Abstract
Different types of cells, such as endothelial cells, tumor-associated fibroblasts, pericytes, and immune cells, release extracellular vesicles (EVs) in the tumor microenvironment. The components of EVs include proteins, DNA, RNA, and microRNA. One of the most important functions of EVs is the transfer of aforementioned bioactive molecules, which in cancer cells may affect tumor growth, progression, angiogenesis, and metastatic spread. Furthermore, EVs affect the presentation of antigens to immune cells via the transfer of nucleic acids, peptides, and proteins to recipient cells. Recent studies have also explored the potential application of EVs in cancer treatment. This review summarizes the mechanisms by which EVs regulate melanoma development, progression, and their potentials to be applied in therapy. We initially describe vesicle components; discuss their effects on proliferation, anti-melanoma immunity, and drug resistance; and finally focus on the effects of EV-derived microRNAs on melanoma pathobiology. This work aims to facilitate our understanding of the influence of EVs on melanoma biology and initiate ideas for the development of novel therapeutic strategies.
1. Melanoma
Human skin is the first layer of defense, protecting us from external factors and providing control of body temperature and storage of moisture and fat. Skin cancer is a common malignancy, with three major types (basal cell carcinoma, squamous cell carcinoma, and melanoma), which have different precursor cells. Basal cell carcinoma and squamous cell carcinoma are classified as non-melanoma skin cancers []. Malignant melanoma which is derived from melanocytes is the most aggressive, invasive, and life-threatening skin cancer []. Risk factors for melanoma development include fair skin and exposure to sunlight while ultraviolet light exposure is one of the main causes of the development of melanoma []. According to the World Health Organization, approximately 132,000 new cases of melanoma are diagnosed each year worldwide. Especially in white (Caucasian) people, melanoma is becoming more common, mostly due to their less pigmented skin, which renders this population more susceptible to ultraviolet light exposure [].
Metastasis is a multistep process. The first step involves tumor cells invading the basal membrane and entering the blood vessels. Then, these tumor cells circulate in the blood stream until their attachment at the site of metastasis and initiate subsequent extravasation. Finally, they colonize and grow in distal host organs. In melanoma, vascular invasion occurs predominantly via lymphatic vessels. Vascular invasion is associated with factors indicative of a poor prognosis, including stage, increased Breslow thickness, and ulceration []. Cells can communicate by different types of EVs, which include exosomes, apoptotic bodies, and microvesicles (MVs) []. EVs are important mediators of intercellular communication between cells and distal organs and are crucial to cancer progression []. The role of EVs in melanoma remains unclear. Research focusing on understanding the functions and effects of EVs is essential to improve the treatment for melanoma patients and reduce the risk of melanoma metastasis. This review will help scientists understand more about the relationship between EVs and aggressive cancer and inspire in-depth research to make greater progress and breakthroughs.
2. Classification and Biology of Extracellular Vesicles
EVs can mediate intercellular communication during many cellular processes, and this role of EVs has piqued the interest of the scientific community. Evidence of the existence and functions of EVs was first collected in 1946 through a combination of ultracentrifugation, electron microscopy, and functional studies []. In 1970, the term “extracellular vesicle” was used in a manuscript title for the first time []. In the 1970s–1980s, several independent studies identified the release of plasma membrane vesicles from rectal adenoma microvillus cells [] and discovered virus-like particles in human cell cultures and bovine serum as preliminary findings of exosomes []. In 1983, detailed ultrastructural studies indicated that EVs are released through the fusion of multivesicular bodies (MVBs) with the cell membrane during immature red blood cell differentiation []. Since 2006, several reports have indicated that nucleic acids, proteins, and other molecules can be transferred between cells via EVs []. Through this shuttle-like mechanism, EVs modulate the activity of recipient cells and participate in various physiological and pathological processes, including tumor development, growth, progression, metastasis, and the development of drug resistance []. EVs express specific membrane proteins that facilitate EV interactions with particular recipient cells. This process was shown to be involved in organotropic metastatic spread []. Following these findings, researchers have isolated EVs from most cell types and biological fluids. The rapid development of the EV research community has been due to the establishment of the International Society of Extracellular Vesicles (ISEV) in the early 2000s, which conducted rigorous and standardized work in this area, including the establishment of the Journal of Extracellular Vesicles. Commercial investment in EV diagnosis and treatment has also increased, and many companies have developed several cancer diagnostic tests based on EVs.
Based on their size and biogenesis, EVs can be classified into exosomes, MVs, and apoptotic bodies (Figure 1). Exosomes and MVs can be released by normal cells or cancer cells, although they differ in several aspects. Exosomes are nanosized vesicles of endocytic origin that bud from MVBs toward the lumen of the compartment and are released into the extracellular space. Their size varies from 30 to 100 nm and is limited by the size of the MVBs (40–200 nm) []. The content of exosomes includes proteins, DNA, mRNA, and microRNA. In particular, Rab GTPases, soluble N-ethylmaleimide-sensitive factor activating protein receptors (SNAREs), Annexin, and Flotillin are enriched in EVs. Moreover, three transmembrane protein (CD9, CD63, and CD81) families are known to accumulate in the plasma membrane domain and are highly expressed in exosomes, thereby serving as biomarkers for exosomes [].
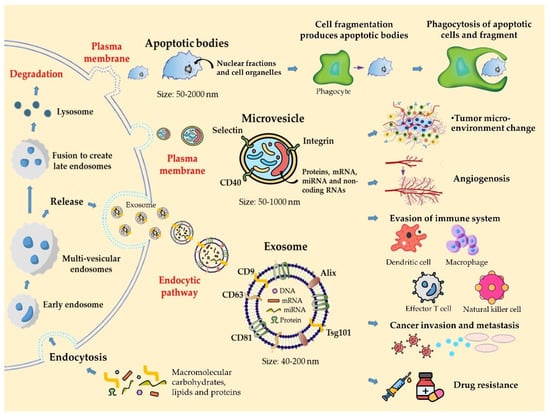
Figure 1.
Origin, classification, composition, and biological function of EVs.
EVs are known to facilitate intercellular communication between adjacent cells and distant cells [,]. EVs can be released by immune cells as antigen-presenting vesicles to stimulate antitumor immune responses or to induce carcinogenesis via suppressing inflammatory responses. EVs derived from tumor cells have also been shown to promote cancer cell proliferation by inactivating cytotoxic T lymphocytes (CTLs) or natural killer (NK) cells to suppress the immune response and promote regulatory CTL differentiation []. Within the nervous system, EVs are thought to be involved in the formation of myelin, the growth of neurites, and the survival of neurons []. In addition, several pathogenic proteins, such as viruses and β-amyloid peptides, have been reported to be transferred to other cells via EVs []. An important finding is that the roles of mRNAs and miRNAs in EVs from different sources are completely different from each other. Some studies have shown that EVs also circulate in various body fluids, including blood and urine, and their mRNAs and miRNAs can be transferred to recipient cells and participate in many biologically relevant processes, including immune response and angiogenesis [].
To investigate the characteristics of EVs, a purification protocol is required to be constructed. Early researchers widely used differential ultracentrifugation (DUC) as the method for EV isolation, with its extensive applicability, large capacity, ease of scaling up, and relatively high purification quality. By using DUC, large particles, such as whole cells, cell debris, subcellular structures, and other contaminants, can be removed under a low centrifugal speed. Thenceforth, scientists elevated the centrifugal force to precipitate EVs. Recently, it was verified that DUC outplays five commercial purification kits in terms of vesicle purity, which was a suitable approach for EV research up to the present. However, DUC-isolated EVs may suffer from the contaminants of other non-vesicular particles. Owing to the distinction in particle density between EVs and nano-contamination, density-gradient ultracentrifugation (DGUC) was applied to increase the purity of EVs. According to the minimal information from studies of extracellular vesicles 2018 (MISEV2018), several components, especially sucrose gradient solution, were applied as the medium for EV purification. Configuring the separation medium with gradient densities across the density range of EVs, crude isolated samples are loaded onto the top of the medium and undergo a longer ultracentrifugation period for further purification. Despite all of the advantages of DUC and DGUC, the pricey equipment and time-consuming process are a matter of concern. The centrifugal- and shearing-force-induced structural damage or aggregation also impede the downstream application. Nonetheless, the enhanced purity and quality of EVs provide a step forward in the research on nanoparticles [].
3. EVs Derived from Melanoma and Their Role in Cancer Progression
Biological information between adjacent tumor cells can be transmitted through tumor-derived EVs in a paracrine manner. This signal transduction between malignant cells not only promotes cancer growth and metastasis but also can interfere with normal signaling pathways []. Tumor cells may metastasize to distant organs in the body and regulate the tumor microenvironment to form pre-metastatic niches; in these cases, tumor-derived EVs may be potential biomarkers for tumor progression and invasion []. In addition, tumor-derived EVs are expected to be used as carriers for cell-free vaccines and for the delivery of specific tumor therapeutic molecules. In this section, we focus on the role of tumor-derived EVs in melanoma development and metastasis and their potential applications in advancing the diagnosis and treatment of melanoma and personalized medicine.
3.1. Growth and Angiogenesis
The literature indicates that the addition of EVs to a human cell culture enhances EV production and supports cell proliferation []. The biodistribution of cancer-derived EVs in tumor tissues is an important factor in determining the role of EVs in tumor proliferation []. In vivo experiments have shown that B16BL6 melanoma cells secrete and absorb B16BL6 cell-secreted EVs to induce their own proliferation and inhibit their own apoptosis, promoting tumor progression []. EV uptake by target cells relies on the integrity of plasma membrane microdomains, namely lipid rafts, which are known to be enriched with cholesterol. Scavenger receptor type-B1 (SR-B1) is a high-affinity receptor for mature high-density lipoproteins (HDLs), and SR-B1 maintains cholesterol equilibrium, uptakes extracellular material, and promotes cell signaling [,]. The expression of SR-B1 in melanoma enhances EV formation and cellular uptake, promoting a metastatic phenotype. SR-B1 is associated with the expression of microphthalmia-associated transcription factor (MITF) and the regulation of proto-oncogene mesenchymal-to-epithelial transition (MET) factor. SR-B1 is a key molecule for regulating EV uptake and cancer growth []. Wnt Family Member 5A (WNT5A) regulates the release of EVs containing the immunomodulatory cytokine IL-6 and proangiogenic factors IL-8, VEGF, and MMP2 from melanoma cells (MeWo, SKmel28, A2058, A375, and HTB63). This effect increases angiogenic processes and facilitates metastatic spread []. Hood et al. indicated that melanoma EVs can boost endothelial angiogenic responses to create a premetastatic niche []. A previous report indicated that miR-155 in melanoma-derived EVs can induce reprogramming of fibroblasts into CAFs (cancer-associate fibroblast) and trigger the proangiogenic switch of these CAFs [].
3.2. Migration and Invasion
Studies on melanoma cell migration and invasion and on the underlying molecular mechanisms are essential for improving melanoma diagnosis, prognosis, and therapy. EVs play an important role in this regard and regulate the migratory and invasive capacity of melanoma cells. Several studies have demonstrated that EVs can increase migratory and invasive capacities []. EVs derived from melanoma cells have also been shown to increase type I interferon receptor (IFNAR1) and cholesterol 25-hydroxylase (CH25H) in normal cells, thus facilitating EV uptake and pre-metastatic niche development []. Matrix metalloproteinases (ADAM) and ADAM with thrombospondin motifs (ADAMTS) are enriched in melanoma-derived EVs. These proteins are critical for degrading the extracellular matrix of cancer cells and increasing metastatic spread []. Insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) is a multifunctional RNA-binding protein that has been linked to the development of a variety of malignancies. According to previous research, EVs derived from IGF2BP1-overexpressing melanoma cells exacerbate EV-induced metastasis []. Xiao et al. showed an increase in invasiveness when normal melanocytes were treated with melanoma EVs []. Melanoma usually metastasizes to the lungs, bones, liver, and brain and rarely to other organs. The current mechanism of this pattern needs further understanding, but it is likely that EVs play an important role. For example, melanoma cells are exposed to bone-derived soluble factors, which are related to the molecular activation pathway of stromal-cell-derived factor 1 (SDF-1)/CXC chemokine receptor type 4 (CXCR4)/type 7 CXC chemokine receptor (CXCR7). To this end, EVs reprogram the innate osteotropism of melanoma cells by upregulating their CXCR7 expression []. These results suggest that melanoma-cell-derived EVs contribute to melanoma metastasis. In addition, adipocytes secrete EVs, which are oxidized by fatty acids and are absorbed by tumor cells, resulting in increased metastasis and invasion of melanoma []. EVs from melanoma cells with poor metastatic potential potently inhibit metastasis to the lung and trigger immune surveillance, resulting in the elicitation of a broad range of monocyte (PMO)-reliant innate immune responses. Furthermore, Plebanek et al. suggested that cancerous cells are cleared at the pre-metastatic niche [].
3.3. Tumor Microenvironment
The interactions of cancer cells with their environment determines whether the primary tumor is contained, metastasizes, or establishes dormant micrometastases. EVs play essential roles in the interstitial transport and intercellular communication within the tumor microenvironment (TME). Metastatic tumor cells show increased ability to sort EV cargo (i.e., proteins and microRNAs) and to release EVs. EV cargo is then transferred to stromal cells, including those that are present in premetastatic niches. Furthermore, EVs promote tumorigenesis and invasion through a variety of mechanisms, resulting in premetastatic niche formation. The following table describes the roles of EVs in the TME [] (Table 1).

Table 1.
The mechanisms of tumor microenvironment regulations in cancers.
3.4. Immune System
The tumor microenvironment controls immune surveillance and anti-tumor immunity [], mainly through intra- and extracellular signaling. Immunoediting is a complex process that includes intra- and extracellular signals. EVs play an important role in immune escape, both directly and indirectly. The direct modulation of either immune cells or their immature precursors is mostly driven by EV-mediated anti-apoptotic or pro-apoptotic signaling during the melanoma cell migration. The indirect roles of EVs include the expansion and differentiation of negative regulators of the immune system, such as myeloid-derived suppressor cells (MDSCs) and regulatory T lymphocytes (Tregs), thus promoting tumor cell escape from immune surveillance [,]. Several effects, i.e., mechanisms link EVs and the immune system (Table 2). Studies have shown that EVs secreted by tumor cells protect and maintain the growth of cancer cells, while EVs produced by normal cells, especially stem cells, inhibit tumor growth and suppress cancer progression []. Homing of melanoma exosomes to sentinel lymph nodes imposes synchronized molecular signals that effect melanoma cell recruitment, extracellular matrix deposition, and vascular proliferation in the lymph nodes []. In addition, tumor-derived EVs were shown to interfere with immunization by inducing loss of antigen expression, suppression of immune effector cells, exchange of nucleic acids, changes in recipient cell transcription, and inhibition of the immune cell response []. Other studies point out that tumor cells and tumor-infiltrating immune cells form a highly tolerant microenvironment, increasing tumor growth and allowing metastatic spread. Studies of anti-tumor immunity have explored the host’s immune responses and promote the development of new therapies and novel methods for use in future therapeutic methods [].
Table 2.
The effect of tumor-derived EVs in immune systems.
3.5. Drug Resistance and Clinical Treatment
EVs are involved in the development and regulation of different cancer-related processes. Drug resistance of cancer cells is a huge clinical problem and requires further investigation. Nevertheless, it is known that drug-resistant tumor cells are able to enclose chemotherapeutic agents in EVs and transfer anticancer drugs out of tumor cells. Therefore, understanding the molecular mechanisms and signaling pathways of EV-mediated drug resistance will help in the design of novel cancer treatments.
A large number of studies indicate that EVs play a crucial role in the development of the drug resistance of cancer cells (Table 3). Previous research has indicated that the use of BRAF kinase inhibitors (vemurafenib and dabrafenib) to treat melanoma patients bearing the BRAF-activating mutation V600E results in tumor regression, followed by quick development of drug resistance. Receptor tyrosine kinases (RTKs) are upregulated and activate the PI3K-Akt signaling pathway. EVs from drug-resistant melanoma cells were enriched with the RTK PDGFRβ, and delivering EVs rich in PDGFRβ to metastatic melanoma cells with the BRAF inhibitor-sensitive phenotype activated the PI3K/AKT pathway and resulted in the development of drug resistance []. Moreover, a novel truncated form of anaplastic lymphoma kinase (ALK) named ALKRES was found to be secreted in EVs. The transfer of ALKRES to sensitive, ALK-negative melanoma cells caused activation of the MAPK signaling pathway and transferred the characteristics of drug resistance to the recipient cells [].
Table 3.
The effect of tumor-derived EVs on drug resistance.
3.6. Small RNA (microRNA)
MicroRNAs (miRNAs) constitute a class of small single-stranded noncoding RNAs (~22 nt in length) that suppress gene expression. miRNAs are transcribed in the nucleus by RNA polymerase II or III. Primary miRNA transcripts (pri-miRNAs) are cleaved through a complex that consists of the endonuclease Drosha and the RNA-binding protein DGCR8. Hairpin pre-miRNAs are exported to the cytoplasm and are cleaved by the endonuclease Dicer to form dsRNA–miRNA duplexes. The complementary strand of the mature miRNA sequence is degraded, facilitating miRNA-induced silencing complex (RISC) formation and targeting the complementary sequences in the 3′ UTR of target mRNAs inhibit translation. miRNAs regulate many physiological and pathophysiological processes, such as growth, differentiation, and cancer progression. miRNAs regulate hundreds of genes; thus, miRNAs can cause complex phenotypic changes []. The loss of certain miRNAs facilitates cancer growth, whereas overexpression of other miRNAs promotes cancer progression []. miRNAs change the phenotype of melanoma cells and metabolic pathways during melanoma progression. They also affect the extracellular matrix (ECM), which includes fibroblasts, endothelial cells, and immune system cells []. miRNAs have different functions in each step of the development of different cancers []. Cells have the ability to selectively sort miRNAs into EVs for secretion to nearby or distant targets. Moreover, certain disease states have also identified dysregulated EV-miRNA content, shedding light on the potential role of selective sorting in pathogenesis. The latest findings regarding the roles of EVs-relevant miRNAs in melanoma pathobiology are summarized in Table 4.
Table 4.
The mechanisms and target locations of microRNA in melanoma.
4. Therapeutic Applications of Extracellular Vesicles
There are several studies on EVs in therapeutic applications. Interestingly, tumor cells release subpopulations of EVs that differ in their molecular and biological characteristics. These differences are essential for the precise transfer of biological information between cells. Accordingly, different components of EVs derived from different cells have different effects depending on their source. Based on these features, monitoring EV phenotypes during treatment enables the discovery of specific EV profiles and an understanding of how these correlate with drug resistance development in melanoma patients. Further analysis of EV heterogeneity will help in understanding the biology of EVs in health and disease and accelerate the development of EV-based diagnostic and therapeutic approaches. Melanoma is diagnosed with cancer-specific EV phenotypes from melanoma patient plasma by a multiplex EV phenotype analyzer chip that incorporates a nano-mixing-enhanced microchip and a multiplex surface-enhanced Raman scattering (SERS) nanotag system []. For these reasons, the therapeutic potential of EVs deserves further consideration in the context of drug delivery and regenerative medicine []. For example, EVs combined with liposomes and nanoparticles offer novel therapeutic delivery methods. Specifically, EVs derived from cancer cells can be carriers of drugs for delivery and can effectively inhibit tumor proliferation because of their ability to transfer biologically active components and overcome biological barriers []. In addition to being considered as potential therapeutics, EVs have the ability to enhance tissue regeneration and serve as potential replacements for stem cell therapy, playing a role in reimmunization, which promotes regeneration and inhibits pathogens []. These properties can lead to a wide range of therapeutic applications, including vaccination, treatment for autoimmune diseases, cancer, and tissue damage (Table 5). In recent years, many drugs for melanoma have been developed, but stimulating cancer cell death is still the major strategy []. If cancer cells acquire drug resistance, therapeutic treatment becomes challenging and the mortality rate significantly increases. The injected EVs derived from colon cancer through the tail vein of NOD.CB17-Prkdcscid/NcrCrlBltw mice determined neoplastic transformation and metastases in the lungs of the mice []. Another study proved that the timing of EV administration is as critical as that of oral administration after resection of the primary tumor reversed the pro-metastatic effects of milk-derived EVs in breast cancer models []. EVs from a highly metastatic clonal variant of the osteosarcoma cell line were internalized by a poorly metastatic clonal variant of the same cell line and induced a migratory and invasive phenotype. It was pointed out that EVs derived from highly metastatic clonal variants drive metastatic behaviors []. EVs originated from the brain carry messages to cancer cells that modify glioma cell metabolism, reducing lactate, nitric oxide (NO), and glutamate (Glu) release. EVs affect Glu homeostasis, increasing the expression of Glu transporter Glt-1 on astrocytes []. Recently, there is increasing evidence showing that EVs promote cancer progression and metastasis. It is suggested that clinicians effectively control the secretion of pernicious exosomes and melanoma will be remedied comprehensively.
Table 5.
The therapeutic applications of EVs in cancer.
5. Conclusions
EVs are important modulators of inter- and intracellular communications. EVs regulate diverse cellular processes, and they also contribute to cancer development and metastasis. The EV components can be transferred to other cells, affecting the physiological processes of the recipient cells and influencing the entire tumor microenvironment. EVs offer a valuable alternative to the current therapeutic options. They serve as nanoscale vehicles for drug delivery and have great potentials in this regard, not only because of their high biocompatibility, but also due to their low cytotoxicity. Tumor-cell-derived extracellular vesicle surface antigens have a huge effect on the immune system and can be modified by various agents to directly affect tumor cells or regulate antitumor immunity. The high EV heterogeneity is a problem in exploring their full therapeutic potential. Regarding their application as drug carriers, the disadvantages include low transfection efficiency and high dependence on cell division for cases in which cell cycle manipulation is required. In addition, the experimental conditions must be precisely controlled during nanoparticle delivery to avoid vesicle rupture. Thus, verifying the roles of EVs in clinical practice is not a simple endeavor, and more research is needed before EVs can be practically applied as therapeutic tools (Figure 2). Nevertheless, EVs offer a fully new approach for treating melanoma and other cancers. Understanding their regulation and biological features has a high potential to improve cancer treatment in the future.
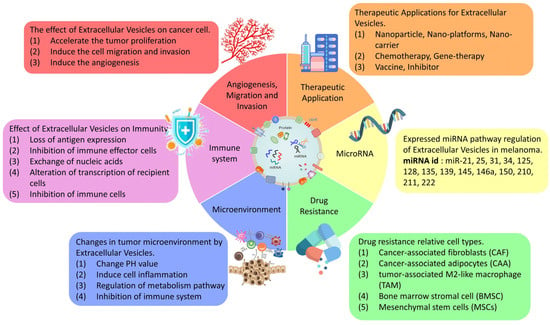
Figure 2.
The summary of EV effectiveness in cancer therapeutic treatments.
Author Contributions
Conceptualization, Y.-C.C., Y.-A.C., Y.-J.C. and H.-M.D.W.; figures and tables, Y.-C.C., Y.-A.C. and Y.-J.C.; writing, Y.-C.C., Y.-A.C., Y.-J.C., H.-L.S., Y.-L.H., H.-M.S., I.B. and H.-M.D.W. All authors have read and agreed to the published version of the manuscript.
Funding
We thank the Ministry of Science and Technology for the grant (Taiwan, MOST 109-2221-E-005-012), the Kaohsiung Armed Forces General Hospital (KAFGH_D_109014), the National Chung Hsing University (ENABLE: 108ST001B and 110ST001G), the Kasetsart University Joint Research Project (108RA129A), and the NCHU-MIRDC Bilateral Joint Research Project (110S0703B). H.M.S. and I.B. acknowledge the financial support by SFB1190 P17.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hogue, L.; Harvey, V.M. Basal cell carcinoma, squamous cell carcinoma, and cutaneous melanoma in skin of color patients. Dermatol. Clin. 2019, 37, 519–526. [Google Scholar] [CrossRef]
- Bandarchi, B.; Ma, L.; Navab, R.; Seth, A.; Rasty, G. From Melanocyte to Metastatic Malignant Melanoma. Dermatol. Res. Pract. 2010, 2010, 583748. [Google Scholar] [CrossRef]
- Volkovova, K.; Bilanicova, D.; Bartonova, A.; Letašiová, S.; Dusinska, M. Associations between environmental factors and incidence of cutaneous melanoma. Review. Env. Health-Glob. 2012, 11, S12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elder, D.E.; Bastian, B.C.; Cree, I.A.; Massi, D.; Scolyer, R.A. The 2018 World Health Organization classification of cutaneous, mucosal, and uveal melanoma: Detailed analysis of 9 distinct subtypes defined by their evolutionary pathway. Arch. Pathol. Lab. Med. 2020, 144, 500–522. [Google Scholar] [CrossRef] [Green Version]
- Storr, S.J.; Safuan, S.; Mitra, A.; Elliott, F.; Walker, C.; Vasko, M.J.; Ho, B.; Cook, M.; Mohammed, R.A.; Patel, P.M.; et al. Objective assessment of blood and lymphatic vessel invasion and association with macrophage infiltration in cutaneous melanoma. Mod. Pathol. 2012, 25, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Steinbichler, T.B.; Dudás, J.; Riechelmann, H.; Skvortsova, I.-I. The role of exosomes in cancer metastasis. Semin. Cancer Biol. 2017, 44, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Gowda, R.; Robertson, B.M.; Iyer, S.; Barry, J.; Dinavahi, S.S.; Robertson, G.P. The role of exosomes in metastasis and progression of melanoma. Cancer Treat Rev. 2020, 85, 101975. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Witwer, K.W.; Théry, C. Extracellular vesicles or exosomes? On primacy, precision, and popularity influencing a choice of nomenclature. J. Extracell Vesicles 2019, 8, 1648167. [Google Scholar] [CrossRef]
- De Broe, M.; Wieme, R.; Roels, F. Letter: Membrane fragments with koinozymic properties released from villous adenoma of the rectum. Lancet 1975, 2, 1214–1215. [Google Scholar] [CrossRef]
- Benz, E.W., Jr.; Moses, H.L. Small, virus-like particles detected in bovine sera by electron microscopy. J. Natl. Cancer Inst. 1974, 52, 1931–1934. [Google Scholar] [CrossRef]
- Pan, B.T.; Johnstone, R.M. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: Selective externalization of the receptor. Cell 1983, 33, 967–978. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, P.; Almeida, F. Role of Exosomal miRNAs and the Tumor Microenvironment in Drug Resistance. Cells 2020, 9, 1450. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Coscia, C.; Parolini, I.; Sanchez, M.; Biffoni, M.; Boussadia, Z.; Zanetti, C.; Fiani, M.L.; Sargiacomo, M. Generation, Quantification, and Tracing of Metabolically Labeled Fluorescent Exosomes. Methods Mol. Biol. 2016, 1448, 217–235. [Google Scholar]
- Surman, M.; Stępień, E.; Przybyło, M. Melanoma-Derived Extracellular Vesicles: Focus on Their Proteome. Proteomes 2019, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Desrochers, L.M.; Bordeleau, F.; Reinhart-King, C.A.; Cerione, R.A.; Antonyak, M.A. Microvesicles provide a mechanism for intercellular communication by embryonic stem cells during embryo implantation. Nat. Commun. 2016, 7, 11958. [Google Scholar] [CrossRef] [Green Version]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, J.L. Natural melanoma-derived extracellular vesicles. Semin. Cancer Biol. 2019, 59, 251–265. [Google Scholar] [CrossRef]
- Pegtel, D.M.; Peferoen, L.; Amor, S. Extracellular vesicles as modulators of cell-to-cell communication in the healthy and diseased brain. Philos. Trans. R Soc. Lond. B Biol. Sci. 2014, 369, 20130516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Diamond, M.P.; Al-Hendy, A. The emerging role of extracellular vesicle-derived miRNAs: Implication in cancer progression and stem cell related diseases. J. Clin. Epigenet 2016, 2. [Google Scholar]
- Dong, L.; Zieren, R.C.; Horie, K.; Kim, C.J.; Mallick, E.; Jing, Y.; Feng, M.; Kuczler, M.D.; Green, J.; Amend, S.R.; et al. Comprehensive evaluation of methods for small extracellular vesicles separation from human plasma, urine and cell culture medium. J. Extracell. Vesicles 2020, 10, e12044. [Google Scholar] [CrossRef]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alipoor, S.D.; Mortaz, E.; Varahram, M.; Movassaghi, M.; Kraneveld, A.D.; Garssen, J.; Adcock, I.M. The potential biomarkers and immunological effects of tumor-derived exosomes in lung cancer. Front. Immunol. 2018, 9, 819. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.S.; Won, J.H.; Lim, G.J.; Han, J.; Lee, J.Y.; Cho, K.O.; Bae, Y.K. A novel population of extracellular vesicles smaller than exosomes promotes cell proliferation. Cell Commun. Signal 2019, 17, 95. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, A.; Takahashi, Y.; Nishikawa, M.; Sano, K.; Morishita, M.; Charoenviriyakul, C.; Saji, H.; Takakura, Y. Accelerated growth of B16BL6 tumor in mice through efficient uptake of their own exosomes by B16BL6 cells. Cancer Sci. 2017, 108, 1803–1810. [Google Scholar] [CrossRef]
- Valacchi, G.; Sticozzi, C.; Lim, Y.; Pecorelli, A. Scavenger receptor class B type I: A multifunctional receptor. Ann. N.Y. Acad. Sci. 2011, 1229, E1–E7. [Google Scholar] [CrossRef]
- Simons, K.; Vaz, W.L. Model systems, lipid rafts, and cell membranes. Annu. Rev. Biophys. Biomol. Struct. 2004, 33, 269–295. [Google Scholar] [CrossRef]
- Kinslechner, K.; Schütz, B.; Pistek, M.; Rapolter, P.; Weitzenböck, H.P.; Hundsberger, H.; Mikulits, W.; Grillari, J.; Röhrl, C.; Hengstschläger, M.; et al. Loss of SR-BI Down-Regulates MITF and Suppresses Extracellular Vesicle Release in Human Melanoma. Int. J. Mol. Sci. 2019, 20, 1063. [Google Scholar] [CrossRef] [Green Version]
- Ekström, E.J.; Bergenfelz, C.; von Bülow, V.; Serifler, F.; Carlemalm, E.; Jönsson, G.; Andersson, T.; Leandersson, K. WNT5A induces release of exosomes containing pro-angiogenic and immunosuppressive factors from malignant melanoma cells. Mol. Cancer 2014, 13, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, J.L.; Pan, H.; Lanza, G.M.; Wickline, S.A. Consortium for Translational Research in Advanced, I.; Nanomedicine, Paracrine induction of endothelium by tumor exosomes. Lab. Invest. 2009, 89, 1317–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Yan, T.; Huang, C.; Xu, Z.; Wang, L.; Jiang, E.; Wang, H.; Chen, Y.; Liu, K.; Shao, Z.; et al. Melanoma cell-secreted exosomal miR-155-5p induce proangiogenic switch of cancer-associated fibroblasts via SOCS1/JAK2/STAT3 signaling pathway. J. Exp. Clin. Cancer Res. 2018, 37, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, N.; Walzl, A.; Unger, C.; Rosner, M.; Krupitza, G.; Hengstschläger, M.; Dolznig, H. In vitro cell migration and invasion assays. Mutat. Res. Rev. Mutat. Res. 2013, 752, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Gui, J.; Zahedi, F.; Yu, P.; Cho, C.; Bhattacharya, S.; Carbone, C.J.; Yu, Q.; Katlinski, K.V.; Katlinskaya, Y.V.; et al. An Interferon-Driven Oxysterol-Based Defense against Tumor-Derived Extracellular Vesicles. Cancer Cell 2019, 35, 33–45.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehead, B.; Wu, L.; Hvam, M.L.; Aslan, H.; Dong, M.; Dyrskjøt, L.; Ostenfeld, M.S.; Moghimi, S.M.; Howard, K.A. Tumour exosomes display differential mechanical and complement activation properties dependent on malignant state: Implications in endothelial leakiness. J. Extracell Vesicles 2015, 4, 29685. [Google Scholar] [CrossRef]
- Ghoshal, A.; Rodrigues, L.C.; Gowda, C.P.; Elcheva, I.A.; Liu, Z.; Abraham, T.; Spiegelman, V.S. Extracellular vesicle-dependent effect of RNA-binding protein IGF2BP1 on melanoma metastasis. Oncogene 2019, 38, 4182–4196. [Google Scholar] [CrossRef]
- Xiao, D.; Ohlendorf, J.; Chen, Y.; Taylor, D.D.; Rai, S.N.; Waigel, S.; Zacharias, W.; Hao, H.; McMasters, K.M. Identifying mRNA, MicroRNA and Protein Profiles of Melanoma Exosomes. PLoS ONE 2012, 7, e46874. [Google Scholar] [CrossRef] [Green Version]
- Mannavola, F.; Tucci, M.; Felici, C.; Passarelli, A.; D’Oronzo, S. Tumor-derived exosomes promote the in vitro osteotropism of melanoma cells by activating the SDF-1/CXCR4/CXCR7 axis. J. Transl. Med. 2019, 17. [Google Scholar] [CrossRef] [Green Version]
- Lazar, I.; Clement, E.; Dauvillier, S.; Milhas, D.; Ducoux-Petit, M.; LeGonidec, S.; Moro, C.; Soldan, V.; Dalle, S.; Balor, S.; et al. Adipocyte Exosomes Promote Melanoma Aggressiveness through Fatty Acid Oxidation: A Novel Mechanism Linking Obesity and Cancer. Cancer Res. 2016, 76, 4051–4057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plebanek, M.P.; Angeloni, N.L.; Vinokour, E.; Li, J.; Henkin, A.; Martinez-Marin, D.; Filleur, S.; Bhowmick, R.; Henkin, J.; Miller, S.D.; et al. Pre-metastatic cancer exosomes induce immune surveillance by patrolling monocytes at the metastatic niche. Nat. Commun. 2017, 8, 1319. [Google Scholar] [CrossRef]
- Tung, K.H.; Ernstoff, M.S.; Allen, C.; Shu, S. A Review of Exosomes and their Role in The Tumor Microenvironment and Host-Tumor "Macroenvironment". J. Immunol. Sci. 2019, 3, 4–8. [Google Scholar] [CrossRef] [Green Version]
- Logozzi, M.; Mizzoni, D.; Angelini, D.F.; Di Raimo, R.; Falchi, M.; Battistini, L.; Fais, S. Microenvironmental pH and Exosome Levels Interplay in Human Cancer Cell Lines of Different Histotypes. Cancers 2018, 10, 370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Lee, S.; Shin, E.; Seong, K.M.; Jin, Y.W.; Youn, H.; Youn, B. The Emerging Roles of Exosomes as EMT Regulators in Cancer. Cells 2020, 9, 861. [Google Scholar] [CrossRef]
- Gener Lahav, T.; Adler, O.; Zait, Y.; Shani, O.; Amer, M.; Doron, H.; Abramovitz, L.; Yofe, I.; Cohen, N.; Erez, N. Melanoma-derived extracellular vesicles instigate proinflammatory signaling in the metastatic microenvironment. Int. J. Cancer 2019, 145, 2521–2534. [Google Scholar] [CrossRef]
- Shu, S.L.; Yang, Y.; Allen, C.L.; Maguire, O.; Minderman, H.; Sen, A.; Ciesielski, M.J.; Collins, K.A.; Bush, P.J.; Singh, P.; et al. Metabolic reprogramming of stromal fibroblasts by melanoma exosome microRNA favours a pre-metastatic microenvironment. Sci. Rep. 2018, 8, 12905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siveen, K.S.; Raza, A.; Ahmed, E.I.; Khan, A.Q.; Prabhu, K.S.; Kuttikrishnan, S.; Mateo, J.M.; Zayed, H.; Rasul, K.; Azizi, F.; et al. The Role of Extracellular Vesicles as Modulators of the Tumor Microenvironment, Metastasis and Drug Resistance in Colorectal Cancer. Cancers 2019, 11, 746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, X.; Shen, H.; Li, Z.; Wang, T.; Wang, S. Tumor-derived exosomes, myeloid-derived suppressor cells, and tumor microenvironment. J. Hematol. Oncol. 2019, 12, 84. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, T. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucci, M.; Mannavola, F.; Passarelli, A.; Stucci, L.S.; Cives, M.; Silvestris, F. Exosomes in melanoma: A role in tumor progression, metastasis and impaired immune system activity. Oncotarget 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Düchler, M.; Czernek, L.; Peczek, L.; Cypryk, W.; Sztiller-Sikorska, M.; Czyz, M. Melanoma-Derived Extracellular Vesicles Bear the Potential for the Induction of Antigen-Specific Tolerance. Cells 2019, 8, 665. [Google Scholar] [CrossRef] [Green Version]
- Isola, L.A.; Chen, S. Exosomes: The messengers of health and disease. Curr. Neuropharmacol. 2017, 15, 157–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, J.L.; San, R.S.; Wickline, S.A. Exosomes released by melanoma cells prepare sentinel lymph nodes for tumor metastasis. Cancer Res. 2011, 71, 3792–3801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passarelli, A.; Mannavola, F.; Stucci, L.S.; Tucci, M.; Silvestris, F. Immune system and melanoma biology: A balance between immunosurveillance and immune escape. Oncotarget 2017, 8, 106132–106142. [Google Scholar] [CrossRef]
- Ventola, C.L. Cancer Immunotherapy, Part 1: Current Strategies and Agents. Pharm. Ther. 2017, 42, 375–383. [Google Scholar]
- Wieckowski, E.U.; Visus, C.; Szajnik, M.; Szczepanski, M.J.; Storkus, W.J.; Whiteside, T.L. Tumor-derived microvesicles promote regulatory T cell expansion and induce apoptosis in tumor-reactive activated CD8+ T lymphocytes. J. Immunol. 2009, 183, 3720–3730. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Yang, Y.; Wang, W.; Zhang, Y.; Chen, Z.; Hao, C.; Zhang, J. Melanoma-released exosomes directly activate the mitochondrial apoptotic pathway of CD4+ T cells through their microRNA cargo. Exp. Cell Res. 2018, 371, 364–371. [Google Scholar] [CrossRef]
- Yin, Y.; Cai, X.; Chen, X.; Liang, H.; Zhang, Y.; Li, J.; Wang, Z.; Chen, X.; Zhang, W.; Yokoyama, S.; et al. Tumor-secreted miR-214 induces regulatory T cells: A major link between immune evasion and tumor growth. Cell Res. 2014, 24, 1164–1180. [Google Scholar] [CrossRef]
- Hiltbrunner, S.; Larssen, P.; Eldh, M.; Martinez-Bravo, M.J.; Wagner, A.K.; Karlsson, M.C.; Gabrielsson, S. Exosomal cancer immunotherapy is independent of MHC molecules on exosomes. Oncotarget 2016, 7, 38707–38717. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al. Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 2018, 560, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Deng, W.; McGinley, E.C.; Klinke, D.J., 2nd. Melanoma exosomes deliver a complex biological payload that upregulates PTPN11 to suppress T lymphocyte function. J. Extracell Vesicles 2017, 30, 203–218. [Google Scholar]
- Cheng, L.; Wang, Y.; Huang, L. Exosomes from M1-Polarized Macrophages Potentiate the Cancer Vaccine by Creating a Pro-inflammatory Microenvironment in the Lymph Node. Mol. Ther. 2017, 25, 1665–1675. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Diergaarde, B.; Ferrone, S.; Kirkwood, J.M.; Whiteside, T.L. Melanoma cell-derived exosomes in plasma of melanoma patients suppress functions of immune effector cells. Sci. Rep.-UK 2020, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Vella, L.J.; Behren, A.; Coleman, B.; Greening, D.W.; Hill, A.F.; Cebon, J. Intercellular Resistance to BRAF Inhibition Can Be Mediated by Extracellular Vesicle–Associated PDGFRβ. Neoplasia 2017, 19, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Cesi, G.; Philippidou, D.; Kozar, I.; Kim, Y.J.; Bernardin, F.; Van Niel, G.; Wienecke-Baldacchino, A.; Felten, P.; Letellier, E.; Dengler, S.; et al. A new ALK isoform transported by extracellular vesicles confers drug resistance to melanoma cells. Mol. Cancer 2018, 17, 145. [Google Scholar] [CrossRef] [PubMed]
- Vu, L.T.; Gong, J.; Pham, T.T.; Kim, Y.; Le, M.T.N. microRNA exchange via extracellular vesicles in cancer. Cell Proliferat. 2020, 53, e12877. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, E.; Hershkovitz, L.; Itzhaki, O.; Hajdu, S.; Nemlich, Y.; Ortenberg, R.; Gefen, N.; Edry, L.; Modai, S.; Keisari, Y.; et al. Regulation of cancer aggressive features in melanoma cells by microRNAs. PLoS ONE 2011, 6, e18936. [Google Scholar] [CrossRef]
- Gajos-Michniewicz, A.; Czyz, M. Role of miRNAs in melanoma metastasis. Cancers 2019, 11, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajos-Michniewicz, A.; Duechler, M.; Czyz, M. MiRNA in melanoma-derived exosomes. Cancer Lett. 2014, 347, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Svedman, F.C.; Lohcharoenkal, W.; Bottai, M.; Brage, S.E.; Sonkoly, E.; Hansson, J.; Pivarcsi, A.; Eriksson, H. Extracellular microvesicle microRNAs as predictive biomarkers for targeted therapy in metastastic cutaneous malignant melanoma. PLoS ONE 2018, 13, e0206942. [Google Scholar] [CrossRef]
- Lee, J.H.; Dindorf, J.; Eberhardt, M.; Lai, X.; Ostalecki, C.; Koliha, N.; Gross, S.; Blume, K.; Bruns, H.; Wild, S.; et al. Innate extracellular vesicles from melanoma patients suppress β-catenin in tumor cells by miRNA-34a. Life Sci. Alliance 2019, 2, e201800205. [Google Scholar] [CrossRef]
- Dror, S.; Sander, L.; Schwartz, H.; Sheinboim, D.; Barzilai, A.; Dishon, Y.; Apcher, S.; Golan, T.; Greenberger, S.; Barshack, I.; et al. Melanoma miRNA trafficking controls tumour primary niche formation. Nat. Cell Biol. 2016, 18, 1006–1017. [Google Scholar] [CrossRef]
- Felicetti, F.; De Feo, A.; Coscia, C.; Puglisi, R.; Pedini, F.; Pasquini, L.; Bellenghi, M.; Errico, M.C.; Pagani, E.; Carè, A. Exosome-mediated transfer of miR-222 is sufficient to increase tumor malignancy in melanoma. J. Transl. Med. 2016, 14, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- House, I.G.; Petley, E.V.; Beavis, P.A. Tumor-derived exosomes modulate T cell function through transfer of RNA. FEBS J. 2018, 285, 1030–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chen, J.; Wang, S.; Li, P.; Zheng, C.; Zhou, X.; Tao, Y.; Chen, X.; Sun, L.; Wang, A.; et al. Blockage of transferred exosome-shuttled miR-494 inhibits melanoma growth and metastasis. J. Cell Physiol. 2019, 234, 15763–15774. [Google Scholar] [CrossRef]
- Huber, V.; Vallacchi, V.; Fleming, V.; Hu, X.; Cova, A.; Dugo, M.; Shahaj, E.; Sulsenti, R.; Vergani, E.; Filipazzi, P.; et al. Tumor-derived microRNAs induce myeloid suppressor cells and predict immunotherapy resistance in melanoma. J. Clin. Invest. 2018, 128, 5505–5516. [Google Scholar] [CrossRef] [Green Version]
- Luan, W.; Ding, Y.; Xi, H.; Ruan, H.; Lu, F.; Ma, S.; Wang, J. Exosomal miR-106b-5p derived from melanoma cell promotes primary melanocytes epithelial-mesenchymal transition through targeting EphA4. J.Exp. Clin. Cancer Res. 2021, 40, 107. [Google Scholar] [CrossRef]
- Dar, A.A.; Majid, S.; de Semir, D.; Nosrati, M.; Bezrookove, V.; Kashani-Sabet, M. miRNA-205 Suppresses Melanoma Cell Proliferation and Induces Senescence via Regulation of E2F1 Protein*. J. Biol. Chem. 2011, 286, 16606–16614. [Google Scholar] [CrossRef] [Green Version]
- Segura, M.F.; Hanniford, D.; Menendez, S.; Reavie, L.; Zou, X.; Alvarez-Diaz, S.; Zakrzewski, J.; Blochin, E.; Rose, A.; Bogunovic, D.; et al. Aberrant miR-182 expression promotes melanoma metastasis by repressing FOXO3 and microphthalmia-associated transcription factor. P. Natl. A. Sci. Bel. Agr. 2009, 106, 1814–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shull, A.Y.; Latham-Schwark, A.; Ramasamy, P.; Leskoske, K.; Oroian, D.; Birtwistle, M.R.; Buckhaults, P.J. Novel Somatic Mutations to PI3K Pathway Genes in Metastatic Melanoma. PLoS ONE 2012, 7, e43369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Q.; He, Q.; Wei, J. MicroRNA-342 prohibits proliferation and invasion of melanoma cells by directly targeting Zinc-finger E-box binding homeobox 1. Oncol. Res. 2018, 26, 1447. [Google Scholar] [CrossRef]
- Wang, J.; Wuethrich, A.; Sina, A.A.; Lane, R.E.; Lin, L.L.; Wang, Y.; Cebon, J.; Behren, A.; Trau, M. Tracking extracellular vesicle phenotypic changes enables treatment monitoring in melanoma. Sci. Adv. 2020, 6, eaax3223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melling, G.E.; Carollo, E.; Conlon, R.; Simpson, J.C.; Carter, D.R.F. The Challenges and Possibilities of Extracellular Vesicles as Therapeutic Vehicles. Eur. J. Pharm. Biopharm. 2019, 144, 50–56. [Google Scholar] [CrossRef]
- Willis, G.R.; Kourembanas, S.; Mitsialis, S.A. Therapeutic Applications of Extracellular Vesicles: Perspectives from Newborn Medicine. Methods Mol. Biol. 2017, 1660, 409–432. [Google Scholar]
- Wiklander, O.P.B.; Brennan, M.Á.; Lötvall, J.; Breakefield, X.O.; EL Andaloussi, S. Advances in therapeutic applications of extracellular vesicles. Sci. Transl. Med. 2019, 11, eaav8521. [Google Scholar] [CrossRef]
- Luke, J.J.; Flaherty, K.T.; Ribas, A.; Long, G.V. Targeted agents and immunotherapies: Optimizing outcomes in melanoma. Nat. Rev. Clin. Oncol. 2017, 14, 463–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdouh, M.; Floris, M.; Gao, Z.H.; Arena, V.; Arena, M.; Arena, G.O. Colorectal cancer-derived extracellular vesicles induce transformation of fibroblasts into colon carcinoma cells. J. Exp. Clin. Cancer Res. 2019, 38, 257. [Google Scholar] [CrossRef] [PubMed]
- Samuel, M.; Fonseka, P.; Sanwlani, R.; Gangoda, L.; Chee, S.H.; Keerthikumar, S.; Spurling, A.; Chitti, S.V.; Zanker, D.; Ang, C.-S.; et al. Oral administration of bovine milk-derived extracellular vesicles induces senescence in the primary tumor but accelerates cancer metastasis. Nat. Commun. 2021, 12, 3950. [Google Scholar] [CrossRef]
- Macklin, R.; Wang, H.; Loo, D.; Martin, S.; Cumming, A.; Cai, N.; Lane, R.; Ponce, N.S.; Topkas, E.; Inder, K.; et al. Extracellular vesicles secreted by highly metastatic clonal variants of osteosarcoma preferentially localize to the lungs and induce metastatic behaviour in poorly metastatic clones. Oncotarget 2016, 7, 43570–43587. [Google Scholar] [CrossRef] [Green Version]
- Serpe, C.; Monaco, L.; Relucenti, M.; Iovino, L.; Familiari, P.; Scavizzi, F.; Raspa, M.; Familiari, G.; Civiero, L.; D’Agnano, I.; et al. Microglia-Derived Small Extracellular Vesicles Reduce Glioma Growth by Modifying Tumor Cell Metabolism and Enhancing Glutamate Clearance through miR-124. Cells 2021, 10, 2066. [Google Scholar] [CrossRef]
- Iessi, E.; Logozzi, M.; Lugini, L.; Azzarito, T.; Federici, C.; Spugnini, E.P.; Mizzoni, D.; Di Raimo, R.; Angelini, D.F.; Battistini, L.; et al. Acridine Orange/exosomes increase the delivery and the effectiveness of Acridine Orange in human melanoma cells: A new prototype for theranostics of tumors. J. Enzyme Inhib. Med. Chem. 2017, 32, 648–657. [Google Scholar] [CrossRef]
- Lin, Q.; Qu, M.; Zhou, B.; Patra, H.K.; Sun, Z.; Luo, Q.; Yang, W.; Wu, Y.; Zhang, Y.; Li, L.; et al. Exosome-like nanoplatform modified with targeting ligand improves anti-cancer and anti-inflammation effects of imperialine. J. Control. Release. 2019, 311-312, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, V.V.; Svistunov, A.A.; Chubarev, V.N.; Dostdar, S.A.; Sokolov, A.V.; Brzecka, A.; Sukocheva, O.; Neganova, M.E.; Klochkov, S.G.; Somasundaram, S.G.; et al. Extracellular vesicles in cancer nanomedicine. Semin. Cancer Biol. 2019, 69. [Google Scholar] [CrossRef] [PubMed]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Koyama, Y.; Ito, T.; Hasegawa, A.; Eriguchi, M.; Inaba, T.; Ushigusa, T.; Sugiura, K. Exosomes derived from tumor cells genetically modified to express Mycobacterium tuberculosis antigen: A novel vaccine for cancer therapy. Biotechnol. Lett. 2016, 38, 1857–1866. [Google Scholar] [CrossRef]
- Erkan, E.P.; Saydam, N.; Chen, C.C.; Saydam, O. Extracellular vesicles as carriers of suicide mRNA and/or protein in cancer therapy. Methods Mol. Biol. 2019, 1895, 87–96. [Google Scholar] [PubMed]
- Chao, O.S.; Chang, T.C.; Di Bella, M.A.; Alessandro, R.; Anzanello, F.; Rappa, G.; Goodman, O.B.; Lorico, A. The HDAC6 Inhibitor Tubacin Induces Release of CD133(+) Extracellular Vesicles From Cancer Cells. J. Cell Biochem. 2017, 118, 4414–4424. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).