Different LED Light Intensities and 6-Benzyladenine Concentrations in Relation to Shoot Development, Leaf Architecture, and Photosynthetic Pigments of Gerbera jamesonii Bolus In Vitro



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experiments Design
2.2. Statistical Analysis
2.3. Energy Consumption
3. Results and Discussion
3.1. Light Intensity
3.2. Exogenous Cytokinin and Interaction with Light
3.3. Predictive Regression Model
3.4. Energy Consumption
4. Conclusions
- The usage of combination of red (70%) and blue (30%) LEDs in specific light intensities (40–120 µmol m−2 s−1) and different BA concentration in the media (1–5 µM) could be an effective and economical tool for modifying the potency of gerbera shoot multiplication and for controlling plant morphometry and photosynthetic pigment content.
- Cytokinin in the media influenced the shoot propagation of gerbera; with the increase BA concentration, the shoot multiplication rate intensified in all light intensities. A PPFD of 80 µmol m−2 s−1 and 5 µM BA is suggested as efficient for shoot propagation and economically viable simultaneously.
- A PPFD of 120 µmol m−2 s−1 decreased plantlet height. The interaction between light intensities and BA concentration was observed in terms of the effect on plant height. With PPFD 80 µmol m−2 s−1, a decreased concentration of BA (1 µM) enhanced the plantlet height, unlike with PPFD 40 µmol m−2 s−1, where the highest plants were noted with 5 µM BA in media.
- PPFD of 120 µmol m−2 s−1 increased some leaf parameters: blade area and width, and circularity and elongation ratios; the shape ratio decreased with an increasing light intensity.
- The dry weight content decreased with increasing cytokinin concentration, and the largest was observed in shoots multiplicated on media with 1 µM BA under PPFD µmol 120 m−2 s−1 and under control fluorescent lamps.
- PPFD 80 µmol m−2 s−1 increased the photosynthetic pigment content in the leaves in comparison to the standard intensity of light. Increased BA concentration raises the content of chlorophyll a.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- George, E.F.; Davies, W. Effects of the physical environment. In Plant propagation by Tissue Culture, 3rd ed.; George, E.F., Hall, M.A., de Klerk, G.-J., Eds.; Springer: AA Dordrecht, The Netherlands, 2008; pp. 423–464. [Google Scholar]
- Banerjee, A.; Roychoudhury, A. Plant responses to light stress: oxidative damages, photoprotection, and role of phytohormones. In Plant Hormones under Challenging Environmental Factors; Ahammed, G.J., Yu, J.Q., Eds.; Springer: AA Dordrecht, The Netherlands, 2016; pp. 181–214. [Google Scholar] [CrossRef]
- Miler, N.; Kulus, D.; Woźny, A.; Rymarz, D.; Hajzer, M.; Wierzbowski, K.; Nelke, R.; Szeffs, L. Application of wide-spectrum light-emitting diodes in micropropagation of popular ornamental plant species: a study on plant quality and cost reduction. Vitr. Cell. Dev. Biol. -Plant 2019, 55, 99–108. [Google Scholar] [CrossRef]
- Moe, R.; Grimstad, S.O.; Gislerød, H.R. The use of artificial light in year round production of greenhouse crops in Norway. Acta Hortic. 2006, 711, 35–42. [Google Scholar] [CrossRef]
- Alvarenga, I.C.A.; Pacheco, F.V.; Silva, S.T.; Bertolucci, S.K.V.; Pinto, J.E.B.P. In vitro culture of Achillea millefolium L.: Quality and intensity of light on growth and production of volatiles. Plant Cell, Tissue Organ Cult. 2015, 122, 299–308. [Google Scholar] [CrossRef]
- Silva, S.T.; Bertolucci, S.K.V.; da Cunha, S.H.B.; Lazzarini, L.E.S.; Tavares, M.C.; Pinto, J.E.B.P. Effect of light and natural ventilation systems on the growth parameters and carvacrol content in the in vitro cultures of Plectranthus amboinicus (Lour.) Spreng. Plant Cell, Tissue Organ Cult. 2017, 129, 501–510. [Google Scholar] [CrossRef]
- Soontornchainaksaeng, P.; Chaicharoen, S.; Sirijuntarut, M.; Kruatrachue, M. In vitro studies on the effect of light intensity on plant growth of Phaius tankervilliae (Banks ex L ’ Herit.) Bl. and Vanda coerulea Griff. Sci. Asia 2001, 27, 233–237. [Google Scholar] [CrossRef]
- Liu, C.; Guo, C.; Wang, Y.; Ouyang, F. Effect of light irradiation on hairy root growth and artemisinin biosynthesis of Artemisia annua L. Process Biochem. 2002, 38, 581–585. [Google Scholar] [CrossRef]
- Morelli, G.; Ruberti, I. Signaling shade avoidance responses. Driving auxin along lateral routes. Plant Physiol. 2000, 122, 621–626. [Google Scholar] [CrossRef]
- Folta, K.M.; Childers, K.S. Light as a growth regulator: Controlling plant biology with narrow-bandwidth solid-state lighting systems. HortScience 2008, 43, 1957–1964. [Google Scholar] [CrossRef]
- Moon, H.K.; Park, S.-Y.; Kim, Y.W.; Kim, C.S. Growth of Tsuru-rindo (Tripterospermum japonicum) cultured in vitro under various sources of light-emitting diode (LED) irradiation. J. Plant Biol. 2006, 49, 174–179. [Google Scholar] [CrossRef]
- Nhut, D.T.; Takamura, T.; Watanabe, H.; Okamoto, K.; Tanaka, M. Responses of strawberry plantlets cultured in vitro under superbright red and blue light-emitting diodes (LEDs). Plant Cell, Tissue Organ Cult. 2003, 73, 43–52. [Google Scholar] [CrossRef]
- Silva, M.M.A.; De Oliveira, A.L.B.; Oliveira-Filho, R.A.; Gouveia-Neto, A.S.; Camara, T.J.R.; Willadino, L.G. Effect of blue/red LED light combination on growth and morphogenesis of Saccharum officinarum plantlets in vitro. Prog. Biomed. Opt. Imaging - Proc. SPIE 2014, 89471. [Google Scholar] [CrossRef]
- OuYang, F.; Mao, J.-F.; Wang, J.; Zhang, S.; Li, Y. Transcriptome analysis reveals that red and blue light regulate growth and phytohormone metabolism in Norway Spruce [Picea abies (L.) Karst.]. PLOS One 2015, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.M.; Kerbauy, G.B. Effects of light and ethylene on endogenous hormones and development of Catasetum fimbriatum (Orchidaceae). Brazilian J. Plant Physiol. 2006, 18, 359–365. [Google Scholar] [CrossRef]
- Aksenova, N.P.; Konstantinova, T.N.; Sergeeva, L.I.; Macháčková, I.; Golyanovskaya, S.A. Morphogenesis of potato plants in vitro. I. Effect of light quality and hormones. J. Plant Growth Regul. 1994, 13, 143–146. [Google Scholar] [CrossRef]
- Karnachuk, R.A.; Tishchenko, S.Y.; Golovatskaya, I.F. Endogenous phytohormones and regulation of morphogenesis of Arabidopsis thaliana by blue light. Russ. J. Plant Physiol. 2001, 48, 226–230. [Google Scholar] [CrossRef]
- Bourget, C.M. An introduction to light-emitting diodes. HortScience 2008, 43, 1944–1946. [Google Scholar] [CrossRef]
- Dutta Gupta, S. Light Emitiing Diodes for Agriculture-Smart Lighting, 1st ed.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. (Amsterdam) 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Hanus-Fajerska, E.; Wojciechowska, R. Impact of Light-Emitting Diodes (LEDs) on propagation of orchids in tissue culture. In Light Emitting Diodes for Agriculture; Gupta, S.D., Ed.; Springer: Singapore, 2017; pp. 305–320. [Google Scholar]
- Viršilė, A.; Olle, M.; Duchovskis, P. LED Lighting in Horticulture. In Light emitiing diodes for agriculture - smart lighting, 1st ed.; Gupta, S., Ed.; Springer: Singapore, 2017; pp. 113–147. [Google Scholar] [CrossRef]
- Morrow, R.C. LED lighting in horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef]
- Minerva, G.; Kumar, S. Micropropagation of Gerbera (Gerbera jamesonii Bolus). In Protocols for Micropropagation of Selected Economically-Important Horticultural Plants; Lambardi, M., Ozudogru, E.A., Jain, S.M., Eds.; Springer: New York, NY, USA, 2013; pp. 305–316. [Google Scholar] [CrossRef]
- Sevelius, N. Photosynthetic features of three gerbera cultivars in low light. Acta Hortic. 2003, 624, 297–302. [Google Scholar] [CrossRef]
- Keditsu, R. Performance of open field Gerbera in resposne to planting time. African J. Agri. Res 2013, 8, 3068–3074. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tabacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bornwaßer, T.; Tantau, H.J. Evaluation of LED lighting systems in in vitro cultures. Acta Hortic. 2012, 956, 555–562. [Google Scholar] [CrossRef]
- Robinson, A.S. Plant Light Stress. Encycl. Life Sci. 2001, 1–6. [Google Scholar] [CrossRef]
- Singh, A.S.; Jones, A.M.P.; Shukla, M.R.; Saxena, P.K. High light intensity stress as the limiting factor in micropropagation of sugar maple (Acer saccharum Marsh.). Plant Cell. Tissue Organ Cult. 2017, 129, 209–221. [Google Scholar] [CrossRef]
- Wolf, S.; Kalman-Rotem, N.; Yakir, D.; Ziv, M. Autotrophic and heterotrophic carbon assimilation of in vitro grown potato (Solanum tuberosum L.) plants. J. Plant Physiol. 1998, 153, 574–580. [Google Scholar] [CrossRef]
- Pawłowska, B.; Żupnik, M.; Szewczyk-Taranek, B.; Cioć, M. Impact of LED light sources on morphogenesis and levels of photosynthetic pigments in Gerbera jamesonii grown in vitro. Hortic. Environ. Biotechnol. 2018, 59, 115–123. [Google Scholar] [CrossRef]
- Vanneste, S.; Friml, J. Auxin: A trigger for change in plant development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef]
- Symons, G.M.; Reid, J.B. Hormone levels and response during de-etiolation in pea. Planta 2003, 216, 422–431. [Google Scholar] [CrossRef]
- Deinum, B.; Sulastri, R.D.; Zeinab, M.H.J.; Maassen, A. Effects of light intensity on growth, anatomy and forage quality of two tropical grasses (Brachiaria brizantha and Panicum maximum var. trichoglume). Netherlands J. Agric. Sci. 1996, 44, 111–124. [Google Scholar]
- Fernandes, V.F.; de Almeida, L.B.; Feijó, E.V.R.; Silva, D.D.; Oliveira, R.A.; Mielke, M.S.; Costa, L.C. Light intensity on growth, leaf micromorphology and essential oil production of Ocimum gratissimum. Brazilian J. Pharmacogn. 2013, 23, 419–424. [Google Scholar] [CrossRef]
- Sáez, P.L.; Bravo, L.A.; Latsague, M.I.; Sánchez, M.E.; Ríos, D.G. Increased light intensity during in vitro culture improves water loss control and photosynthetic performance of Castanea sativa grown in ventilated vessels. Sci. Hortic. (Amsterdam). 2012, 138, 7–16. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, D.; Ma, Z.; Li, X.; Xiao, Y. Growth and photosynthetic capability of Momordica grosvenori plantlets grown photoautotrophically in response to light intensity. HortScience 2009, 44, 757–763. [Google Scholar] [CrossRef]
- Jo, E.A.; Tewari, R.K.; Hahn, E.J.; Paek, K.Y. Effect of photoperiod and light intensity on in vitro propagation of Alocasia amazonica. Plant Biotechnol. Rep. 2008, 2, 207–212. [Google Scholar] [CrossRef]
- Gupta, S.K.; Singh, P.K.; Sawant, S.V.; Chaturvedi, R.; Tuli, R. Effect of light intensity on in vitro multiple shoot induction and regeneration of cotton (Gossypium hirsutum L. cv Khandawa-2). Indian J. Exp. Biol. 2000, 38, 399–401. [Google Scholar] [PubMed]
- Fallah, F.; Kahrizi, D. Effect of Light spectrum and intensity on growth of grape (Vitis vinifera) under in vitro conditions. Orig. Artic. J. Appl. Biotechnol. Rep. Autumn 2016, 3, 495–499. [Google Scholar]
- Andrade, H.B.; Braga, A.F.; Bertolucci, S.K.V.; Hsie, B.S.; Silva, S.T.; Pinto, J.E.B.P. Effect of plant growth regulators, light intensity and LED on growth and volatile compound of Hyptis suaveolens (L.) Poit in vitro plantlets. Acta Hortic. 2017, 1155, 277–284. [Google Scholar] [CrossRef]
- Meziani, R.; Jaiti, F.; Mazri, M.A.; Anjarne, M.; Chitt, M.A.; El Fadile, J.; Alem, C. Effects of plant growth regulators and light intensity on the micropropagation of date palm (Phoenix dactylifera L.) cv. Mejhoul. J. Crop Sci. Biotechnol. 2015, 18, 325–331. [Google Scholar] [CrossRef]
- Zhou, M.; Guan, Q.; Wei, Y.; Zhang, Z. Effects of sucrose concentration and light intensity on growth and photosynthesis of ginger plantlets in vitro. Chinese J. Appl. Environ. Biol. 2008, 14, 356–361. [Google Scholar]
- Sánchez-Sánchez, J.; López-Orenes, A.; Ferrer, M.A.; Calderón, A.A. Subtle changes in light intensity affect in vitro responses but not ex vitro performance of Limonium sinuatum. 3 Biotech 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Kulchin, Y.N.; Nakonechnaya, O.V.; Gafitskaya, I.V.; Grishchenko, O.V.; Epifanova, T.Y.; Orlovskaya, I.Y.; Zhuravlev, Y.N.; Subbotin, E.P. Plant morphogenesis under different light intensity. Defect Diffus. Forum 2018, 386, 201–206. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Nevo, E.; Pavlíek, T.; Beharv, A.; Bolshakova, M.A.; Martyn, G.I.; Musatenko, L.I.; Sytnikl, K.M. Drought and light anatomical adaptive leaf strategies in three woody species caused by microclimatic selection at “Evolution Canyon”, Israel. Isr. J. Plant Sci. 2000, 48, 33–46. [Google Scholar] [CrossRef]
- Guerfel, M.; Baccouri, O.; Boujnah, D.; Chaïbi, W.; Zarrouk, M. Impacts of water stress on gas exchange, water relations, chlorophyll content and leaf structure in the two main Tunisian olive (Olea europaea L.) cultivars. Sci. Hortic. (Amsterdam). 2009, 119, 257–263. [Google Scholar] [CrossRef]
- Xiao, Y.; He, L.; Liu, T.; Yang, Y. Growth promotion of gerbera plantlets in largevessels by using photoautotrophic micropropagation system with forced ventilation. Propag. Ornam. Plants 2005, 5, 179–185. [Google Scholar]
- Yao, X.Y.; Liu, X.Y.; Xu, Z.G.; Jiao, X.L. Effects of light intensity on leaf microstructure and growth of rape seedlings cultivated under a combination of red and blue LEDs. J. Integr. Agric. 2017, 16, 97–105. [Google Scholar] [CrossRef]
- Kubota, S.; Yoneda, K. Effect of light intensity preceding day-night temperatures on the sensitivity of Phalaenopsis to flower (Japanese). J. Japanese Soc. Hortic. Sci. 1993, 62, 595–600. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.S.; Pons, T.L. Plant Physiological Ecology, 2nd ed.; Springer: New York, NY, USA, 2008. [Google Scholar] [CrossRef]
- Takahashi, S.; Murata, N. How do environmental stresses accelerate photoinhibition? Trends Plant Sci. 2008, 13, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 1–7. [Google Scholar] [CrossRef]
- Franck, F.; Schoefs, B.; Barthélemy, X.; Myśliwa-Kurdziel, B.; Strzałka, K.; Popovic, R. Protection of native chlorophyll(ide) forms and of photosystem II against photodamage during early stages of chloroplast differentiation. Acta Physiol. Plant. 1995, 17, 123–132. [Google Scholar]
- Bartley, G.E.; Scolnik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant Cell 1995, 7, 1027–1038. [Google Scholar] [CrossRef]
- Gonçalves, J.F.; Barreto, D.C.; Santos Junior, U.M.; Fernandes, A.V.; Sampaio, P.D.; Buckeridge, M.S. Growth, photosynthesis and stress indicators in young rosewood plants (Aniba rosaeodora Ducke) under different light intensities. Brazilian J. Plant Physiol. 2005, 17, 325–334. [Google Scholar] [CrossRef]
- Mahmood, S.; Reza, R.; Hossain, G.; Hauser, B. Response of cytokinins on in vitro shoot multiplication of rose cv. Frisco. Res. Rev. 2017, 6–11. [Google Scholar]
- Dobránszki, J.; Mendler-Drienyovszki, N. Cytokinin-induced changes in the chlorophyll content and fluorescence of in vitro apple leaves. J. Plant Physiol. 2014, 171, 1472–1478. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A.V.; Belgio, E. The relationship between maximum tolerated light intensity and photoprotective energy dissipation in the photosynthetic antenna: chloroplast gains and losses. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 1–9. [Google Scholar] [CrossRef] [PubMed]
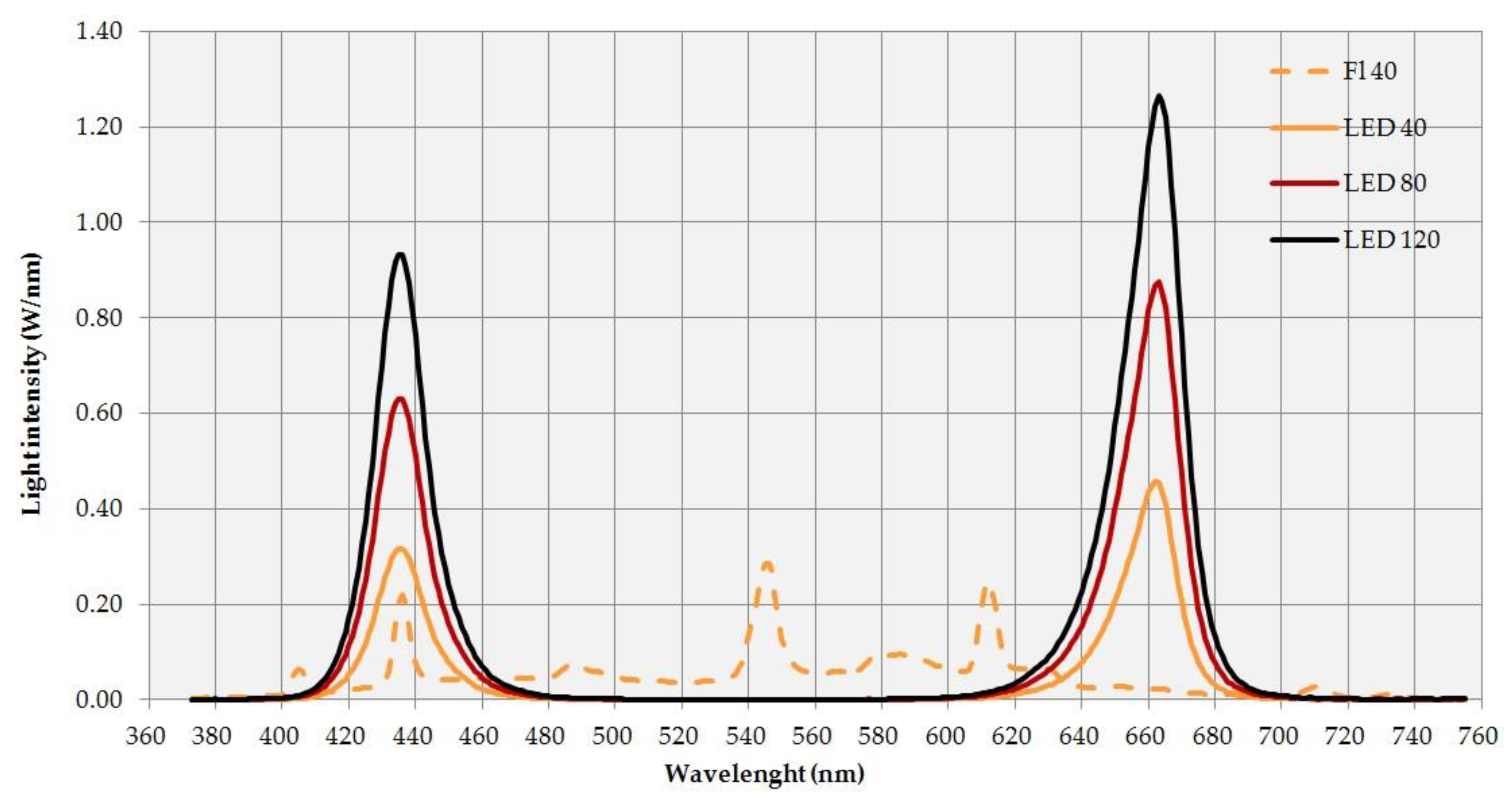





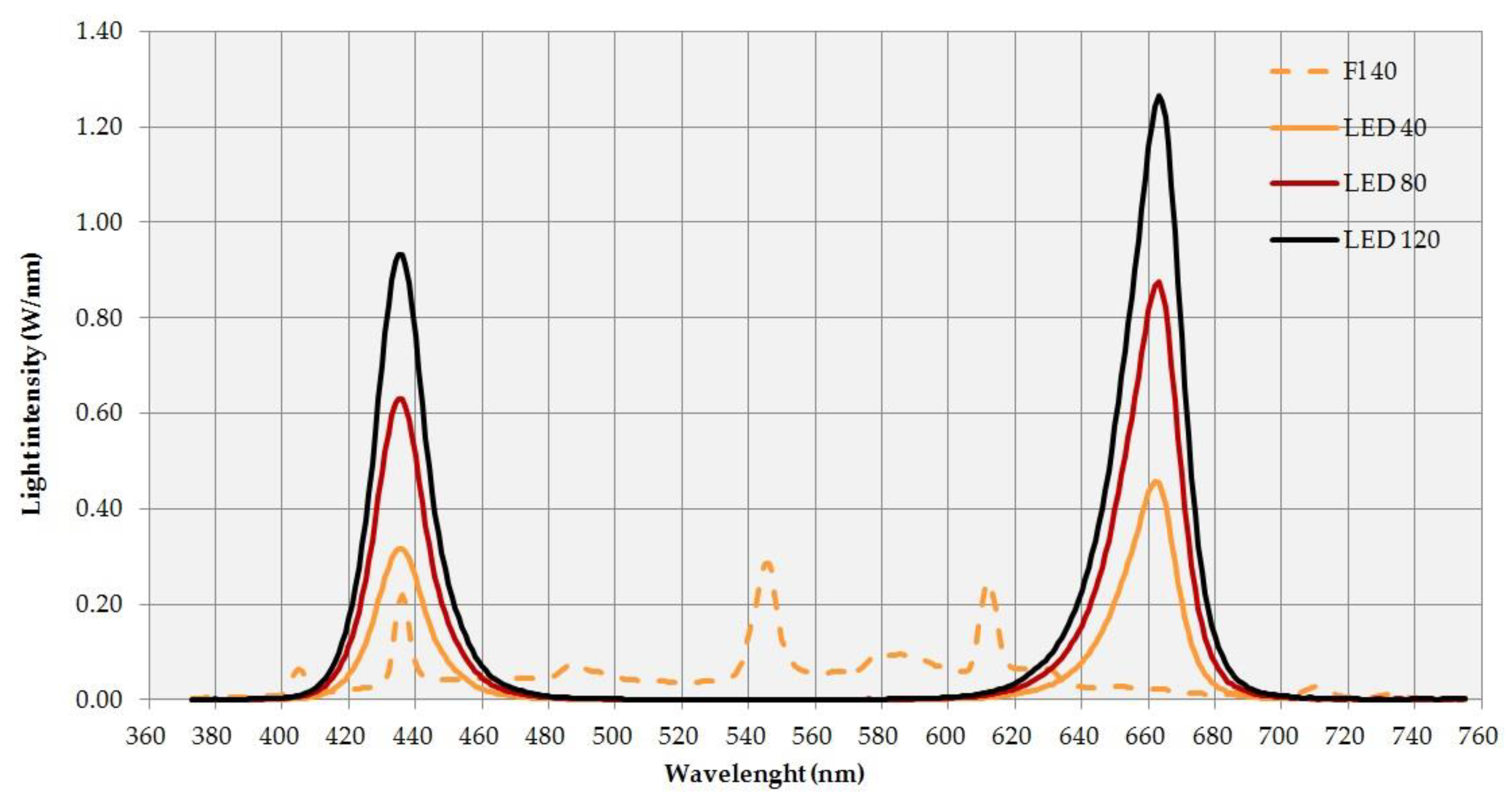{kind=link}
{kind=link}
{kind=link}
{kind=link}
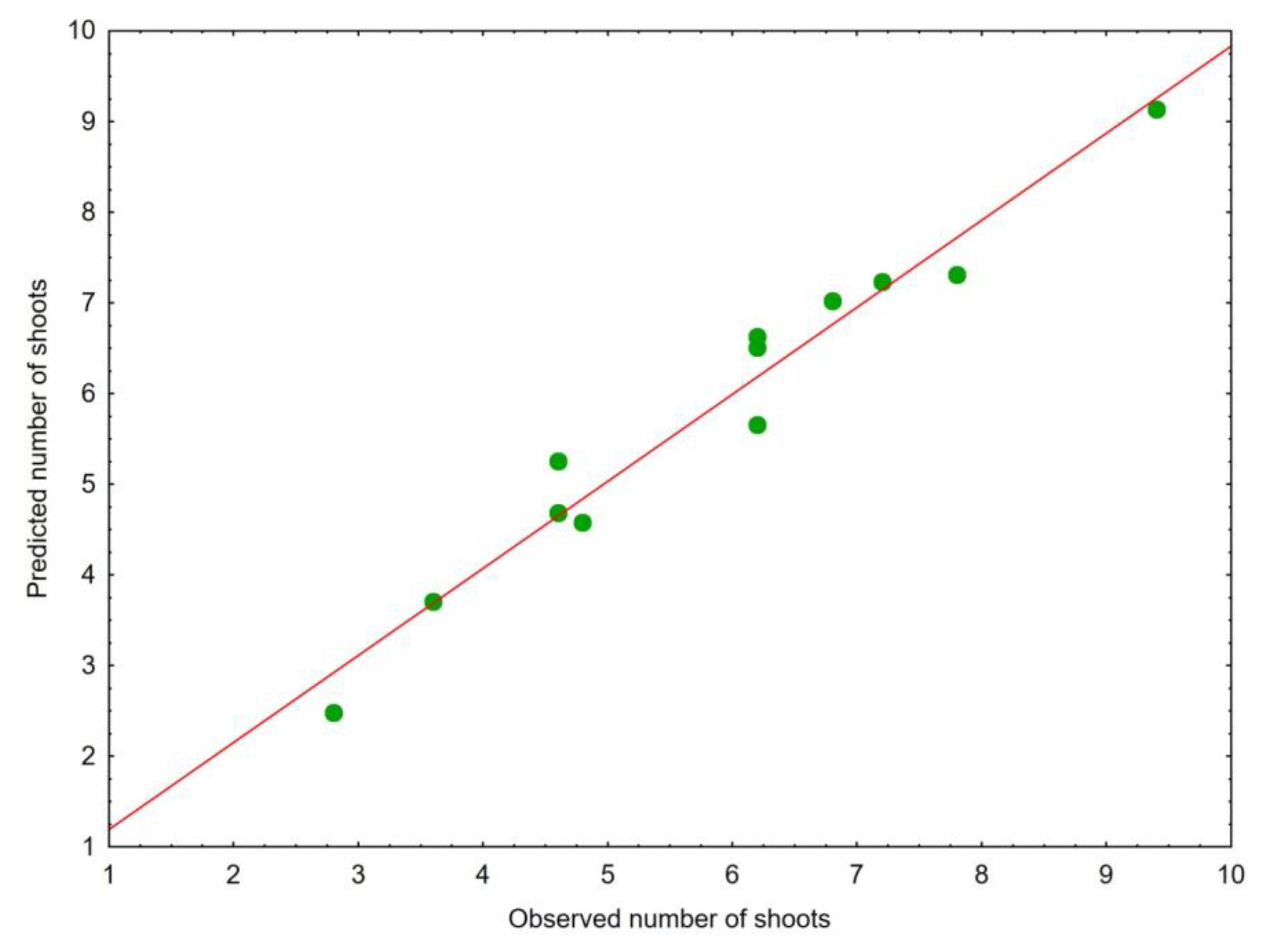{kind=link}
| PPFD (µmol m−2 s−1) | BA (µM) | Biometric Features | ||||
|---|---|---|---|---|---|---|
| Multiplication Rate | Plantlet Height (cm) | Number of Leaves per Shoot | Fresh Weight of Plantlet (g) | Dry Weight (%) | ||
| Fl 40 | 1 | 4.6 ± 0.9 a–c | 4.0 ± 0.9 d | 4.6 ± 0.9 ab | 0.81 ± 0.15 a–c | 8.91 ± 1.15 cd |
| 2.5 | 6.8 ± 0.8 c–e | 3.8 ± 0.8 cd | 4.2 ± 0.3 a | 1.11 ± 0.20 b–e | 7.04 ± 0.07 ab | |
| 5 | 9.4 ± 3.2 e | 3.7 ± 0.5 cd | 5.1 ± 1.3 ab | 1.54 ± 0.33 e | 6.40 ± 0.15 a | |
| LED 40 | 1 | 4. 8 ± 2.2 a–c | 2.9 ± 0.8 a–c | 5.0 ± 0.8 ab | 0.70 ± 0.23 ab | 7.59 ± 0.67 a–c |
| 2.5 | 6.2 ± 1.8 b–d | 3.6 ± 0.4 cd | 4.9 ± 0.3 ab | 1.17 ± 0.45 c–e | 8.29 ± 2.27 b–d | |
| 5 | 6.2 ± 1.3 b–d | 4.1 ± 0.5 d | 5.9 ± 1.4 b | 1.18 ± 0.27 c–e | 6.25 ± 0.43 a | |
| LED 80 | 1 | 3.6 ± 1.1 ab | 3.6 ± 0.4 b–d | 5.2 ± 0.3 ab | 0.88 ± 0.15 a–c | 7.11 ± 0.36 ab |
| 2.5 | 6.2 ± 1.9 b–d | 3.0 ± 0.5 a–c | 5.4 ± 1.8 ab | 1.37 ± 0.32 de | 6.89 ± 0.14 ab | |
| 5 | 7.8 ± 3.5 de | 2.9 ± 0.7 a–c | 4.8 ± 0.6 ab | 1.26 ± 0.44 c–e | 6.65 ± 0.42 a | |
| LED 120 | 1 | 2.8 ± 1.3 a | 2.2 ± 0.9 a | 6.0 ± 1.5 b | 0.64 ± 0.24 a | 9.31 ± 0.30 d |
| 2.5 | 4.6 ± 1.1 a–c | 2.7 ± 0.4 ab | 5.6 ± 1.0 ab | 0.98 ± 0.19 a–d | 6.81 ± 0.68 ab | |
| 5 | 7.2 ± 2.6 c–e | 2.6 ± 0.6 a | 5.4 ± 0.5 ab | 1.20 ± 0.53 c–e | 6.58 ± 0.63 a | |
| Means for PPFD | ||||||
| Fl 40 | 6.9 ± 2.7 b | 3.8 ± 0.7 c | 4.6 ± 0.9 a | 1.15 ± 0.38 a | 7.45 ± 1.27 a | |
| LED 40 | 5.7 ± 1.8 ab | 3.5 ± 0.7 bc | 5.3 ± 1.0 ab | 1.02 ± 0.38 a | 7.38 ± 1.50 a | |
| LED 80 | 5.9 ± 2.9 ab | 3.2 ± 0.6 b | 5.1 ± 1.0 ab | 1.17 ± 0.37 a | 6.88 ± 0.35 a | |
| LED 120 | 4.9 ± 2.5 a | 2.5 ± 0.6 a | 5.7 ± 1.0 b | 0.94 ± 0.40 a | 7.57 ± 1.40 a | |
| Means for BA | ||||||
| 1 | 4.0 ± 1.6 a | 3.2 ± 1.0 a | 5. 2 ± 1.0 a | 0.76 ± 0.20 a | 8.23 ± 1.12 c | |
| 2.5 | 6.0 ± 1.6 b | 3.3 ± 0.7 a | 5.0 ± 1.1 a | 1.15 ± 0.32 b | 7.26 ± 1.19 b | |
| 5 | 7.7 ± 2.8 c | 3.3 ± 0.8 a | 5.3 ± 1.0 a | 1.30 ± 0.40 b | 6.47 ± 0.41 a | |
| Source of variation | ||||||
| PPFD × BA | n.s. | ** | n.s. | n.s. | *** | |
| PPFD | n.s. | *** | n.s. | n.s. | n.s. | |
| BA | *** | n.s. | n.s. | ** | ||
| PPFD (µmol m−2 s−1) | BA (µM) | Leaf blade | Petiole | |||
|---|---|---|---|---|---|---|
| Area (mm2) | Perimeter (mm) | Length (mm) | Width (mm) | Length (mm) | ||
| Fl 40 | 1 | 76.91 ± 37.50 bc | 71.36 ± 24.51 b | 30.04 ± 10.68 e | 6.70 ± 2.19 b–d | 19.64 ± 7.96 bc |
| 2.5 | 59.36 ± 22.42 ab | 68.99 ± 20.81 b | 28.60 ± 8.83 c–e | 5.88 ± 1.81 ab | 21.95 ± 8.46 c | |
| 5 | 46. 30 ± 35.29 a | 55.58 ± 21.52 a | 22.62 ± 8.83 ab | 4.62 ± 3.00 a | 15.92 ± 7.80 b | |
| LED 40 | 1 | 71.33 ± 29.78 ab | 66.04 ± 18.62 ab | 26.97 ± 7.91 b–e | 6.75 ± 1.78 b–d | 17.22 ± 6.18 b |
| 2.5 | 82.20 ± 28.96 bc | 67.94 ± 13.52 ab | 26.81 ± 5.93 b–e | 7.46 ± 2.07 b–d | 16.75 ± 5.21 b | |
| 5 | 61.98 ± 27.13 ab | 61.93 ± 20.06 ab | 23.92 ± 8.63 a–d | 6.19 ± 1.54 bc | 16.19 ± 7.57 b | |
| LED 80 | 1 | 132.81 ± 47.44 e | 74.23 ± 22.75 b | 29.15 ± 9.35 de | 9.50 ± 3.59 fg | 17.04 ± 5.39 b |
| 2.5 | 98.00 ± 35.18 cd | 70.02 ± 14.44 b | 27.93 ± 6.24 b–e | 8.32 ± 2.44 d–f | 16.58 ± 4.43 b | |
| 5 | 78.32 ± 25.15 bc | 64.67 ± 16.78 ab | 25.13 ± 7.37 a–e | 7.63 ± 1.60 c–e | 16.91 ± 5.40 b | |
| LED 120 | 1 | 132.64 ± 63.48 e | 63.44 ± 12.82 ab | 24.38 ± 3.94 a–d | 10.15 ± 3.23 g | 10.58 ± 2.56 a |
| 2.5 | 108.61 ± 44.88 ed | 61.14 ± 10.65 ab | 23.54 ± 4.25 a–c | 9.10 ± 2.49 e–g | 10.22 ± 3.20 a | |
| 5 | 81.06 ± 30.34 bc | 55.07 ± 10.14 a | 21.18 ± 4.31 a | 7.85 ± 1.66 c–e | 10.35 ± 3.46 a | |
| Means for PPFD | ||||||
| Fl 40 | 60.86 ± 34.29 a | 65.31 ± 23.04 ab | 27.08 ± 9.87 b | 5.73 ± 2.50 a | 19.17 ± 8.32 c | |
| LED 40 | 71.84 ± 29.36 a | 65.30 ± 17.51 ab | 25.90 ± 7.58 b | 6.80 ± 1.86 b | 16.72 ± 6.30 b | |
| LED 80 | 103.04 ± 42.93 b | 69.64 ± 18.44 b | 27.40 ± 7.81 b | 8.48 ± 2.74 c | 16.84 ± 5.01 b | |
| LED 120 | 107.44 ± 51.91 b | 59.88 ± 11.63 a | 23.04 ± 4.32 a | 9.03 ± 2.67 c | 10.38 ± 3.05 a | |
| Means for BA | ||||||
| 1 | 103.43 ± 54.18 c | 68.77 ± 20.26 b | 27.63 ± 8.49 b | 8.27 ± 3.16 b | 16.00 ± 6.65 a | |
| 2.5 | 87.04 ± 38.11 b | 67.02 ± 15.42 b | 26.72 ± 6.69 b | 7.69 ± 2.49 b | 16.49 ± 6.97 a | |
| 5 | 66.92 ± 32.36 a | 59.31 ± 17.82 a | 23.21 ± 7.51 a | 6.57 ± 2.39 a | 14.84 ± 6.73 a | |
| Source of variation | ||||||
| PPFD × BA | n.s. | n.s. | n.s. | n.s. | n.s. | |
| PPFD | *** | ** | *** | *** | *** | |
| BA | *** | *** | *** | *** | n.s. | |
| Variable | Equation Parameters | Standard Errors | t Statistic Values | p |
|---|---|---|---|---|
| Intercept | 27.6600 | 4.8766 | 5.6720 | 0.0024 |
| Fresh weight of one shoot (g) | 55.0505 | 14.1478 | 3.8911 | 0.0115 |
| Number of leaves per shoots | −3.1792 | 0.6716 | −4.7340 | 0.0052 |
| Leaf area (cm2) | −0.1801 | 0.0403 | −4.4662 | 0.0066 |
| Leaf perimeter (cm) | −0.2735 | 0.0497 | −5.5059 | 0.0027 |
| Leaf width (cm) | 2.0566 | 0.5746 | 3.5789 | 0.0159 |
| Chlorophyll b (mg g-1 fresh weight) | 12.2295 | 4.6124 | 2.6515 | 0.0454 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cioć, M.; Kalisz, A.; Żupnik, M.; Pawłowska, B. Different LED Light Intensities and 6-Benzyladenine Concentrations in Relation to Shoot Development, Leaf Architecture, and Photosynthetic Pigments of Gerbera jamesonii Bolus In Vitro. Agronomy 2019, 9, 358. https://doi.org/10.3390/agronomy9070358
Cioć M, Kalisz A, Żupnik M, Pawłowska B. Different LED Light Intensities and 6-Benzyladenine Concentrations in Relation to Shoot Development, Leaf Architecture, and Photosynthetic Pigments of Gerbera jamesonii Bolus In Vitro. Agronomy. 2019; 9(7):358. https://doi.org/10.3390/agronomy9070358
Chicago/Turabian StyleCioć, Monika, Andrzej Kalisz, Marek Żupnik, and Bożena Pawłowska. 2019. "Different LED Light Intensities and 6-Benzyladenine Concentrations in Relation to Shoot Development, Leaf Architecture, and Photosynthetic Pigments of Gerbera jamesonii Bolus In Vitro" Agronomy 9, no. 7: 358. https://doi.org/10.3390/agronomy9070358
APA StyleCioć, M., Kalisz, A., Żupnik, M., & Pawłowska, B. (2019). Different LED Light Intensities and 6-Benzyladenine Concentrations in Relation to Shoot Development, Leaf Architecture, and Photosynthetic Pigments of Gerbera jamesonii Bolus In Vitro. Agronomy, 9(7), 358. https://doi.org/10.3390/agronomy9070358