; Ogawa, S.K.; Sun, C.; Obata, Y.; Yoshiki, A.; Tonegawa, S. Distinct Neural Circuits for the Formation and Retrieval of Episodic Memories. Cell 2017, 170, 1000–1012.e19. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, T.; Kitamura, T.; Roy, D.S.; Itohara, S.; Tonegawa, S. Ventral CA1 Neurons Store Social Memory. Science 2016, 353, 1536–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Jia, Z. Disruption of PAK3 Signaling in Social Interaction Induced cFos Positive Cells Impairs Social Recognition Memory. Cells 2021, 10, 3010. https://doi.org/10.3390/cells10113010
Zhou S, Jia Z. Disruption of PAK3 Signaling in Social Interaction Induced cFos Positive Cells Impairs Social Recognition Memory. Cells. 2021; 10(11):3010. https://doi.org/10.3390/cells10113010
Chicago/Turabian StyleZhou, Susan, and Zhengping Jia. 2021. "Disruption of PAK3 Signaling in Social Interaction Induced cFos Positive Cells Impairs Social Recognition Memory" Cells 10, no. 11: 3010. https://doi.org/10.3390/cells10113010
APA StyleZhou, S., & Jia, Z. (2021). Disruption of PAK3 Signaling in Social Interaction Induced cFos Positive Cells Impairs Social Recognition Memory. Cells, 10(11), 3010. https://doi.org/10.3390/cells10113010