SSR Marker-Assisted Management of Parental Germplasm in Sugarcane (Saccharum spp. hybrids) Breeding Programs

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. SSR Genotyping
2.3. Genetic Diversity Analysis
2.4. Population Structure Analysis
2.5. Differentiation Analysis and Genetic Diversity Indices
3. Results
3.1. Polymorphism Revealed by SSR Genotyping
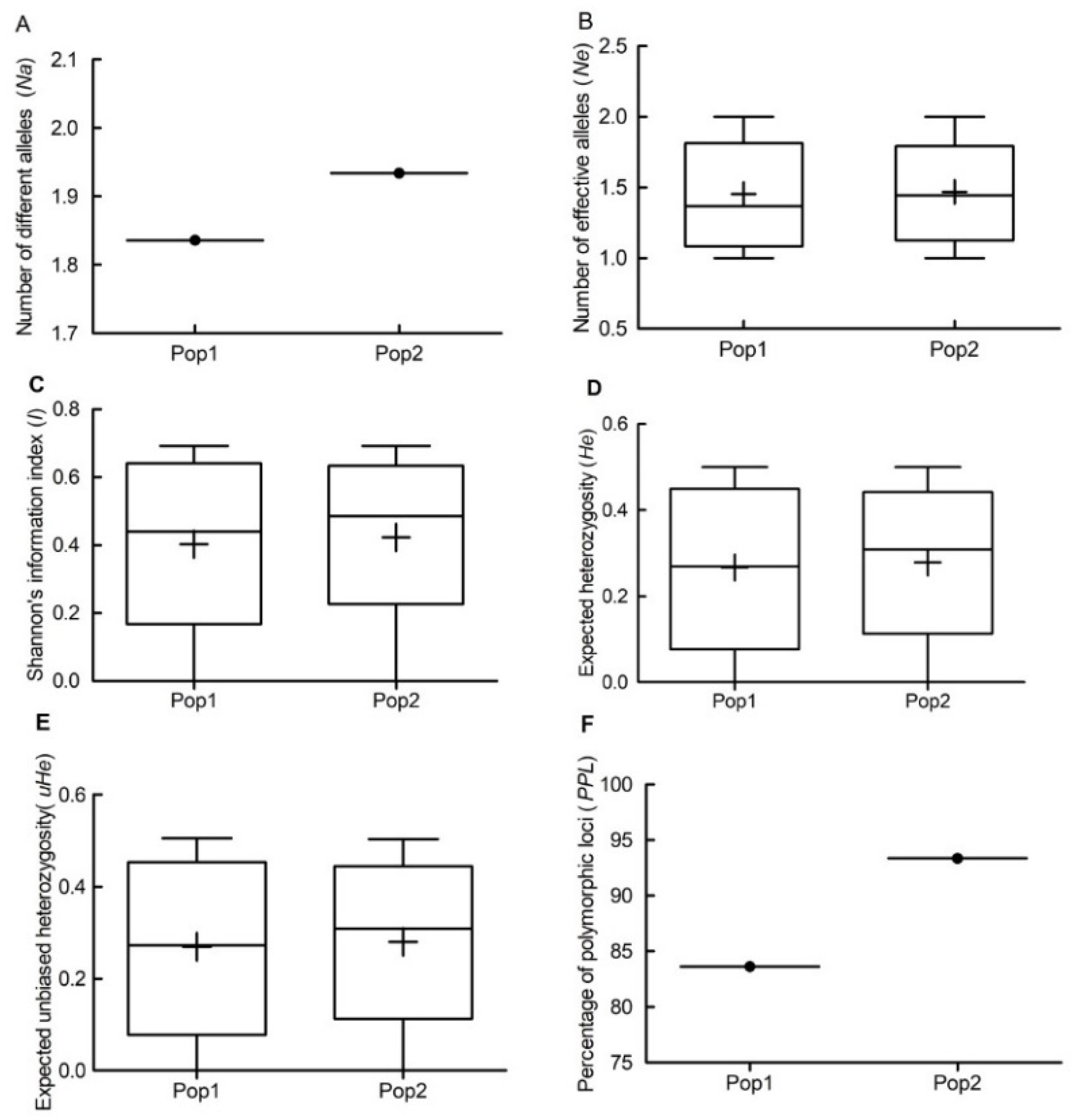
3.2. Genetic Diversity
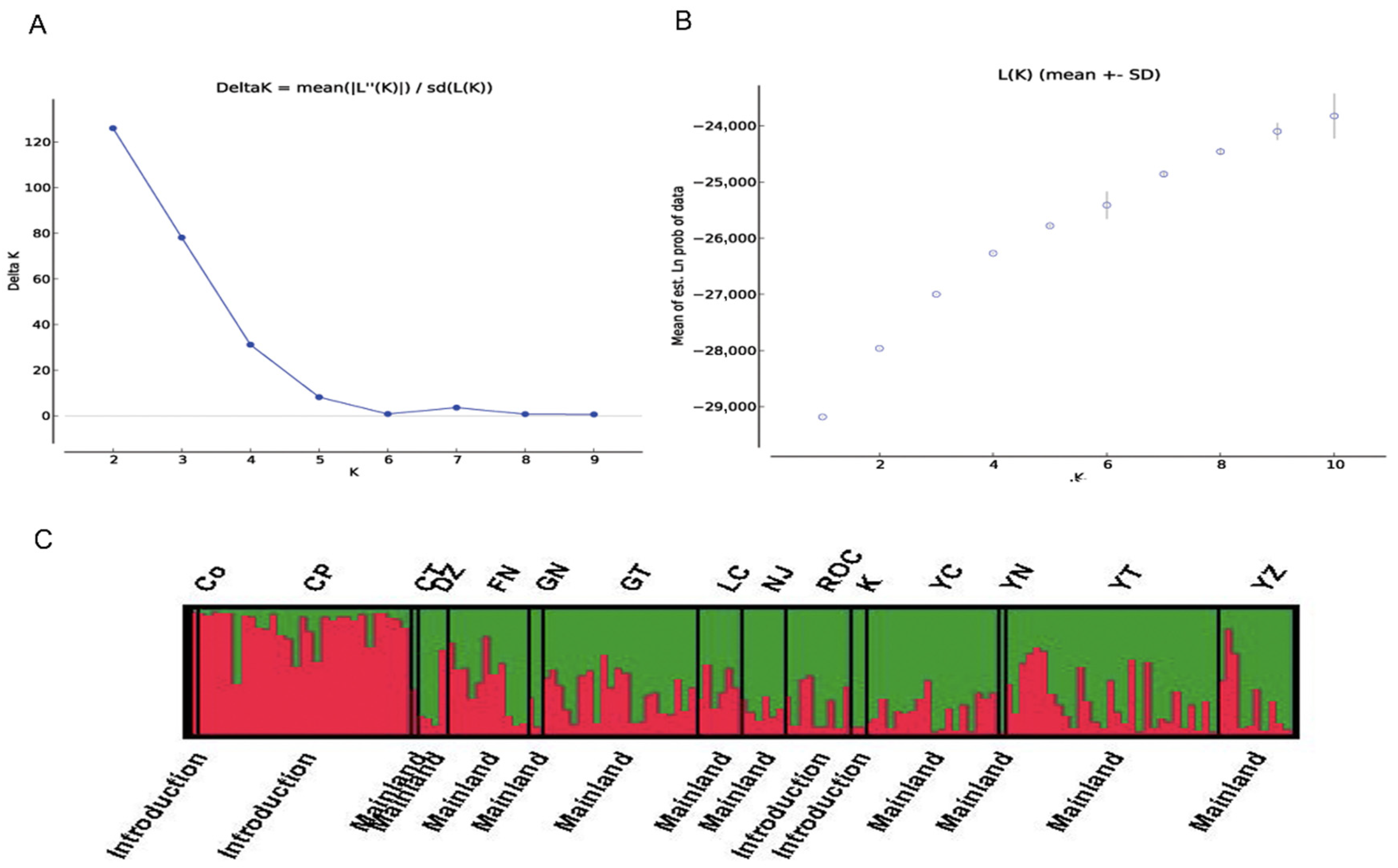
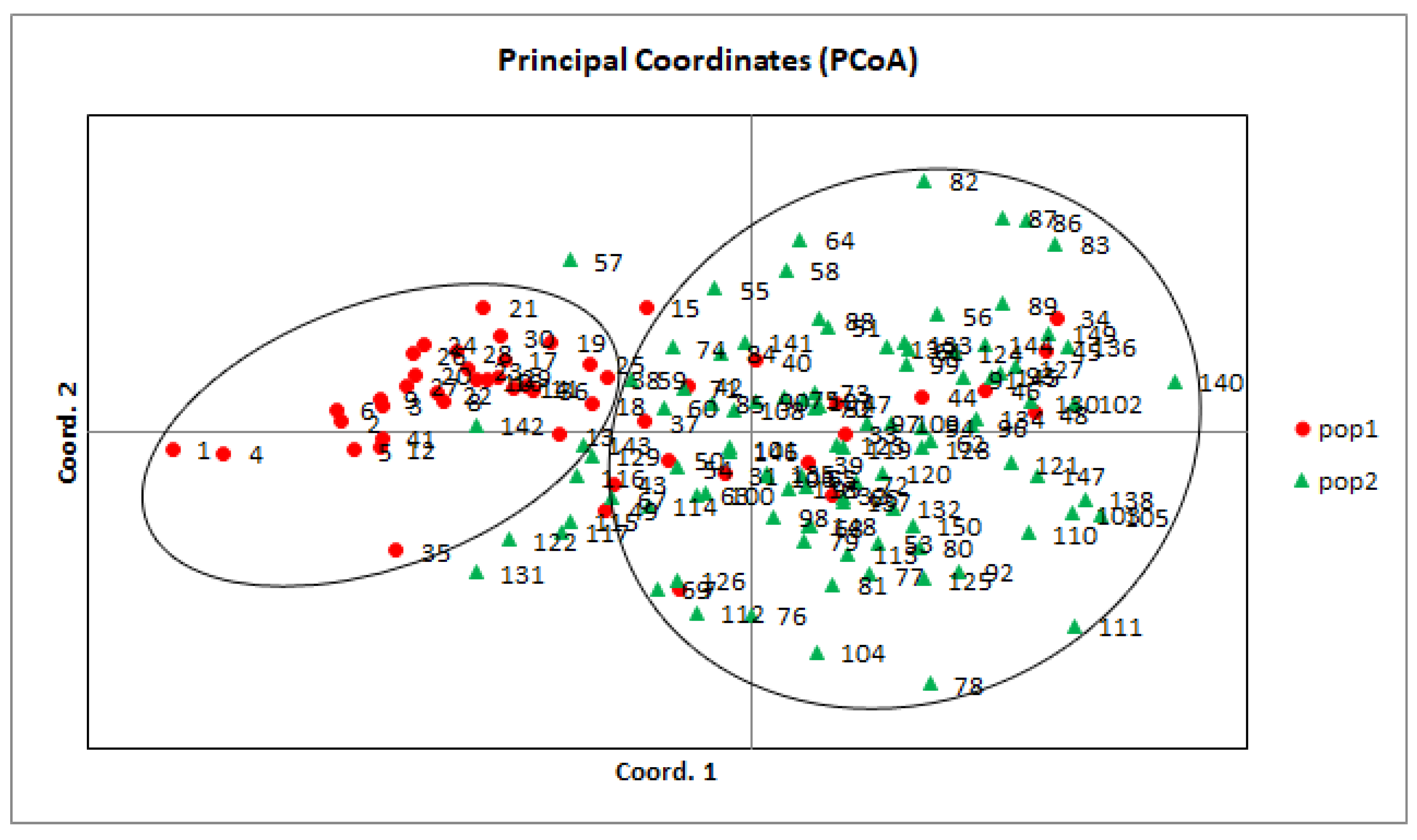
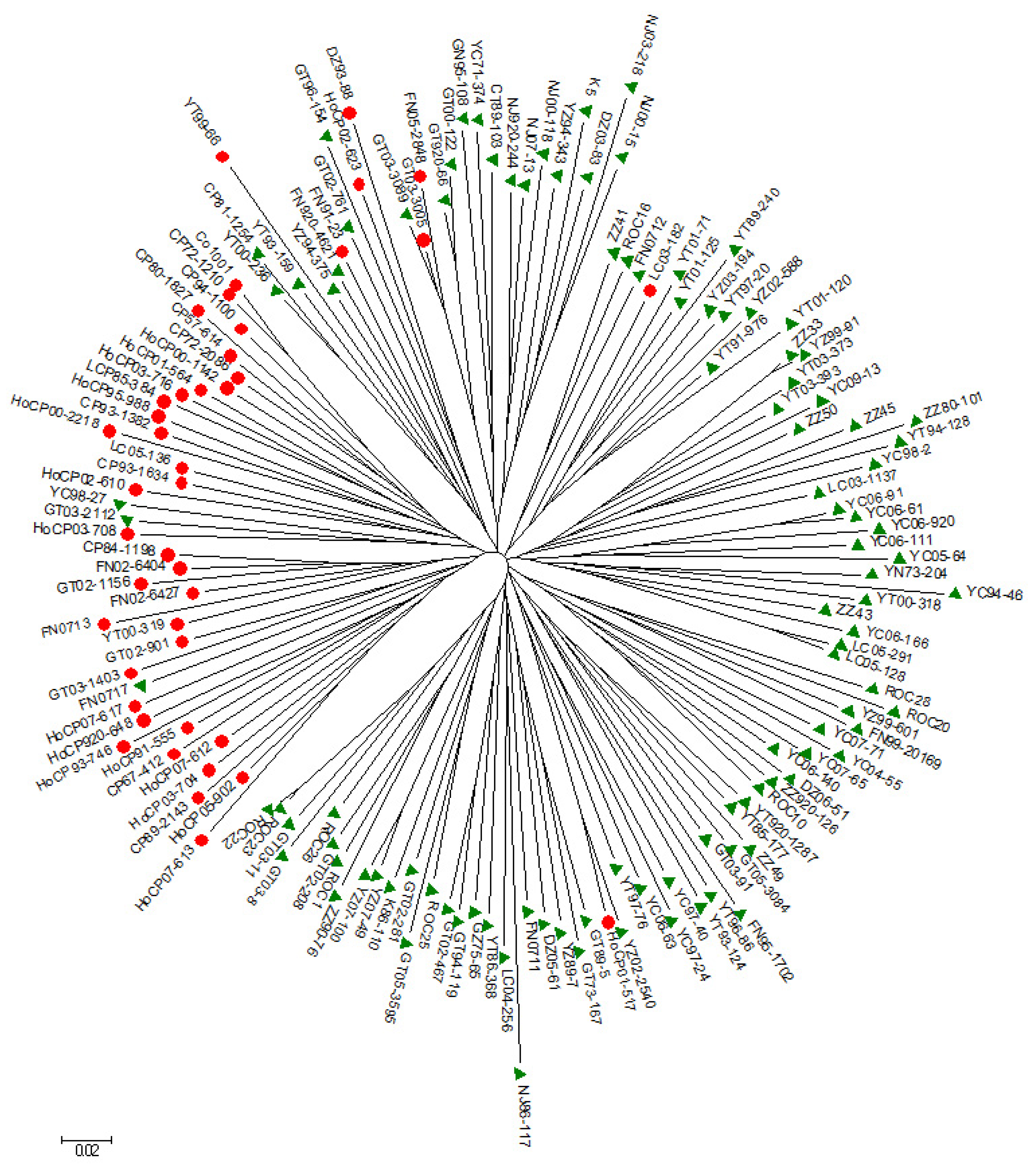
3.3. Population Structure and Phylogeny
3.4. Genetic Differentiation and Allelic Pattern Across Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lima, M.L.A.; Garcia, A.A.F.; Oliveira, K.M.; Matsuoka, S.; Arizono, H.; de Souza, C.L.; de Souza, A.P. Analysis of genetic similarity detected by AFLP and coefficient of parentage among genotypes of sugar cane (Saccharum spp.). Theor. Appl. Genet. 2002, 104, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Ming, R.; Moore, P.H.; Wu, K.; D’Hont, A.; Glaszmann, J.C.; Tew, T.L.; Mirkov, T.E.; Da Silva, J.; Jifon, J.; Rai, M.; et al. Sugarcane Improvement through Breeding and Biotechnology; John Wiley & Sons, Ltd.: New York, NY, USA, 2010; pp. 15–118. [Google Scholar]
- Qi, Y.W.; Pan, Y.B.; Fang, Y.L.; Zhang, C.M.; Fan, L.N.; He, H.Y.; Liu, R.; Wang, Q.N.; Liu, S.M.; Liu, F.Y.; et al. Genetic structure and diversity of parental cultivars involved in China mainland sugarcane breeding programs as inferred from DNA microsatellites. J. Integr. Agric. 2012, 11, 1794–1803. [Google Scholar] [CrossRef]
- Nair, N.V.; Selvi, A.; Sreenivasan, T.V.; Pushpalatha, K.N. Molecular diversity in Indian sugarcane cultivars as revealed by Randomly Amplified DNA polymorphisms. Euphytica 2002, 127, 219–225. [Google Scholar] [CrossRef]
- Creste, S.; Sansoli, D.M.; Tardiani, A.C.S.; Silva, D.N.; Gonçalves, F.K.; Fávero, T.M.; Medeiros, C.N.F.; Festucci, C.S.; Carlini-Garcia, L.A.; Landell, M.G.A.; et al. Comparison of AFLP, TRAP and SSRs in the estimation of genetic relationships in sugarcane. Sugar Tech. 2010, 12, 150–154. [Google Scholar] [CrossRef]
- Jannoo, N.; Grivet, L.; Seguin, M.; Paulet, F.; Domaingue, R.; Rao, P.S.; Dookun, A.; D’Hont, A.; Glaszmann, J.C. Molecular investigation of the genetic base of sugarcane cultivars. Theor. Appl. Genet. 1999, 99, 171–184. [Google Scholar] [CrossRef]
- Silva, J.A.G.D.; Sorrells, M.E.; Burnquist, W.L.; Tanksley, S.D. RFLP linkage map and genome analysis of Saccharum spontaneum. Genome 1993, 36, 782–791. [Google Scholar] [CrossRef]
- Pan, Y.B.; Burner, D.M.; Legendre, B.L.; Grisham, M.P.; White, W.H. An assessment of the genetic diversity within a collection of Saccharum spontaneum L. with RAPD-PCR. Genet. Resour. Crop Evol. 2004, 51, 895–903. [Google Scholar] [CrossRef]
- Singh, P.; Singh, S.P.; Tiwari, A.K.; Sharma, B.L. Genetic diversity of sugarcane hybrid cultivars by RAPD markers. 3 Biotech 2017, 7, 222. [Google Scholar] [CrossRef]
- Garcia, A.A.F.; Mollinari, M.; Marconi, T.G.; Serang, O.R.; Silva, R.R.; Vieira, M.L.C.; Vicentini, R.; Costa, E.A.; Mancini, M.C.; Garcia, M.O.S.; et al. SNP genotyping allows an in-depth characterisation of the genome of sugarcane and other complex autopolyploids. Sci. Rep. UK 2013, 3, 3399. [Google Scholar] [CrossRef]
- Parida, S.K.; Kalia, S.K.; Kaul, S.; Dalal, V.; Hemaprabha, G.; Selvi, A.; Pandit, A.; Singh, A.; Gaikwad, K.; Sharma, T.R.; et al. Informative genomic microsatellite markers for efficient genotyping applications in sugarcane. Theor. Appl. Genet. 2009, 118, 327–338. [Google Scholar] [CrossRef]
- Devarumath, R.M.; Kalwade, S.B.; Kawar, P.G.; Sushir, K.V. Assessment of Genetic Diversity in Sugarcane Germplasm Using ISSR and SSR Markers. Sugar Tech. 2012, 14, 334–344. [Google Scholar] [CrossRef]
- Oliveira, L.A.R.; Machado, C.A.; Cardoso, M.N.; Oliveira, A.C.A.; Amaral, A.L.; Rabbani, A.R.C.; Silva, A.V.C.; Ledo, A.S. Genetic diversity of Saccharum complex using ISSR markers. Genet. Mol. Res. 2017, 16, 1–9. [Google Scholar] [CrossRef][Green Version]
- Pinto, L.R.; Oliveira, K.M.; Marconi, T.; Garcia, A.A.F.; Ulian, E.C.; de Souza, A.P. Characterization of novel sugarcane expressed sequence tag microsatellites and their comparison with genomic SSRs. Plant Breed. 2006, 125, 378–384. [Google Scholar] [CrossRef]
- James, B.T.; Chen, C.; Rudolph, A.; Swaminathan, K.; Murray, J.E.; Na, J.; Spence, A.K.; Smith, B.; Hudson, M.E.; Moose, S.P.; et al. Development of microsatellite markers in autopolyploid sugarcane and comparative analysis of conserved microsatellites in sorghum and sugarcane. Mol. Breed. 2012, 30, 661–669. [Google Scholar] [CrossRef]
- Parthiban, S.; Govindaraj, P.; Senthilkumar, S. Comparison of relative efficiency of genomic SSR and EST-SSR markers in estimating genetic diversity in sugarcane. 3 Biotech 2018, 8, 144. [Google Scholar] [CrossRef]
- Pan, Y.B.; Burner, D.M.; Legendre, B.L. An Assessment of the Phylogenetic Relationship Among Sugarcane and Related Taxa Based on the Nucleotide Sequence of 5S rRNA Intergenic Spacers. Genetica 2000, 108, 285–295. [Google Scholar] [CrossRef]
- Que, Y.X.; Pan, Y.B.; Lu, Y.H.; Yang, C.; Yang, Y.T.; Huang, N.; Xu, L.P. Genetic analysis of diversity within a Chinese local sugarcane germplasm based on start codon targeted polymorphism. Biomed Res. Int. 2014, 2014, 468375. [Google Scholar] [CrossRef]
- Que, Y.X.; Chen, T.; Xu, L.; Chen, R.K. Genetic diversity among key sugarcane clones revealed by TRAP markers. J. Agric. Biotechnol. 2009, 17, 496–503. [Google Scholar]
- Alwala, S.; Suman, A.; Arro, J.A.; Veremis, J.C.; Kimbeng, C.A. Target region amplification polymorphism (TRAP) for assessing genetic diversity in sugarcane germplasm collections. Crop Sci. 2006, 46, 448. [Google Scholar] [CrossRef]
- Khan, M.; Pan, Y.B.; Iqbal, J. Development of an RAPD-based SCAR marker for smut disease resistance in commercial sugarcane cultivars of Pakistan. Crop Prot. 2017, 94, 166–172. [Google Scholar] [CrossRef]
- Ali, A.; Pan, Y.; Wang, Q.; Wang, J.; Chen, J.; Gao, S. Genetic diversity and population structure analysis of Saccharum and Erianthus genera using microsatellite (SSR) markers. Sci. Rep. UK 2019, 9, 395. [Google Scholar] [CrossRef]
- Nayak, S.N.; Song, J.; Villa, A.; Pathak, B.; Ayala-Silva, T.; Yang, X.; Todd, J.; Glynn, N.C.; Kuhn, D.N.; Glaz, B.; et al. Promoting Utilization of Saccharum spp. Genetic Resources through Genetic Diversity Analysis and Core Collection Construction. PLoS ONE 2014, 9, e110856. [Google Scholar] [CrossRef]
- Liu, H.L.; Yang, X.P.; You, Q.; Song, J.; Wang, L.P.; Zhang, J.S.; Deng, Z.H.; Ming, R.; Wang, J.P. Pedigree, marker recruitment, and genetic diversity of modern sugarcane cultivars in China and the United States. Euphytica 2018, 214, 48. [Google Scholar] [CrossRef]
- Pan, Y. Development and Integration of an SSR-Based Molecular Identity Database into Sugarcane Breeding Program. Agronomy 2016, 6, 28. [Google Scholar] [CrossRef]
- Marconi, T.G.; Costa, E.A.; Miranda, H.R.; Mancini, M.C.; Cardososilva, C.B. Functional markers for gene mapping and genetic diversity studies in sugarcane. BMC Res. Notes 2011, 4, 264. [Google Scholar] [CrossRef]
- Andru, S.; Pan, Y.; Thongthawee, S.; Burner, D.M.; Kimbeng, C.A. Genetic analysis of the sugarcane (Saccharum spp.) cultivar ‘LCP 85-384’. I. Linkage mapping using AFLP, SSR, and TRAP markers. Theor. Appl. Genet. 2011, 123, 77–93. [Google Scholar] [CrossRef]
- Banerjee, N.; Siraree, A.; Yadav, S.; Kumar, S.; Singh, J.; Kumar, S.; Pandey, D.K.; Singh, R.K. Marker-trait association study for sucrose and yield contributing traits in sugarcane (Saccharum spp. hybrid). Euphytica 2015, 205, 185–201. [Google Scholar] [CrossRef]
- Racedo, J.; Gutiérrez, L.; Perera, M.F.; Ostengo, S.; Pardo, E.M.; Cuenya, M.I.; Welin, B.; Castagnaro, A.P. Genome-wide association mapping of quantitative traits in a breeding population of sugarcane. BMC Plant Biol. 2016, 16, 142. [Google Scholar] [CrossRef]
- Ukoskit, K.; Posudsavang, G.; Pongsiripat, N.; Chatwachirawong, P.; Klomsa-ard, P.; Poomipant, P.; Tragoonrung, S. Detection and validation of EST-SSR markers associated with sugar-related traits in sugarcane using linkage and association mapping. Genomics 2019, 111, 1–9. [Google Scholar] [CrossRef]
- Pan, Y.B. Highly polymorphic microsatellite DNA markers for sugarcane germplasm evaluation and variety identity testing. Sugar Tech. 2006, 8, 246–256. [Google Scholar] [CrossRef]
- Chen, P.H.; Pan, Y.B.; Chen, R.K.; Xu, L.P.; Chen, Y.Q. SSR marker-based analysis of genetic relatedness among sugarcane cultivars (Saccharum spp. hybrids) from breeding programs in China and other countries. Sugar Tech. 2009, 11, 347–354. [Google Scholar] [CrossRef]
- Liu, P.W.; Que, Y.X.; Pan, Y.B. Highly Polymorphic Microsatellite DNA Markers for Sugarcane Germplasm Evaluation and Variety Identity Testing. Sugar Tech. 2011, 13, 129–136. [Google Scholar] [CrossRef]
- Pan, Y.B.; Liu, P.W.; Que, Y.X. Independently Segregating Simple Sequence Repeats (SSR) Alleles in Polyploid Sugarcane. Sugar Tech. 2015, 17, 235–242. [Google Scholar] [CrossRef]
- You, Q.; Pan, Y.; Xu, L.; Gao, S.; Wang, Q.; Su, Y.; Yang, Y.; Wu, Q.; Zhou, D.; Que, Y. Genetic Diversity Analysis of Sugarcane Germplasm Based on Fluorescence-Labeled Simple Sequence Repeat Markers and a Capillary Electrophoresis-based Genotyping Platform. Sugar Tech. 2016, 18, 380–390. [Google Scholar] [CrossRef]
- Ali, A.; Wang, J.; Pan, Y.; Deng, Z.; Chen, Z.; Chen, R.; Gao, S. Molecular identification and genetic diversity analysis of Chinese sugarcane (Saccharum spp. Hybrids) varieties using SSR markers. Trop. Plant Biol. 2017, 10, 194–203. [Google Scholar] [CrossRef]
- Fu, Y.; Pan, Y.; Lei, C.; Grisham, M.P.; Yang, C.; Meng, Q. Genotype-Specific Microsatellite (SSR) Markers for the Sugarcane Germplasm from the Karst Region of Guizhou, China. Am. J. Plant Sci. 2016, 7, 2209–2220. [Google Scholar] [CrossRef]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from plant tissues. Plant Molecular Biology Manual; Gelvin, S.B., Schilperoort, R.A., Verma, D.P.S., Eds.; Springer: Dordrecht, The Netherlands, 1989. [Google Scholar]
- Pan, Y.B.; Scheffler, B.E.; Richard, E., Jr. High throughput genotyping of commercial sugarcane clones with microsatellite (SSR) DNA markers. Sugar Tech. 2007, 9, 176–181. [Google Scholar]
- Singh, R.K.; Mishra, S.K.; Singh, S.P.; Mishra, N.; Sharma, M.L. Evaluation of microsatellite markers for genetic diversity analysis among sugarcane species and commercial hybrids. Aust. J. Crop Sci. 2010, 4, 116–125. [Google Scholar]
- Parida, S.K.; Pandit, A.; Gaikwad, K.; Sharma, T.R.; Srivastava, P.; Singh, N.K.; Mohapatra, T. Functionally relevant microsatellites in sugarcane unigenes. BMC Plant Biol. 2010, 10, 251. [Google Scholar] [CrossRef]
- Singh, R.K.; Singh, R.B.; Singh, S.P.; Sharma, M.L. Identification of sugarcane microsatellites associated to sugar content in sugarcane and transferability to other cereal genomes. Euphytica 2011, 182, 335–354. [Google Scholar] [CrossRef]
- Pan, Y.B.; Tew, T.L.; Schnell, R.J.; Viator, R.P.; Richard, E.P.; Grisham, M.P.; White, W.H. Microsatellite DNA marker-assisted selection of Saccharum spontaneum cytoplasm-derived germplasm. Sugar Tech. 2006, 8, 23–29. [Google Scholar] [CrossRef]
- Fan, W.Q.; Ge, H.M.; Sun, X.S.; Yang, A.G.; Zhang, Z.F.; Ren, M. DataFormater, A software for SSR data formatting to develop population genetics analysis. Mol. Plant Breed. 2016, 14, 1029–1034. [Google Scholar]
- Liu, K.; Muse, S.V. PowerMaker: An integrated analysis environment for genetic maker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of Population Structure Using Multilocus Genotype Data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2010, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; VonHoldt, B.M. Structure harvester: A website and program for visualizing structure output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. Clumpp: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Luo, Z.; Brock, J.; Dyer, J.M.; Kutchan, T.; Schachtman, D.; Augustin, M.; Ge, Y.; Fahlgren, N.; Abdel-Haleem, H. Genetic Diversity and Population Structure of a Camelina sativa Spring Panel. Front. Plant Sci. 2019, 10, 184. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2016, 30, 2725–2729. [Google Scholar] [CrossRef]
- Jackson, P.A. Breeding for improved sugar content in sugarcane. Field Crop. Res. 2005, 92, 277–290. [Google Scholar] [CrossRef]
- Stevenson, G.C. Genetic and Breeding of Sugarcane; Longmans: London, UK, 1965. [Google Scholar]
- Cordeiro, G.M.; Casu, R.; McIntyre, C.L.; Manners, J.M.; Henry, R.J. Microsatellite markers from sugarcane (Saccharum spp.) ESTs cross transferable to erianthus and sorghum. Plant Sci. 2001, 160, 1115–1123. [Google Scholar] [CrossRef]
- Hameed, U.; Pan, Y.; Muhammad, K.; Afghan, S.; Iqbal, J. Use of simple sequence repeat markers for DNA fingerprinting and diversity analysis of sugarcane (Saccharum spp.) cultivars resistant and susceptible to red rot. Genet. Mol. Res. 2012, 11, 1195. [Google Scholar] [CrossRef]
- You, Q.; Xu, L.P.; Zheng, Y.F.; Que, Y.X. Genetic diversity analysis of sugarcane parents in Chinese breeding programmes using gSSR markers. Sci. World J. 2013, 2013, 613062. [Google Scholar] [CrossRef]
- Szpiech, Z.A.; Rosenberg, N.A. On the size distribution of private microsatellite alleles. Theor. Popul. Biol. 2011, 80, 100–113. [Google Scholar] [CrossRef]
- Slatkin, M. Rare alleles as indicators of gene flow. Evolution 1985, 39, 53–65. [Google Scholar] [CrossRef]
- Kalinowski, S.T. Counting Alleles with Rarefaction: Private Alleles and Hierarchical Sampling Designs. Conserv. Genet. 2004, 5, 539–543. [Google Scholar] [CrossRef]
- Schroeder, K.B.; Schurr, T.G.; Long, J.C.; Rosenberg, N.A.; Crawford, M.H.; Tarskaia, L.A.; Osipova, L.P.; Zhadanov, S.I.; Smith, D.G. A private allele ubiquitous in the Americas. Biol. Lett. 2007, 3, 218–223. [Google Scholar] [CrossRef][Green Version]
- Bird, K.A.; An, H.; Gazave, E.; Gore, M.A.; Pires, J.C.; Robertson, L.D.; Labate, J.A. Population structure and phylogenetic relationships in a diverse panel of Brassica rapa L. Front. Plant Sci. 2017, 8, 321. [Google Scholar] [CrossRef]
- Wright, S. The interpretation of population structure by F-statistics with special regard to systems of mating. Evolution 1965, 19, 395–420. [Google Scholar] [CrossRef]
- Deng, H.H.; Li, Q.W. Utilization of CP72-1210 in sugarcane breeding program in mainland China. Guangdong Agric. Sci. 2007, 11, 18–21. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Accession | Series | No. | Accession | Series | No. | Accession | Series |
|---|---|---|---|---|---|---|---|---|
| 1 | Co1001 | Co | 51 | GZ75-65 | GN | 101 | YC06-92 | YC |
| 2 | CP57-614 | CP | 52 | HoCP00-1142 | CP | 102 | YC07-65 | YC |
| 3 | CP67-412 | CP | 53 | HoCP00-2218 | CP | 103 | YC07-71 | YC |
| 4 | CP72-1210 | CP | 54 | HoCP01-517 | CP | 104 | YC09-13 | YC |
| 5 | CP72-2086 | CP | 55 | HoCP01-564 | CP | 105 | YC71-374 | YC |
| 6 | CP80-1827 | CP | 56 | HoCP02-610 | CP | 106 | YC94-46 | YC |
| 7 | CP81-1254 | CP | 57 | HoCP02-623 | CP | 107 | YC97-24 | YC |
| 8 | CP84-1198 | CP | 58 | HoCP03-704 | CP | 108 | YC97-40 | YC |
| 9 | CP89-2143 | CP | 59 | HoCP03-708 | CP | 109 | YC98-2 | YC |
| 10 | CP93-1382 | CP | 60 | HoCP03-716 | CP | 110 | YC98-27 | YC |
| 11 | CP93-1634 | CP | 61 | HoCP05-902 | CP | 111 | YN73-204 | YN |
| 12 | CP94-1100 | CP | 62 | HoCP07-612 | CP | 112 | YT00-236 | YT |
| 13 | CT89-103 | CT | 63 | HoCP07-613 | CP | 113 | YT00-318 | YT |
| 14 | DZ03-83 | DZ | 64 | HoCP07-617 | CP | 114 | YT00-319 | YT |
| 15 | DZ05-61 | DZ | 65 | HoCP91-555 | CP | 115 | YT01-120 | YT |
| 16 | DZ06-51 | DZ | 66 | HoCP92-648 | CP | 116 | YT01-125 | YT |
| 17 | DZ93-88 | DZ | 67 | HoCP93-746 | CP | 117 | YT01-71 | YT |
| 18 | FN02-6404 | FN | 68 | HoCP95-988 | CP | 118 | YT03-373 | YT |
| 19 | FN02-6427 | FN | 69 | K5 | K | 119 | YT03-393 | YT |
| 20 | FN05-2848 | FN | 70 | K86-110 | K | 120 | YT85-177 | YT |
| 21 | FN0711 | FN | 71 | LC03-1137 | LC | 121 | YT86-368 | YT |
| 22 | FN0712 | FN | 72 | LC03-182 | LC | 122 | YT89-240 | YT |
| 23 | FN0713 | FN | 73 | LC04-256 | LC | 123 | YT91-976 | YT |
| 24 | FN0717 | FN | 74 | LC05-128 | LC | 124 | YT92-1287 | YT |
| 25 | FN91-23 | FN | 75 | LC05-136 | LC | 125 | YT93-124 | YT |
| 26 | FN92-4621 | FN | 76 | LC05-291 | LC | 126 | YT93-159 | YT |
| 27 | FN95-1702 | FN | 77 | LCP85-384 | CP | 127 | YT94-128 | YT |
| 28 | FN99-20169 | FN | 78 | NJ00-118 | NJ | 128 | YT96-86 | YT |
| 29 | GN95-108 | GN | 79 | NJ00-15 | NJ | 129 | YT97-20 | YT |
| 30 | GT00-122 | GT | 80 | NJ03-218 | NJ | 130 | YT97-76 | YT |
| 31 | GT02-1156 | GT | 81 | NJ07-13 | NJ | 131 | YT99-66 | YT |
| 32 | GT02-208 | GT | 82 | NJ86-117 | NJ | 132 | YZ02-2540 | YZ |
| 33 | GT02-281 | GT | 83 | NJ92-244 | NJ | 133 | YZ02-588 | YZ |
| 34 | GT02-467 | GT | 84 | ROC1 | ROC | 134 | YZ03-194 | YZ |
| 35 | GT02-761 | GT | 85 | ROC10 | ROC | 135 | YZ07-100 | YZ |
| 36 | GT02-901 | GT | 86 | ROC16 | ROC | 136 | YZ07-49 | YZ |
| 37 | GT03-11 | GT | 87 | ROC20 | ROC | 137 | YZ89-7 | YZ |
| 38 | GT03-1403 | GT | 88 | ROC22 | ROC | 138 | YZ94-343 | YZ |
| 39 | GT03-2112 | GT | 89 | ROC23 | ROC | 139 | YZ94-375 | YZ |
| 40 | GT03-3005 | GT | 90 | ROC25 | ROC | 140 | YZ99-601 | YZ |
| 41 | GT03-3089 | GT | 91 | ROC26 | ROC | 141 | YZ99-91 | YZ |
| 42 | GT03-8 | GT | 92 | ROC28 | ROC | 142 | ZZ33 | YT |
| 43 | GT03-91 | GT | 93 | YC04-55 | YC | 143 | ZZ41 | YT |
| 44 | GT05-3084 | GT | 94 | YC05-64 | YC | 144 | ZZ43 | YT |
| 45 | GT05-3595 | GT | 95 | YC06-111 | YC | 145 | ZZ45 | YT |
| 46 | GT73-167 | GT | 96 | YC06-140 | YC | 146 | ZZ49 | YT |
| 47 | GT89-5 | GT | 97 | YC06-166 | YC | 147 | ZZ50 | YT |
| 48 | GT92-66 | GT | 98 | YC06-61 | YC | 148 | ZZ80-101 | YT |
| 49 | GT94-119 | GT | 99 | YC06-63 | YC | 149 | ZZ90-76 | YT |
| 50 | GT96-154 | GT | 100 | YC06-91 | YC | 150 | ZZ92-126 | YT |
| Primer Name | Type a | Repeat Motif | Primer Sequence (5′-3′) | Annealing Temperatures (°C) |
|---|---|---|---|---|
| mSSCIR36 | G-SSR | (GA)18GT (GA)4 | CAACAATAACTTAACTGGTA CTGTCCTTTTTATTCTCTTT | 52 |
| mSSCIR46 | G-SSR | (GT)10 | ATGCTCCGCTTCTCACTC AAGGGGAAAATGAAAACC | 52 |
| mSSCIR74 | G-SSR | (CGC)9 | GCGCAAGCCACACTGAGA ACGCAACGCAAAACAACG | 56 |
| SCM4 | E-SSR | (CGGAT)4 | CATTGTTCTGTGCCTGCT CCGTTTCCCTTCCTTCCC | 52 |
| SCM7 | E-SSR | (GCAC)4 | ACGGTGCTCTTCACTGCT GGGCATACTTCCTCCTCTAC | 60 |
| SCM18 | E-SSR | (ATAC)3 | CATCAGTATCATTTCATCTTGG CAGTCACAGTCGGGTAGA | 60 |
| SMC1825LA | G-SSR | (TG)11 | CACGTCCTTCCGCCTTGA TCATCGTTCGTCGCACTG | 56 |
| SMC286CS | G-SSR | (TG)43 | TCAAATGGGACCTTATTGGAG TCCCTCGATCTCCGTTGTT | 52 |
| SMC477CG | G-SSR | (CA)31 | CCAACAACGAATTGTGCATGT CCTGGTTGGCTACCTGTCTTCA | 60 |
| SMC486CG | G-SSR | (CA)14 | GAAATTGCCTCCCAGGATTA CCAACTTGAGAATTGAGATTCG | 60 |
| SMC569CS | G-SSR | (TG)37 | GCGATGGTTCCTATGCAACTT TTCGTGGCTGAGATTCACACTA | 60 |
| SMC597CS | G-SSR | (AG)31 | GCACACCACTCGAATAACGGAT AGTATATCGTCCCTGGCATTCA | 52 |
| SMC334BS | G-SSR | (TG)36 | CAATTCTGACCGTGCAAAGAT CGATGAGCTTGATTGCGAATG | 60 |
| SMC36BUQ | G-SSR | (TTG)7 | GGGTTTCATCTCTAGCCTACC TCAGTAGCAGAGTCAGACGCTT | 56 |
| SMC7CUQ | G-SSR | (CA)10(C)4 | GCCAAAGCAAGGGTCACTAGA AGCTCTATCAGTTGAAACCGA | 60 |
| SEGM285 | G-SSR | (GCAC)4 | AAGAAGAAGACTGAGAAGAACACT TAGCAACAACTTAATTTAGCAATC | 56 |
| UGSM345 | E-SSR | (TG)6 | CTGTACTGGTATTACATGTGACCT TCTACTAATCACAAGAGAAGATGC | 60 |
| UGSM10 | E-SSR | (GGC)11 | GCTACTATGGACAACAGGG ATGAAGAGACGAGACGAAGA | 56 |
| UGSuM50 | E-SSR | (TC)14 | CTACTGCCGAGGAAAGATCG GGAAAAGTTTGTGGCAAGGA | 56 |
| MCSA068G08 | E-SSR | (CAG)6 | CTAATGCCATGCCCCAGAGG GCTGGTGATGTCGCCCATCT | 56 |
| MCSA176C01 | E-SSR | (GGT)5 | GAGTCAGTTGGTGCCGAGATTG GAACAGGTTAAAGCCCATGTC | 56 |
| Primer Name | Allele (No.) | Product Size (bp) | Range of PIC a Values | Mean of PIC Values | PIb | RPc |
|---|---|---|---|---|---|---|
| mSSCIR36 | 21 | 127–168 | 0.01–0.38 | 0.15 | 0.000001 | 7.09 |
| mSSCIR46 | 12 | 146–177 | 0.01–0.37 | 0.15 | 0.002858 | 13.04 |
| mSSCIR74 | 6 | 215–228 | 0.00–0.37 | 0.17 | 0.042135 | 4.69 |
| SCM4 | 25 | 92–209 | 0.01–0.37 | 0.17 | 0.000087 | 4.16 |
| SCM7 | 7 | 155–188 | 0.03–0.37 | 0.18 | 0.048672 | 3.68 |
| SCM18 | 9 | 226–251 | 0.00–0.38 | 0.19 | 0.010157 | 8.67 |
| SMC1825LA | 10 | 91–119 | 0.01–0.37 | 0.20 | 0.001240 | 6.53 |
| SMC286CS | 13 | 128–152 | 0.01–0.37 | 0.21 | 0.000411 | 7.31 |
| SMC477CG | 15 | 115–134 | 0.00–0.36 | 0.21 | 0.000125 | 4.11 |
| SMC486CG | 7 | 222–243 | 0.06–0.36 | 0.22 | 0.051066 | 4.88 |
| SMC569CS | 6 | 166–220 | 0.04–0.38 | 0.24 | 0.071332 | 14.05 |
| SMC597CS | 14 | 143–166 | 0.03–0.37 | 0.24 | 0.000034 | 10.99 |
| SMC334BS | 12 | 145–163 | 0.01–0.38 | 0.24 | 0.000140 | 6.27 |
| SMC36BUQ | 12 | 103–251 | 0.00–0.37 | 0.25 | 0.010448 | 7.49 |
| SMC7CUQ | 7 | 156–170 | 0.00–0.37 | 0.26 | 0.024118 | 9.76 |
| SEGM285 | 13 | 306–389 | 0.03–0.38 | 0.26 | 0.000143 | 21.01 |
| UGSM345 | 8 | 320–334 | 0.01–0.38 | 0.27 | 0.005772 | 13.68 |
| UGSM10 | 10 | 97–125 | 0.00–0.38 | 0.28 | 0.005289 | 9.31 |
| UGSuM50 | 6 | 123–139 | 0.05–0.38 | 0.28 | 0.023095 | 6.24 |
| MCSA068G08 | 8 | 179–202 | 0.06–0.38 | 0.29 | 0.003035 | 15.57 |
| MCSA176C01 | 5 | 427–440 | 0.11–0.38 | 0.29 | 0.026013 | 13.31 |
| Series | Sample Size | ha | Ib | PPLc | Series-Specific Alleles |
|---|---|---|---|---|---|
| CP | 29 | 0.239 | 0.361 | 0.743 | SCM7-188, SCM18-238, SMC486CG-225, SMC486CG-233 |
| DZ | 4 | 0.235 | 0.341 | 0.562 | |
| FN | 11 | 0.245 | 0.365 | 0.677 | mSSCIR46-153 |
| GN | 2 | 0.148 | 0.205 | 0.296 | |
| GT | 21 | 0.251 | 0.376 | 0.721 | |
| LC | 6 | 0.197 | 0.293 | 0.522 | |
| NJ | 6 | 0.205 | 0.302 | 0.527 | SMC36BUQ-125 |
| ROC | 9 | 0.201 | 0.301 | 0.558 | SMC36BUQ-184, SEGM285-359 |
| K | 2 | 0.164 | 0.227 | 0.327 | |
| YC | 18 | 0.261 | 0.397 | 0.805 | mSSCIR46-146, mSSCIR46-149, SCM7-175, SMC569CS-174, SMC569CS-202, SMC36BUQ-106, SMC36BUQ-132, UGSM10-113 |
| YT | 29 | 0.254 | 0.386 | 0.814 | SMC36BUQ-105, SMC36BUQ-139 |
| YZ | 10 | 0.241 | 0.358 | 0.650 | |
| Mean | 0.176 | 0.261 | 0.480 |
| Source of Variation | Degrees of Freedom | Sum of Squares | Mean Sum of Squares | Estimated Variance | Percentage of Variation |
|---|---|---|---|---|---|
| Between sub-Pops | 1 | 546.240 | 546.240 | 6.308 | 5% |
| Within sub-Pop | 148 | 18,601.600 | 125.686 | 125.686 | 95% |
| Total | 149 | 19,147.840 | 131.995 | 100% | |
| Fixation Index | Fst = 0.048 | ||||
| Gene Flow | Nm = 4.981 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Wang, Q.; Xie, J.; Pan, Y.-B.; Zhou, F.; Guo, Y.; Chang, H.; Xu, H.; Zhang, W.; Zhang, C.; et al. SSR Marker-Assisted Management of Parental Germplasm in Sugarcane (Saccharum spp. hybrids) Breeding Programs. Agronomy 2019, 9, 449. https://doi.org/10.3390/agronomy9080449
Wu J, Wang Q, Xie J, Pan Y-B, Zhou F, Guo Y, Chang H, Xu H, Zhang W, Zhang C, et al. SSR Marker-Assisted Management of Parental Germplasm in Sugarcane (Saccharum spp. hybrids) Breeding Programs. Agronomy. 2019; 9(8):449. https://doi.org/10.3390/agronomy9080449
Chicago/Turabian StyleWu, Jiantao, Qinnan Wang, Jing Xie, Yong-Bao Pan, Feng Zhou, Yuqiang Guo, Hailong Chang, Huanying Xu, Wei Zhang, Chuiming Zhang, and et al. 2019. "SSR Marker-Assisted Management of Parental Germplasm in Sugarcane (Saccharum spp. hybrids) Breeding Programs" Agronomy 9, no. 8: 449. https://doi.org/10.3390/agronomy9080449
APA StyleWu, J., Wang, Q., Xie, J., Pan, Y.-B., Zhou, F., Guo, Y., Chang, H., Xu, H., Zhang, W., Zhang, C., & Qiu, Y. (2019). SSR Marker-Assisted Management of Parental Germplasm in Sugarcane (Saccharum spp. hybrids) Breeding Programs. Agronomy, 9(8), 449. https://doi.org/10.3390/agronomy9080449