Double Duty: Mitotic Kinesins and Their Post-Mitotic Functions in Neurons


Abstract
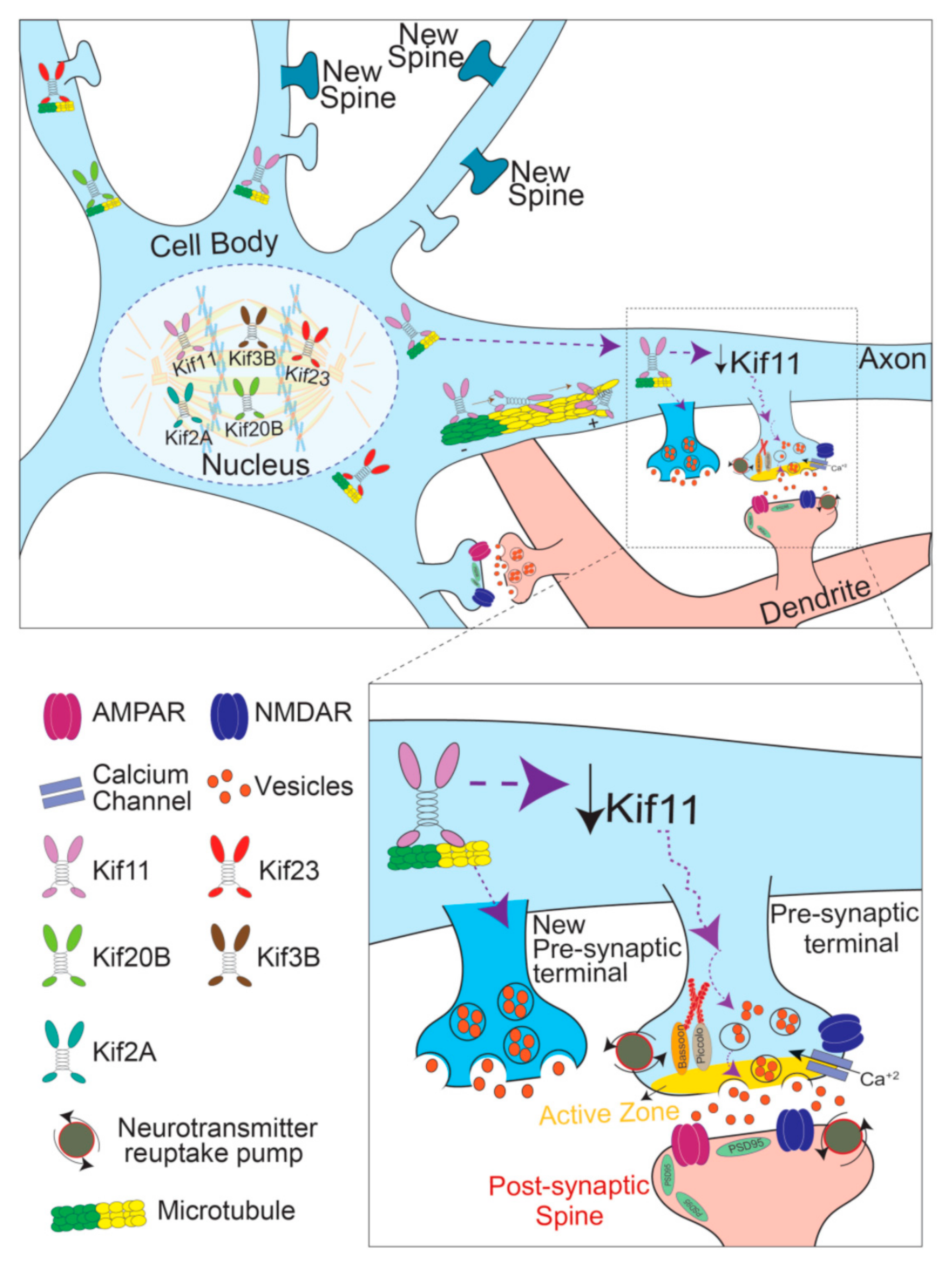
1. Introduction
2. Mitotic Functions of Kinesins
3. Post-Mitotic Function of Kinesins: Microtubule Regulation
4. Structural Plasticity
5. Transport
6. Synaptic Plasticity
7. Higher Order Cognition
8. Conclusions
Funding
Conflicts of Interest
References
- Hirokawa, N.; Niwa, S.; Tanaka, Y. Molecular Motors in Neurons: Transport Mechanisms and Roles in Brain Function, Development, and Disease. Neuron 2010, 68, 610–638. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, N. mRNA Transport in Dendrites: RNA Granules, Motors, and Tracks. J. Neurosci. 2006, 26, 7139–7142. [Google Scholar] [CrossRef] [PubMed]
- Nangaku, M.; Sato-Yoshitake, R.; Okada, Y.; Noda, Y.; Takemura, R.; Yamazaki, H.; Hirokawa, N. KIF1B, a novel microtubule plus end-directed monomeric motor protein for transport of mitochondria. Cell 1994, 79, 1209–1220. [Google Scholar] [CrossRef]
- Setou, M.; Nakagawa, T.; Seog, D.H.; Hirokawa, N. Kinesin superfamily motor protein KIF17 and mLin-10 in NMDA recep-tor-containing vesicle transport. Science 2000, 288, 1796–1802. [Google Scholar] [CrossRef] [PubMed]
- Kanai, Y.; Dohmae, N.; Hirokawa, N. Kinesin Transports RNA. Neuron 2004, 43, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Vale, R.D.; Reese, T.S.; Sheetz, M.P. Identification of a novel force-generating protein, kinesin, involved in microtubule-based motility. Cell 1985, 42, 39–50. [Google Scholar] [CrossRef]
- Brady, S.T. A novel brain ATPase with properties expected for the fast axonal transport motor. Nat. Cell Biol. 1985, 317, 73–75. [Google Scholar] [CrossRef]
- Miki, H.; Setou, M.; Kaneshiro, K.; Hirokawa, N. All kinesin superfamily protein, KIF, genes in mouse and human. Proc. Natl. Acad. Sci. 2001, 98, 7004–7011. [Google Scholar] [CrossRef]
- Marx, A.; Müller, J.; Mandelkow, E. The structure of microtubule motor proteins. Protein Simulations 2005, 71, 299–344. [Google Scholar] [CrossRef]
- Endow, S.A.; Barker, D.S. Processive and Nonprocessive Models of Kinesin Movement. Annu. Rev. Physiol. 2003, 65, 161–175. [Google Scholar] [CrossRef]
- Freixo, F.; Delgado, P.M.; Manso, Y.; Sánchez-Huertas, C.; Lacasa, C.; Soriano, E.; Roig, J.; Lüders, J. NEK7 regulates dendrite morphogenesis in neurons via Eg5-dependent microtubule stabilization. Nat. Commun. 2018, 9, 2330. [Google Scholar] [CrossRef] [PubMed]
- Swarnkar, S.; Avchalumov, Y.; Raveendra, B.L.; Grinman, E.; Puthanveettil, S.V. Kinesin Family of Proteins Kif11 and Kif21B Act as Inhibitory Constraints of Excitatory Synaptic Transmission Through Distinct Mechanisms. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Frade, J.M.; Ovejero-Benito, M.C. Neuronal cell cycle: The neuron itself and its circumstances. Cell Cycle 2015, 14, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Vashishtha, M.; Walther, Z.; Hall, J.L. The kinesin-homologous protein encoded by the Chlamydomonas FLA10 gene is associated with basal bodies and centrioles. J. Cell Sci. 1996, 109. [Google Scholar]
- Haraguchi, K.; Hayashi, T.; Jimbo, T.; Yamamoto, T.; Akiyama, T. Role of the Kinesin-2 Family Protein, KIF3, during Mitosis. J. Biol. Chem. 2006, 281, 4094–4099. [Google Scholar] [CrossRef]
- Cross, R.A.; McAinsh, A.D. Prime movers: The mechanochemistry of mitotic kinesins. Nat. Rev. Mol. Cell Biol. 2014, 15, 257–271. [Google Scholar] [CrossRef]
- Hildebrandt, E.R.; Gheber, L.; Kingsbury, T.; Hoyt, M.A. Homotetrameric Form of Cin8p, aSaccharomyces cerevisiaeKinesin-5 Motor, Is Essential for Itsin VivoFunction. J. Biol. Chem. 2006, 281, 26004–26013. [Google Scholar] [CrossRef]
- Kapitein, L.C.; Peterman, E.J.G.; Kwok, B.H.; Kim, J.H.; Kapoor, T.M.; Schmidt, C.F. The bipolar mitotic kinesin Eg5 moves on both microtubules that it crosslinks. Nat. Cell Biol. 2005, 435, 114–118. [Google Scholar] [CrossRef]
- Sawin, K.E.; Leguellec, K.; Philippe, M.; Mitchison, T.J. Mitotic spindle organization by a plus-end-directed microtubule motor. Nat. Cell Biol. 1992, 359, 540–543. [Google Scholar] [CrossRef]
- Myers, K.A.; Baas, P.W. Kinesin-5 regulates the growth of the axon by acting as a brake on its microtubule array. J. Cell Biol. 2007, 178, 1081–1091. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, Z.; Ouyang, M.; Yang, F.; Hao, H.; Lamb, K.L.; Yang, J.; Yin, Y.; Shen, H.W. PTEN regulates EG5 to control spindle architecture and chromosome congression during mitosis. Nat. Commun. 2016, 7, 12355. [Google Scholar] [CrossRef]
- Xu, M.; Liu, D.; Dong, Z.; Wang, X.; Wang, X.; Liu, Y.; Baas, P.W.; Liu, M. Kinesin-12 influences axonal growth during zebrafish neural development. Cytoskelet. 2014, 71, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Tanenbaum, M.E.; Macůrek, L.; Janssen, A.; Geers, E.F.; Alvarez-Fernández, M.; Medema, R.H. Kif15 Cooperates with Eg5 to Promote Bipolar Spindle Assembly. Curr. Biol. 2009, 19, 1703–1711. [Google Scholar] [CrossRef] [PubMed]
- Boleti, H.; Karsenti, E.; Vernos, I. Xklp2, a Novel Xenopus Centrosomal Kinesin-like Protein Required for Centrosome Separation during Mitosis. Cell 1996, 84, 49–59. [Google Scholar] [CrossRef]
- Vanneste, D.; Takagi, M.; Imamoto, N.; Vernos, I. The Role of Hklp2 in the Stabilization and Maintenance of Spindle Bipolarity. Curr. Biol. 2009, 19, 1712–1717. [Google Scholar] [CrossRef] [PubMed]
- She, Z.-Y.; Yang, W.-X. Molecular mechanisms of kinesin-14 motors in spindle assembly and chromosome segregation. J. Cell Sci. 2017, 130, 2097–2110. [Google Scholar] [CrossRef]
- Cai, S.; Weaver, L.N.; Ems-McClung, S.C.; Walczak, C.E. Kinesin-14 Family Proteins HSET/XCTK2 Control Spindle Length by Cross-Linking and Sliding Microtubules. Mol. Biol. Cell 2009, 20, 1348–1359. [Google Scholar] [CrossRef]
- Wei, Y.-L.; Yang, W.-X. Kinesin-14 motor protein KIFC1 participates in DNA synthesis and chromatin maintenance. Cell Death Dis. 2019, 10, 402. [Google Scholar] [CrossRef]
- Ali, A.; Veeranki, S.N.; Chinchole, A.; Tyagi, S. MLL/WDR5 Complex Regulates Kif2A Localization to Ensure Chromosome Congression and Proper Spindle Assembly during Mitosis. Dev. Cell 2017, 41, 605–622. [Google Scholar] [CrossRef]
- Ganem, N.J.; Upton, K.; Compton, D.A. Efficient Mitosis in Human Cells Lacking Poleward Microtubule Flux. Curr. Biol. 2005, 15, 1827–1832. [Google Scholar] [CrossRef]
- Ganem, N.J.; Compton, D.A. The KinI kinesin Kif2a is required for bipolar spindle assembly through a functional relationship with MCAK. J. Cell Biol. 2004, 166, 473–478. [Google Scholar] [CrossRef]
- Janisch, K.M.; Vock, V.M.; Fleming, M.S.; Shrestha, A.; Grimsley-Myers, C.M.; Rasoul, B.A.; Neale, S.A.; Cupp, T.D.; Kinchen, J.M.; Liem, K.F.; et al. The vertebrate-specific Kinesin-6, Kif20b, is required for normal cytokinesis of polarized cortical stem cells and cerebral cortex size. Dev. 2013, 140, 4672–4682. [Google Scholar] [CrossRef]
- Abaza, A.; Soleilhac, J.-M.; Westendorf, J.; Piel, M.; Crevel, I.; Roux, A.; Pirollet, F. M Phase Phosphoprotein 1 Is a Human Plus-end-directed Kinesin-related Protein Required for Cytokinesis. J. Biol. Chem. 2003, 278, 27844–27852. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, L.; Bassi, Z.I.; Geymonat, M.; Todesca, S.; Copoiu, L.; Enright, A.J.; Callaini, G.; Riparbelli, M.G.; Yu, L.; Choudhary, J.S.; et al. The midbody interactome reveals unexpected roles for PP1 phosphatases in cytokinesis. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Neef, R.; Klein, U.R.; Kopajtich, R.; Barr, F.A. Cooperation between Mitotic Kinesins Controls the Late Stages of Cytokinesis. Curr. Biol. 2006, 16, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Hunter, A.W.; Wordeman, L. How motor proteins influence microtubule polymerization dynamics. J. Cell Sci. 2000, 113. [Google Scholar]
- van Riel, W.E.; Rai, A.; Bianchi, S.; Katrukha, E.A.; Liu, Q.; Heck, A.J.R.; Hoogenraad, C.C.; Steinmetz, M.O.; Kapitein, L.C.; Akhmanova, A. Kinesin-4 KIF21B is a potent microtubule pausing factor. eLife 2017, 6, e24746. [Google Scholar] [CrossRef]
- Ebernardinelli, Y.; Enikonenko, I.; Muller, D. Structural plasticity: Mechanisms and contribution to developmental psychiatric disorders. Front. Neuroanat. 2014, 8, 123. [Google Scholar] [CrossRef]
- Butz, M.V.; Wörgötter, F.; Van Ooyen, A. Activity-dependent structural plasticity. Brain Res. Rev. 2009, 60, 287–305. [Google Scholar] [CrossRef]
- Puthanveettil, S.V.; Antonov, I.; Kalachikov, S.; Rajasethupathy, P.; Choi, Y.-B.; Kohn, A.B.; Citarella, M.; Yu, F.; Karl, K.A.; Kinet, M.; et al. A strategy to capture and characterize the synaptic transcriptome. Proc. Natl. Acad. Sci. 2013, 110, 7464–7469. [Google Scholar] [CrossRef]
- Puthanveettil, S.V.; Monje, F.J.; Miniaci, M.C.; Choi, Y.-B.; Karl, K.A.; Khandros, E.; Gawinowicz, M.A.; Sheetz, M.P.; Kandel, E.R. A New Component in Synaptic Plasticity: Upregulation of Kinesin in the Neurons of the Gill-Withdrawal Reflex. Cell 2008, 135, 960–973. [Google Scholar] [CrossRef] [PubMed]
- Guillaud, L.; Setou, M.; Hirokawa, N. KIF17 Dynamics and Regulation of NR2B Trafficking in Hippocampal Neurons. J. Neurosci. 2003, 23, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Muhia, M.; Thies, E.; Labonté, D.; Ghiretti, A.E.; Gromova, K.V.; Xompero, F.; Lappe-Siefke, C.; Hermans-Borgmeyer, I.; Kuhl, D.; Schweizer, M.; et al. The Kinesin KIF21B Regulates Microtubule Dynamics and Is Essential for Neuronal Morphology, Synapse Function, and Learning and Memory. Cell Rep. 2016, 15, 968–977. [Google Scholar] [CrossRef]
- Takeda, S.; Yamazaki, H.; Seog, D.-H.; Kanai, Y.; Terada, S.; Hirokawa, N. Kinesin Superfamily Protein 3 (Kif3) Motor Transports Fodrin-Associating Vesicles Important for Neurite Building. J. Cell Biol. 2000, 148, 1255–1266. [Google Scholar] [CrossRef]
- Leterrier, C. A dual role for βII-spectrin in axons. Proc. Natl. Acad. Sci. 2019, 116, 15324–15326. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, D.N.; Badea, A.; Zhou, R.; Mohler, P.J.; Zhuang, X.; Bennett, V. βII-spectrin promotes mouse brain connectivity through stabilizing axonal plasma membranes and enabling axonal organelle transport. Proc. Natl. Acad. Sci. 2019, 116, 15686–15695. [Google Scholar] [CrossRef]
- Alsabban, A.H.; Morikawa, M.; Tanaka, Y.; Takei, Y.; Hirokawa, N. Kinesin Kif3b mutation reduces NMDAR subunit NR 2A trafficking and causes schizophrenia-like phenotypes in mice. EMBO J. 2020, 39, e101090. [Google Scholar] [CrossRef]
- Muralidharan, H.; Baas, P.W. Mitotic Motor KIFC1 Is an Organizer of Microtubules in the Axon. J. Neurosci. 2019, 39, 3792–3811. [Google Scholar] [CrossRef]
- Braun, M.; Drummond, D.R.; Cross, R.; McAinsh, A.D. The kinesin-14 Klp2 organizes microtubules into parallel bundles by an ATP-dependent sorting mechanism. Nat. Cell Biol. 2009, 11, 724–730. [Google Scholar] [CrossRef]
- Maor-Nof, M.; Homma, N.; Raanan, C.; Nof, A.; Hirokawa, N.; Yaron, A. Axonal Pruning Is Actively Regulated by the Microtubule-Destabilizing Protein Kinesin Superfamily Protein 2A. Cell Rep. 2013, 3, 971–977. [Google Scholar] [CrossRef]
- Buster, D.W.; Baird, D.H.; Yu, W.; Solowska, J.M.; Chauvière, M.; Mazurek, A.; Kress, M.; Baas, P.W. Expression of the mitotic kinesin Kif15 in postmitotic neurons: Implications for neuronal migration and development. J. Neurocytol. 2003, 32, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Homma, N.; Takei, Y.; Tanaka, Y.; Nakata, T.; Terada, S.; Kikkawa, M.; Noda, Y.; Hirokawa, N. Kinesin Superfamily Protein 2A (KIF2A) Functions in Suppression of Collateral Branch Extension. Cell 2003, 114, 229–239. [Google Scholar] [CrossRef]
- McNeely, K.C.; Cupp, T.D.; Little, J.N.; Janisch, K.M.; Shrestha, A.; Dwyer, N.D. Mutation of Kinesin-6 Kif20b causes defects in cortical neuron polarization and morphogenesis. Neural Dev. 2017, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Sapir, T.; Levy, T.; Sakakibara, A.; Rabinkov, A.; Miyata, T.; Reiner, O. Shootin1 Acts in Concert with KIF20B to Promote Polarization of Migrating Neurons. J. Neurosci. 2013, 33, 11932–11948. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Cook, C.; Sauter, C.; Kuriyama, R.; Kaplan, P.L.; Baas, P.W. Depletion of a Microtubule-Associated Motor Protein Induces the Loss of Dendritic Identity. J. Neurosci. 2000, 20, 5782–5791. [Google Scholar] [CrossRef] [PubMed]
- Joseph, N.F.; Grinman, E.; Swarnkar, S.; Puthanveettil, S.V. Molecular Motor KIF3B Acts as a Key Regulator of Dendritic Architecture in Cortical Neurons. Front. Cell. Neurosci. 2020, 14, 521199. [Google Scholar] [CrossRef] [PubMed]
- Delandre, C.; Amikura, R.; Moore, A.W. Microtubule nucleation and organization in dendrites. Cell Cycle 2016, 15, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Kahn, O.I.; Ha, N.; Baird, M.A.; Davidson, M.W.; Baas, P.W. TPX2 regulates neuronal morphology through kinesin-5 interaction. Cytoskelet. 2015, 72, 340–348. [Google Scholar] [CrossRef]
- Silverman, M.; Kaech, S.; Ramser, E.M.; Lu, X.; Lasarev, M.R.; Nagalla, S.; Banker, G. Expression of kinesin superfamily genes in cultured hippocampal neurons. Cytoskelet. 2010, 67, 784–795. [Google Scholar] [CrossRef]
- Noda, Y.; Niwa, S.; Homma, N.; Fukuda, H.; Imajo-Ohmi, S.; Hirokawa, N. Phosphatidylinositol 4-phosphate 5-kinase alpha (PIPK ) regulates neuronal microtubule depolymerase kinesin, KIF2A and suppresses elongation of axon branches. Proc. Natl. Acad. Sci. 2012, 109, 1725–1730. [Google Scholar] [CrossRef]
- Ogawa, T.; Hirokawa, N. Microtubule Destabilizer KIF2A Undergoes Distinct Site-Specific Phosphorylation Cascades that Differentially Affect Neuronal Morphogenesis. Cell Rep. 2015, 12, 1774–1788. [Google Scholar] [CrossRef] [PubMed]
- Sharp, D.J.; Yu, W.; Ferhat, L.; Kuriyama, R.; Rueger, D.C.; Baas, P.W. Identification of a Microtubule-associated Motor Protein Essential for Dendritic Differentiation. J. Cell Biol. 1997, 138, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Liu, M.; Mozgova, O.I.; Yu, W.; Baas, P.W. Mitotic Motors Coregulate Microtubule Patterns in Axons and Dendrites. J. Neurosci. 2012, 32, 14033–14049. [Google Scholar] [CrossRef] [PubMed]
- 64 Liu, X.-A.; Kadakkuzha, B.; Pascal, B.; Steckler, C.; Akhmedov, K.; Yan, L.; Chalmers, M.; Puthanveettil, S.V. New approach to capture and characterize synaptic proteome. Proc. Natl. Acad. Sci. 2014, 111, 16154–16159. [Google Scholar] [CrossRef]
- Yamazaki, H.; Nakata, T.; Okada, Y.; Hirokawa, N. KIF3A/B: A heterodimeric kinesin superfamily protein that works as a microtubule plus end-directed motor for membrane organelle transport. J. Cell Biol. 1995, 130, 1387–1399. [Google Scholar] [CrossRef]
- Hebb, D.O. The Organization of Behavior: A Neuropsychological Theory; John Willey & Sons: New York, NY, USA, 1949. [Google Scholar]
- Citri, A.; Malenka, R.C. Synaptic Plasticity: Multiple Forms, Functions, and Mechanisms. Neuropsychopharmacol. 2007, 33, 18–41. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Kamboj, S.; Ehlers, M.D.; Rosen, K.R.; Fischbach, G.D.; Huganir, R.L. Activity-Dependent Modulation of Synaptic AMPA Receptor Accumulation. Neuron 1998, 21, 1067–1078. [Google Scholar] [CrossRef]
- Murthy, V.N.; Schikorski, T.; Stevens, C.F.; Zhu, Y. Inactivity produces increases in neurotransmitter release and synapse size. Neuron 2001, 32, 673–682. [Google Scholar] [CrossRef]
- Bullock, J.M.; Medway, C.; Cortina-Borja, M.; Turton, J.; Prince, J.A.; Ibrahim-Verbaas, C.A.; Schuur, M.; Breteler, M.M.; Van Duijn, C.M.; Kehoe, P.G.; et al. Discovery by the Epistasis Project of an epistatic interaction between the GSTM3 gene and the HHEX/IDE/KIF11 locus in the risk of Alzheimer’s disease. Neurobiol. Aging 2013, 34, 1309.e1–1309.e7. [Google Scholar] [CrossRef]
- Yin, X.; Feng, X.; Takei, Y.; Hirokawa, N. Regulation of NMDA Receptor Transport: A KIF17-Cargo Binding/Releasing Underlies Synaptic Plasticity and Memory In Vivo. J. Neurosci. 2012, 32, 5486–5499. [Google Scholar] [CrossRef]
- Rath, O.; Kozielski, F. Kinesins and cancer. Nat. Rev. Cancer 2012, 12, 527–539. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Mitotic Kinesin | KIF Family | Function in Neurons | Ref |
|---|---|---|---|
| KIF1C | kinesin-14 | Sorts MT into parallel bundles and cross link MTs; promotes axonal growth and the stability of the growth cones | [27,48,49] |
| KIF2A | kinesin-13 | MT depolymerase; promotes axonal pruning | [50] |
| KIF3B | kinesin-2 | Traffics component of the plasma membrane and NR2A to the synapse; reduces the density of dendritic spines | [44,47] |
| KIF11 | kinesin-5 | MT stabilizer; impedes dendritic spine growth and dendritic branching; suppresses axonal growth | [11,12,20] |
| KIF15 | kinesin-12 | MT depolymerase; suppresses the growth of axonal collateral branches | [51,52] |
| KIF20B | kinesin-6 | May bundle MTs into tight arrays; regulates neuron polarization, and axon and dendrite branching, outgrowth | [53,54] |
| KIF23 | kinesin-6 | Transports minus-end MTs into nascent dendrites | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joseph, N.F.; Swarnkar, S.; Puthanveettil, S.V. Double Duty: Mitotic Kinesins and Their Post-Mitotic Functions in Neurons. Cells 2021, 10, 136. https://doi.org/10.3390/cells10010136
Joseph NF, Swarnkar S, Puthanveettil SV. Double Duty: Mitotic Kinesins and Their Post-Mitotic Functions in Neurons. Cells. 2021; 10(1):136. https://doi.org/10.3390/cells10010136
Chicago/Turabian StyleJoseph, Nadine F., Supriya Swarnkar, and Sathyanarayanan V Puthanveettil. 2021. "Double Duty: Mitotic Kinesins and Their Post-Mitotic Functions in Neurons" Cells 10, no. 1: 136. https://doi.org/10.3390/cells10010136
APA StyleJoseph, N. F., Swarnkar, S., & Puthanveettil, S. V. (2021). Double Duty: Mitotic Kinesins and Their Post-Mitotic Functions in Neurons. Cells, 10(1), 136. https://doi.org/10.3390/cells10010136