Distributions of Alien Invasive Weeds under Climate Change Scenarios in Mountainous Bhutan




Abstract
1. Introduction
2. Materials and Methods
2.1. Defining Survey Site and Species Record Sampling
2.2. Environmental Data Processing
2.3. Model Calibration
2.4. Statistical Testing of Model Predicted Areas of Invasion
2.5. Tracking Directional Distributions between Two Climatic Conditions
3. Results and Discussion
3.1. Model Evaluation
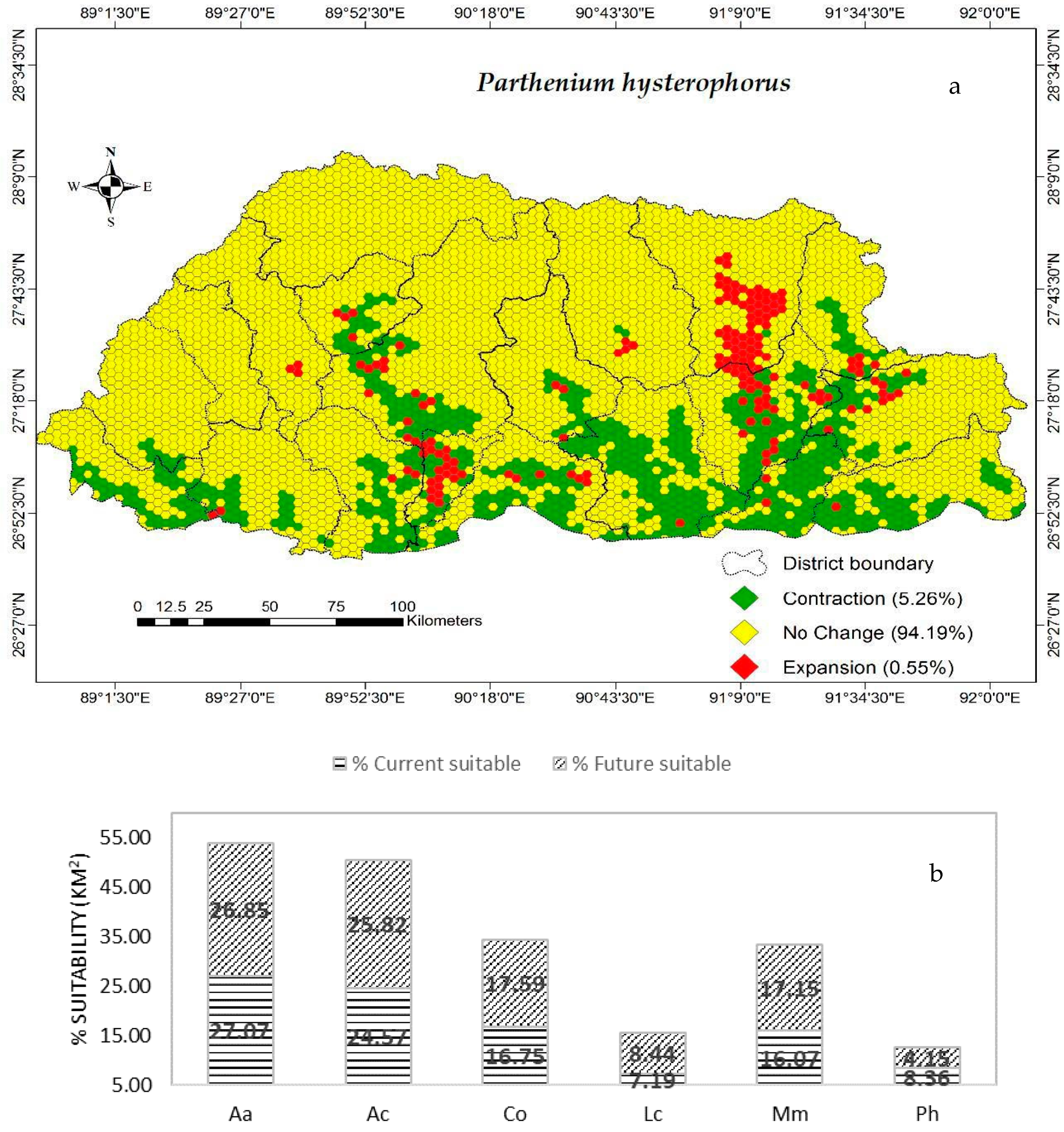
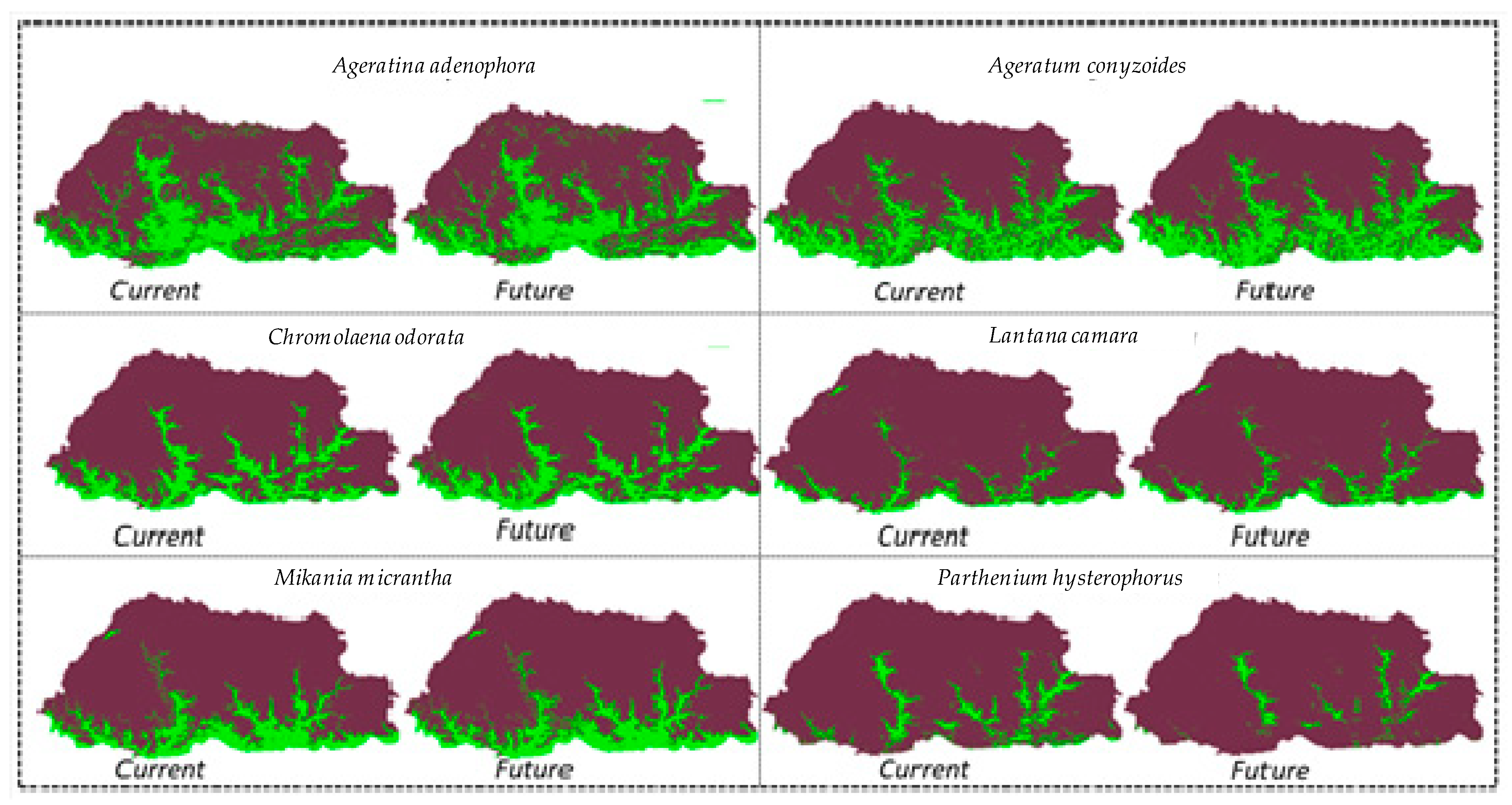
3.2. Spatial Change Analysis
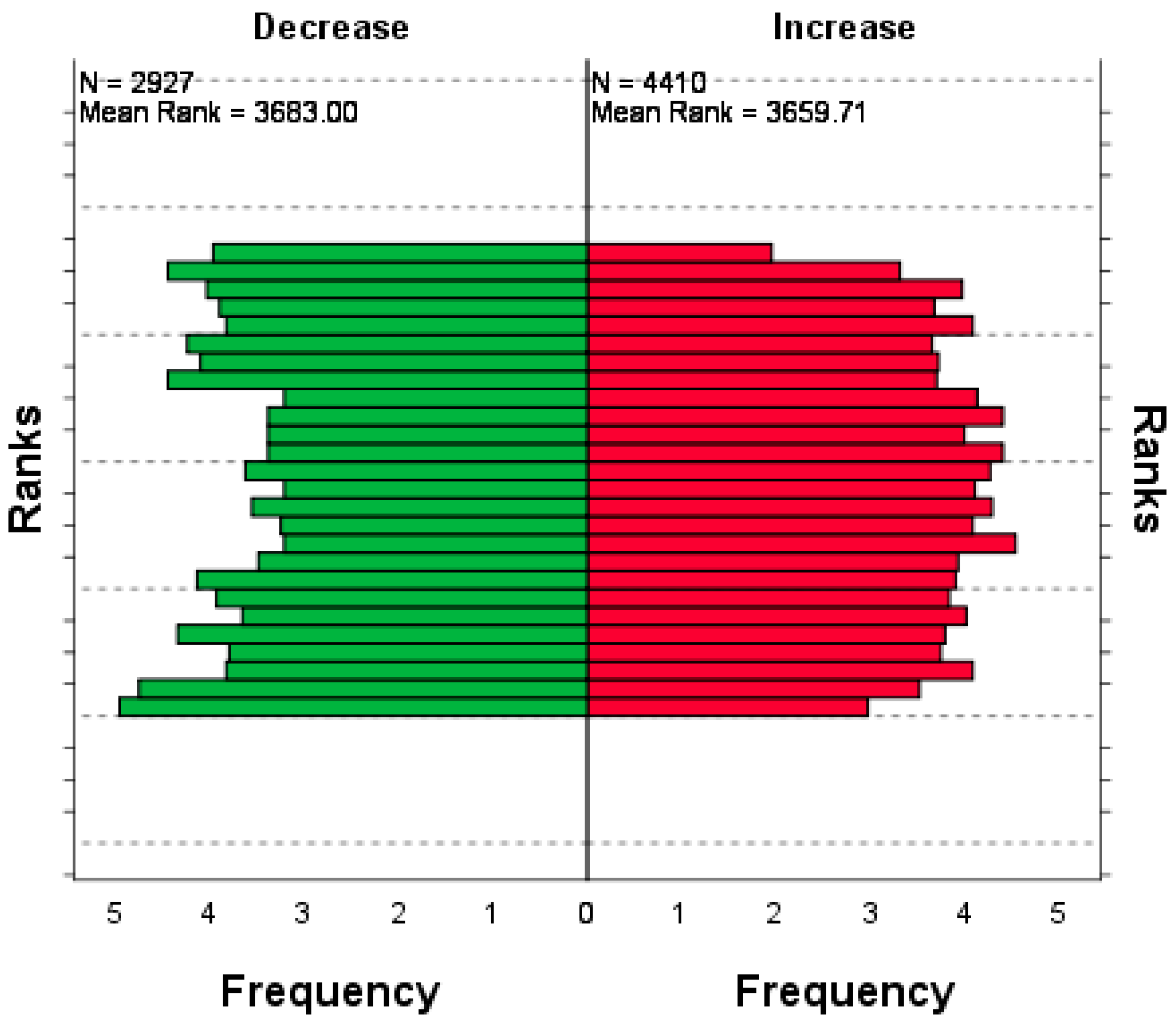
3.3. Testing the Statistical Significance of Changes
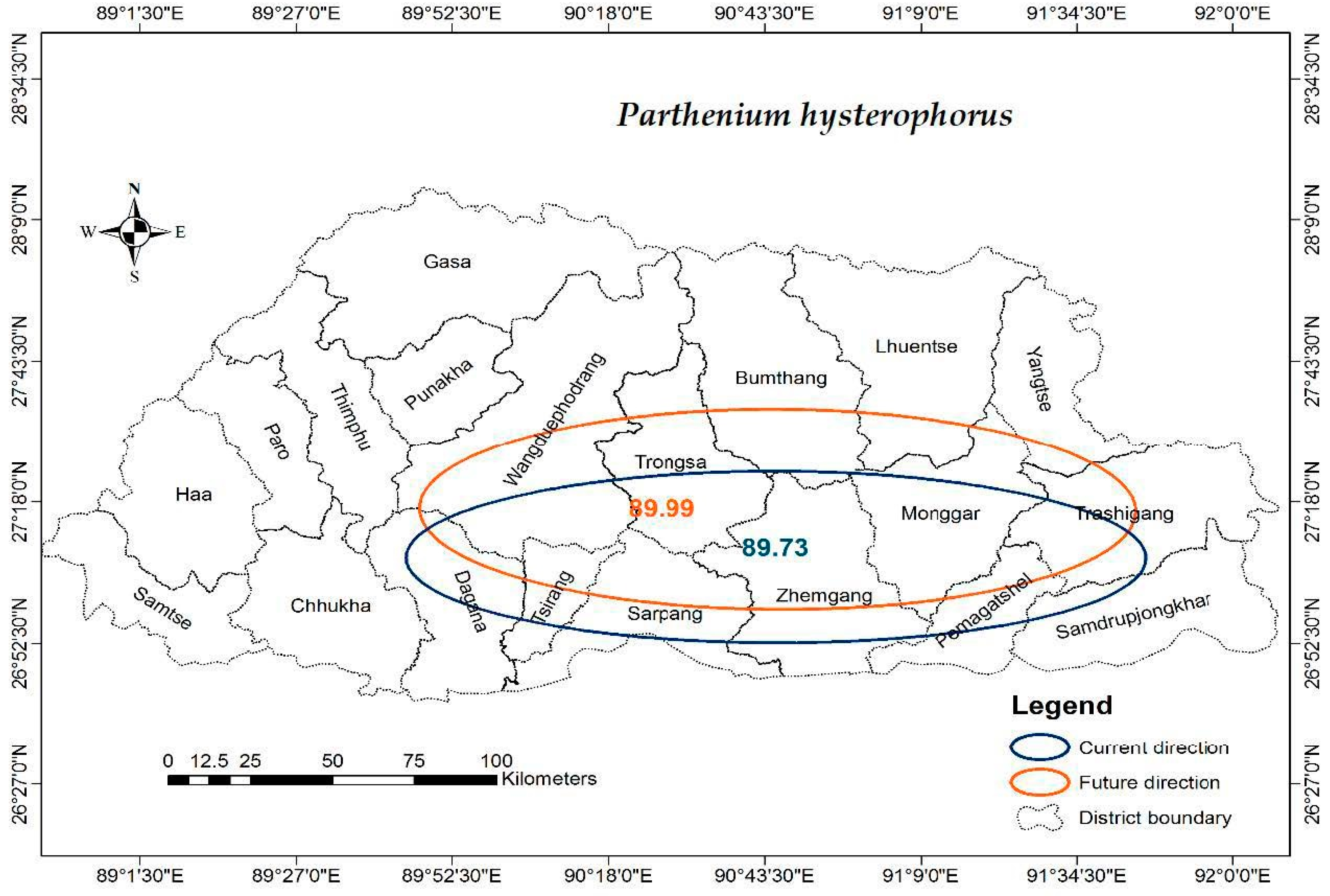
3.4. Cardinal Directions of Distributions
3.5. Limitations and Caveats of the Model
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | Variable | Scaling Factor | Units |
|---|---|---|---|
| BIO1 | Annual mean Temperature | 10 | Degree Celsius |
| BIO2 | Mean Diurnal Range (Mean of monthly (max temp-min temp)) | 10 | Degree Celsius |
| BIO3 | Isothermality (BIO2/BIO7) | 100 | Degree Celsius |
| BIO4 | Temperature seasonality (standard deviation) | 100 | Degree Celsius |
| BIO5 | Max Temperature of Warmest Month | 10 | Degree Celsius |
| BIO6 | Min Temperature of Coldest Month | 10 | Degree Celsius |
| BIO7 | Temperature annual Range (BIO5-BIO6) | 10 | Degree Celsius |
| BIO8 | Mean Temperature of Wettest Quarter | 10 | Degree Celsius |
| BIO9 | Mean Temperature of Driest Quarter | 10 | Degree Celsius |
| BIO10 | Mean Temperature of Warmest Quarter | 10 | Degree Celsius |
| BIO11 | Mean Temperature of Coldest Quarter1 | 10 | Degree Celsius |
| BIO12 | Annual 1Precipitation | 1 | Millimetres |
| BIO13 | Precipitation of Wettest Month | 1 | Millimetres |
| BIO14 | Precipitation of Driest Month | 1 | Millimetres |
| BIO15 | Precipitation Seasonality (Coefficient of variation) | 100 | Fraction |
| BIO16 | Precipitation of Wettest Quarter | 1 | Millimetres |
| BIO17 | Precipitation of Driest Quarter | 1 | Millimetres |
| BIO18 | Precipitation of Warmest Quarter | 1 | Millimetres |
| BIO19 | Precipitation of Coldest Quarter | 1 | Millimetres |

References
- Ihlow, F.; Courant, J.; Secondi, J.; Herrel, A.; Rebelo, R.; Measey, G.J.; Lillo, F.; De Villiers, F.A.; Vogt, S.; De Busschere, C.; et al. Impacts of climate change on the global invasion potential of the African clawed frog Xenopus laevis. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [PubMed]
- Lourenco-de-Moraes, R.; Lansac-Toha, F.M.; Schwind, L.T.F.; Arrieira, R.L.; Rosa, R.R.; Terribile, L.C.; Lemess, P.; Rangel, T.F.; Diniz, J.A.F.; Bastosz, R.P.; et al. Climate change will decrease the range size of snake species under negligible protection in the Brazilian Atlantic forest hotspot. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, J.; Singh, C.P.; Hamid, M.; Verma, A.; Semwal, S.C.; Gajmer, B.; Khuroo, A.A.; Kumar, A.; Nautiyal, M.C.; Sharma, N.; et al. Modelling Betula utilis distribution in response to climate-warming scenarios in Hindu-Kush Himalaya using random forest. Biodivers. Conserv. 2019, 28, 2295–2317. [Google Scholar] [CrossRef]
- Priyanka, N.; Joshi, P.K. Effects of climate change on invasion potential distribution of Lantana camara. J. Earth Sci. Clim. Chang. 2013, 4, 164. [Google Scholar] [CrossRef]
- Rathore, P.; Roy, A.; Karnatak, H. Assessing the vulnerability of Oak (Quercus) forest ecosystems under projected climate and land use land cover changes in Western Himalaya. Biodivers. Conserv. 2019, 28, 2275–2294. [Google Scholar] [CrossRef]
- Ruaro, R.; Conceicao, E.O.; Silva, J.C.; Cafofo, E.G.; Angulo-Valencia, M.A.; Mantovano, T.; Pineda, A.; de Paula, A.C.M.; Zanco, B.F.; Capparros, E.M.; et al. Climate change will decrease the range of a keystone fish species in La Plata river basin, South America. Hydrobiologia 2019, 836, 1–19. [Google Scholar] [CrossRef]
- Saupe, E.E.; Myers, C.E.; Peterson, A.T.; Soberon, J.; Singarayer, J.; Valdes, P.; Qiao, H.J. Non-random latitudinal gradients in range size and niche breadth predicted by spatial patterns of climate. Glob. Ecol. Biogeogr. 2019, 28, 928–942. [Google Scholar] [CrossRef]
- Wang, C.J.; Wan, J.Z.; Zhang, Z.X. Will Global climate change facilitate plant invasions in conservation areas? Pak. J. Bot. 2019, 51, 1395–1403. [Google Scholar] [CrossRef]
- Bugmann, H.; Gurung, A.B.; Ewert, F.; Haeberli, W.; Guisan, A.; Fagre, D.; Kaab, A.; Participants, G. Modeling the biophysical impacts of global change in mountain biosphere reserves. Mt. Res. Dev. 2007, 27, 66–77. [Google Scholar] [CrossRef]
- De la Hoz, C.F.; Ramos, E.; Puente, A.; Juanes, J.A. Climate change induced range shifts in seaweeds distributions in Europe. Mar. Environ. Res. 2019, 148, 1–11. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Global Warming of 1.5 C; Report; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- Geng, Y.P.; Pan, X.Y.; Xu, C.Y.; Zhang, W.J.; Li, B.; Chen, J.K. Phenotypic plasticity of invasive Alternanthera philoxeroides in relation to different water availability, compared to its native congener. Acta Oecol. 2006, 30, 380–385. [Google Scholar] [CrossRef]
- Keser, L.H.; Dawson, W.; Song, Y.B.; Yu, F.H.; Fischer, M.; Dong, M.; van Kleunen, M. Invasive clonal plant species have a greater root-foraging plasticity than non-invasive ones. Oecologia 2014, 174, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Oduor, A.M.; Zhang, Z.; Manea, A.; Tooth, I.M.; Leishman, M.R.; Xu, X.; Van Kleunen, M. Do invasive alien plants benefit more from global environmental change than native plants? Glob. Chang. Biol. 2017, 23, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Chown, S.L.; Hodgins, K.A.; Griffin, P.C.; Oakeshott, J.G.; Byrne, M.; Hoffmann, A.A. Biological invasions, climate change and genomics. Evol. Appl. 2015, 8, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Bradley, B.A.; Wilcove, D.S.; Oppenheimer, M. Climate change increases risk of plant invasion in the Eastern United States. Biol. Invasions 2010, 12, 1855–1872. [Google Scholar] [CrossRef]
- Keller, S.; Taylor, D. Genomic admixture increases fitness during a biological invasion. J. Evol. Biol. 2010, 23, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Tecco, P.A.; Pais-Bosch, A.I.; Funes, G.; Marcora, P.I.; Zeballos, S.R.; Cabido, M.; Urcelay, C. Mountain invasions on the way: Are there climatic constraints for the expansion of alien woody species along an elevation gradient in Argentina? J. Plant Ecol. 2015, 9, 380–392. [Google Scholar] [CrossRef]
- Eckholm, E.P. The deterioration of mountain environments. Science 1975, 189, 764–770. [Google Scholar] [CrossRef]
- McDougall, K.L.; Khuroo, A.A.; Loope, L.L.; Parks, C.G.; Pauchard, A.; Reshi, Z.A.; Rushworth, I.; Kueffer, C. Plant Invasions in mountains: Global lessons for better management. Mt. Res. Dev. 2011, 31, 380–387. [Google Scholar] [CrossRef]
- Lamsal, P.; Kumar, L.; Aryal, A.; Atreya, K. Invasive alien plant species dynamics in the Himalayan region under climate change. Ambio 2018, 47, 697–710. [Google Scholar] [CrossRef]
- Shrestha, U.B.; Sharma, K.P.; Devkota, A.; Siwakoti, M.; Shrestha, B.B. Potential impact of climate change on the distribution of six invasive alien plants in Nepal. Ecol. Indic. 2018, 95, 99–107. [Google Scholar] [CrossRef]
- Thapa, S.; Chitale, V.; Rijal, S.J.; Bisht, N.; Shrestha, B.B. Understanding the dynamics in distribution of invasive alien plant species under predicted climate change in Western Himalaya. PLoS ONE 2018, 13, e0195752. [Google Scholar] [CrossRef] [PubMed]
- Hoy, A.; Katel, O. Status of Climate Change and Implications to Ecology and Community Livelihoods in the Bhutan Himalaya. In Environmental Change in the Himalayan Region; Springer: Cham, Switzerland, 2019; pp. 23–45. [Google Scholar]
- Penuelas, J.; Boada, M. A global change-induced biome shift in the Montseny mountains (NE Spain). Glob. Chang. Biol. 2003, 9, 131–140. [Google Scholar] [CrossRef]
- Suberi, B.; Tiwari, K.R.; Gurung, D.B.; Bajracharya, R.M.; Sitaula, B.K. People’s perception of climate change impacts and their adaptation practices in Khotokha valley, Wangdue, Bhutan. Indian J. Tradit. Knowl. 2018, 17, 97–105. [Google Scholar]
- McDougall, K.L.; Alexander, J.M.; Haider, S.; Pauchard, A.; Walsh, N.G.; Kueffer, C. Alien flora of mountains: Global comparisons for the development of local preventive measures against plant invasions. Divers. Distrib. 2011, 17, 103–111. [Google Scholar] [CrossRef]
- Seldon, P. First ever Bhutan Climate Report Predicts a Hotter and Wetter Bhutan. The Bhutanese. Available online: https://thebhutanese.bt/first-ever-bhutan-climate-report-predicts-a-hotter-and-wetter-bhutan/ (accessed on 3 March 2019).
- Fort, M. Impact of climate change on mountain environment dynamics. An introduction. J. Alp. Res. 2015. [Google Scholar] [CrossRef]
- Beniston, M. Mountain Environments in Changing Climates; Beniston, M., Ed.; Routledge: London, UK; New York, NY, USA, 2002. [Google Scholar]
- International Centre for Integrated Mountain Development (Nepal) (ICIMOD). Climate Change Impacts and Vulnerabilities in the Eastern Himalayas; ICIMOD: Lalitpur, Nepal, 2009. [Google Scholar]
- Malanson, G.P.; Fagre, D.B. Spatial contexts for temporal variability in alpine vegetation under ongoing climate change. Plant Ecol. 2013, 214, 1309–1319. [Google Scholar] [CrossRef]
- Williamson, M.H.; Fitter, A. The characters of successful invaders. Biol. Conserv. 1996, 78, 163–170. [Google Scholar] [CrossRef]
- Rejmanek, M.; Richardson, D.M. What attributes make some plant species more invasive? Ecology 1996, 77, 1655–1661. [Google Scholar] [CrossRef]
- Sutherland, S. What makes a weed a weed: Life history traits of native and exotic plants in the USA. Oecologia 2004, 141, 24–39. [Google Scholar] [CrossRef]
- Devin, S.; Beisel, J.N. Biological and ecological characteristics of invasive species: A gammarid study. Biol. Invasions 2007, 9, 13–24. [Google Scholar] [CrossRef]
- Maron, J.L.; Vilà, M.; Bommarco, R.; Elmendorf, S.; Beardsley, P. Rapid evolution of an invasive plant. Ecol. Monogr. 2004, 74, 261–280. [Google Scholar] [CrossRef]
- Higgins, S.I.; Richardson, D.M. Invasive plants have broader physiological niches. Proc. Natl. Acad. Sci. USA 2014, 111, 10610–10614. [Google Scholar] [CrossRef] [PubMed]
- Funk, J.L. The physiology of invasive plants in low-resource environments. Conserv. Physiol. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Royal Government of Bhutan. Forest and Nature Conservation Rules and Regulations of Bhutan; Department of Forest and Park Services: Thimphu, Butan, 2017.
- Chhetri, P.B.; Tenzin, K. Bhutan: The State of the World’s Forest Genetic Resources—FAO; RNR Research and Development Centre: Yusipang, Bhutan, 2012.
- National Biodiversity Centre (NBC). Bhutan-Biodiversity Action Plan 2009—UNDP in Bhutan; NBC: Thimphu, Bhutan, 2009. [Google Scholar]
- Grierson, A.J.C.; Long, D.G. Flora of Bhutan. NORDIC J. Bot. 2001, 2, 456. [Google Scholar] [CrossRef]
- Parker, C. Weeds of Bhutan; National Palnt Protection Centre: Thimphu, Bhutan, 1992.
- Bhutan Biodiversity Portal. Available online: https://biodiversity.bt/ (accessed on 1 August 2019).
- Pallewatta, N.; Reaser, J.; Gutierrez, A. Invasive Alien Species in South-Southeast Asia: National Reports and Directory of Resources; Global Invasive Species Programme: Cape Town, South Africa, 2003. [Google Scholar]
- Tallis, H.; Kareiva, P.; Marvier, M.; Chang, A. An ecosystem services framework to support both practical conservation and economic development. Proc. Natl. Acad. Sci. USA 2008, 105, 9457–9464. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Navarro-Sigüenza, A.G.; Martínez-Meyer, E.; Cuervo-Robayo, A.P.; Berlanga, H.; Soberón, J. Twentieth century turnover of Mexican endemic avifaunas: Landscape change versus climate drivers. Sci. Adv. 2015, 1, e1400071. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, M.R.; Deb, P.; Singha, H.; Chakdar, B.; Medhi, M. Predicting the probable distribution and threat of invasive Mimosa diplotricha Suavalle and Mikania micrantha Kunth in a protected tropical grassland. Ecol. Eng. 2016, 97, 23–31. [Google Scholar] [CrossRef]
- Keith, D.A.; Akcakaya, H.R.; Thuiller, W.; Midgley, G.F.; Pearson, R.G.; Phillips, S.J.; Regan, H.M.; Araujo, M.B.; Rebelo, T.G. Predicting extinction risks under climate change: Coupling stochastic population models with dynamic bioclimatic habitat models. Biol. Lett. 2008, 4, 560–563. [Google Scholar] [CrossRef] [PubMed]
- López-Darias, M.; Lobo, J.M.; Gouat, P. Predicting potential distributions of invasive species: The exotic Barbary ground squirrel in the Canarian archipelago and the west Mediterranean region. Biol. Invasions 2007, 10, 1027–1040. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Stiels, D.; Schidelko, K.; Engler, J.O.; van den Elzen, R.; Rödder, D. Predicting the potential distribution of the invasive Common Waxbill Estrilda astrild (Passeriformes: Estrildidae). J. Ornithol. 2011, 152, 769–780. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Merow, C.; Allen, J.M.; Aiello-Lammens, M.; Silander, J.A. Improving niche and range estimates with Maxent and point process models by integrating spatially explicit information. Glob. Ecol. Biogeogr. 2016, 25, 1022–1036. [Google Scholar] [CrossRef]
- Ray, D.; Behera, M.D.; Jacob, J. Evaluating Ecological Niche Models: A Comparison Between Maxent and GARP for Predicting Distribution of Hevea brasiliensis in India. Proc. Indian Natl. Sci. Acad. Part B Biol. Sci. 2018, 88, 1337–1343. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, S.; Sun, P.; Wang, T.; Wang, G.; Zhang, X.; Wang, L. Consensus forecasting of species distributions: The effects of niche model performance and niche properties. PLoS ONE 2015, 10, e0120056. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Ashraf, U.; Peterson, A.T.; Chaudhry, M.N.; Ashraf, I.; Saqib, Z.; Ahmad, S.R.; Ali, H. Ecological niche model comparison under different climate scenarios: A case study of Olea spp. in Asia. Ecosphere 2017, 8, e01825. [Google Scholar] [CrossRef]
- Kramer-Schadt, S.; Niedballa, J.; Pilgrim, J.D.; Schröder, B.; Lindenborn, J.; Reinfelder, V.; Stillfried, M.; Heckmann, I.; Scharf, A.K.; Augeri, D.M. The importance of correcting for sampling bias in MaxEnt species distribution models. Divers. Distrib. 2013, 19, 1366–1379. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Ohsawa, M.E. Life Zone Ecology of the Bhutan Himalaya; Laboratory of Ecology, Chiba University: Chiba, Japan, 1987. [Google Scholar]
- Pearson, R.G. Species’ Distribution Modeling for Conservation Educators and Practitioners. Lessons Conserv. 2008, 3, 54–89. [Google Scholar]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Hijmans, R.; Cameron, S.; Parra, J.; Jones, P.; Jarvis, A.; Richardson, K. WorldClim, version 1.3; University of California: Berkeley, CA, USA, 2005. [Google Scholar]
- Phillips, S.J.; Dudik, M.; Schapire, R. MaxEnt, Version 3.3. 3k; AT & T Labs-Research, Princeton University: Princeton, NJ, USA, 2012. [Google Scholar]
- Environmental Systems Research Institute (ESRI). ArcGIS Desktop: Release 10.1. Available online: https://www.esri.com/news/arcnews/spring12articles/introducing-arcgis-101.html (accessed on 26 January 2015).
- Pitt, J.P.W.; Kriticos, D.J.; Dodd, M.B. Temporal limits to simulating the future spread pattern of invasive species: Buddleja davidii in Europe and New Zealand. Ecol. Model. 2011, 222, 1880–1887. [Google Scholar] [CrossRef]
- Wan, F.; Liu, W.; Guo, J.; Qiang, S.; Li, B.; Wang, J.; Yang, G.; Niu, H.; Gui, F.; Huang, W. Invasive mechanism and control strategy of Ageratina adenophora (Sprengel). Sci. China Life Sci. 2010, 53, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- He, J.Y.; Qiang, S.; Song, X.L.; Jin, H.Y. Comparison of the stem and leaf morphological structures of 18 communities of the foreign plant Eupatorium adenophorum. Acta Bot. Sin. 2005, 25, 1089. [Google Scholar]
- Kaur, A.; Batish, D.R.; Kaur, S.; Singh, H.P.; Kohli, R.K. Phenological behaviour of Parthenium hysterophorus in response to climatic variations according to the extended BBCH scale. Ann. Appl. Biol. 2017, 171, 316–326. [Google Scholar] [CrossRef]
- Ura, K.; Kinga, S. Bhutan—Sustainable Development through Good Governance; World Bank: Washington, DC, USA, 2004. [Google Scholar]
- Regmi, B.; Pandit, A.; Pradhan, B.; Kovats, S.; Lama, P. Capacity Strengthening in the Least Developed Countries (LDCs) for Adaptation to Climate Change (CLACC), Climate Change and Health in Nepal. Available online: https://pubs.iied.org/pdfs/G02664.pdf? (accessed on 1 May 2019).
- Penjore, D.; Rapten, P. Trends of forestry policy concerning local participation in Bhutan. Forest 2004, 25, 21–27. [Google Scholar]
- Department of Forests and Park Services. Forest Facts and Figures. Available online: http://www.dofps.gov.bt/wp-content/uploads/2017/07/ForestBookletFinal.pdf (accessed on 18 May 2019).
- Franklin, J. Moving beyond static species distribution models in support of conservation biogeography. Divers. Distrib. 2010, 16, 321–330. [Google Scholar] [CrossRef]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Mechanistic and correlative models of ecological niches. Eur. J. Ecol. 2015, 1, 28–38. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distribution; Cambridge University Press: New York, NY, USA, 2009. [Google Scholar]
- Jarnevich, C.S.; Stohlgren, T.J.; Kumar, S.; Morisette, J.T.; Holcombe, T.R. Caveats for correlative species distribution modeling. Ecol. Inform. 2015, 29, 6–15. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Li, J.; Peterson, A.T.; Graham, C.H.; Guisan, A. NCEAS Predicting Species Distributions Working Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar] [CrossRef]
- Guisan, A.; Graham, C.H.; Elith, J.; Huettmann, F.; NCEAS Species Distribution Modelling Group. Sensitivity of predictive species distribution models to change in grain size. Divers. Distrib. 2007, 13, 332–340. [Google Scholar] [CrossRef]
- Naimi, B.; Skidmore, A.K.; Groen, T.A.; Hamm, N.A.S. Spatial autocorrelation in predictors reduces the impact of positional uncertainty in occurrence data on species distribution modeling. J. Biogeogr. 2011, 38, 1497–1509. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]



| Species | Native Range | Invaded Area in Bhutan | Impact | Reference |
|---|---|---|---|---|
| Ageratina adenophora | Mexico | Punakha, Trongsa, Samtse, Wangdue, and Phuentsholing | Toxic to animal health, and displaces native vegetation | Grierson and Long [43] |
| Ageratum conyzoides | Central America and South America | Mongar, Sarpang, and Wangdue | Causes liver cancer in humans and inhibits the growth of other plants | Parker [44] |
| Chromolaena odorata | Southeastern USA, Mexico, and South America | Punakha, Trongsa Wangdue, Mongar, and Phuentsholing | Toxic to animal health and causes asthma in humans | Bhutan Biodiversity Portal [45] |
| Lantana camara | Mexico, Central America, the Caribbean, and tropical South America | Wangdue and Phuentsholing | Causes allergenic rhinititis, a type of child cirrhosis via pollen-contaminated milk | Bhutan Biodiversity Portal [45] |
| Mikania micrantha | Mexico, Central America, the Caribbean, and tropical South America | Chhukha, Mongar, Samtse, and Sarpang | Can damage perennial crops by twinning and entanglement, and reduces biodiversity through competition | Parker [44] |
| Parthenium hysterophorus | Mexico, Central America, the Caribbean, and South America | Mongar, Tashigang, Tongsa, and Wangdi | Causes severe allergic illness in adult human males, when in contact with the pollen. | Parker [44] |
| Species | Pre-Survey | Post-Survey | Rarified Record |
|---|---|---|---|
| Ageratina adenophora | 132 | 336 | 48 |
| Ageratum conyzoides | 170 | 172 | 27 |
| Chromolaena odorata | 200 | 89 | 30 |
| Lantana camara | 200 | 71 | 18 |
| Mikania micrantha | 200 | 147 | 26 |
| Parthenium hysterophorus | 282 | 167 | 20 |
| Total | 1184 | 982 | 169 |
| Bio | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | Avg |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 0.9 | 0 | 1 | 1 | 1 | 0.9 | 1 | 1 | 1 | 1 | 0.9 | 0.9 | 0.9 | 0.3 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | |
| 2 | 0.3 | 1 | 0.9 | 0.9 | 1 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | 0.2 | 0.9 | 1 | 1 | 0.9 | 0.8 | ||
| 3 | 0.1 | 0.1 | 0 | 0.2 | 0 | 0 | 0 | 0 | 0.2 | 0.2 | 0.1 | 0.3 | 0.2 | 0.2 | 0.2 | 0.2 | 0.1 | |||
| 4 | 0.9 | 1 | 1 | 0.9 | 1 | 0.9 | 1 | 1 | 0.9 | 0.9 | 0.2 | 1 | 1 | 1 | 0.9 | 0.9 | ||||
| 5 | 1 | 0.9 | 1 | 1 | 1 | 1 | 0.9 | 0.9 | 0.9 | 0.3 | 0.9 | 0.9 | 0.9 | 0.8 | 0.9 | |||||
| 6 | 1 | 1 | 1 | 1 | 1 | 0.9 | 0.9 | 0.9 | 0.3 | 0.9 | 1 | 0.9 | 0.9 | 0.9 | ||||||
| 7 | 0.9 | 1 | 0.9 | 0.9 | 1 | 0.9 | 0.9 | 0.2 | 0.9 | 1 | 1 | 0.9 | 0.9 | |||||||
| 8 | 1 | 1 | 1 | 0.9 | 0.9 | 0.9 | 0.3 | 0.9 | 0.9 | 0.9 | 0.8 | 0.9 | ||||||||
| 9 | 1 | 1 | 0.9 | 0.9 | 0.9 | 0.3 | 0.9 | 0.9 | 0.9 | 0.9 | 0.9 | |||||||||
| 10 | 1 | 0.9 | 0.9 | 0.9 | 0.3 | 0.9 | 0.9 | 0.9 | 0.8 | 0.8 | ||||||||||
| 11 | 0.9 | 0.9 | 0.9 | 0.3 | 0.9 | 0.9 | 0.9 | 0.9 | 0.8 | |||||||||||
| 12 | 1 | 0.9 | 0.1 | 1 | 0.9 | 1 | 0.9 | 0.8 | ||||||||||||
| 13 | 0.9 | 0 | 1 | 0.9 | 1 | 0.8 | 0.8 | |||||||||||||
| 14 | 0.2 | 0.9 | 0.9 | 0.9 | 0.9 | 0.8 | ||||||||||||||
| 15 | 0 | 0.3 | 0 | 0.4 | 0.2 | |||||||||||||||
| 16 | 0.9 | 1 | 0.8 | 0.9 | ||||||||||||||||
| 17 | 0.9 | 1 | 0.9 | |||||||||||||||||
| 18 | 0.8 | 0.8 | ||||||||||||||||||
| 19 | 0.8 |
| Species | N | Ratio AUCc | Ratio AUC f | PAUCc | PAUCf | FAUCc |
| Ageratina adenophora | 48 | 1.76 ± 0.05 | 1.71 ± 0.07 | 0.88 | 0.86 | 0.79 |
| Ageratum conyzoides | 27 | 1.76 ± 0.08 | 1.74 ± 0.08 | 0.88 | 0.87 | 0.77 |
| Chromolaena odorata | 30 | 1.82 ± 0.09 | 1.76 ± 0.09 | 0.91 | 0.88 | 0.83 |
| Lantana camara | 18 | 1.93 ± 0.04 | 1.92 ± 0.04 | 0.97 | 0.96 | 0.86 |
| Mikania micrantha | 26 | 1.85 ± 0.05 | 1.81 ± 0.04 | 0.93 | 0.90 | 0.81 |
| Parthenium hysterophorus | 20 | 1.94 ± 0.03 | 1.92 ± 0.03 | 0.97 | 0.96 | 0.93 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thiney, U.; Banterng, P.; Gonkhamdee, S.; Katawatin, R. Distributions of Alien Invasive Weeds under Climate Change Scenarios in Mountainous Bhutan. Agronomy 2019, 9, 442. https://doi.org/10.3390/agronomy9080442
Thiney U, Banterng P, Gonkhamdee S, Katawatin R. Distributions of Alien Invasive Weeds under Climate Change Scenarios in Mountainous Bhutan. Agronomy. 2019; 9(8):442. https://doi.org/10.3390/agronomy9080442
Chicago/Turabian StyleThiney, Ugyen, Poramate Banterng, Santimaitree Gonkhamdee, and Roengsak Katawatin. 2019. "Distributions of Alien Invasive Weeds under Climate Change Scenarios in Mountainous Bhutan" Agronomy 9, no. 8: 442. https://doi.org/10.3390/agronomy9080442
APA StyleThiney, U., Banterng, P., Gonkhamdee, S., & Katawatin, R. (2019). Distributions of Alien Invasive Weeds under Climate Change Scenarios in Mountainous Bhutan. Agronomy, 9(8), 442. https://doi.org/10.3390/agronomy9080442